Introduction
Prednisolone is a corticosteroid drug, predominantly
comprising glucocorticoid, and is a widely used therapeutic for
immune suppression (1). The drug
abrogates the expression of inflammatory genes by inhibiting the
transcriptional promoting activity of the AP-1 and NF-κB
transcription factors, and also enhances the release of
anti-inflammatory proteins such as IL-4, IL-13 or IL-10 (2,3),
suggesting that prednisolone modulates tissue responses by
regulating gene regulation at sites of inflammation (4). Data of animal studies suggest that
prednisolone, the active metabolite of prednisone, exerts its
pharmacological effects in cholesterol-rich environments.
Prednisone, which is metabolized by the liver to prednisolone,
inhibits development of inflammatory lesions in the aortas of
cholesterol-fed rabbits, without lowering serum cholesterol levels
(5). Site-specific targeting of
nanoparticles of prednisolone reduces inflammation and formation of
neo-intima in a high cholesterol diet rabbit model of established
atheroma (6). However, it is yet to
be established how prednisolone affects tissue responses occurring
in a milieu rich in cholesterol molecules.
Oxysterols, the oxygenated derivatives of
cholesterol, induce expression of inflammatory molecules by
monocytes/macrophages and are recognized as strong inducers of
inflammation (7–9). 27-Hydroxycholesterol (27OHChol) is the
most abundant oxysterol in circulation and tissues under
hypercholesterolemic conditions (10). 27OHChol promotes migration of
monocytic cells and T lymphocytes expressing CCR5 (11,12),
enhances the production of molecules involved in various
inflammatory processes, including TNF-α and CXCL8 (13–15), and
increases the expression of MHC I and II molecules and pattern
recognition receptors, thereby augmenting responses of immune cells
to pathogen-associated molecular patterns (16–18).
These findings suggest that 27OHChol steers macrophages/monocytes
to an immunostimulatory phenotype.
As the key innate immune effector cells, macrophages
are highly heterogeneous and are capable of rapidly changing their
functions in response to local microenvironmental signals (19). The activated macrophages (M1) driven
by interferon-γ and lipopolysaccharide (LPS) release inflammatory
and immunostimulatory cytokines (20). The alternatively activated
macrophages (M2) are elicited by IL-4, immune complexes, or
glucocorticoids, in combination or not with transforming growth
factor-β, and act to restrict these inflammatory responses through
secretion of immunoregulatory cytokines (20), thereby affecting angiogenesis and
invasiveness (21). Controlling the
macrophage polarization results in altered disease progression,
indicating that macrophage polarization serves as a novel
therapeutic approach.
In the present study, we used human THP-1 monocytic
cells to examine whether prednisolone modifies the
27OHChol-mediated polarization and responses of
monocytes/macrophages. Dexamethasone was employed as a positive
control because it is a potent, long-acting steroid product and is
reported to modify cellular responses to oxysterol (22,23).
This study determines the new biological activities of prednisolone
that contribute to pharmacological effects of the drug.
Materials and methods
Cells and reagents
The human THP-1 monocyte/macrophage cell line was
purchased from the American Type Culture Collection (ATCC) and
maintained as suggested by the ATCC. Prednisolone and LPS were
purchased from Sigma-Aldrich and InvivoGen, respectively. 27OHChol
and antibodies against CD14, p65, phosphorylated p65, and β-actin
were obtained from Santa Cruz Biotechnology Inc.
Reverse transcription (RT) -
polymerase chain reaction (PCR)
Total RNA was reverse-transcribed for 1 h at 42°C
using the Moloney murine leukemia virus reverse transcriptase, and
real-time quantitative PCR was performed in triplicate using a
LightCycler® 96 Real-Time PCR System (Roche), as
previously reported (24). Each 20
µl reaction mixture consisted of 10 µl of SYBR Green Master Mix and
2 µl each of 10 pM forward and reverse primers of the gene to be
quantified. The thermal cycling conditions were as follows: 95°C
for 10 min, followed by 45 cycles at 95°C for 10 sec, 50°C for 10
sec, and 72°C for 10 sec. The LightCycler® 96 software
(v1.1.0.1320; Roche) was applied to calculate the relative
expression of each gene as the ratio to the housekeeping gene,
glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Target gene mRNA
levels were normalized to GAPDH using the 2−ΔΔCt method
(25). The primers used for
real-time PCR were as follows: TNF-α, 5′-CCCAGGGACCTCTCTCTAATC-3′
(forward) and 5′-ATGGGCTACAGGCTTGTCACT-3′ (reverse); IL-1β,
5′-TGAGCTCGCCAGTGAAATGA (forward) and 5′-AGATTCGTAGCTGGATGCCG-3′
(reverse); CXCL10, 5′-TGTACGCTGTACCTGCATCA-3′ (forward) and
5′-GGACAAAATTGGCTTGCAGGA-3′ (reverse); CXCL11,
5′-AAGCAGTGAAAGTGGCAGAT-3′ (forward) and 5′-TAAGCCTTGCTTGCTTCGAT-3′
(reverse); CD80, 5′-GCAGGGAACATCACCATCCA-3′ (forward) and
5′-TCACGTGGATAACACCTGAACA-3′ (reverse); CD86,
5′-GGACTAGCACAGACACACGGA-3′ (forward) and
5′-CTTCAGAGGAGCAGCACCAGA-3′ (reverse); CD163,
5′-AAAAAGCCACAACAGGTCGC-3′ (forward) and 5′-CTTGAGGAAACTGCAAGCCG-3′
(reverse); CD206, 5′-TGAATTGTACTGGTCTGTCCT-3′ (forward) and
5′-CTGTGGTGCTGTGCATTTATCT-3′ (reverse); CCL2,
5′-CAGCCAGATGCAATCAATGCC-3′ (forward) and
5′-TGGAATCCTGAACCCACTTCT-3′ (reverse); matrix metalloprotease-9
(MMP-9), 5′-GCACGACGTCTTCCAGTACC-3′ (forward) and
5′-CAGGATGTCATAGGTCACGTAGC-3′ (reverse); CD14,
5′-ACGCCAGAACCTTGTGAGC-3′ (forward) and
5′-GCATGGATCTCCACCTCTACTG-3′ (reverse); and GAPDH,
5′-GAAGGTGAAGGTCGGAGT-3′ (forward) and 5′-GAAGATGGTGATGGGATTTC-3′
(reverse).
