Introduction
Rheumatoid arthritis (RA), a chronic inflammatory
autoimmune disease, arises from a breakdown in self-tolerance which
is characterized by the infiltration of massive inflammatory cells
into both synovial tissues and synovial fluid, leading to chronic
synovitis, progressive erosions and cartilage destruction
(synovium, cartilage, and bone of multiple joints (1). CD4+CD25+
regulatory T cells (Tregs), a lymphocyte subpopulation, are engaged
in peripheral tolerance and anti-autoimmunity by directly or
indirectly suppressing self-reactive T cells and B cells. The
decreased number of Tregs in circulation, as well as deficiency of
their suppressive and anti-inflammatory activity in the joints have
been regarded as an indispensable factor of RA (2,3).
Although Tregs are unable to undergo mitosis, they do proliferate
in response to exogenous stimulation (4). Induction or amplification of Tregs may
serve as helpful and innovative therapies for RA (5).
In humans, Tregs have been identified in the
peripheral circulation and in the thymus (4). Tregs express certain chemokines and
chemokine receptors (such as CCR4, −5, and −6), rendering it
migrate into rheumatic joint to retard the activation of
auto-aggressive cells that have escaped central tolerance
mechanisms (6). The
macrophage-derived chemokine (MDC)/CCL22 belongs to the CC family,
and selectively interactes with the C-C chemokine receptor 4
(CCR4). Evidence suggests that CCL22 is significantly enhanced in
both synovial fluid and plasma in patients with RA (7). However, whether and how CCL22
functioned in RA remain largely unknown.
Signal transducer and activator of transcription 5
(STAT5) is implicated in maintaining immune function, particularly
in Tregs development and function (8,9).
Moreover, high expression of FOXP3 and the in vitro suppressive
activity of Tregs was maintained in the presence of
STAT5-activating cytokines, such as IL-2 and IL-15 (10). Evidence suggests that the expression
of chemokine CCL1 in primary human dermal fibroblasts is depended
on the activation of STAT5 (11).
To study the effect of chemokine CCL22 on Tregs,
CD4+ T cells obtained from RA patients and healthy
control were treated with anti-CCL22 antibody and recombinant CCL22
protein, respectively. STAT5 inhibitor (CAS 285986-31-4) was used
to block the STAT5 pathway (12).
Our data suggested that CCL22 decreased the number of Tregs and
FOXP3 expression via STAT5 pathway in human RA. Further, we found
that Sinomenine, which has been used in RA therapy for several
decades (13), regulated Tregs by
CCL22/STAT5 pathway.
Patients and methods
Patients
This study was approved by the Ethics Committee of
Shuguang Hospital Affiliated to Shanghai University of TCM
(Shanghai, China), and written informed consent from the
participants was obtained. Blood samples were collected from 30
pairs of RA patients and age-matched healthy controls for isolation
of serum.
Enzyme-linked immunosorbent assay
(ELISA)
Human Macrophage-Derived Chemokine, MDC ELISA Kit
96T/48T (XY-E10131, X-Y Biotechnology) was used to determine serum
concentration of CCL22 (pg/ml) in accordance with the
manufacturer's procedure.
Isolation and stimulation of
CD4+ T cells
Peripheral blood mononuclear cells (PBMCs) were
isolated from blood of RA patients (n=15) and healthy controls
(n=10) by using density gradient centrifugation method.
CD4+ T cells were isolated from PBMCs, using MagCellect
Human CD4+ T cell Isolation Kit (MAGH102; R&D
Systems), and then cultured in a medium of RPMI media (HyClone)
with 10% fetal calf serum (Gibco; Thermo Fisher Scientific, Inc.)
and 100 U/ml penicillin (Solarbio).
For cytokine responses, CD4+ T cells from
RA patients and healthy controls were exposed to anti-CCL22
antibody (ab9847; Abcam) at a dose of 0, 0.1, 0.2, 0.5, 1.0 and 2.0
µg/ml, and recombinant CCL22 (ab243277; Abcam) at a dose of 0, 2,
10 and 50 ng/ml, respectively. To study the involvement of STAT5
pathway in the promoted effect of anti-CCL22 antibody on Tregs
population, CD4+ T cells from RA patients were treated
with 50 ng/ml of anti-CCL22 antibody (0.5 µg/ml) plus 1 µM of STAT5
inhibitor (Millipore) or vehicle (DMSO). To study the effect of
sinomenine (SIN), CD4+ T cells from RA patients were
treated with 0, 1.0, 2.5, 5.0, 10 and 20 µg/l of SIN (SS8560;
Solarbio).
Flow cytometric analysis of Tregs
CD4+ T cells were stained with Anti-Human
FOXP3 Staining Kit (560133; BD Biosciences), and the percentage of
CD4+CD25+FOXP3+ cells in
CD4+ T cell population was analyzed on Accuri C6 flow
cytometer (BD Biosciences) with FlowJo 7.6.1 software (Tree Star
Inc.).
