Introduction
Essential oil is a mixture of highly complex,
naturally occurring, volatile aroma compounds synthesized in
medicinal and aromatic plants as secondary metabolites (1). Essential oils can be extracted from
leaves, flowers, and stems by solvent extraction,
hydrodistillation, or steam distillation (2). In recent years, there has been
increasing interest worldwide in the use of alternative/herbal
medicines for the prevention and treatment of various illnesses
(3). Essential oils are widely used
in pharmaceutical, cosmetic, sanitary, food industry, and
agriculture for their antibacterial, antiviral, antifungal,
antiparasitic, insecticidal, anticancer, neuroprotective,
psychophysiological, and anti-aging activities (1). However, much of the research into
essential oils is still in its early state, and a systematic and
rigorous approach to the study of biological activities of
potential phytotherapeutics has only been initiated in recent
decades. This is particularly true regarding the cytotoxic effects
of phytocomplexes (4). Because of
the increasing popularity of essential oils and the prevalence of
essential oil-based self-care practices targeting a wide variety of
ailments in the United States, healthcare professionals need to be
prepared to address concerns about the agents' safety and efficacy.
Thorough literature evaluation of a purported therapeutic requires
the ability to discern the quality of an oil, the safety of its
administration, and the validity of its declared use (5). However, there is considerable anecdotal
information about the biological activity of essential oils, and
much of that information has not been substantiated by scientific
or clinical evidence (6). A recent
study has shown that, in eukaryotic cells, essential oils can act
as prooxidants affecting inner cell membranes and organelles such
as mitochondria. Depending on the type and concentration, essential
oils can exhibit cytotoxic effects on living cells, but they are
usually non-genotoxic (7).
The cell cycle is divided into discrete phases: G1
(gap 1) is the interval or gap between mitosis (M phase) and DNA
synthesis (S phase). During G1, the cell is subject to stimulation
by extracellular mitogens and growth factors, and, in response to
these stimuli, the cell passes through G1 and proceeds with DNA
synthesis in the S phase (8). G2
(gap 2) is the interval between the completion of DNA synthesis and
mitosis, and the M phase is marked by the generation of bipolar
mitotic spindles, segregation of sister chromatids, and cell
division (8). The G1/S and G2/M
cell-cycle checkpoints maintain genomic stability in eukaryotes in
response to genotoxic stress (9).
The balance between cyclin-dependent protein kinase (CDK)
activation and inactivation determines whether cells proceed
through G1 into the S phase, as well as from G2 to M, through
regulatory mechanisms that are conserved in more complex eukaryotes
(10).
The formation of cyclin D/CDK complexes results in
phosphorylation and activation of the CDKs (11). The critical role for CDK4 and CDK6
during cell-cycle progression is to phosphorylate the tumor
suppressor pRb, which consequently permits the transition from G1
to the S phase (12). As the cell
progresses through late G1, there is increased expression of cyclin
E, and cyclin E/CDK2 complexes are required for the transition from
G1 to the S phase. Increased expression of cyclin A occurs at the
G1/S transition and persists through the S phase. With the binding
of cyclin A to CDK2, DNA synthesis proceeds. In the latter part of
the S phase, cyclin A associates with CDK1. A checkpoint in G2
responds to the presence of DNA damage or incomplete DNA synthesis;
as a consequence, progression into mitosis is delayed allowing DNA
repair or the cycle is aborted. Increased levels of cyclins A and B
complexed with CDK1 propel the cell through mitosis (n.b.,
historically, CDK1 was referred to as cdc2) (11). Cyclin A/CDK2 complexes are active
from the onset of the S phase through the G2 phase and into the
early M phase, at which point the complex is inactivated by cyclin
A degradation (12). The active
cyclin B1/CDK1 complex first appears at the centrosome, suggesting
that the centrosome may facilitate the activation of mitotic
regulators that are required for the commitment of cells to proceed
to mitosis (13).
Interleukin 1-converting enzyme-like proteases
(caspases) are crucial components of cell-death pathways (14). Caspase-3 normally exists in the
cytosolic fraction of cells as an inactive precursor that is
activated proteolytically when cells are signaled to undergo
apoptosis (15). The activation of
apoptosis occurs in two stages: First, cytosolic caspase-3 is
cleaved and activated in a reaction that is triggered by cytochrome
c released from mitochondria; and second, the activated caspase-3
interacts with other cytosolic proteins to generate DNA
fragmentation when it is added to isolated nuclei (16,17).
Thus, caspase-3 is essential for certain processes associated with
the dismantling of the cell and the formation of apoptotic bodies,
but it may also function before, or at the stage when, the
commitment to loss of cell viability is made (18).
In this study, to determine the appropriateness of
using several plant essential oils clinically, we identified the
IC50 values of essential oils extracted from Pinus
densiflora for. multicaulis Uyeki, Trifolium repens,
Ligularia fischeri, Abies nephrolepis, Illicium anisatum,
Zanthoxylum coreanum, Abies koreana, Lindera obtusiloba,
Chamaecyparis obtuse, Pinus densiflora, Magnolia kobus, Picea
koraiensis, Picea abies, Abies holophylla, and Platycladus
orientalis in human lung (A549) and human skin (Detroit 551)
cells. In addition, the mechanism of cytotoxicity of each of the
essential oils was determined by examining changes in the gene and
protein expressions of four cell-cycle markers (cyclin A, cyclin B,
cyclin D, and cyclin E) and one cell-death marker (caspase-3).
Materials and methods
Essential oil extraction from
plants
The plants come from National Experimental Forest in
Pocheon city, Gyeonggi-do and Seogwipo city, JeJu-do, Republic of
Korea. The sampling was performed with the permission of operations
officer of experimental forest of National Institute of Forest
Science for the purpose of experiments. The
threatened/endangered/near threatened species such as Abies
koreana, Chamaecyparis obtuse, Abies holophylla, Platycladus
orientalis samples came from byproduct with the cutback. All
the threatened/endangered/near threatened species sample was
obtained non-lethal, sustainable, responsible collecting which is
following International Union for Conservation of Nature Species
Survival Commission recommendations with the government official
from National Institute of Forest Science of Korea. Taxonomical
identifications were established by the ecologist Dr. Jae-Min Chung
from Korean National Institute of Forest and the voucher specimens
was preserved in Korea National Arboretum.
Hydrodistillation was used for extracting essential
oils from plant samples. The essential oil from one kilogram of
freshly cut plant leaves is obtained by steam distillation using a
manufactured apparatus with a condenser at 105°C. Distillation
(EAMS 9501; Misung Scientific Co. Ltd.) was continued for 7 h at
100°C and the volatile compounds containing the water-soluble
fraction were allowed to settle for 20 min. The extracted essential
oils were added Na2SO4 (98.5%; SAMCHUN) to
subduct water from essential oil. The essential oil layer was
separated and finally purified using a microfilter (pore size, 0.45
µm) and the water vapor distillation method.
Cell proliferation assay
Human lung (A549) and human skin (Detroit 551) cells
were purchased from the Korean Cell Line Bank (KCLB). Cells were
seeded at a density of 0.5×103 cells per well in 96-well
plate and 100 µl of media used for cell culture which are DMEM
high-glucose media (Biowest) supplemented with
penicillin-streptomycin solution (Biowest), Plasmocin™ prophylactic
(Biowest), and fetal bovine serum. Cells were incubated in a
CO2 atmosphere at 37°C for 24 h prior to treating with
an essential oil. The plant essential oils were prepared in four
concentrations 10−8, 10−6, 10−4,
and 10−2% and distilled water as a vehicle (specimen
volume/media volume). After 24 h, the cells are treated with
diluted plant essential oil as indicated and incubated for 24 h.