Chemotaxis assay
Cell migration was measured using Transwell
Permeable Supports (Costar) as previously described (12). Briefly, 5×105 cells in 100
µl 0.1% BSA were loaded into the top chamber of 5-µm-pore
polycarbonate transwell inserts. Transwell chambers were inserted
into wells filled with supernatant harvested from THP-1 cells
treated with 27OHChol, with or without prednisolone. After
incubation for 3 h at 37°C, the number of cells that migrate to the
bottom chamber was counted using a Vi-Cell cell counter (Beckman
Coulter, Inc.).
MMP-9 gelatinolytic activity in
supernatants
Supernatants isolated from THP-1 cells were
collected and concentrated 30-fold using Vivaspin 2 Centricon, as
previously described (26). The
concentrated medium was then electrophoretically separated onto an
8% polyacrylamide gel containing 0.15% gelatin. After
electrophoresis, the gel was washed, activated, and stained with
0.2% Coomassie brilliant blue R-250, followed by destaining. Clear
zones against the blue background indicate gelatinolytic
activity.
Flow cytometric analysis
Surface levels of CD molecules were evaluated as
previously reported (18). Briefly,
THP-1 cells were harvested by centrifugation, followed by
incubation for 40 min at 4°C with anti-CD14 antibody conjugated
with a green fluorescent dye (Santa Cruz Biotechnology Inc.), FITC
anti-human CD163 and PE anti-human CD206 (BioLegend). After washing
twice with phosphate-buffered saline (PBS), cells were re-suspended
in 1% paraformaldehyde in PBS. Fluorescence was analyzed by flow
cytometry.
Enzyme-linked immunosorbent assay
The levels of CCL2, sCD14, and MMP-9 secreted into
the culture media were determined using commercially available
enzyme-linked immunosorbent assay (ELISA) kits as per the
manufacturer's instructions (R&D Systems), following the
previously described protocols (17).
Western blot analysis
Cell lysates were separated by 10% SDS-PAGE and
subsequently transferred to nitrocellulose membranes. After
blocking for 1 h in 1% skim milk (prepared in TBS containing 0.05%
Tween-20), membranes were incubated overnight at 4°C, with primary
antibodies against CD14, phosphorylated p65, p65 subunit, or
β-actin. After three washes with TBS-T, the membranes were
incubated for 1 h with HRP-conjugated secondary antibodies at room
temperature. Bands were detected using chemiluminescent detection
reagents.
Statistical analysis
One-way ANOVA followed by Tukey's multiple
comparison tests were performed using PRISM (version 5.0) (GraphPad
Software Inc.). Data are presented as the mean ± SD, and are
representative of three independent experiments. Null hypotheses of
no difference were rejected for P-values less than 0.05.
Results
Prednisolone inhibits the expression
of M1 markers in monocytes/macrophages
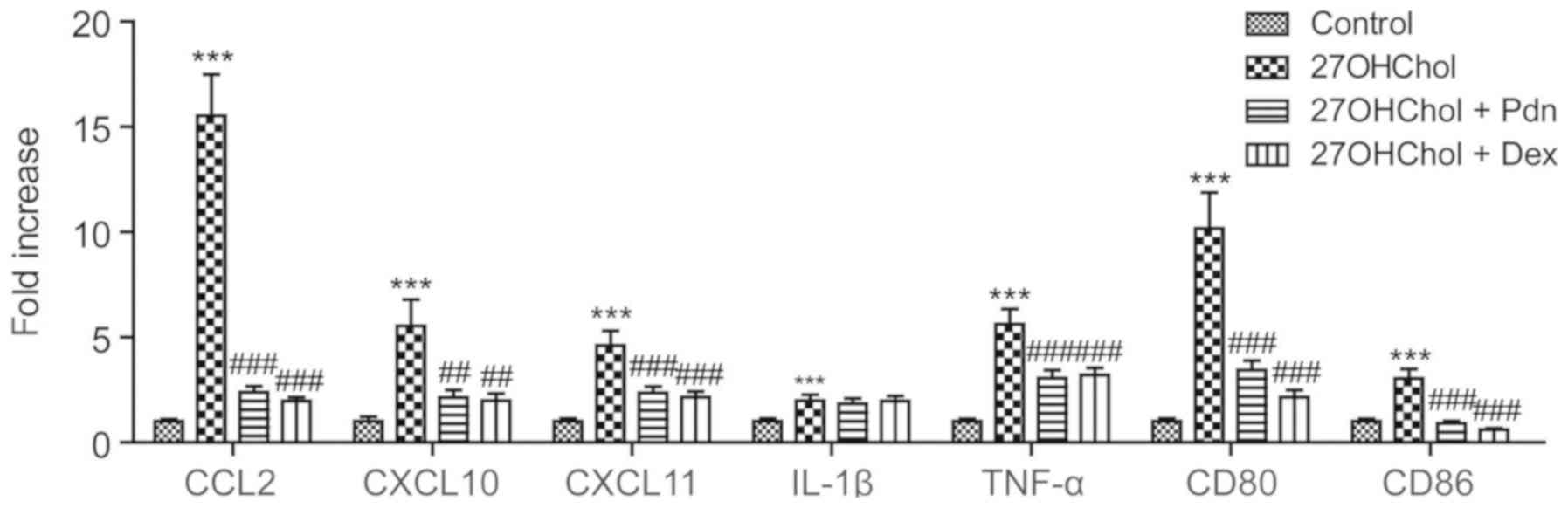
We determined whether prednisolone affects the
expression of the M1 phenotype markers (Fig. 1). Stimulation of monocyte/macrophage
cells with 27OHChol results in increased expression of molecules
associated with the M1 phenotype, such as CCL2, CXCL10, CXCL11,
IL-1β, TNF-α, CD80 and CD86, but the 27OHChol-induced expression of
M1 markers (except IL-1β) is significantly suppressed following
exposure to prednisolone. The inhibition of M1 marker expressions
is similar to that exerted by dexamethasone, which is used as a
positive control due to its potent, long-acting activity. These
results indicate that prednisolone regulates polarization to the M1
phenotype.