Real-time (RT)-PCR
Total RNA from CD4+ T cells was extracted
by TRIzol reagent (Sigma-Aldrich; Merck KGaA) and reverse
transcribed using cDNA synthesis kit (Takara). The primers for
CCL22 (NCBI NM_002990.5) were: 5′-CCTGCTTAAACCCTTCCATGAC-3′ and
5′-TTGGAGAACAGGGAGCTAGAAC-3′; primers for CCR4 (NCBI NM_005508.4)
were: 5′-CCTTCCTGGCTTTCTGTTC-3′ and 5′-CATCTTCACCGCCTTGTTC-3′,
primers for FOXP3 (NCBI NM_001114377.1) were:
5′-AGGAGGATGGACGAACAG-3′ and 5′-GGCAAGACAGTGGAAACC-3′; and primers
for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (NCBI
NM_001256799.2) were: 5′-AATCCCATCACCATCTTC-3′ and
5′-AGGCTGTTGTCATACTTC-3′. SYBR-Green PCR Kit (Thermo Fisher
Scientific, Inc.) with an ABI 7300 system (Applied Biosystems) was
used for analysis. mRNA levels of CCL22, CCR4 and FOXP3 were
normalized by GAPDH.
Western blot analysis
After fully lysed of CD4+ T cells, total
protein was quantified using BCA protein assay kit (Thermo Fisher
Scientific, Inc.), and 25 µg of which was separated using 15%
SDS-PAGE. After transferring onto PVDF membranes (Millipore), the
membrane were incubated with antibody against CCL22 (Ab9847), CCR4
(Ab832F50), FOXP3 (Ab20034), STAT5 (Ab230670), antibody against
phosphorylated (p)-STAT5 (Ab32364) (all from Abcam) and antibody
against GAPDH (#5174; Cell Signaling Technology) at 4°C overnight
followed by secondary antibodies (A0208 and A0216; Beyotime
Institute of Biotechnology) for 1 h at 25°C. ECL system (GE
Healthcare/Amersham Biosciences) was used for analysis.
Statistical analysis
Data were calculated using GraphPad Prism 7.0
software (GraphPad Software, Inc.) and described as mean ± standard
error of the mean (SEM). Comparison between RA and HC group, as
well as CD4+ T cells with various stimulations and the
corresponding Control group were conducted by one-way ANOVA
analysis with post-hoc Tukey's test, and P-value <0.05 was
considered significant.
Results
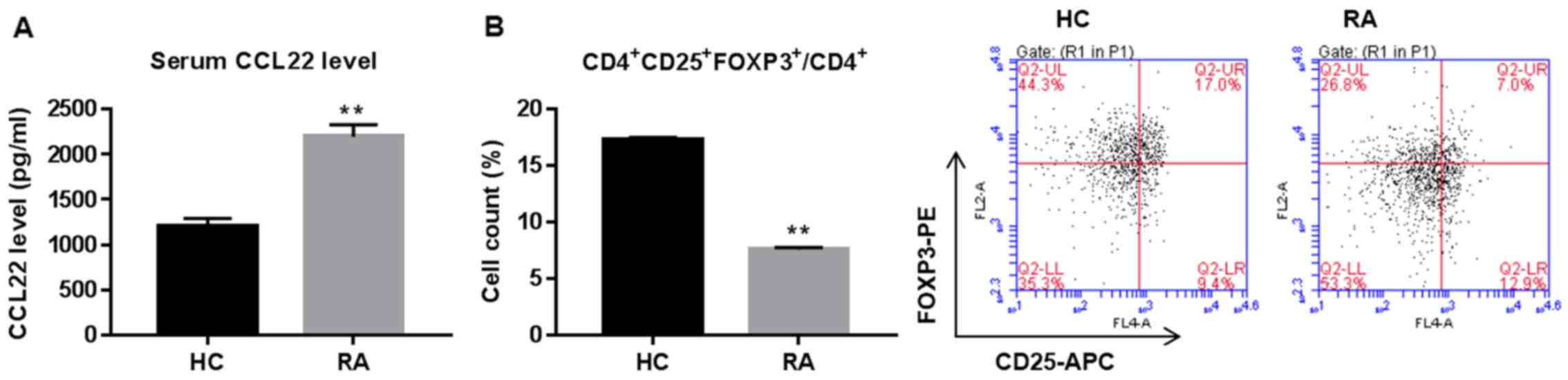
Serum CCL22 is upregulated while Tregs
are downregulated in RA
Serum concentration of CCL22 and Tregs proportion in
CD4+ T cells are shown in Fig. 1. CCL22 was significantly increased
while peripheral Tregs were reduced in RA patients when compared
with healthy controls (HC).
