Introduction
Proliferative vitreoretinopathy (PVR) is a serious
complication that is caused by rhegmatogenous retinal detachment
and is the leading cause of retinal detachment surgery failure
(1,2). It is physiologically characterised by
increased cell proliferation, migration and secretion of
extracellular matrix (ECM) proteins, which results in the formation
of fibrotic membranes in response to retinal detachment. Retinal
pigment epithelial (RPE) cells are one of the major cellular
components of the fibrotic membrane. Of the number of cytokines and
growth factors that have been previously reported to contribute to
PVR pathogenesis, transforming growth factor (TGF)-β is of
particular importance (3).
Gremlin is a highly conserved 184-amino-acid protein
that contains a cysteine-rich region (4–7).
Structurally, it is a member of the cysteine knot superfamily,
which can be present in both soluble and cell-associated forms
(8–11). Gremlin belongs to a family of bone
morphogenetic protein antagonists that participate in a number of
physiological processes, including cell survival, differentiation,
growth and development (4,8–12).
Gremlin is predominantly localised to the outer retina, and high
levels of its expression have been demonstrated in bovine retinal
pericytes in response to elevated glucose levels compared with
control treated pericytes (13). The
dysfunction of gremlin has also been observed to be associated with
a number of diseases, such as diabetic fibrotic disease (4,8–12,14).
Although it has been previously shown that, during
PVR, gremlin induces epithelial-to-mesenchymal transition (EMT) in
RPE cells (15), information on the
potential link between gremlin and the expression of pro-fibrogenic
factors in human RPE cells remain limited. The present study
demonstrated that gremlin increased the proliferation and
expression of fibronectin and type I collagen in human RPE cells,
whereas gremlin knockdown by small interfering (si)RNA expression
significantly reduced TGF-β1- and TGF-β2-induced expression of
fibronectin and type I collagen in human RPE cells.
Materials and methods
Reagents
Gremlin-1 (cat. no. SRP3285) and DAPI (cat. no.
D9542) were purchased from Sigma-Aldrich; Merck KGaA. Recombinant
human TGF-β1 and TGF-β2 were obtained from Cell Signaling
Technology, Inc. Anti-fibronectin (cat. no. ab2413) and anti-type I
collagen (cat. no. ab34710) antibodies, type I collagen (human
pro-collagen Iα, cat. no. ab229389) and fibronectin (cat. no.
ab219046) ELISA kits were purchased from Abcam. TRITC conjugated
goat anti-rabbit IgG secondary antibody (cat. no. ZF-0316) was
obtained from Zhongshan Golden Bridge Biotechnology Co., Ltd. Human
gremlin siRNA and siRNA control (cat. no. siP06969473939) were
purchased from Guangzhou RiboBio Co., Ltd. The Gremlin siRNA
sequence is CCACCTACCAAGAAGAAGA.
Cell culture
Human RPE cells (ARPE-19; CRL-2302) were purchased
from the American Type Culture Collection. The cells were cultured
in DMEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 ng/ml streptomycin at 37°C and 5% CO2
in a humidified atmosphere. ARPE-19 cells were transfected as
described below. Transfected ARPE-19 cells were incubated with
recombinant human TGF-β1 or TGF-β2.
Gremlin siRNA transfection
The cells were transfected with gremlin siRNA or
control siRNA (50 nM each). Transfections were performed using the
riboFECT™ CP regeant according to the manufacturer' protocol.
Assays were performed 48 h after transfection, including assessment
of mRNA and protein levels and immunofluorescent staining.
Cell viability and proliferation
assay
Cell viability and proliferation was measured using
MTT and 5′bromo-2-deoxyuridine (BrdU) assays. For MTT assay, RPE
cells were cultured in 96-well plates at 1.0×104
cells/well for 24 h and were partially starved in DMEM supplemented
with 1% FBS for 12 h, following which they were treated with
various concentrations (25, 50 and 100 ng/ml) of gremlin at various
time points (12, 24 and 48 h). MTT (5 mg/ml; 20 µl) was then added
to the culture medium, and the cells were incubated for an
additional 4 h at 37°C. DMSO was then added to each well to
dissolve the formazan crystals formed. Absorbance in each well was
measured at 490 nm using a microplate reader. The BrdU assay was
performed according to the manufacturer's protocol. A total of
5.0×103 cells were seeded in a 96-well plate for 24 h
and were partially starved in DMEM supplemented with 1% FBS for 12
h, and then treated with or without tested substances for 48 h at
37°C. A 10 µM concentration of BrdU was added to the medium.