Prednisolone impairs migration of
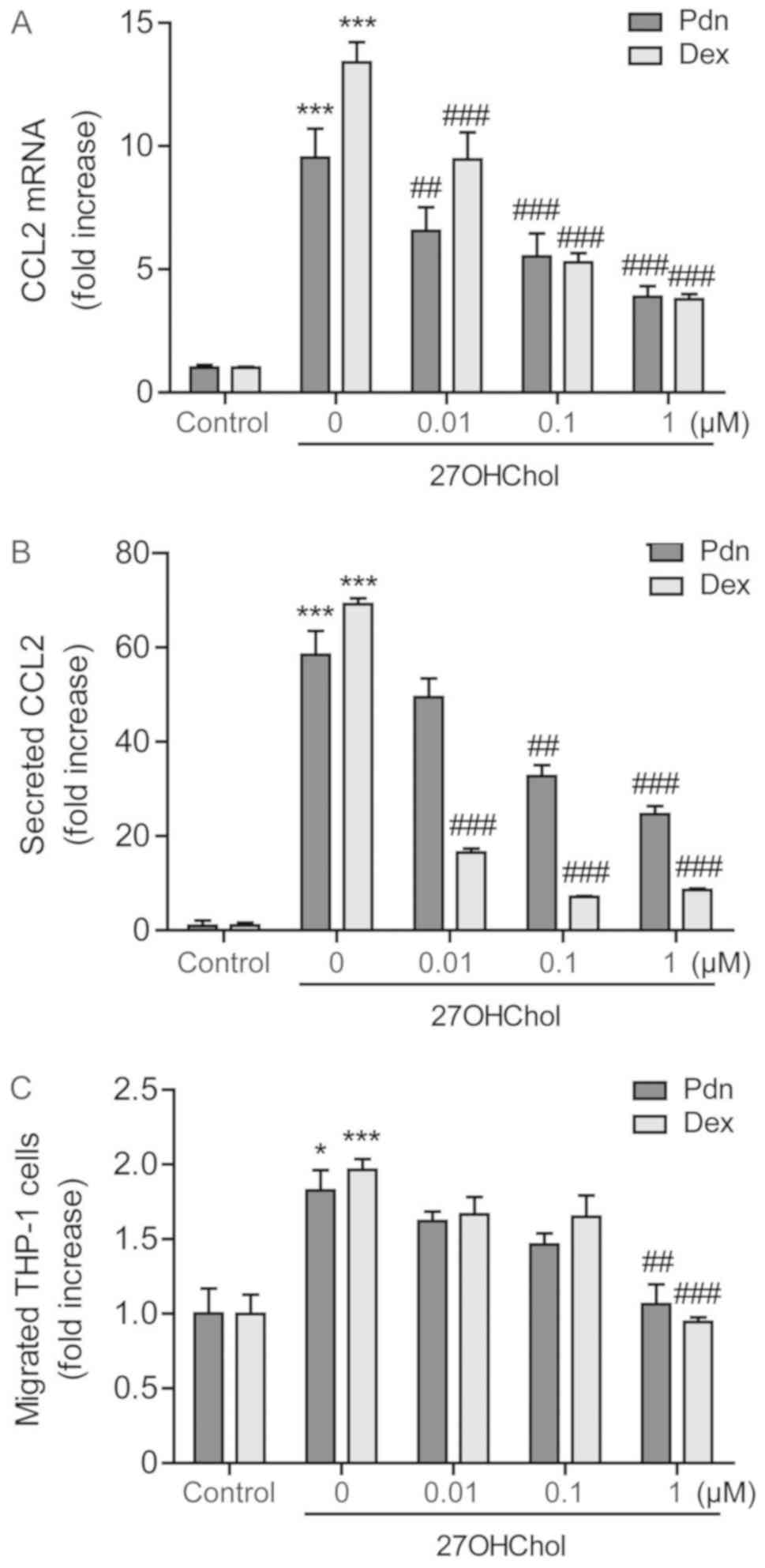
monocytic cells via inhibiting CCL2 expression
Since CCL2 is the key M1 molecule regulating
migration of monocytes/macrophages, we examined the effects of
prednisolone, in parallel with dexamethasone, on CCL2 expression at
the transcriptional and protein levels. The 27OHChol-induced
transcription of CCL2 is attenuated in a dose-dependent manner
after treatment with prednisolone, and which is comparable to that
obtained with dexamethasone (Fig.
2A). Exposure to prednisolone also significantly reduces the
amount of secreted CCL2. Of the two steroids, CCL2 is reduced to a
greater extent after exposure to dexamethasone, indicating that
dexamethasone is more efficacious in inhibiting the CCL2 secretion
(Fig. 2B). We further determined the
influence of prednisolone on cell migration. Significant increase
of monocytic cell migration is observed in response to the
supernatants harvested following stimulation with 27OHChol. The
cell migration is reduced when cells are exposed to supernatants
isolated from cells cultured with 27OHChol plus 1 µM prednisolone
or dexamethasone. The reduction caused by prednisolone is
comparable to that obtained by dexamethasone (Fig. 2C). These results indicate the
impairment of 27OHChol-induced CCL2 expression and cell migration
following exposure to prednisolone.