Roles of CCL22 in regulating the
number of Tregs in CD4+ T cells
To study the roles of CCL22 in regulating Tregs
expansion, CD4+ T cells isolated from RA were stimulated
with anti-CCL22 antibody (0, 0.1, 0.2, 0.5, 1.0 and 2.0 µg/ml)
while CD4+ T cells, isolated from HC, were stimulated
with exogenous CCL22 protein (0, 2, 10 and 50 ng/ml), then the
percentage of CD4+CD25+FOXP3+
regulatory T cells, and the expression of CCL22, CCR4 and FOXP3
within CD4+ T cells were determined. Our data showed
that, in comparison to their corresponding control, the percentage
of CD4+CD25+FOXP3+ regulatory T
cells in CD4+ T cell population from RA patients was
significantly enhanced 1, 65, 129, 157 and 228% under a stimulation
dose of anti-CCL22 antibody at 0.1, 0.2, 0.5, 1.0 and 2.0 µg/ml,
respectively (all P<0.01) (Fig.
2A). On the contrary, the percentage of
CD4+CD25+FOXP3+ regulatory T cells
in CD4+ T cell population from HC was significantly
reduced to 11, 23 and 40% under a stimulation dose of recombinant
CCL22 protein at 2, 10 and 50 ng/ml, respectively (all P<0.01)
(Fig. 2B). FOXP3, the Tregs function
associated maker, was dose-dependently upregulated by anti-CCL22
antibody, however, downregulated by recombinant CCL22 protein
(Fig. 2C and D) further confirming
the roles of CCL22 on Tregs function at a molecular level.
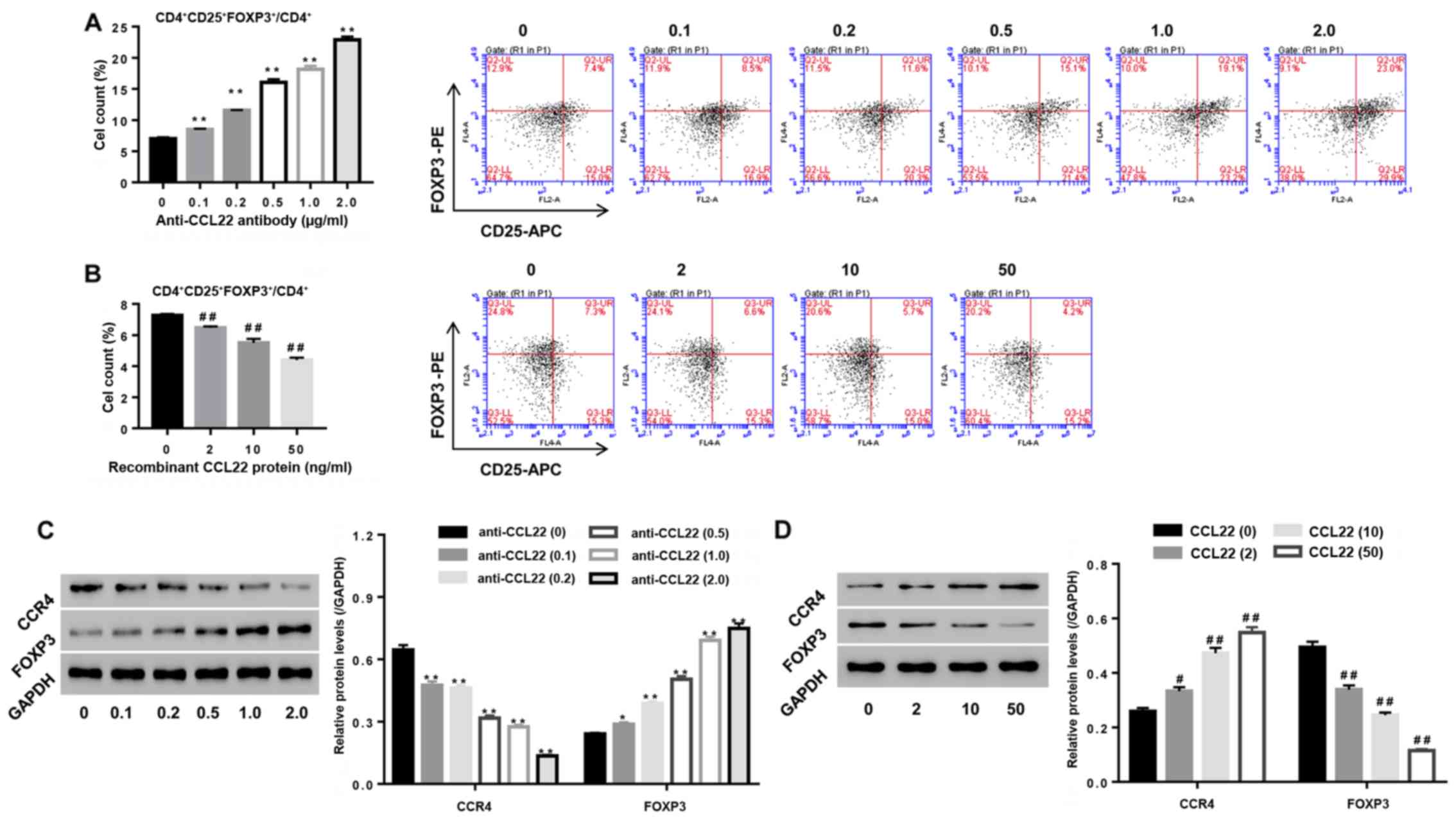 | Figure 2.Roles of CCL22 in regulating the
number and function associated makers of Tregs. (A and C)
Stimulation of CD4+ T cells of RA patients with
anti-CCL22 antibody (0.1, 0.2, 0.5, 1.0 and 2.0 µg/ml) caused
apromoted effect on the number of Tregs, as well as a decrease of
CCR4 while an increase in FOXP3 (C). (B and D) Stimulation of
CD4+ T cells of HC with CCL22 protein (2, 10 and 50
ng/ml) caused inhibition of the number of Tregs (B), as well as
increase in CCR4 while decreased FOXP3 (D). *P<0.05, **P<0.01
vs. control CD4+ T cells from RA; #P<0.05,
##P<0.01 vs. control CD4+ T cells from HC.