Subsequently, the plant essential oil and the medium treating the
cells were removed and the cells washed with PBS solution
(Welgene). EZ-Cytox enhanced cell viability assay reagent
(DoGenBio) at the recommended concentration was then placed in each
well. One hour later, the absorbance value at 450 nm was measured
with an Epoch microplate spectrophotometer (BioTek). Cell viability
(%) was determined by comparing optical density (OD) values via the
formula ODsample/ODcontrol ×100 for each
concentration range. A cell survival curve was calculated from the
obtained values, and the oil's IC50 value was
established. After conducting the CCK assay, the same procedure was
repeated with plant essential oils that were diluted to
concentrations of 10−6, 10−5,
10−4, 10−3, 10−2. All experiment
was replicated four times. The IC50 values were
calculated with Graph Pad Prism (v.5.0; GraphPad Software) with
nonlinear regression (curve fit) in the 95% confidence
interval.
RNA extraction
Cells were seeded in 6-well plates at a density of
0.3×104 cells per well and incubated at 37°C in 5%
CO2 atmosphere. Plant essential oils in DMEM
high-glucose media (Biowest) were prepared in three concentrations:
IC50 from the CCK assay, 10 times more than the
IC50, and 10 times less than the IC50. When
cell confluency was 70%, the cells were treated with the
appropriate concentration of plant essential oil. Harvesting of
cells and RNA extraction were accomplished by using TRI reagent
(Invitrogen) at 24 h after plant essential oil treatment. The
purity and concentration of the extracted RNA were measured by
using an Epoch microplate spectrophotometer (BioTek).
Synthesis of cDNA
The extracted RNA was synthesized by reverse
transcription (RT). The concentration of RNA was titrated to 1
µg/µl and DEPC-treated water was added to adjust the total volume
to 10.5 µl. The mixture contained Moloney murine leukemia virus
reverse transcriptase (Invitrogen), Ribonuclease inhibitor
(Invitrogen), random primers (6-mers; TaKaRa), 100 mM DTT
(Invitrogen), dNTP, and 5× first-strand buffer. The reaction
mixture was incubated at 37°C for 60 min. After incubation, the
cells were incubated further at 95°C for 5 min for inactivation of
the enzyme. After cooling for 5 min on ice, the cDNA was stored at
−20°C prior to use.
Quantitative PCR
For qPCR, a master mix of the following reaction
components was prepared: cDNA sample (2 µl), 0.25 µl 50× ROX dye,
6.25 µl 2× prime Q-mastermix SYBR (GeNetBio), 10 pM forward and
reverse primer. QPCR was performed using a 7300 RT-PCR system
(Applied Biosystems), and the procedure sequence was as follows:
cDNA denatured at 95°C for 30 sec and annealed at 58°C for 30 sec,
elongation performed at 72°C for 30 sec, and the whole cycle is
repeated 40 times. The degree of gene expression was determined by
using RQ software (Applied Biosystems). The primer sequences used
are as follows: 5′-CATACTCCACAGCACCTGGTTA-3′ (forward) and
5′-CTGTTGCCACCTTTCGGTTA-3′ (reverse) for caspase-3;
5′-TACTTTCTGCATCAGCAGCC-3′ (forward) and 5′-TGATTCAGGCCAGCTTTGTC-3′
(reverse) for cyclin A; 5′-CTTTGCACTTCCTTCGGAGA-3′ (forward) and
5′-GTAGAGTTGGTGTCCATTCACC-3′ (reverse) for cyclin B;
5′-CCTCGGTGTCCTACTTCA-3′ (forward) and 5′-CTCCTCGCACTTCTGTTC-3′
(reverse) for cyclin D; and 5′-GGTTTCAGGGRATCAGTGGT-3′ (forward)
and 5′-TTTCTTTGCTCGGGGCTTTG-3′ (reverse) for cyclin E. The primer
sequence for Rn18s, 5′-CTCAACACGGGAAACCTCAC-3′ (forward) and
5′-CGCTCCACCAACTAAGAACG-3′ (reverse), was used as the primer for
the control group. All primers were obtained from Macrogen.
Western blot analysis
Proteins were extracted using Pro-prep (InTron Inc.)
according to the manufacturer's instructions. Proteins (30 µg per
lane) were separated on a 10% SDS-PAGE gel and transferred to a
polyvinylidene fluoride transfer membrane (Perkin Elmer Co.) in a
TransBlot Cell (TE-22; Hoefer Co.) according to the manufacturer's
instructions. The resulting blot was blocked with TBS-T containing
5% skimmed milk for 60 min, then incubated with primary antibody:
Caspase-3 (mouse monoclonal, 1:500, cat. no. 9665), Cyclin A2
(mouse monoclonal, 1:500, cat. no. 4656), Cyclin B1 (rabbit
polyclonal, 1:500, cat. no. 4138), Cyclin D1 (rabbit monoclonal,
1:500, cat. no. 2978), Cyclin E1 (mouse monoclonal, 1:500, cat. no.
4129), or β-actin (rabbit monoclonal, 1:1,000, cat. no. 4970; all
purchased from Cell Signaling Technology). After washing in buffer,
the membranes were incubated with the appropriate horseradish
peroxidase-conjugated secondary antibodies (anti-goat, 1:2,000 or
anti-mouse, 1:4,000; Santa Cruz) for 1 h at room temperature. Next,
the blots were developed by incubation in ECL chemiluminescence
reagent (Santa Cruz) and subsequently exposed to the GeneGnome
Bioimaging System (Syngene) for development, after which images
were captured by using the GENESys software (Syngene). Signal
specificity was confirmed by blotting in the absence of primary
antibody, and bands were normalized to β-actin immunoreactive bands
visualized on the same membrane after stripping. The density of
each band was measured using the NIH ImageJ software.
Statistical analysis
All data were analyzed by applying nonparametric
one-way analysis of variance followed by Turkey's test for multiple
comparisons. All experiments consisted of three separate trials.
Statistical analysis was performed by using Graph Pad Prism
(GraphPad Software).
Results
Two-step identification of
IC50 value of each essential oil
CCK-8 assays were used to determine IC50
values of 15 essential oils. To determine the test concentration,
the cells were treated initially with four concentrations, followed
by testing of further concentrations in CCK-8 assays. The
IC50 values of the 15 essential oils are presented in
Figs. 1 and 2. For the initial testing of concentrations
of the essential oils (i.e., 10−8,
10−6, 10−4, 10−2, 1%), the
IC50 values (%) were determined to be 0.2001 (Pinus
densiflora for. multicaulis Uyeki), 0.0521 (Trifolium
repens), 0.6011 (Ligularia fischeri), 0.1127 (Abies
nephrolepis), 0.3551 (Illicium anisatum), 0.0391
(Zanthoxylum coreanum), 0.0946 (Abies koreana),
0.8551 (Lindera obtusiloba), 0.0339 (Chamaecyparis
obtusa), 0.0065 (Pinus densiflora), 0.553 (Magnolia
kobus), 0.0526 (Picea koraiensis), 0.0364 (Picea
abies), 0.0473 (Abies holophylla), and 0.0987
(Platycladus orientalis) in A549 cells and as 0.3321
(Pinus densiflora for. multicaulis Uyeki), 0.2869
(Trifolium repens), 0.4013 (Ligularia fischeri),
1.037 (Abies nephrolepis), 0.3440 (Illicium
anisatum), 0.4661 (Zanthoxylum coreanum), 1.190
(Abies koreana), 0.8859 (Lindera obtusiloba), 0.1246
(Chamaecyparis obtusa), 0.2863 (Pinus densiflora),
0.0748 (Magnolia kobus), 0.0460 (Picea koraiensis),
0.0274 (Picea abies), 0.1031 (Abies holophylla),
0.0165 (Platycladus orientalis) in Detroit 551 cells.