Prednisolone down-regulates
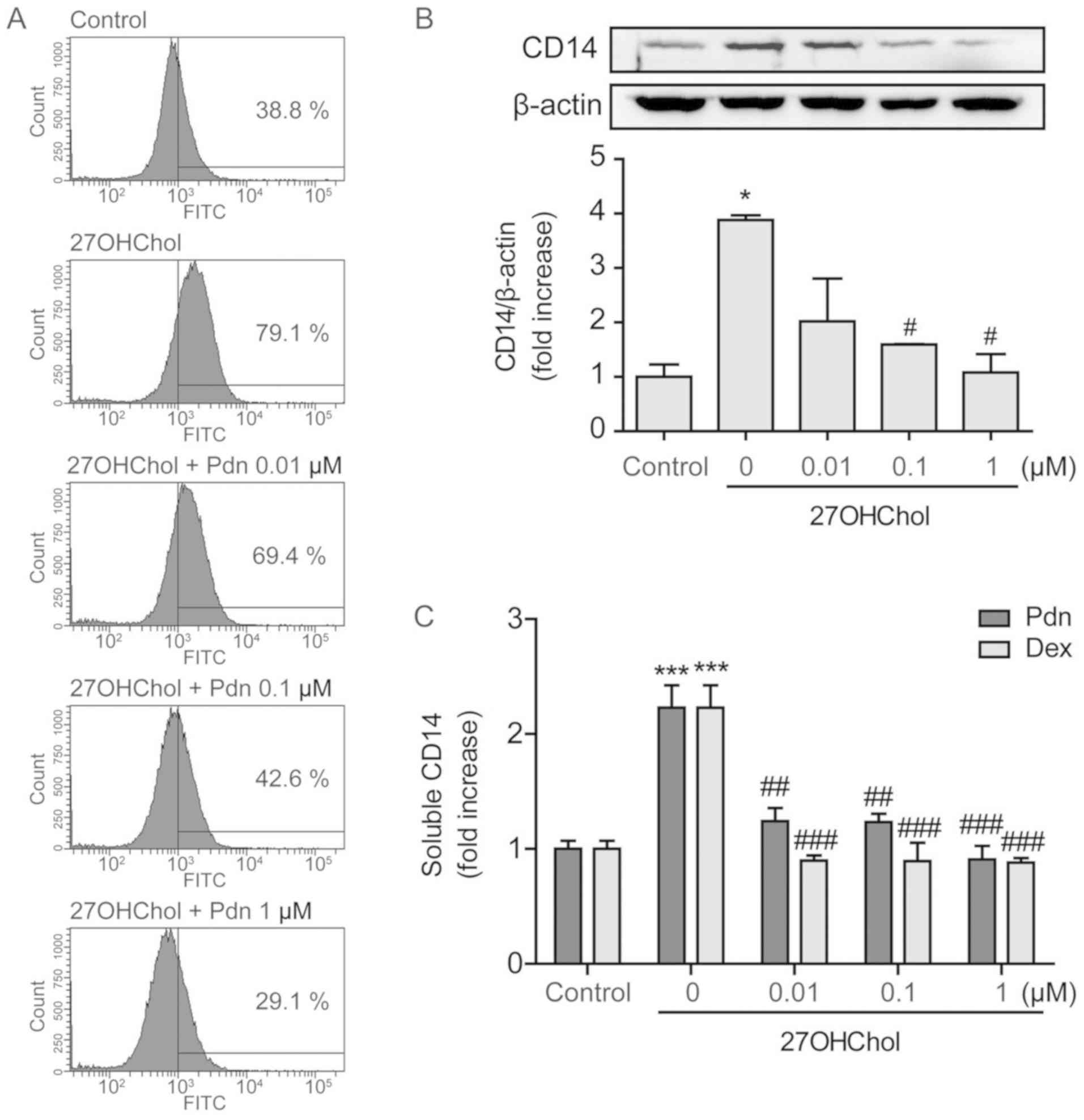
27OHChol-induced CD14 expression and weakens LPS response
We next investigated whether prednisolone influences
the expression of CD14. Stimulation with 27OHChol results in
upregulation of CD14 on the monocyte/macrophage cell surface, as
indicated by the increased percentage of CD14-positive cells, but
the percentage decreases in a dose-dependent manner after treatment
with prednisolone (Fig. 3A). The
levels of CD14 protein were also evaluated using Western blot
analysis. 27OHChol exposure increases the levels of cellular CD14
protein, which reduce to the basal level and lower, following
treatment with 0.1 and 1 µM of prednisolone, respectively (Fig. 3B). However, we were unable to obtain
conclusive data that prednisolone affects the levels of CD14
transcripts (Fig. S1). We further
investigated the effects of prednisolone on secretion of soluble
CD14 (sCD14). The 27OHChol-induced sCD14 release is almost
completely inhibited after treatment with prednisolone, which is
comparable to results obtained with dexamethasone (Fig. 3C). These results indicate that
prednisolone affects CD14 expression at the protein level.
The effects of prednisolone were also determined on
LPS stimulation, by measuring the CCL2 expression (Fig. S2). Levels of CCL2 transcripts were
observed to increase 33.8- and 11.1-folds after stimulation with
27OHChol and LPS, respectively. Addition of LPS to 27OHChol-exposed
cells resulted in further elevation of CCL2 transcripts by 115.4-
fold, which reduces to 79.2-, 29.8- and 9.8-folds in the presence
of 0.01, 0.1 and 1 µM of prednisolone, respectively. Collectively,
our data indicate that prednisolone down-regulates CD14 and thereby