Tregs, regulatory T cells; CCR4, C-C chemokine receptor 4. |
In addition, Fig. 2C and
D indicated that anti-CCL22 antibody reduced the expression of
CCR4 in a dose-dependent manner, however, recombinant CCL22 protein
resulted in the opposite effects (all P<0.01), confirming
inhibition of CCR4 expression by anti-CCL22 antibody, while
stimulation of CCR4 expression by exogenous CCL22 protein.
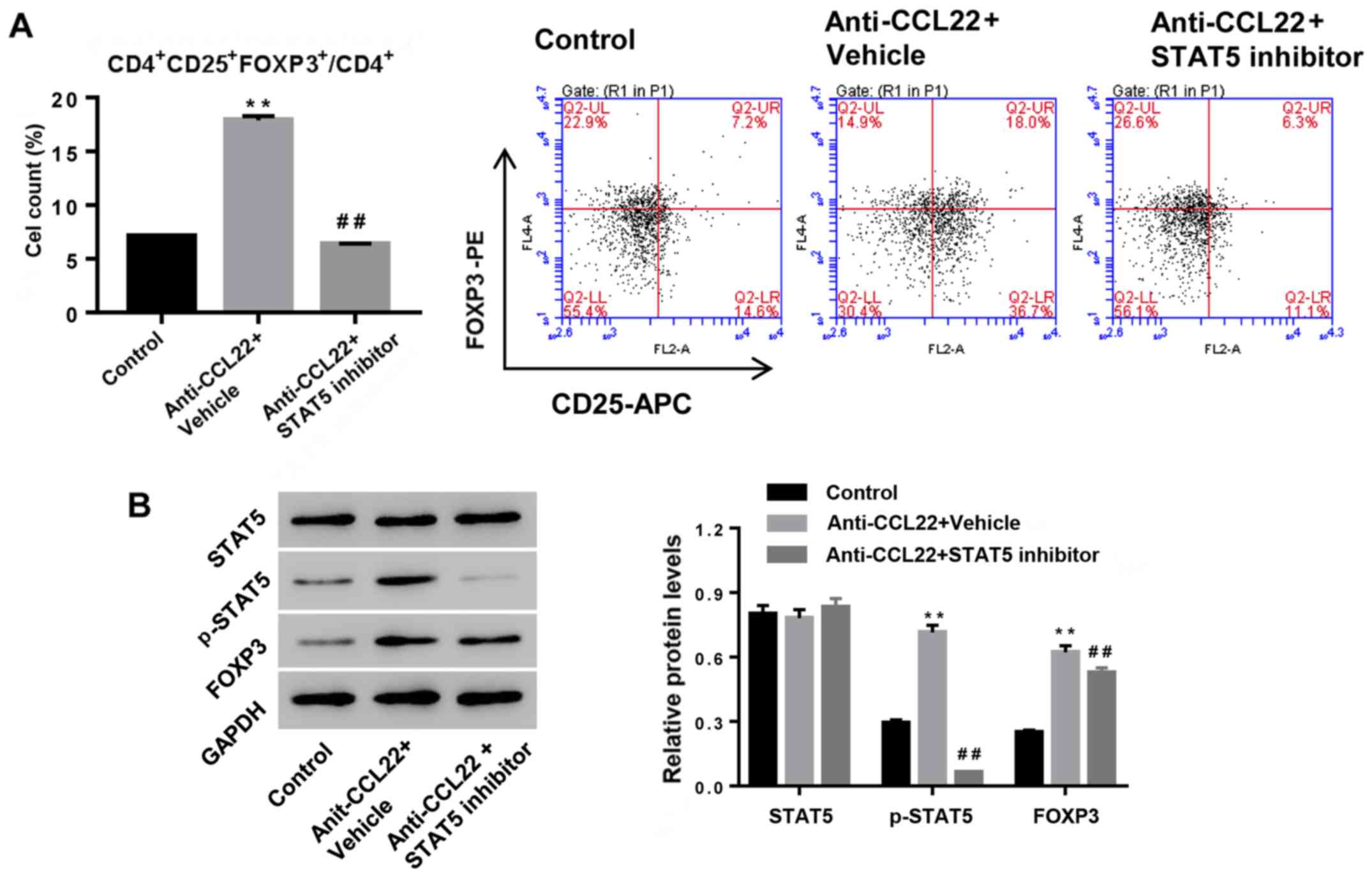
Anti-CCL22 antibody enhances Tregs
content by increasing p-STAT5
To study the possible mechanism, CD4+ T
cells, isolated from RA patients, were exposed to anti-CCL22
antibody (0.5 µg/ml) in the presence of STAT5 inhibitor (1 µM) or
vehicle, and then Tregs content and expression of p-STAT5 and FOXP3
were determined. Fig. 3 shows that
additional STAT5 inhibitor significantly reversed the effects of
anti-CCL22 antibody on Tregs content and the expression of p-STAT5
and FOXP3, suggesting that activation of STAT5 was the mechanism,
by which anti-CCL22 exerted promoting effect on the number of Tregs
in CD4+ T cells.