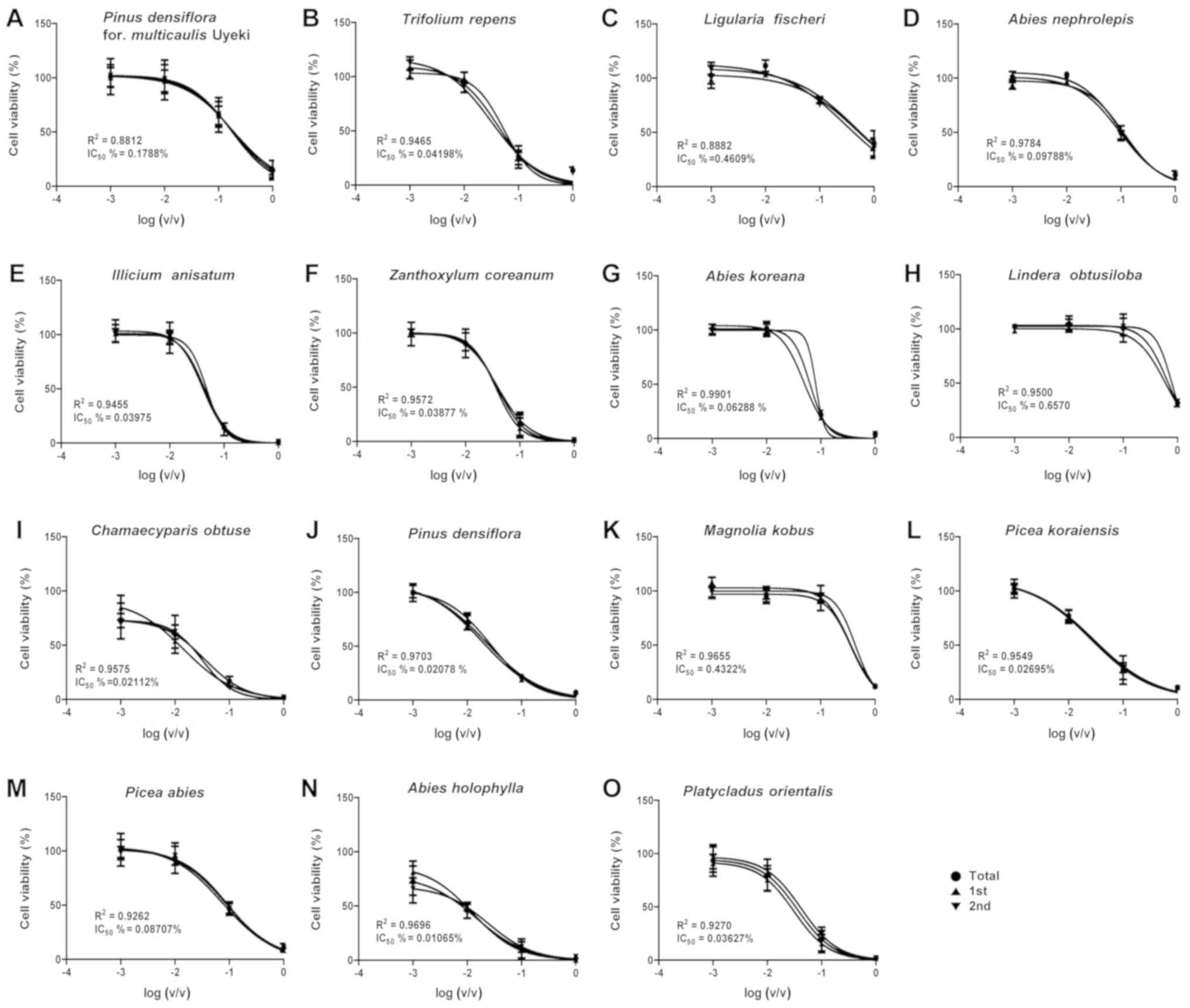 | Figure 1.A549 cell survival curves for the
determination of essential oil IC50 values. After cells
were treated with an essential oil, the OD was measured by
performing a cell counting kit-8 assay. Cell viability was
determined using the following formula:
ODsample/ODcontrol ×100 (%), after which the
IC50 value was determined through the obtained survival
curve. (A) Pinus densiflora for. multicaulis Uyeki,
(B) Trifolium repens, (C) Ligularia fischeri, (D)
Abies nephrolepis, (E) Illicium anisatum, (F)
Zanthoxylum coreanum, (G) Abies koreana, (H)
Lindera obtusiloba, (I) Chamaecyparis obtuse, (J)
Pinus densiflora, (K) Magnolia kobus, (L) Picea
koraiensis, (M) Picea abies, (N) Abies
holophylla, (O) Platycladus orientalis. OD, optical
density. |
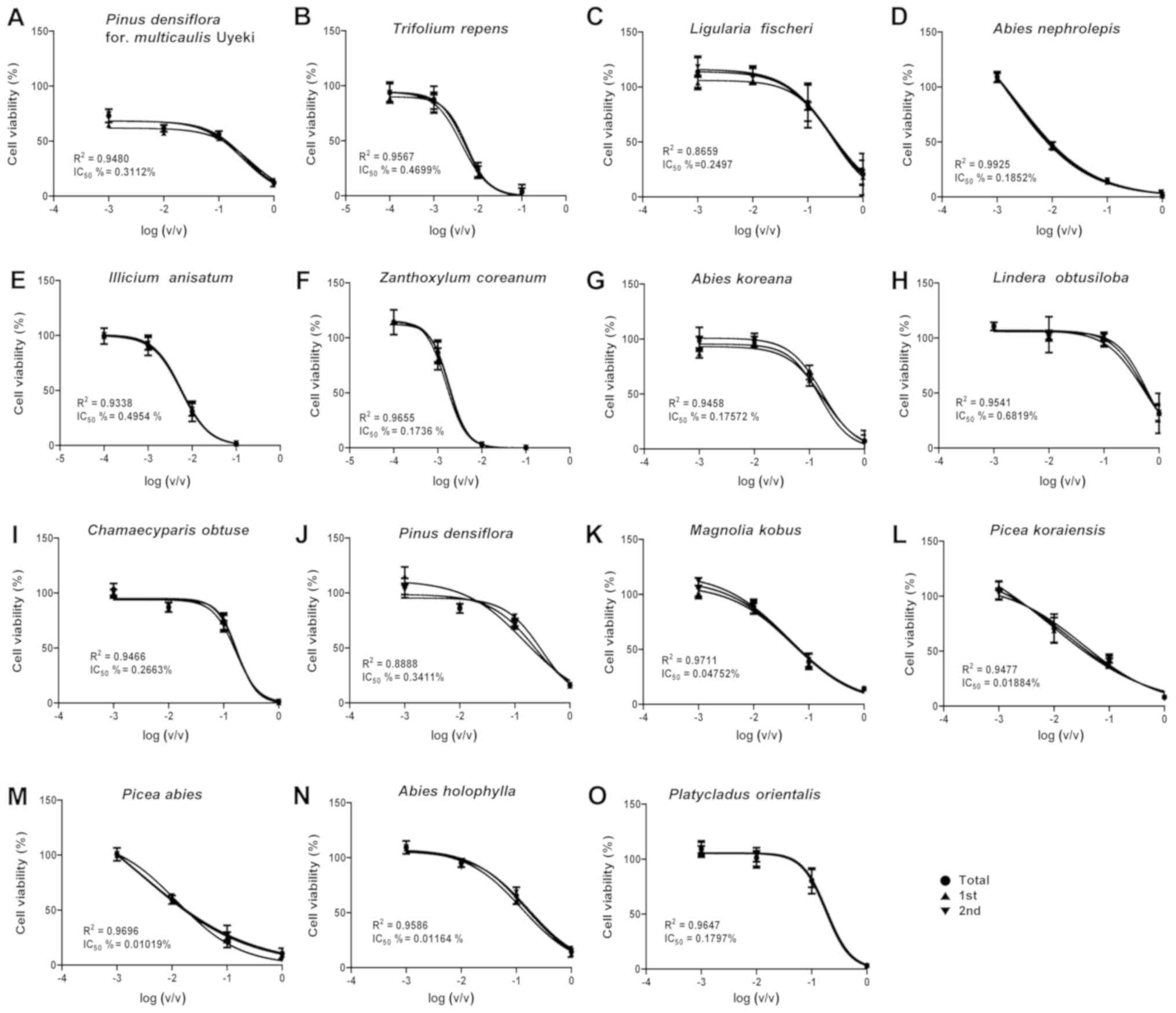 | Figure 2.Detroit 551 cell survival curves for
the determination of essential oil IC50 values. After
the cells were treated with an essential oil, Cell viability was
determined using the following formula:
ODsample/ODcontrol ×100 (%), after which the
IC50 value was determined through the obtained survival
curve. (A) Pinus densiflora for. multicaulis Uyeki,
(B) Trifolium repens, (C) Ligularia fischeri, (D)
Abies nephrolepis, (E) Illicium anisatum, (F)
Zanthoxylum coreanum, (G) Abies koreana, (H)
Lindera obtusiloba, (I) Chamaecyparis obtuse, (J)
Pinus densiflora, (K) Magnolia kobus, (L) Picea
koraiensis, (M) Picea abies, (N) Abies
holophylla, (O) Platycladus orientalis. OD, optical
density. |
For the testing of further subdivisions of the
concentrations of the essential oils (i.e., 10−5,
10−4, 10−3, 10−2, 1%), the
IC50 values (%) were determined to be 0.1788 (Pinus
densiflora for. multicaulis Uyeki), 0.0420 (Trifolium
repens), 0.4609 (Ligularia fischeri), 0.0979 (Abies
nephrolepis), 0.0398 (Illicium anisatum), 0.0388
(Zanthoxylum coreanum), 0.0629 (Abies koreana),
0.6570 (Lindera obtusiloba), 0.0211 (Chamaecyparis
obtusa), 0.0207 (Pinus densiflora), 0.4322 (Magnolia
kobus), 0.0270 (Picea koraiensis), 0.0871 (Picea
abies), 0.0107 (Abies holophylla), and 0.0363
(Platycladus orientalis) in A549 cells (Table I) and as 0.3112 (Pinus
densiflora for. multicaulis Uyeki), 0.4699 (Trifolium
repens), 0.2497 (Ligularia fischeri), 0.1852 (Abies
nephrolepis), 0.4954 (Illicium anisatum), 0.1736
(Zanthoxylum coreanum), 0.1757 (Abies koreana),
0.6819 (Lindera obtusiloba), 0.2663 (Chamaecyparis
obtusa), 0.3411 (Pinus densiflora), 0.0475 (Magnolia
kobus), 0.0188 (Picea koraiensis), 0.0102 (Picea
abies), 0.1164 (Abies holophylla), 0.1797
(Platycladus orientalis) in Detroit 551 cells (Table II).