inhibits the 27OHChol-enhanced LPS response.
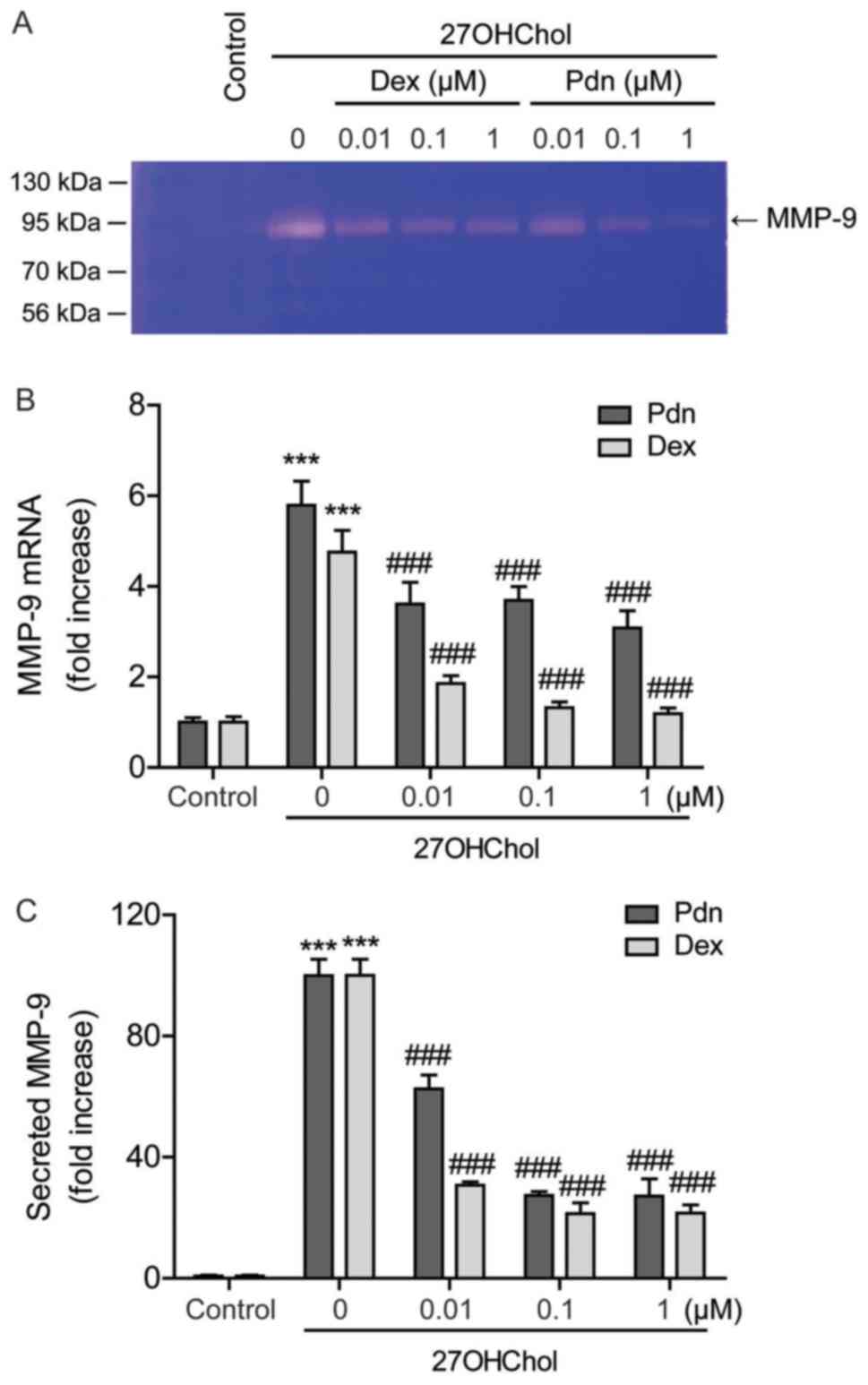
Prednisolone inhibits 27OHChol-induced
MMP-9 expression
Since prednisolone inhibits the sCD14 release, we
evaluated the effects of the drug on MMP-9 activity. 27OHChol
enhances MMP-9 activity in the cell supernatant, which decreases
after exposure to prednisolone, as demonstrated by gelatin
zymography (Fig. 4A). Furthermore,
evaluating the effects of prednisolone on MMP-9 expression reveals
elevated levels of MMP-9 transcripts after 27OHChol exposure, which
is suppressed by treatment with prednisolone (Fig. 4B). The 27OHChol-induced MMP-9
secretion is also significantly suppressed following treatment with
prednisolone, as determined by ELISA (Fig. 4C). Compared to prednisolone,
dexamethasone inhibits the transcription and secretion of MMP-9
with higher-potency (Fig. 4B and C).
These results indicate that although less effective than
dexamethasone, prednisolone inhibits MMP-9 expression at the
transcriptional and protein levels.
Prednisolone regulates molecular
signaling enhanced by 27OHChol
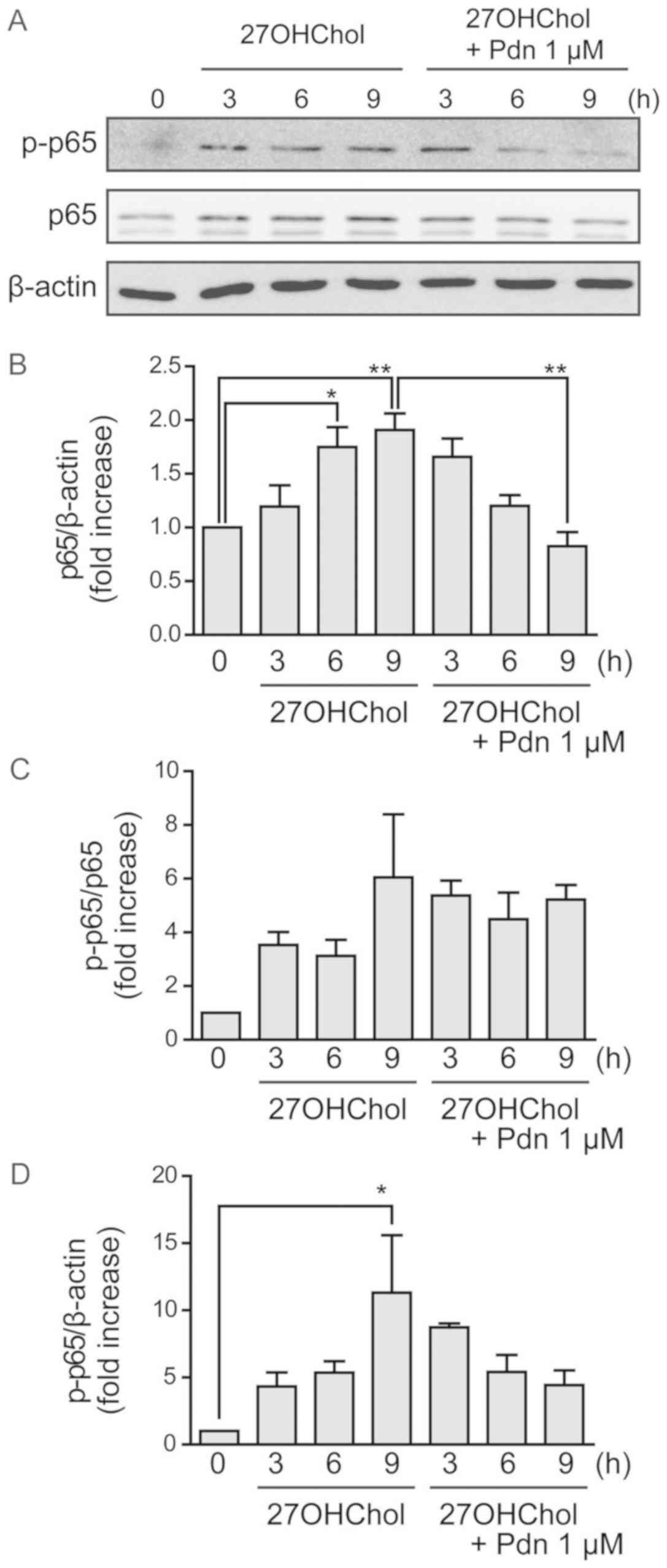
We investigated the effects of prednisolone on
expression levels of the NF-κB p65 subunit and its phosphorylated
form by performing Western blot analyses. 27OHChol increases the
levels of p65 subunit, which is suppressed following treatment with
prednisolone (Fig. 5A and B). The
phosphorylated form of p65 (p-p65) may only elevate because total
p65 expression increases, rather than increased p65 phosphorylation
(Fig. 5C and D). These results
suggest that prednisolone suppresses the 27OHChol-induced activity
of the transcription factor NF-κB.