Sinomenine promotes the number of
Tregs and enhances CCL22 expression in CD4+ T cells
Sinomenine has been used in RA therapy for several
decades (13). To study the roles of
SIN in Tregs and its effect on CCL22 expression in CD4+
T cells, isolated CD4+ T cells from RA patients were
stimulated with SIN (1, 2.5, 5, 10 and 20 µg/l), and then the
number of Tregs was assessed, as well as expression of CCL22, CCR4
and FOXP3 in CD4+ T cells. Fig. 4A indicates that SIN promoted Treg
contents, and was associated with the downregulation of CCL22 and
CCR4, and the upregulation of FOXP3, substantiating that SIN
enhanced the number of Tregs and Tregs function at the molecular
level, and the involvement of CCL22 and CCR4 in this process.
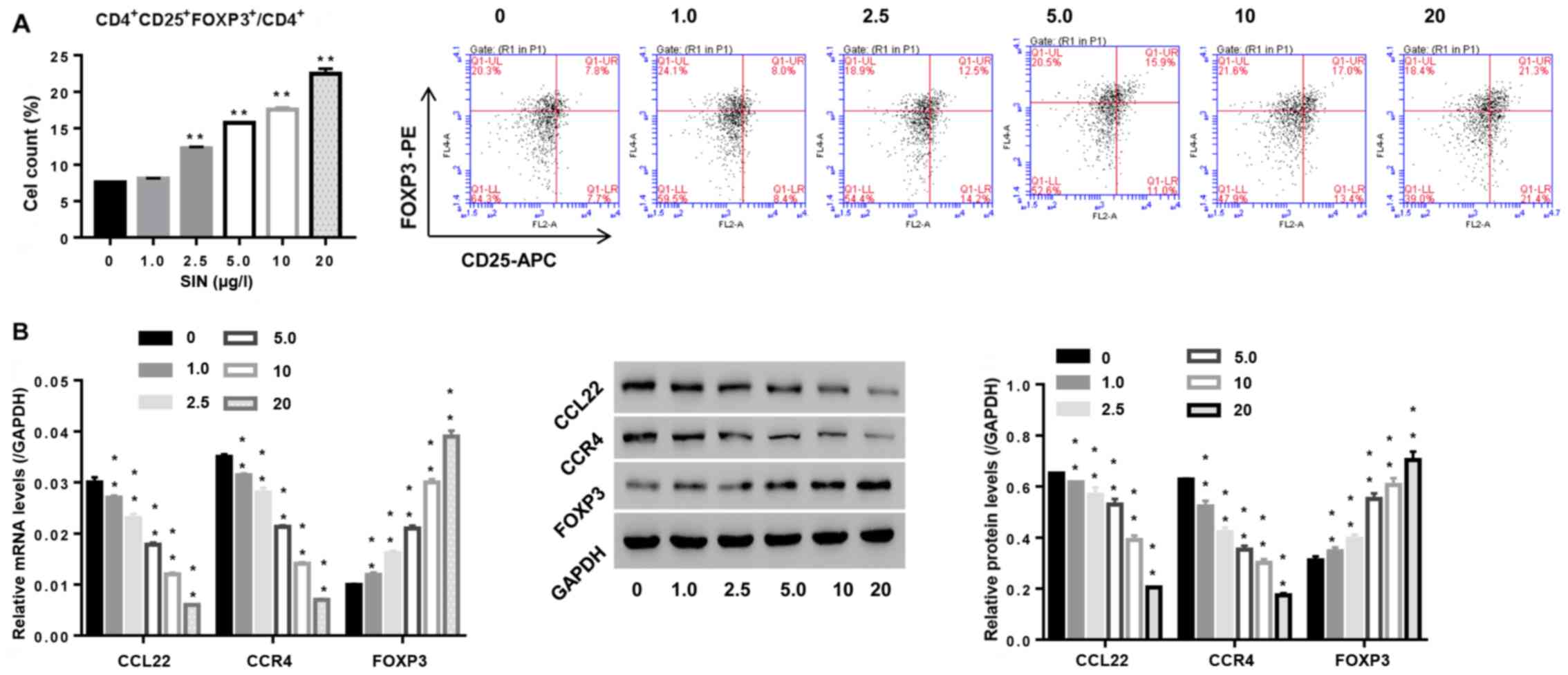 | Figure 4.Effect of sinomenine (SIN) on the
number of Tregs and function-associated makers. CD4+ T
cells, isolated from peripheral blood of RA patients, were exposed
to SIN (0, 1.0, 2.5, 5.0, 10 and 20 µg/l). (A) After 24 h, the
number of Tregs in CD4+ T cells was measured by flow
cytometric analysis. (B) 48 h later, mRNA and protein levels of
FOXP3, CCL22 and CCR4 were assessed by real-time PCR and western
blotting, respectively. **P<0.01 vs. 0 (CD4+ T cells
stimulated with vehicle PBS). Tregs, regulatory T cells; CCR4, C-C
chemokine receptor 4. |
Sinomenine promotes the number of
Tregs via CCL22/STAT5 pathway
We studied whether CCL22 and STAT5 was the
underlying mechanism, by which SIN regulated the number of Tregs in
CD4+ T cells. Tregs were treated with SIN, CCL22, STAT5
inhibitor, SIN plus CCL22, or SIN plus STAT5 inhibitor. As shown in
Fig. 5, SIN significantly enhanced