 | Table I.IC50 values of essential
oils in A549 cell line. |
Table I.
IC50 values of essential
oils in A549 cell line.
| Essential oils | IC50 (%,
v/v) |
|---|
| Pinus
densiflora for. multicaulis Uyeki | 0.1788 |
| Trifolium
repens | 0.04198 |
| Ligularia
fischeri | 0.4609 |
| Abies
nephrolepis | 0.09788 |
| Illicium
anisatum | 0.03975 |
| Zanthoxylum
coreanum | 0.03877 |
| Abies
koreana | 0.06288 |
| Lindera
obtusiloba | 0.6570 |
| Chamaecyparis
obtusa | 0.02112 |
| Pinus
densiflora | 0.02078 |
| Magnolia
kobus | 0.4322 |
| Picea
koraiensis | 0.02695 |
| Picea
abies | 0.08707 |
| Abies
holophylla | 0.01065 |
| Platycladus
orientalis | 0.03627 |
| Table II.IC50 values of essential
oils in Detroit 551 cells. |
Table II.
IC50 values of essential
oils in Detroit 551 cells.
| Essential oils | IC50 (%,
v/v) |
|---|
| Pinus
densiflora for. multicaulis Uyeki | 0.3112 |
| Trifolium
repens | 0.4699 |
| Ligularia
fischeri | 0.2497 |
| Abies
nephrolepis | 0.1852 |
| Illicium
anisatum | 0.4954 |
| Zanthoxylum
coreanum | 0.1736 |
| Abies
koreana | 0.17572 |
| Lindera
obtusiloba | 0.6819 |
| Chamaecyparis
obtusa | 0.2663 |
| Pinus
densiflora | 0.3411 |
| Magnolia
kobus | 0.04752 |
| Picea
koraiensis | 0.01884 |
| Picea
abies | 0.01019 |
| Abies
holophylla | 0.1164 |
| Platycladus
orientalis | 0.1797 |
Expression of caspase-3 gene affected
by plant essential oils
Expression levels of the caspase-3 gene, an
apoptosis marker gene, were determined by performing reverse
transcription-quantitative PCR (RT-qPCR) in essential oil-treated
cell lines (Fig. 3). In the A549
human lung cell line, the expression level of caspase-3 was
decreased by high concentrations of all of the tested essential
oils. However, in the Detroit 551 human skin cell line, the
expression level of caspase-3 was increased only in high
concentrations of Zanthoxylum coreanum and Ligularia
fischeri, while it was decreased in high concentrations of
Lindera obtusiloba, Abies koreana, and Abies
nephrolepis.
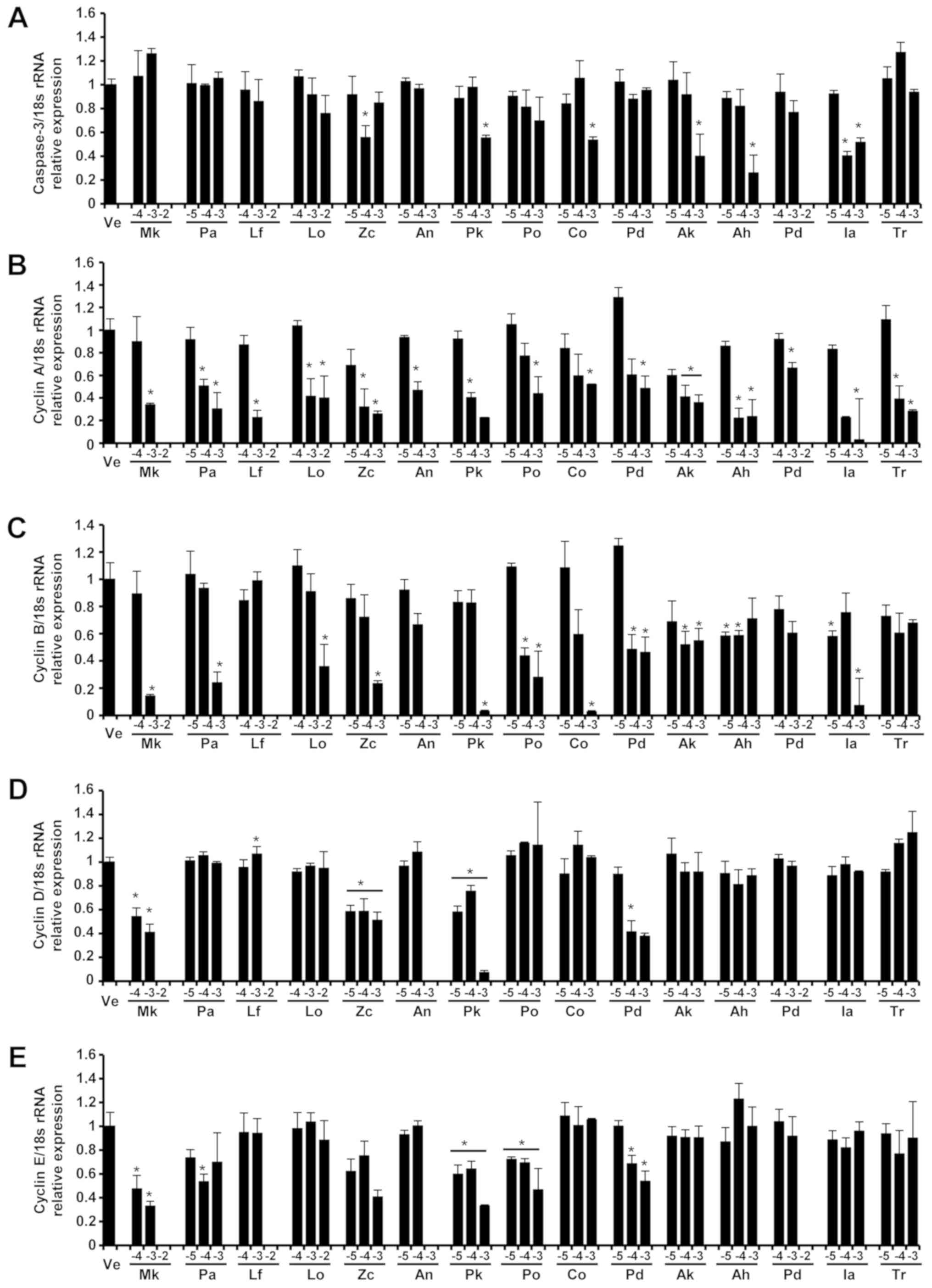 | Figure 3.Expression of (A) caspase-3, (B)
cyclin A, (C) cyclin B, (D) cyclin D and (E) cyclin E genes in A549
cells. The levels of caspase-3, cyclin A, cyclin B, cyclin D and
cyclin E mRNA in the A549 cells of each group were determined via
reverse transcription-quantitative PCR after treatment with three
concentrations of essential oil. The control group was treated with
vehicle (distilled water). Concentrations of −5, −4, −3 and −2
represented 10−5, 10−4, 10−3 and
10−2, respectively. Data are presented as the mean ±
standard deviation. *P<0.05 vs. control. |
The expression levels of caspase-3 protein, an
apoptosis marker, in the A549 and Detroit 551 cells were determined
by analyzing western blots (Fig. 4).