Measurement of cell proliferation was quantified by BrDU ELISA kit
(Roche Diagnostics). Absorbance in each well was measured at 450 nm
with a microplate reader (Bio-Rad Laboratories, Inc.)
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to extract total RNA from the RPE cells.
cDNA synthesis was performed with 2 µg total RNA using RevertAid
First Strand cDNA Synthesis kit (Fermentas; Thermo Fisher
Scientific, Inc.) according to manufacturer's protocols. The
reaction mixtures contained the following: 1 µl oligo(dT) primer, 4
µl 5× reaction buffer, 1 µl Ribolock™ RNase Inhibitor, 2 µl 10 mM
dNTP mix and 1 µl RevertAid™ M-MuLV Reverse Transcriptase in a
total volume of 20 µl. Incubation was for 60 min at 42°C before the
reaction was terminated by heating at 70°C for 5 min. The ABI
Sequence Detector System 7500 (Thermo Fisher Scientific, Inc.) was
used to perform RT-qPCR. The PCR solution consisted of specific
primers (0.3 µM each), 2.5 µl cDNA, and 12.5 µl Maxima SYBR-Green
qPCR Master Mix (Fermentas; Thermo Fisher Scientific, Inc.) to a
final reaction volume of 25 µl. The sequences of the primers used
were as follows: Human fibronectin forward,
5′-GATAAATCAACAGTGGGAGC-3′ and reverse, 5′-CCCAGATCATGGAGTCTTTA-3′;
human type I collagen forward, 5′-TGGTGGTTATGACTTTGGTTACGAT-3 and
reverse, 5′-TGTGCGAGCTGGGTTCTTTCTA-3′; human GAPDH forward,
5′-TGTTCGACAGTCAGCCGCAT-3′ and reverse, 5′-ACTCCGACCTTCACCTTCCC-3′;
and human gremlin forward, 5′-AAGCGAGACTGGTGCAAAAC-3′ and reverse,
5′-CTTGCAGAAGGAGCAGGACT-3′. The thermocycling conditions were as
follows: Initial denaturation at 95°C for 10 min, followed by 39
cycles of 95°C for 15 sec, 61°C for 30 sec and 72°C for 30 sec. The
relative mRNA expression levels were calculated as the difference
between target and GAPDH expression levels using the
2−ΔΔCq method (16).
Immunofluorescence staining
Human RPE cells were fixed with 4% paraformaldehyde
for 20 min at 23°C, washed three times with PBS and permeabilised
using 0.5% Triton X-100 for 15 min at 23°C. The cells were
subsequently blocked with 10% goat serum (Zhongshan Golden Bridge
Biotechnology) for 1 h at 23°C before incubation with rabbit
anti-fibronectin (dilution, 1:100) or rabbit anti-collagen I
(dilution, 1:100) primary antibodies, which were diluted in PBS
supplemented with 10% goat serum, overnight at 4°C. The next day,
the cells were incubated with TRITC-conjugated goat anti-rabbit IgG
(dilution, 1:200) secondary antibodies at room temperature for 1 h.
The samples were counterstained with DAPI (1:1,000; cat. no. D9542,
Sigma-Aldrich; Merck KGaA) at room temperature for 10 min. The
samples were examined using a fluorescence microscope (DS-Ril-U2;
Nikon Corporation) and photographed at ×200 magnification in five
random fields (DS-U2; Nikon Corporation).
ELISA
The measurement of fibronectin protein levels of
using ELISA have been previously reported (17–19).