the number of Tregs and protein levels of FOXP3 and p-STAT5, and
the effects were weakened by additional CCL22 or STAT5 inhibitor
treatment. Thus, our data indicated that SIN increased the number
of Tregs probably via CCL22/STAT5 pathway.
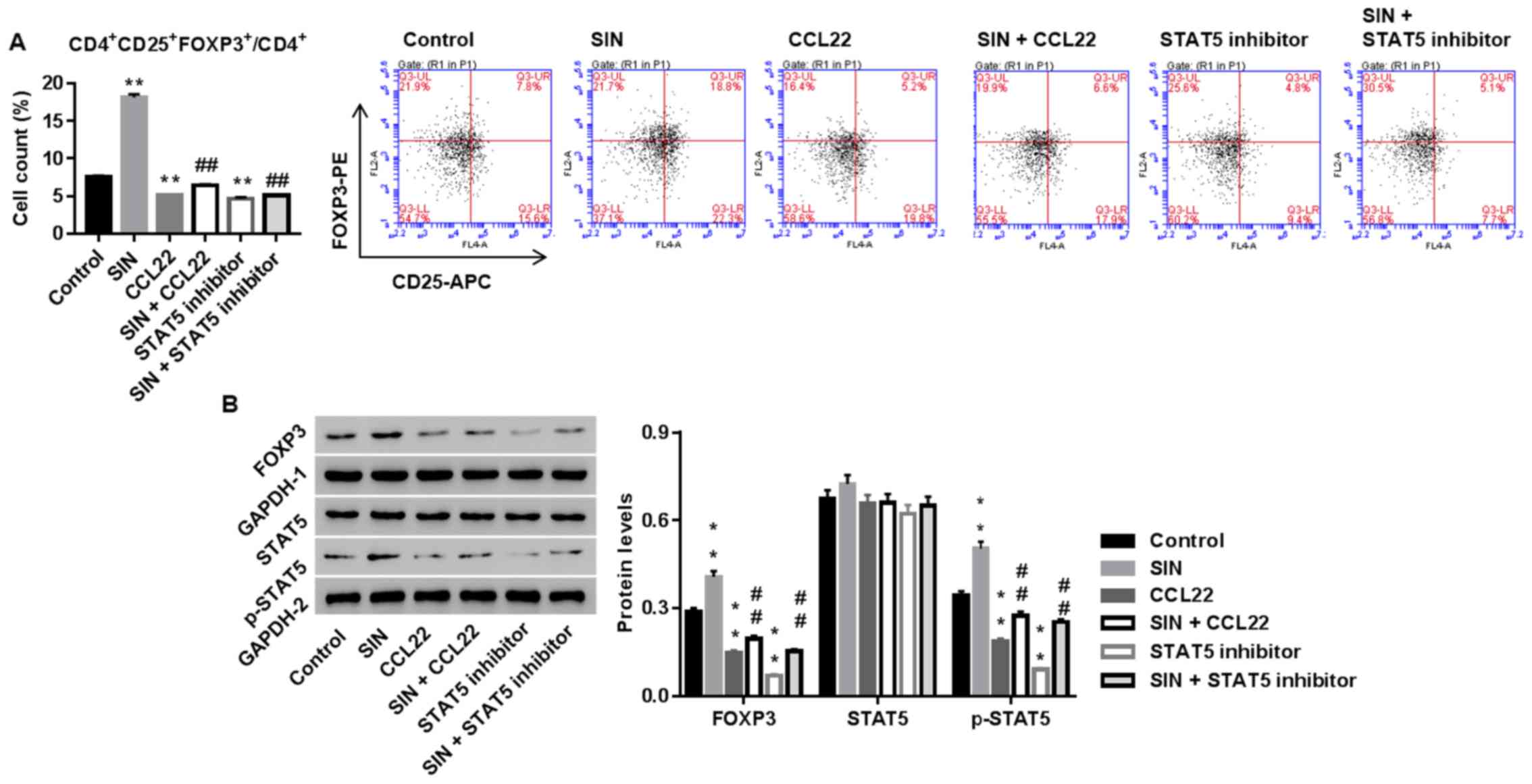 | Figure 5.Sinomenine (SIN) promotes the number
of Tregs via CCL22/STAT5 pathway. CD4+ T cells, isolated
from RA, were treated with vehicle, CCL22 (50 ng/ml) + vehicle,
CCL22 + SIN (5 µg/l), STAT5 inhibitor (1 µM), or SIN + STAT5
inhibitor. Twenty-four hours later, the number of Tregs (A) and
protein levels of FOXP3, STAT5 and p-STAT5 (B) were assessed.
GAPDH, loading control. **P<0.01 vs. control;
##P<0.01 vs. CCL22. Tregs, regulatory T cells; STAT5,
signal transducer and activator of transcription 5. |
Discussion
A hallmark of RA is the reduction of Tregs function
in peripheral blood, and Tregs expansion and transfer have been
involved in therapeutic implications in RA (2,4). FOXP3,
a specific maker for Tregs function, is responsible for Tregs
specific detection and enumeration. In the present study, we
confirmed that peripheral Tregs were decreased while serum CCL22
was increased in RA which was in line with a reported study
(Fig. 1) (7). Evidence suggests that injection of
CCL22 induces accumulation of CCR4 expressing T cells in vivo
(14). Given the roles of CCL22 in
regulating Tregs proliferation in tumor immunity (15), we studied whether CCL22 function in
modulating the number of Tregs in RA. Our data elucidated that
CCL22 increased CCR4 expression, and decreased the number of Tregs
and expression of FOXP3 in CD4+ T cells of healthy
controls, and anti-CCL22 showed the reverse effect in