In the Detroit 551 cells, the expression level of caspase-3 protein
was increased at concentrations near the IC50
values.
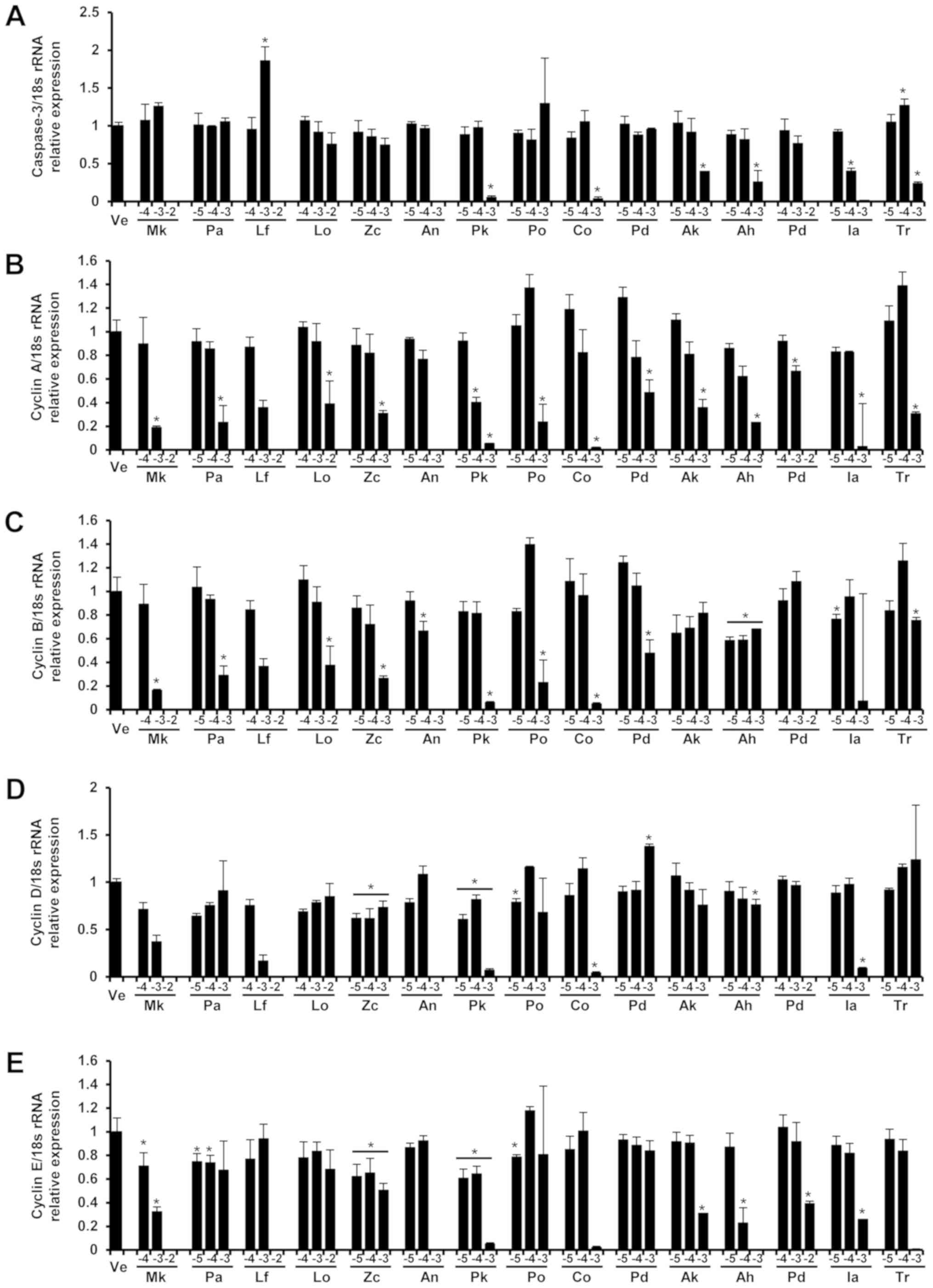 | Figure 4.Expression levels of (A) caspase-3,
(B) cyclin A, (C) cyclin B, (D) cyclin D and (E) cyclin E genes in
Detroit 551 cells. The levels of caspase-3, cyclin A, cyclin B,
cyclin D and cyclin E mRNA in the Detroit 551 cells of each group
were determined by via reverse transcription-quantitative PCR after
treatment with three concentrations of essential oil. The control
group was treated with vehicle (distilled water). Concentrations of
−5, −4, −3 and −2 represented 10−5, 10−4,
10−3 and 10−2, respectively. Data are
presented as the mean ± standard deviation. *P<0.05 vs.
control. |
Expressions of cyclin A, cyclin B,
cyclin D, and cyclin E affected by plant essential oils
The gene expression levels of cyclin A, cyclin B,
cyclin D, and cyclin E in the A549 and Detroit 551 cell lines after
treatment with different concentrations of the essential oils
tested in this study are shown in Figs.
3 and 4.
For all the tested essential oils, the gene
expression of cyclin A, a cell-cycle regulation marker, showed
concentration-dependent decreases in A549 cells. The expression of
cyclin B decreased after treatment with all oils except
Ligularia fischeri in which the expression level of cyclin B
increased in A549 cells. The cyclin D expression in A549 cells
showed decreases when they were treated with Zanthoxylum
coreanum and Abies koreana but showed increases with
Ligularia fischeri and Abies nephrolepis treatment.
Treatment with the essential oil of Lindera obtusiloba
produced no tendency for change in cyclin D expression in A549
cells. Expressions of cyclin E in A549 cells slightly decreased in
Ligularia fischeri and Abies koreana. Similarly, the
expressions of cyclin E in A549 cells showed no tendency for change
after treatment with essential oils of Lindera obtusiloba,
Zanthoxylum coreanum, and Abies nephrolepis.
In Detroit 551 cells, the gene expression of cyclin
A decreased in all samples except Ligularia fischeri in
which the expression of cyclin A increased. The expression of
cyclin B in Detroit 551 cells showed a decrease with Lindera
obtusiloba treatment but showed increases after Ligularia
fischeri, Abies koreana, and Abies nephrolepis
treatment. There was no tendency for change in cyclin B expression
in Detroit 551 cells after treatment with Zanthoxylum
coreanum. The expression levels of cyclin D in Detroit 551
cells decreased after Lindera obtusiloba and Abies
koreana treatment but increased following Zanthoxylum
coreanum, Ligularia fischeri, and Abies nephrolepis. The
expression of cyclin E in Detroit 551 cells decreased with
Ligularia fischeri and Abies nephrolepis treatment.