Cells were seeded at 4×103 cells per well in 12-well
plates. After reaching a confluency of approximately 85–90%, the
medium was changed to a serum-free medium for 12 h. Cells were
transfected with control siRNA or gremlin siRNA using the riboFECT™
CP reagent. After transfection, cells were incubated with
recombinant human TGF-β1 or TGF-β2 for 48 h at 37°C. The samples
were collected after the cells were treated. Fibronectin and type I
collagen levels in the culture supernatants of RPE cells were
measured using fibronectin and type I collagen ELISA kits according
to manufacturer's protocols.
Statistical analysis
All statistical differences were evaluated using
one-way analysis of variance followed by Tukey's test. All data are
presented as the mean ± SD and were analysed using SPSS 17.0
software (SPSS, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Gremlin induces human RPE cell
proliferation and increases fibronectin and type I collagen
expression
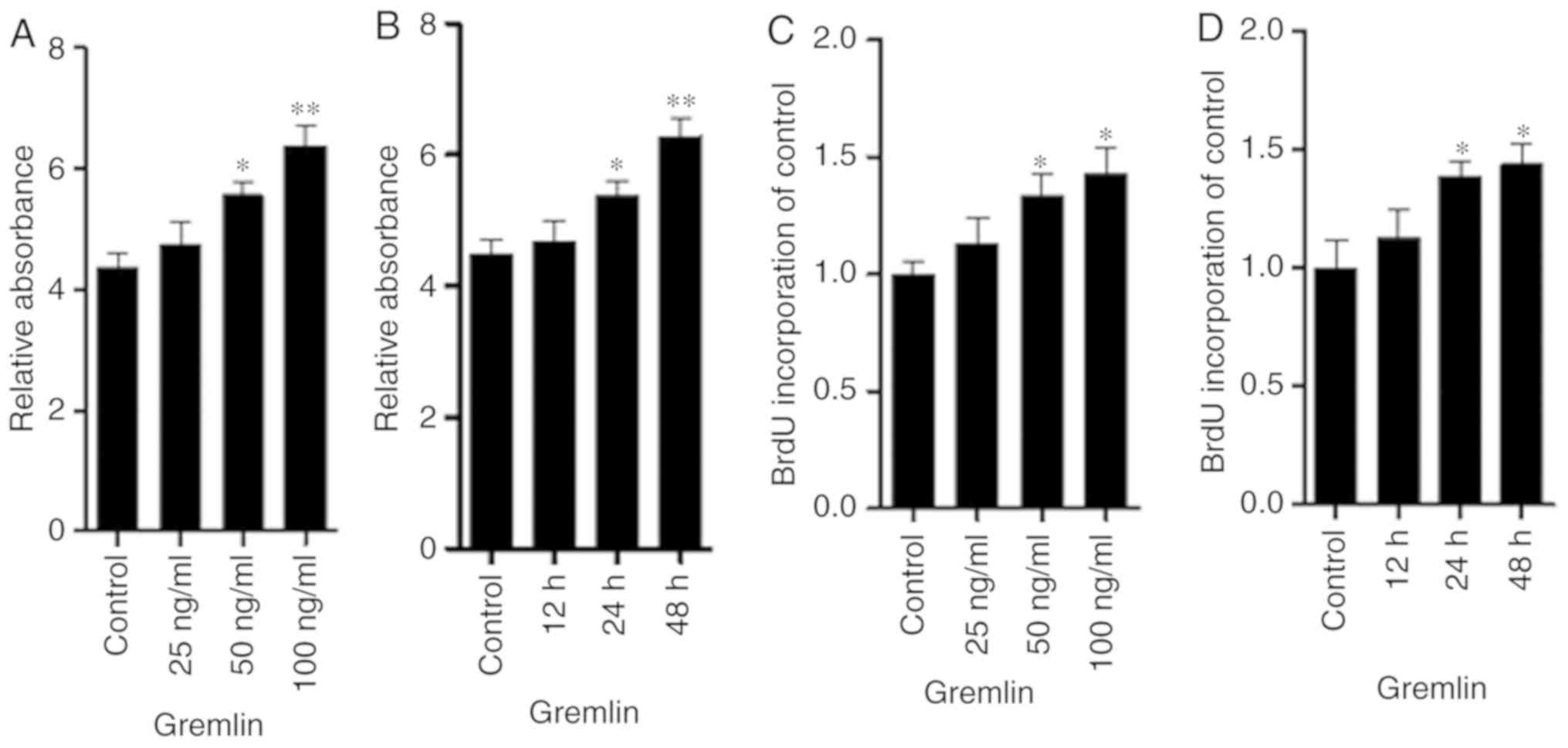
To examine the role of gremlin in the proliferation
and expression of fibronectin and type I collagen in human RPE
cells, cells were treated with ascending concentrations of gremlin