CD4+ T cells from RA patients (Fig. 2). Our data indicated that the
elevated CCL22/CCR4 may involve in the pathogenesis of RA by
decreasing the number of Tregs.
Activation of STAT5 is sufficient to increase the
number of Tregs and favors Tregs homeostasis and self-tolerance
(16). Tregs with the high levels of
p-STAT5 is frequently associated with the high suppressive activity
in vitro (17). Expression levels of
p-STAT5 is down regulated in Tregs in RA patients (17). In this study, we suggest that the
activation of STAT5 was accelerated by anti-CCL22 antibody
(Fig. 3). The STAT5 inhibitor
abolished anti-CCL22 and increased the number of Tregs and FOXP3
expression. Thus, we deduced that CCL22 inactivated STAT5 signals,
which lead to the reduction of the number of Tregs in
CD4+ T cells of RA patients.
Alkaloid SIN, known as an anti-arthritis drug, has
been used in RA therapy for several decades. Li et al
(13) reported that treatment with
SIN reduced the proportion of Th17
(CD4+IL-17+) and elevated the proportion of
Tregs in PBMC of RA patients. Tong et al (18) suggested that SIN treatment suppressed
collagen-induced arthritis by regulating Th17/Treg cells in
intestinal lymph nodes. This study substantiated the promotion
effect of SIN on the number of Tregs and FOXP3 expression in
CD4+ T cells of RA patients in vitro simultaneously with
the decreased CCL22 and CCR4 (Fig.
4). Further experiments showed that recombinant CCL22 and STAT5
inhibitor blocked the effect of SIN (Fig. 5), suggesting that CCL22/CCR4/STAT5
axis mediated the function of SIN on Tregs. Thus, compounds which
can modulate CCL22/CCR4/STAT5 axis may be applied for the treatment
of RA.