Prednisolone affects transcript levels
of CD163 and CD206
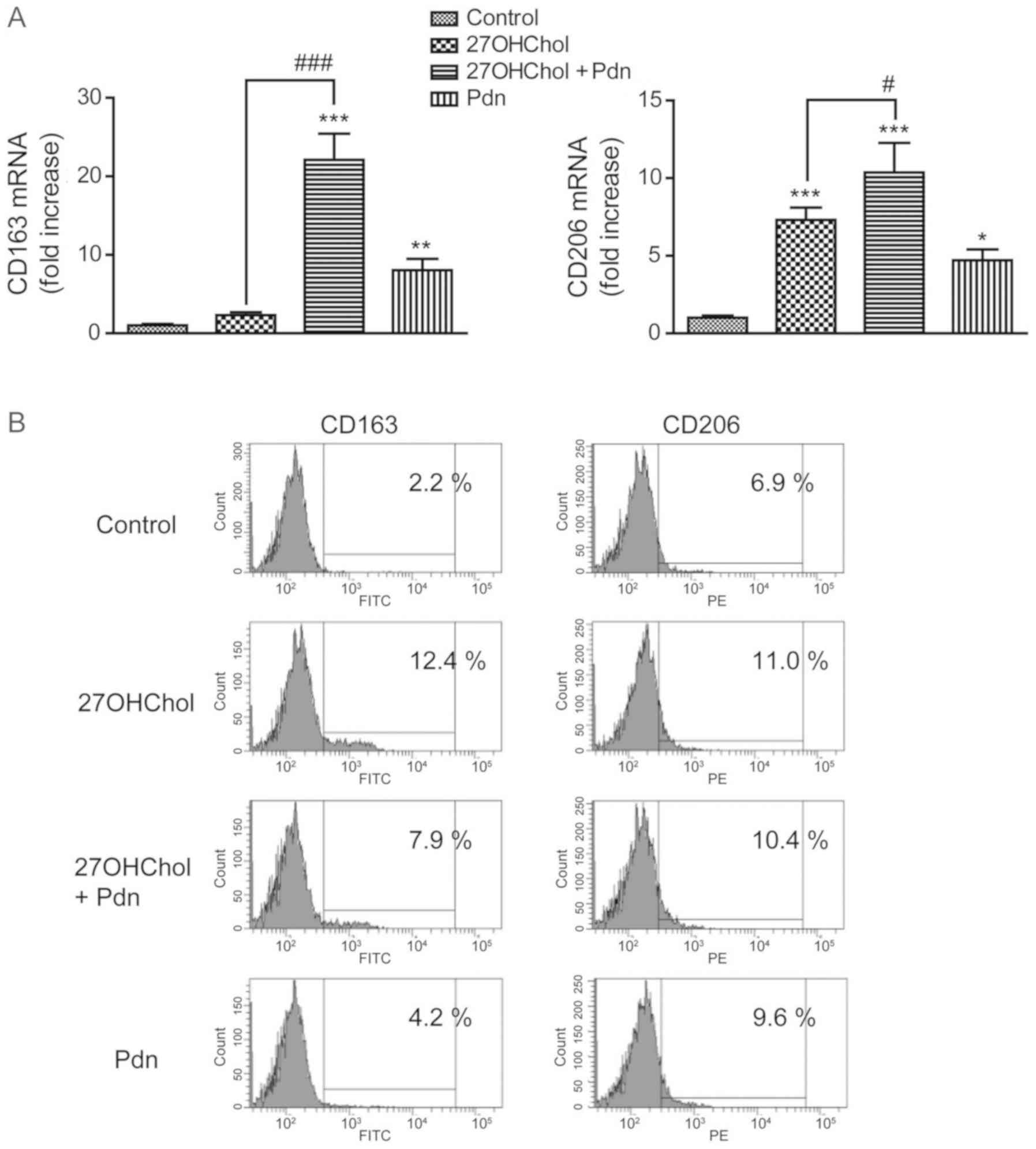
We subsequently examined the effects of prednisolone
on the 27OHChol-induced expression of M2 markers (Fig. 6). Prednisolone exposure increases the
transcript levels of CD163 and augments the 27OHChol-induced
transcription of the CD163 gene. However, although an increase is
observed in the transcript levels of CD206, there was no
amplification in the 27OHChol-induced transcription of CD206 gene;
the transcript levels of CD206 following cotreatment with 27OHChol
and prednisolone were comparable to the summation of the levels
observed with each treatment alone (Fig.
6A). We further determined whether prednisolone affects the
expression levels of CD163 and CD206 molecules on the cell surface
(Fig. 6B). We observed an increase
in the surface levels of CD163 and CD206 after exposure to 27OHChol
or prednisolone. Investigating the levels of CD68 after 27OHChol
and prednisolone exposure shows no increase in the expression of
CD68 (Fig. S3). Taken together,
these results suggest that prednisolone differentially regulates
the transcript levels and surface expression of M2 makers in the
presence of 27OHChol.
Discussion
27OHChol enhances the expression of
anti-inflammatory and inflammatory molecules, and cytokines and
chemokines of monocytic cells (12,27,28),
indicating that 27OHChol is involved in the polarization of
monocytes/macrophages. However, the effects of 27OHChol on M1/M2
polarization have been elusive. We therefore investigated the
expressions of M1 and M2 markers, to understand the overall
influence of 27OHChol on monocyte/macrophage polarization.
Increases in M1 markers validate that 27OHChol is an active
molecule with inflammatory functions, whereas the increased
transcription of M2 markers (such as CD163 and CD206) (24) is in agreement with a previous study
that reported polarization of human macrophages toward the M2
immunomodulatory phenotype after short-term exposure to this
oxysterol (27). These findings
suggest that 27OHChol affects both M1 and M2 polarization of
monocytes/macrophages. Besides, 27OHChol does not change CD68
expression, which agrees with the fact that CD68 is expressed both
in M1 and M2 macrophages (29), and
indicates that 27OHChol is unlikely to cause differentiation or
polarization of monocytic cells into other lineages because CD68
can be used as a pan-macrophage marker. Of the two markers,
expression of M1 markers is more strongly enhanced following
treatment with 27OHChol. The preferential expression of M1 markers
could help explain the dominance of immunostimulatory and
inflammatory responses in a milieu rich in 27OHChol in spite of its
liver X receptor agonistic activity which suppresses inflammatory
signaling in macrophages (30,31).