The essential oils of Lindera obtusiloba, Zanthoxylum
coreanum, and Ligularia fischeri did not produce a
tendency for change in the expression level of cyclin E in Detroit
551 cell lines.
The protein expression levels of cyclin A, cyclin B,
cyclin D, and cyclin E in the A549 and Detroit 551 cells at
different concentrations of the essential oils are presented in
Figs. 5 and 6. In the A549 cells, the protein expression
levels of cyclin A, cyclin B, cyclin D, and cyclin E decreased in
all essential oils except Ligularia fischeri in which the
expression levels of cyclins A and B proteins increased. In the
Detroit 551 cells, the protein expression levels of cyclin A,
cyclin B, cyclin D, and cyclin E decreased after treatment with all
essential oils except Ligularia fischeri for which the
expression level of cyclin B increased and for Abies
nephrolepis in which cyclin E protein expression increased.
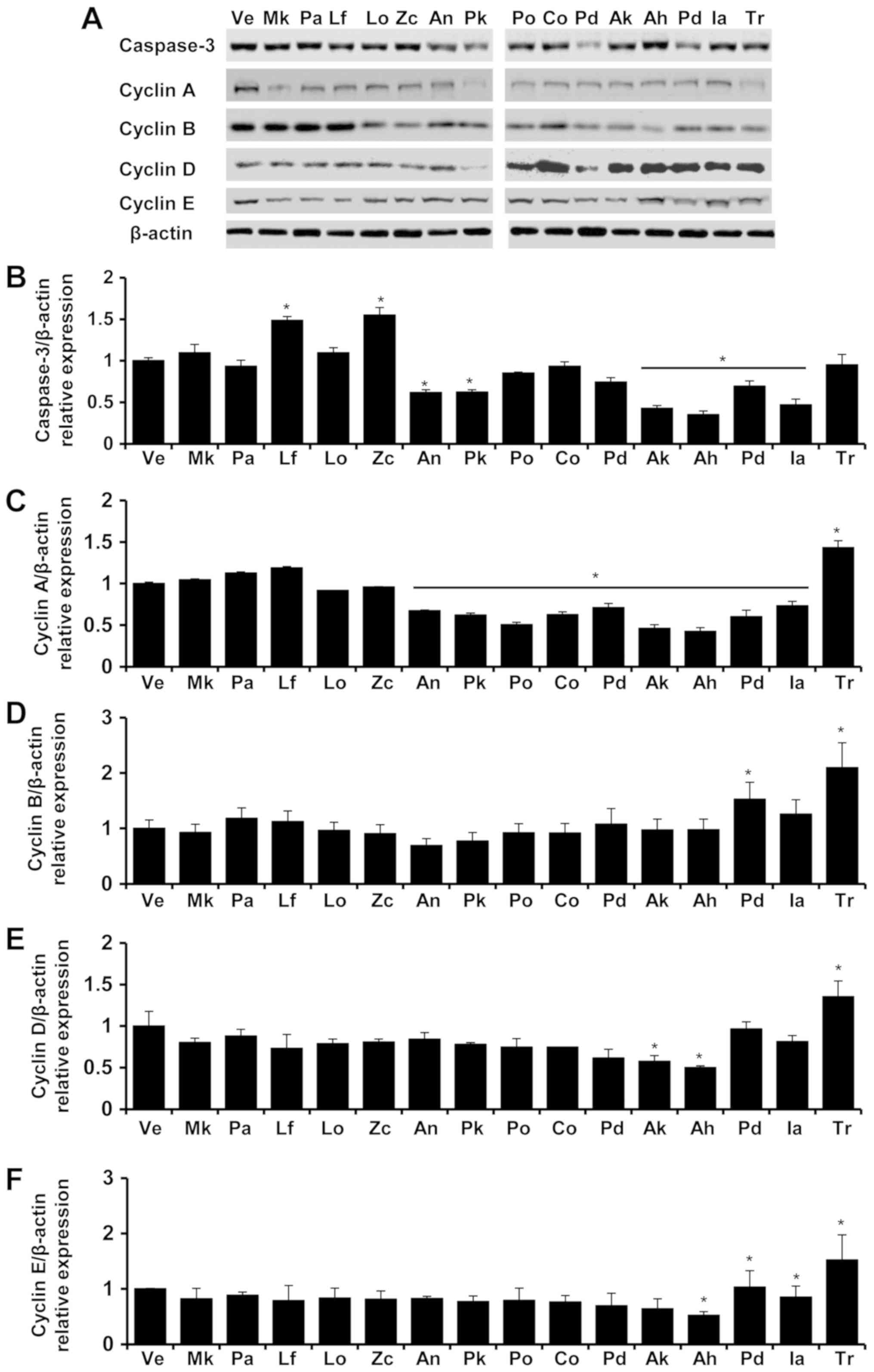 | Figure 5.Expression of caspase-3, cyclin A,
cyclin B, cyclin D and cyclin E proteins in A549 cells. Levels of
caspase-3, cyclin A, cyclin B, cyclin D and cyclin E protein in the
A549 cells of each group were determined via Western blot analysis
after treatment with tessential oils (A). The protein expression
level of Caspase-3 (B), Cyclin A (C), Cyclin B (D), Cyclin D (E),
Cyclin E (F)/β-actin in vehicle (Ve), Magnolia kobus (Mk), Picea
abies (Pa), Ligularia fischeri (Lf), Lindera obtusiloba (Lo),
Zanthoxylum coreanum (Zc), Abies nephrolepis (A.n), Picea
koraiensis (Pk), Platycladus orientalis (Po), Chamaecyparis obtuse
(Co), Pinus densiflora (Pd), Abies koreana (Ak), Abies holophylla
(Ah), Pinus densiflora for. multicaulis Uyeki (Pd), Illicium
anisatum (Ia), Trifolium repens (Tr) were presented. The control
group was treated with vehicle (distilled water). Concentrations of
−5, −4, −3 and −2 represented 10-5, 10-4, 10-3 and 10-2,
respectively. Data are presented as the mean ± standard deviation.