(0, 25, 50 and 100 ng/ml) for 48 h. At a concentration of 50 ng/ml
or 100 ng/ml, gremlin significantly increased RPE cell
proliferation compared with the control group (Fig. 1A and C). In addition, RPE cell
proliferation was significantly enhanced following incubation with
100 ng/ml gremlin for 24 and 48 h compared with the control group
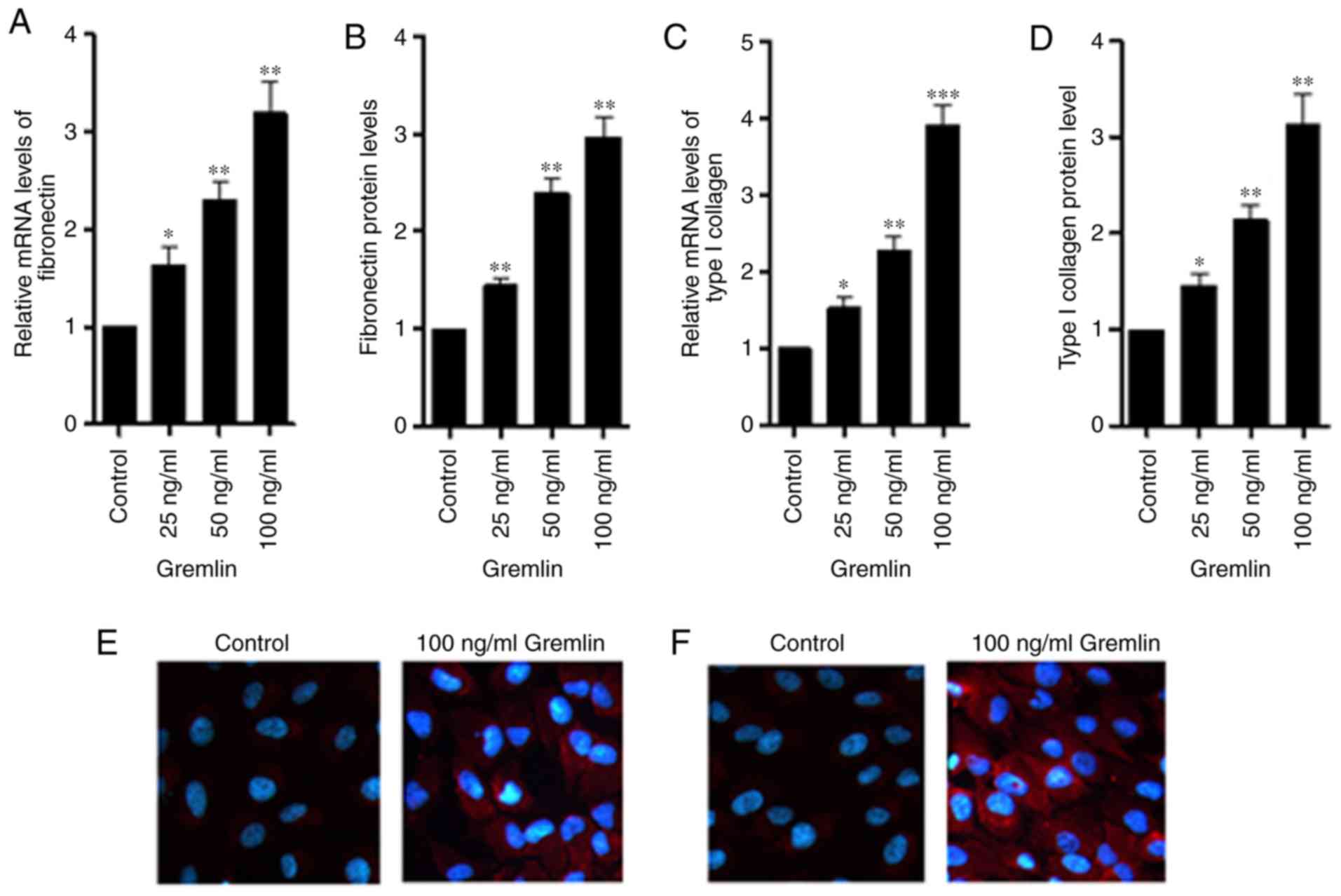
(Fig. 1B and D). Subsequently, the
mRNA and protein levels of fibronectin and type I collagen were
measured using RT-qPCR, ELISA and immunofluorescence staining. At a
concentration of 25, 50 or 100 ng/ml, recombinant gremlin
significantly increased the expression of fibronectin (Fig. 2A and B) and type I collagen (Fig. 2C and D) compared with the control
group. In addition, strong positive staining of fibronectin
(Fig. 2E) and type I collagen
(Fig. 2F) was observed following
treatment of RPE cells with gremlin (100 ng/ml, 48 h).
TGF-β1 and TGF-β2 increase the
expression of gremlin, fibronectin and type I collagen in human RPE
cells
The protein and mRNA levels of gremlin, fibronectin
and type I collagen were measured using ELISA and RT-qPCR following
treatment of RPE cells with TGF-β. TGF-β1 and TGF-β2 treatment