We next attempted to determine the effects of
prednisolone on M1/M2 polarization under hypercholesterolemic
conditions. We observed suppressed expression of 27OHChol-induced
M1 markers, and upregulation of the transcription and cell surface
expression of CD163 and CD206, without further enhancement of the
27OHChol-induced expression of molecules. These results are
consistent with previous reports that glucocorticoids generate M2
macrophages (20) and enhance
transdifferentiation of macrophages towards the immune regulatory
M2 phenotype (32). Taken together
with previous publications, our findings suggest that prednisolone
differentially regulates M1/M2 polarization of
monocytes/macrophages. The differential effects are likely to
contribute to the immune suppressive activity of the drugs in
27OHChol-rich conditions.
Prednisolone and dexamethasone exhibit not only high
effectiveness but also differences in their potency with respect to
suppression of 27OHChol-mediated immune stimulation. Dexamethasone
more effectively inhibits secretion of CCL2 and transcription of
MMP-9, than prednisolone. We believe that the differences in the
inhibitory activity are in line with pharmacokinetics and
pharmacological activity of the drugs. Prednisolone is an
intermediate acting steroid with a half-life of 12 to 36 h, whereas
dexamethasone is a long acting corticosteroid with a biological
half-life between 36 and 72 h; furthermore, dexamethasone is five
to six times as potent as prednisolone in terms of
anti-inflammatory potential (23,33).
NF-κB is one of the most important regulators of
pro-inflammatory gene expressions such as TNF-α, IL-1β, and IL-6
(34); also, the activation of
macrophages in response to multiple M1 polarizing stimuli is
regulated primarily via NF-κB (35,36).
Therefore, we investigated the possible involvement of NF-κB in
27OHChol-induced M1 polarization. Increased expression of M1
markers coincides with enhanced phosphorylation of the p65 subunit
of NF-κB following 27OHChol treatment, and prednisolone suppresses
both the expression of M1 markers and phosphorylation of p65,
without affecting M2 markers. These results suggest a correlation
between activity of inducible NF-κB and regulation of M1
polarization in the presence of 27OHChol and prednisolone.
During an inflammatory response, the expression and
secretion of MMP-9 is elevated by macrophages, and its activity is
required for migration of macrophages (37). CCL2 is a key molecule recruiting
monocytes to the sites of inflammation (38). Prednisolone not only suppresses MMP-9
expression but also decreases monocytic cell migration coupled with
CCL2 production. These results are in line with the findings by
Wong et al, who reported decreased MMP-9 expression in
macrophages and reduced infiltration of inflammatory cells
following treatment with prednisolone (39). MMP-9 is also involved in the
post-translational processing of CD14. Proteolytic cleavage of
mCD14 by MMP-9 results in sCD14 shedding (40). CD14 binds to LPS, and the LPS-CD14
complex triggers macrophage activation, culminating in inflammatory
responses by enhancing the production of multiple inflammatory
molecules (41). Our studies
determined that prednisolone down-regulates CD14 and attenuates the
LPS response. Taken together, these results indicate that MMP-9 may
be one of the key molecules that mediate the 27OHChol-induced
inflammatory and immune responses.
This study reports a new pharmacological effect of
prednisolone: The differential regulation of M1 and M2 markers in a
milieu rich in 27OHChol. We believe that differential regulation of
M1/M2 polarization of monocyte/macrophage cells is a promising
strategy for suppression of the immune reactions activated due to
cholesterol oxidation products.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Biomedical
Research Institute of Pusan National University Hospital (grant no.
2019B004).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
KK and MSK designed the study and analyzed the data.
BYK and YS performed the experiments and analyzed the data. BYK and
KK wrote the manuscript. All authors approved the final version of
the manuscript for publication.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Czock D, Keller F, Rasche FM and Häussler
U: Pharmacokinetics and pharmacodynamics of systemically
administered glucocorticoids. Clin Pharmacokinet. 44:61–98. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blotta MH, DeKruyff RH and Umetsu DT:
Corticosteroids inhibit IL-12 production in human monocytes and
enhance their capacity to induce IL-4 synthesis in CD4+
lymphocytes. J Immunol. 158:5589–5595. 1997.PubMed/NCBI
|
|
3
|
Chung KF and Adcock IM: Signalling and
transcriptional regulation in inflammatory and immune cells:
Importance in lung biology and disease. Eur Respir J. 26:762–763.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Parrillo JE and Fauci AS: Mechanisms of
glucocorticoid action on immune processes. Annu Rev Pharmacol
Toxicol. 19:179–201. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bailey JM and Butler J: Anti-inflammatory
drugs in experimental atherosclerosis. I. Relative potencies for
inhibiting plaque formation. Atherosclerosis. 17:515–522. 1973.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Joner M, Morimoto K, Kasukawa H,
Steigerwald K, Merl S, Nakazawa G, John MC, Finn AV, Acampado E,
Kolodgie FD, et al: Site-specific targeting of nanoparticle
prednisolone reduces in-stent restenosis in a rabbit model of
established atheroma. Arterioscler Thromb Vasc Biol. 28:1960–1966.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lemaire-Ewing S, Prunet C, Montange T,
Vejux A, Berthier A, Bessède G, Corcos L, Gambert P, Néel D and
Lizard G: Comparison of the cytotoxic, pro-oxidant and
pro-inflammatory characteristics of different oxysterols. Cell Biol
Toxicol. 21:97–114. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
van Reyk DM, Brown AJ, Hult'en LM, Dean RT
and Jessup W: Oxysterols in biological systems: Sources, metabolism
and pathophysiological relevance. Redox Rep. 11:255–262. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vejux A and Lizard G: Cytotoxic effects of
oxysterols associated with human diseases: Induction of cell death
(apoptosis and/or oncosis), oxidative and inflammatory activities,
and phospholipidosis. Mol Aspects Med. 30:153–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carpenter KL, Taylor SE, Ballantine JA,
Fussell B, Halliwell B and Mitchinson MJ: Lipids and oxidised
lipids in human atheroma and normal aorta. Biochim Biophys Acta.
1167:121–130. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim SM, Kim BY, Lee SA, Eo SK, Yun Y, Kim
CD and Kim K: 27-Hydroxycholesterol and 7alpha-hydroxycholesterol
trigger a sequence of events leading to migration of
CCR5-expressing Th1 lymphocytes. Toxicol Appl Pharmacol.
274:462–470. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim SM, Lee SA, Kim BY, Bae SS, Eo SK and
Kim K: 27-Hydroxycholesterol induces recruitment of monocytic cells
by enhancing CCL2 production. Biochem Biophys Res Commun.