*P<0.05 vs. control.. |
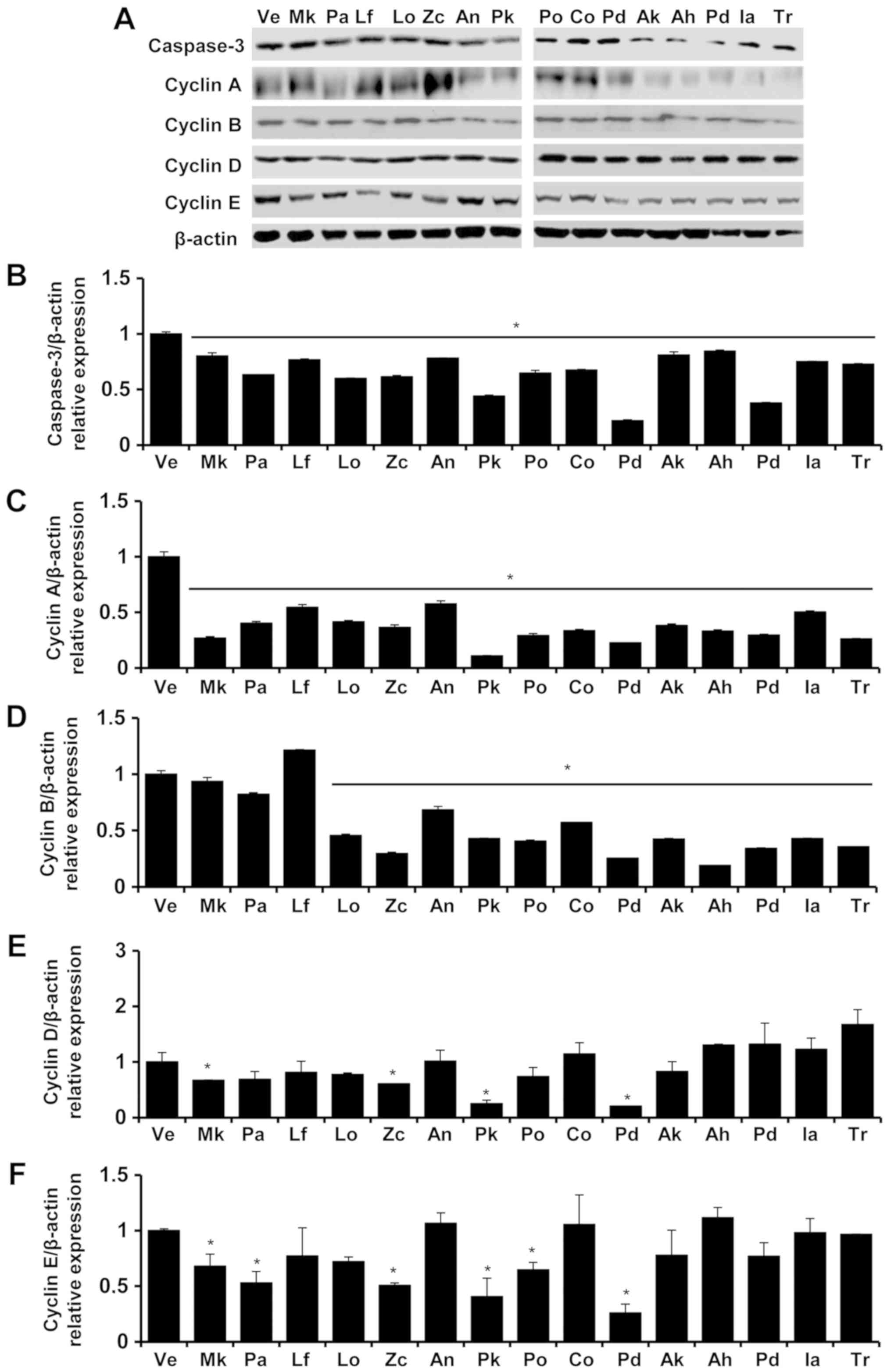 | Figure 6.Expression of caspase-3, cyclin A,
cyclin B, cyclin D and cyclin E proteins in Detroit 551 cells.
Levels of caspase-3, cyclin A, cyclin B, cyclin D and cyclin E
protein in the Detroit 551 cells of each group were determined via
Western blot analysis after treatment with essential oils (A). The
protein expression level of Caspase-3 (B), Cyclin A (C), Cyclin B
(D), Cyclin D (E), Cyclin E (F)/β-actin in vehicle (Ve), Magnolia
kobus (Mk), Picea abies (Pa), Ligularia fischeri (Lf), Lindera
obtusiloba (Lo), Zanthoxylum coreanum (Zc), Abies nephrolepis
(A.n), Picea koraiensis (Pk), Platycladus orientalis (Po),
Chamaecyparis obtuse (Co), Pinus densiflora (Pd), Abies koreana
(Ak), Abies holophylla (Ah), Pinus densiflora for. multicaulis
Uyeki (Pd), Illicium anisatum (Ia), Trifolium repens (Tr) were
presented. The control group was treated with vehicle (distilled
water). Concentrations of −5, −4, −3 and −2 represented 10-5, 10-4,
10-3 and 10-2, respectively. Data are presented as the mean ±
standard deviation. *P<0.05 vs. control. |
As indicated by the increased expression levels of
caspase-3, apoptosis is the cell-death mechanism associated with
the essential oils of Zanthoxylum coreanum and Ligularia
fischeri in the Detroit 551 cell line. As indicated by the
decreased expression levels of cyclin A, cyclin B, cyclin D, and
cyclin E, the cell-death mechanism associated with the rest of the
tested essential oils is cell-cycle arrest.
Discussion
A plant essential oil is a volatile natural compound
obtained as a secondary product of the plant and is characterized
by a strong smell (12). The
components of a plant essential oil depend on the species of plant
undergoing essential oil extraction and from which part of the
plant the oil is extracted (17).
Oils may be extracted from different parts of the plant, such as
leaves, fruit peels, seeds, bark, wood, and flowers, and are
usually obtained via hydro- or steam distillation processes.
Essential oils are complex mixtures of volatile compounds produced
by living organisms. and are isolated by physical means
(compression and/or distillation) from part of whole plants of
known taxonomic origin (19).
Biological properties of various essential oils have
been described in several studies. In India, Trifolium
repens is fed to livestock and considered a folk medicine
against intestinal helminthic worms (4), Illicium anisatum cause severe
inflammation of the kidneys, urinary tract, and digestive organs
(5). A Lindera obtusiloba
water extract inhibited mast-cell-derived allergic inflammation
in vitro and in vivo, suggesting it may have possible
application in allergic diseases such as allergic rhinitis, asthma
and atopic dermatitis (20).
Chamaecyparis obtuse and Pinus densiflora exhibited
anti-inflammatory effects in an in vivo study (12). Picea abies has been used as a
traditional Austrian medicine internally (as syrup or tea) and
externally (for bathing, for inhalation, and as an ointment, resin
application, or tea) for treatment of disorders of the respiratory
tract, skin, locomotory system, and gastrointestinal tract, as well
as for infection treatment (21).
Although there is extensive usage of essential oils and some of
their effects have been described, there is a lack of studies on
the cytotoxic effects of essential oils and their mechanisms of
action.
Determining the IC50 value is regarded as
a standard when assessing the toxicity of chemical compounds. In
this study, the IC50 value for each essential oil was
calculated by using a CCK-8 assay. At concentrations close to the
oil's IC50 value, cell viability (as %) was shown to be
markedly decreased in both human lung cells (A549) and human skin
cells (Detroit 551). Cytostatic effects, identified by determining
IC50 values, of essential oils from Pinus
densiflora for. multicaulis Uyeki, Trifolium repens,
Ligularia fischeri, Abies nephrolepis, Illicium anisatum,
Zanthoxylum coreanum, Abies koreana, Lindera obtusiloba,
Chamaecyparis obtuse, Pinus densiflora, Magnolia kobus, Picea
koraiensis, Picea abies, Abies holophylla, and Platycladus
orientalis were identified in A549 human lung cells and Detroit
551 human skin cells. Furthermore, at the determined cytotoxicity
levels, we examined the expression levels of marker genes and
proteins (caspase-3, cyclin A, cyclin B, cyclin D, cyclin E) to
elucidate each oil's cytotoxic mechanism. Various mechanisms may be
account for the reported cytotoxic effects of some essential oils
or their constituents, including induction of cell death by
apoptosis and/or necrosis, cell-cycle arrest, and loss of function
of key organelles (4). Caspase-3 is
required for some typical hallmarks of apoptosis and is
indispensable for apoptotic chromatin condensation and DNA
fragmentation in all cell types examined and is considered a
cell-death marker gene. Thus, caspase-3 is essential for certain
processes associated with the dismantling of the cell and the
formation of apoptotic bodies, but it may also function before, or
at, the stage when commitment to loss of cell viability is made
(22). In Detroit 551 cells,
expression levels of caspase-3 increased under high concentration
treatments of Zanthoxylum coreanum, and Ligularia
fischeri indicating that cell death can occur via treatment by
essential oils from Zanthoxylum coreanum and Ligularia
fischeri. In contrast, under high concentration treatment of
Lindera obtusiloba, Abies koreana, and Abies
nephrolepis essential oils, the expression levels of caspase-3
decreased. In A549 cells, expression levels of caspase-3 decreased
under high concentration treatment by all oil samples. This
suggests that the cytotoxicity of these oils in lung cell lines is
due to inhibition of cell proliferation rather than to apoptosis.