In conclusion, CCL22 plays a role in regulating the
number of Tregs and the function, and blocking STAT5 activation is
the underlying mechanism. Drugs targeting CCL22/CCR4/STAT5 axis
might represent the immunomodulatory effect in the long-term
treatment of RA. Our study provides a novel strategy for RA
treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LW (first in the author list) conceived the study
and drafted the manuscript. PH and QC acquired the data; ZZ and LW
(second in the author list) analyzed the data and revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Shuguang Hospital Affiliated to Shanghai University of TCM
(Shanghai, China). Patients who participated in this research had
complete clinical data. Signed informed consents were obtained from
the patients and/or guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kurkó J, Besenyei T, Laki J, Glant TT,
Mikecz K and Szekanecz Z: Szekanecz, immunology, genetics of
rheumatoid arthritis - A comprehensive review. Clin Rev Allergy
Immunol. 45:170–179. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hong Q, Song XY, Huang JY and Guo Y:
CD4+CD25+Foxp3+ regulatory T cell
content in peripheral blood and synovial fluid of patients with
rheumatoid arthritis and osteoarthritis. Zhongguo Shengwuzhipinxue
Zazhi. 24:169–172. 2011.(In Chinese).
|
|
3
|
Bayry J, Sibéril S, Triebel F, Tough DF
and Kaveri SV: Rescuing CD4+CD25+ regulatory
T-cell functions in rheumatoid arthritis by cytokine-targeted
monoclonal antibody therapy. Drug Discov Today. 12:548–552. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Leipe J, Skapenko A, Lipsky PE and
Schulze-Koops H: Regulatory T cells in rheumatoid arthritis.
Arthritis Res Ther. 7:932005. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu Q, Xu M, Yu F and Jin Y: CD4(+)CD25(+)
regulatory T cells as a therapeutic target in rheumatoid arthritis.
Cent Eur J Immunol. 39:100–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yi H and Zhao Y: Chemokines, chemokine
receptors and CD4+CD25+ regulatory T cells.
Expert Rev Clin Immunol. 3:343–349. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Flytlie HA, Hvid M, Lindgreen E,
Kofod-Olsen E, Petersen EL, Jørgensen A, Deleuran M, Vestergaard C
and Deleuran B: Expression of MDC/CCL22 and its receptor CCR4 in
rheumatoid arthritis, psoriatic arthritis and osteoarthritis.
Cytokine. 49:24–29. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Burchill MA, Yang J, Vogtenhuber C, Blazar
BR and Farrar MA: IL-2 receptor beta-dependent STAT5 activation is
required for the development of Foxp3+ regulatory T
cells. J Immunol. 178:280–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mahmud SA, Manlove LS and Farrar MA:
Interleukin-2 and STAT5 in regulatory T cell development and
function. JAK-STAT. 2:e231542013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Passerini L, Allan SE, Battaglia M, Di
Nunzio S, Alstad AN, Levings MK, Roncarolo MG and Bacchetta R:
STAT5-signaling cytokines regulate the expression of FOXP3 in
CD4+CD25+ regulatory T cells and
CD4+CD25− effector T cells. Int Immunol.
20:421–431. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hintzen C, Haan C, Tuckermann JP, Heinrich
PC and Hermanns HM: Oncostatin M-induced and constitutive
activation of the JAK2/STAT5/CIS pathway suppresses CCL1, but not
CCL7 and CCL8, chemokine expression. J Immunol. 181:7341–7349.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Betts BC, Veerapathran A, Pidala J, Yu XZ
and Anasetti C: STAT5 polarization promotes iTregs and suppresses
human T-cell alloresponses while preserving CTL capacity. J Leukoc
Biol. 95:205–213. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li N, Deng Y, Zhou JR, Song GZ and Wang
JY: Effect of sinomenine on the proportion of T helpers 17 and
regulatory T cells in puerperal blood mononuclear cells from
patients with rheumatoid arthritis. Shanghai Med J. 36:254–258.
2013.
|
|
14
|
Fahy O, Porte H, Sénéchal S, Vorng H,
McEuen AR, Buckley MG, Walls AF, Wallaert B, Tonnel AB and
Tsicopoulos A: Chemokine-induced cutaneous inflammatory cell
infiltration in a model of Hu-PBMC-SCID mice grafted with human
skin. Am J Pathol. 158:1053–1063. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nishikawa H and Sakaguchi S: Regulatory T
cells in tumor immunity. Int J Cancer. 127:759–767. 2010.PubMed/NCBI
|
|
16
|
Antov A, Yang L, Vig M, Baltimore D and
Van Parijs L: Essential role for STAT5 signaling in
CD25+CD4+ regulatory T cell homeostasis and
the maintenance of self-tolerance. J Immunol. 171:3435–3441. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang Z, Chen L, Yang B, Chen J and Wang
L: Decreased phosphostat5 contributed to impairment of
CD4+CD25+Foxp3+ Tregs in RA
patients. Ann Rheum Dis. 75 (Suppl 2):9872016. View Article : Google Scholar
|
|
18
|
Tong B, Yu J, Wang T, Dou Y, Wu X, Kong L,
Dai Y and Xia Y: Sinomenine suppresses collagen-induced arthritis
by reciprocal modulation of regulatory T cells and Th17 cells in
gut-associated lymphoid tissues. Mol Immunol. 65:94–103. 2015.
View Article : Google Scholar : PubMed/NCBI
|