442:159–164. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jurisic V, Terzic T, Colic S and Jurisic
M: The concentration of TNF-alpha correlate with number of
inflammatory cells and degree of vascularization in radicular
cysts. Oral Dis. 14:600–605. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim SM, Jang H, Son Y, Lee SA, Bae SS,
Park YC, Eo SK and Kim K: 27-Hydroxycholesterol induces production
of tumor necrosis factor-alpha from macrophages. Biochem Biophys
Res Commun. 430:454–459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim SM, Lee CW, Kim BY, Jung YS, Eo SK,
Park YC and Kim K: 27-Oxygenated cholesterol induces expression of
CXCL8 in macrophages via NF-kappaB and CD88. Biochem Biophys Res
Commun. 463:1152–1158. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Heo W, Kim SM, Eo SK, Rhim BY and Kim K:
FSL-1, a toll-like receptor 2/6 agonist, induces expression of
interleukin-1α in the presence of 27-hydroxycholesterol. Korean J
Physiol Pharmacol. 18:475–480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim SM, Kim BY, Eo SK, Kim CD and Kim K:
27-Hydroxycholesterol up-regulates CD14 and predisposes monocytic
cells to superproduction of CCL2 in response to lipopolysaccharide.
Biochim Biophys Acta. 1852:442–450. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Son Y, Kim SM, Lee SA, Eo SK and Kim K:
Oxysterols induce transition of monocytic cells to phenotypically
mature dendritic cell-like cells. Biochem Biophys Res Commun.
438:161–168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Murray PJ and Wynn TA: Protective and
pathogenic functions of macrophage subsets. Nat Rev Immunol.
11:723–737. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Colvin EK: Tumor-associated macrophages
contribute to tumor progression in ovarian cancer. Front Oncol.
4:1372014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takeuchi H, Tanaka M, Tanaka A, Tsunemi A
and Yamamoto H: Predominance of M2-polarized macrophages in bladder
cancer affects angiogenesis, tumor grade and invasiveness. Oncol
Lett. 11:3403–3408. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Son Y, Kim BY, Eo SK, Park YC and Kim K:
Dexamethasone suppresses oxysterol-induced differentiation of
monocytic cells. Oxid Med Cell Longev. 2016:29153822016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sparrow A and Geelhoed G: Prednisolone
versus dexamethasone in croup: A randomised equivalence trial. Arch
Dis Child. 91:580–583. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee J, Kim BY, Son Y, Giang DH, Lee D, Eo
SK and Kim K: 4′-O-Methylalpinumisoflavone inhibits the activation
of monocytes/macrophages to an immunostimulatory phenotype induced
by 27-hydroxycholesterol. Int J Mol Med. 43:2177–2186.
2019.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Radenkovic S, Konjevic G, Jurisic V,
Karadzic K, Nikitovic M and Gopcevic K: Values of MMP-2 and MMP-9
in tumor tissue of basal-like breast cancer patients. Cell Biochem
Biophys. 68:143–152. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Marengo B, Bellora F, Ricciarelli R, De
Ciucis C, Furfaro A, Leardi R, Colla R, Pacini D, Traverso N,
Moretta A, et al: Oxysterol mixture and, in particular,
27-hydroxycholesterol drive M2 polarization of human macrophages.
Biofactors. 42:80–92. 2016.PubMed/NCBI
|
|
28
|
Umetani M, Ghosh P, Ishikawa T, Umetani J,
Ahmed M, Mineo C and Shaul PW: The cholesterol metabolite
27-hydroxycholesterol promotes atherosclerosis via proinflammatory
processes mediated by estrogen receptor alpha. Cell Metab.
20:172–182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bertani FR, Mozetic P, Fioramonti M,
Iuliani M, Ribelli G, Pantano F, Santini D, Tonini G, Trombetta M,
Businaro L, et al: Classification of M1/M2-polarized human
macrophages by label-free hyperspectral reflectance confocal
microscopy and multivariate analysis. Sci Rep. 7:89652017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fu X, Menke JG, Chen Y, Zhou G, MacNaul
KL, Wright SD, Sparrow CP and Lund EG: 27-hydroxycholesterol is an
endogenous ligand for liver X receptor in cholesterol-loaded cells.
J Biol Chem. 276:38378–38387. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schulman IG: Liver X receptors link lipid
metabolism and inflammation. FEBS Lett. 591:2978–2991. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Paulus P, Holfeld J, Urbschat A, Mutlak H,
Ockelmann PA, Tacke S, Zacharowski K, Reissig C, Stay D and
Scheller B: Prednisolone as preservation additive prevents from
ischemia reperfusion injury in a rat model of orthotopic lung
transplantation. PLoS One. 8:e732982013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ito C, Evans WE, McNinch L, Coustan-Smith
E, Mahmoud H, Pui CH and Campana D: Comparative cytotoxicity of
dexamethasone and prednisolone in childhood acute lymphoblastic
leukemia. J Clin Oncol. 14:2370–2376. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tak PP and Firestein GS: NF-kappaB: A key
role in inflammatory diseases. J Clin Invest. 107:7–11. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mantovani A, Sica A, Sozzani S, Allavena
P, Vecchi A and Locati M: The chemokine system in diverse forms of
macrophage activation and polarization. Trends Immunol. 25:677–686.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kawai T and Akira S: Signaling to
NF-kappaB by Toll-like receptors. Trends Mol Med. 13:460–469. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hanania R, Sun HS, Xu K, Pustylnik S,
Jeganathan S and Harrison RE: Classically activated macrophages use
stable microtubules for matrix metalloproteinase-9 (MMP-9)
secretion. J Biol Chem. 287:8468–8483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Deshmane SL, Kremlev S, Amini S and Sawaya
BE: Monocyte chemoattractant protein-1 (MCP-1): An overview. J
Interferon Cytokine Res. 29:313–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wong C, Bezhaeva T, Rothuizen TC,
Metselaar JM, de Vries MR, Verbeek FP, Vahrmeijer AL, Wezel A, van
Zonneveld AJ, Rabelink TJ, et al: Liposomal prednisolone inhibits
vascular inflammation and enhances venous outward remodeling in a
murine arteriovenous fistula model. Sci Rep. 6:304392016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Senft AP, Korfhagen TR, Whitsett JA,
Shapiro SD and LeVine AM: Surfactant protein-D regulates soluble
CD14 through matrix metalloproteinase-12. J Immunol. 174:4953–4959.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kielian TL and Blecha F: CD14 and other
recognition molecules for lipopolysaccharide: A review.
Immunopharmacology. 29:187–205. 1995. View Article : Google Scholar : PubMed/NCBI
|