This suggests that there can be other mechanisms involved in
essential oil-related cell death. Cell-cycle progression is
controlled by the activities of complexes that are comprised of
cyclins (A, B, D, E) bound to CDKs (20). The expression levels of cyclin A,
cyclin B, cyclin D, cyclin E, which are cell-cycle regulation
checkpoint markers for different cell-cycle stages, can be used to
determine the timing of cell-cycle arrest. CyclinA is associated
with the G1/S transition (the G1 checkpoint), cyclin B with the S
phase progression (the intra-S phase checkpoint), cyclin D with the
G2/M boundary (the G2/M checkpoint), and cyclin E with the spindle
checkpoint at the transition from metaphase to anaphase during
mitosis. Checkpoint activation results either in cell death or
improved cell survival, and deregulation of these cyclin-based
critical signaling pathways may lead to the disruption of essential
cellular functions (20). Therefore,
decreases in the levels of specific cyclins can be used to indicate
the specific phase of the cell cycle that is being inhibited by the
oil being tested. There is little information on the medical and
daily uses of some of the plant essential oils that were examined
in this study. In our previous study, the essential oils of
Chamaecyparis obtuse and Pinus densiflora have
anti-inflammatory effects that differ for skin and lung cells.
Illicium anisatum oil has toxic to proinflammatory effects
dependent on its concentration (23). Therefore, determination of the
appropriate concentration of a plant essential oil use is important
when it is being used for clinical purposes.
In this study, we evaluated the cytotoxicity of
several essential oils by performing CCK-8 assays and assessed the
mechanism of their cytotoxic effect by conducting RT-qPCR of the
cell-cycle-related genes, cyclin E, cyclin D, cyclin A, cyclin B,
and caspase-3. Each essential oil had a distinctive IC50
value and they produced different expression patterns for cyclin E,
cyclin D, cyclin A, cyclin B, caspase-3. Moreover, in the same
essential oil, the IC50 values and expression patterns
of the cell proliferation markers and the cell-death marker
differed depending on the cell type being tested. Essential oils
and their volatile constituents are used widely to prevent and
treat human disease (22); however,
medical use of a plant essential oil has the potential to cause
adverse effects due to using an inappropriate dose of the oil. The
lack of sufficiently convincing evidence regarding the
effectiveness of a plant essential oil combined with its potential
to cause adverse effects questions the usefulness of using this
treatment modality under any conditions (24). Regardless, with insufficient
evidence, plant essential oils can be effective in medical and
daily use. For example, conditions such as atopic dermatitis or
asthma can be alleviated with plant essential oils (12). In this study, to determine their
cytotoxicity, we calculated the IC50 values for 15 plant
essential oils from CCK-8 assay results and identified their
cytotoxic mechanisms by examining the gene and protein expression
levels of five marker genes (caspase-3, cyclin A, cyclin B, cyclin
D, and cyclin E). The results support the conclusion that cell
lines of human origin can be used to determine the safe
concentration of a plant essential oil for skin and lung cell
treatments (20). These results
should provide users of these essential oils with guidelines for
their clinically appropriate and safe use.
Acknowledgements
The authors give thanks to the ecologist Dr. Jae-Min
Chung from The Korean National Institute for Taxonomical
identifications of samples
Funding
The present study was supported by the National
Institute of Forest Science funded by the Korean government (grant
no. FP0700-2015-02).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
C. A and E-B. J conceived the study, C. A and J-H. L
performed and analyzed the experiments, YMY interpreted the data
and finalized as well as proofread the manuscript. M-J. P, J-W. K
and J. Y prepared the essential oil and analyzed the experiments.
E-B. J supervised the study.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kumar Y, Prakash O, Tripathi H, Tandon S,
Gupta MM, Rahman LU, Lal RK, Semwal M, Darokar MP and Khan F:
AromaDb: A database of medicinal and aromatic Plant's aroma
molecules with phytochemistry and therapeutic potentials. Front
Plant Sci. 9:10812018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Diaz-Maroto MC, Diaz-Maroto Hidalgo IJ,
Sanchez-Palomo E and Pérez-Coello MS: Volatile components and key
odorants of fennel (Foeniculum vulgare Mill.) and thyme (Thymus
vulgaris L.) oil extracts obtained by simultaneous
distillation-extraction and supercritical fluid extraction. J Agric
Food Chem. 53:5385–5389. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huie CW: A review of modern
sample-preparation techniques for the extraction and analysis of
medicinal plants. Anal Bioanal Chem. 373:23–30. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Russo R, Corasaniti MT, Bagetta G and
Morrone LA: Exploitation of cytotoxicity of some essential oils for
translation in cancer therapy. Evid Based Complement Alternat Med.
2015:3978212015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Manion CR and Widder RM: Essentials of
essential oils. Am J Health Syst Pharm. 74:e153–e162. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cavanagh HM and Wilkinson JM: Biological
activities of lavender essential oil. Phytother Res. 16:301–308.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bakkali F, Averbeck S, Averbeck D and
Idaomar M: Biological effects of essential oils-a review. Food Chem
Toxicol. 46:446–475. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Israels ED and Israels LG: The cell cycle.
Stem Cells. 19:88–91. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang XW, Zhan Q, Coursen JD, Khan MA,
Kontny HU, Yu L, Hollander MC, O'Connor PM, Fornace AJ Jr and
Harris CC: GADD45 induction of a G2/M cell cycle checkpoint. Proc
Natl Acad Sci USA. 96:3706–3711. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bertoli C, Skotheim JM and de Bruin RA:
Control of cell cycle transcription during G1 and S phases. Nat Rev
Mol Cell Biol. 14:518–528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vermeulen K, Van Bockstaele DR and
Berneman ZN: The cell cycle: A review of regulation, deregulation
and therapeutic targets in cancer. Cell Prolif. 36:131–149. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Donjerkovic D and Scott DW: Regulation of
the G1 phase of the mammalian cell cycle. Cell Res. 10:1–16. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kramer A, Mailand N, Lukas C, Syljuåsen
RG, Wilkinson CJ, Nigg EA, Bartek J and Lukas J:
Centrosome-associated Chk1 prevents premature activation of
cyclin-B-Cdk1 kinase. Nat Cell Biol. 6:884–891. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jänicke RU, Sprengart ML, Wati MR and
Porter AG: Caspase-3 is required for DNA fragmentation and
morphological changes associated with apoptosis. J Biol Chem.
273:9357–9360. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li P, Nijhawan D and Wang X: Mitochondrial
activation of apoptosis. Cell. 116 (2 Suppl):S57–S59. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo X, Budihardjo I, Zou H, Slaughter C
and Wang X: Bid, a Bcl2 interacting protein, mediates cytochrome c
release from mitochondria in response to activation of cell surface
death receptors. Cell. 94:481–490. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang J, Liu X, Bhalla K, Kim CN, Ibrado
AM, Cai J, Peng TI, Jones DP and Wang X: Prevention of apoptosis by
Bcl-2: Release of cytochrome c from mitochondria blocked. Science.
275:1129–1132. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gurib-Fakim A: Medicinal plants:
Traditions of yesterday and drugs of tomorrow. Mol Aspects Med.
27:1–93. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
El-Aouar Filho RA, Nicolas A, De Paula
Castro TL, Deplanche M, De Carvalho Azevedo VA, Goossens PL, Taieb
F, Lina G, Le Loir Y and Berkova N: Heterogeneous family of
cyclomodulins: Smart weapons that allow bacteria to hijack the
eukaryotic cell cycle and promote infections. Front Cell Infect
Microbiol. 7:2082017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barile FA and Cardona M: Acute
cytotoxicity testing with cultured human lung and dermal cells. In
Vitro Cell Dev Biol Anim. 34:631–635. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Edris AE: Pharmaceutical and therapeutic
potentials of essential oils and their individual volatile
constituents: A review. Phytother Res. 21:308–323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim JY, Kim SS, Oh TH, Baik JS, Song G,
Lee NH and Hyun CG: Chemical composition, antioxidant,
anti-elastase, and anti-inflammatory activities of Illicium
anisatum essential oil. Acta Pharm. 59:289–300. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Posadzki P, Alotaibi A and Ernst E:
Adverse effects of aromatherapy: A systematic review of case
reports and case series. Int J Risk Saf Med. 24:147–161. 2012.
View Article : Google Scholar : PubMed/NCBI
|