increased the expression of gremlin in human RPE cells, which was
significantly reversed by gremlin siRNA (Fig. 3A and B). TGF-β1 (10 mg/ml) increased
fibronectin (Fig. 3C and D) and type
I collagen (Fig. 3E and F)
expression in human RPE cells and their levels in the cell
supernatant. Additionally, TGF-β2 (5 ng/ml) significantly
upregulated the levels of fibronectin (Fig. 3G and H) and type I collagen (Fig. 3I and J) in human RPE cells and RPE
cell supernatant.
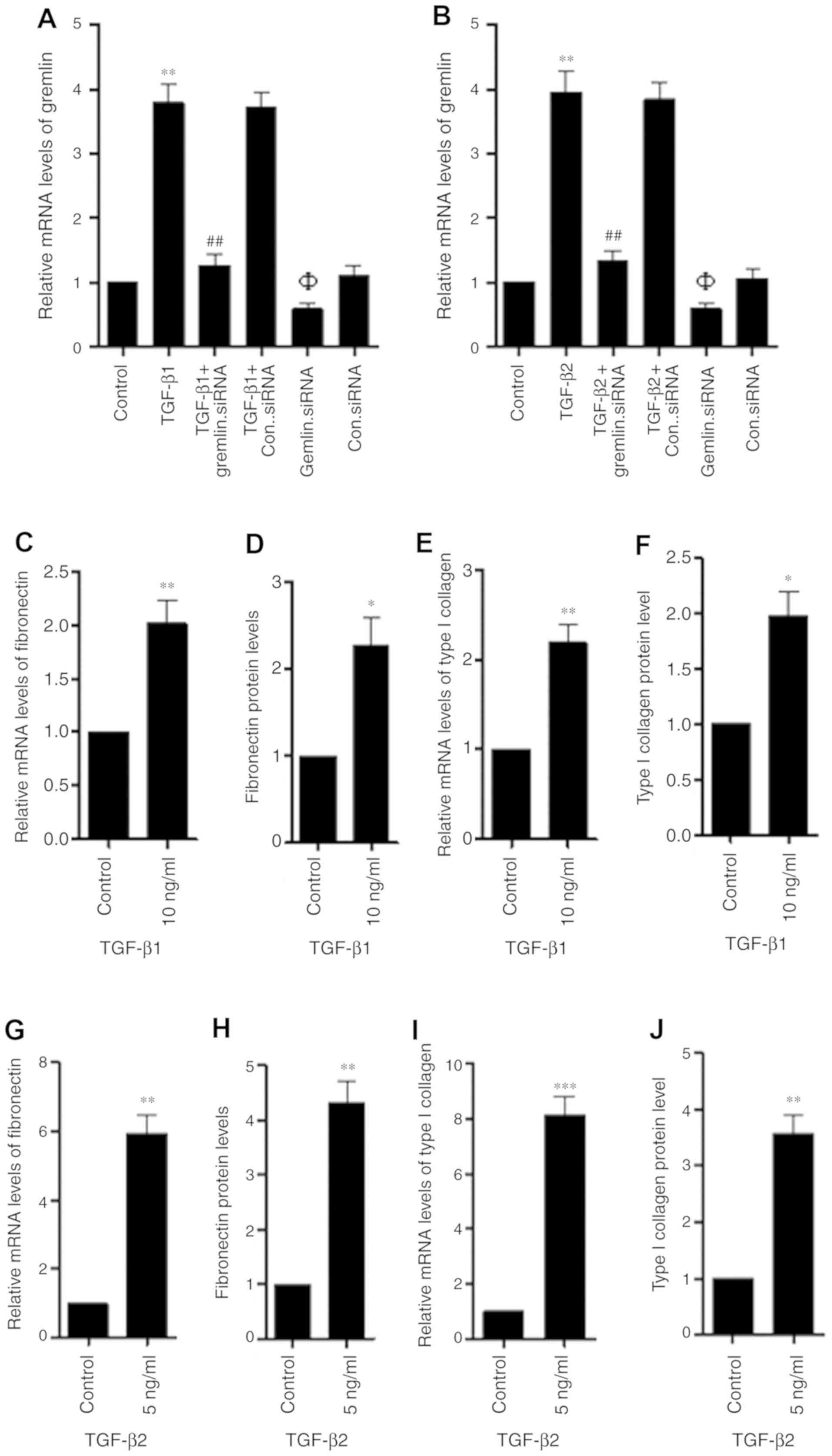 | Figure 3.TGF-β1 and TGF-β2 increase the
expression and secretion of gremlin, fibronectin and type I
collagen in human RPE cells. (A) RPE cells were subjected to TGF-β1
and/or gremlin siRNA transfection or (B) TGF-β2 and/or gremlin
siRNA transfection for 48 h, following which the relative
expression of gremlin mRNA was measured using reverse
transcription-quantitative PCR. (C) RPE cells were first treated
with 10 ng/ml TGF-β1 for 48 h before fibronectin mRNA expression,
(D) fibronectin secretion, (E) type I collagen mRNA expression and
(F) type I collagen secretion were measured. (G) RPE cells were
first treated with 5 ng/ml TGF-β2 for 48 h before fibronectin mRNA
expression, (H) fibronectin secretion, (I) type I collagen mRNA
expression and (J) type I collagen secretion were measured. The
data represent the mean ± SD of three independent experiments.
*P<0.05, **P<0.01 and ***P<0.001 vs. control;
##P<0.01 vs. respective TGF-β1 or TGF-β2 group;
ΦP<0.05 vs. Control group. TGF-β, transforming growth
factor-β; RPE, retinal pigment epithelial cells; siRNA, small
interfering RNA. Con. represents Control. |
Gremlin mediated TGF-β induces the
expression of fibronectin and type I collagen in human RPE
cells
Gremlin knockdown following TGF-β-treatment
decreased the expression of fibronectin and type I collagen in
human RPE cells. RT-qPCR and ELISA were used to measure the
expression or secretion of fibronectin and type I collagen. Gremlin
knockdown significantly reversed the TGF-β1-induced expression and
secretion of fibronectin (Fig. 4A and
B) and type I collagen (Fig. 4C and
D) compared with the TGF-β1-treated group. Compared with the
TGF-β2-treated group, gremlin knockdown also significantly reversed
TGF-β2 induced expression and secretion of fibronectin (Fig. 4E and F) and type I collagen (Fig. 4G and H).
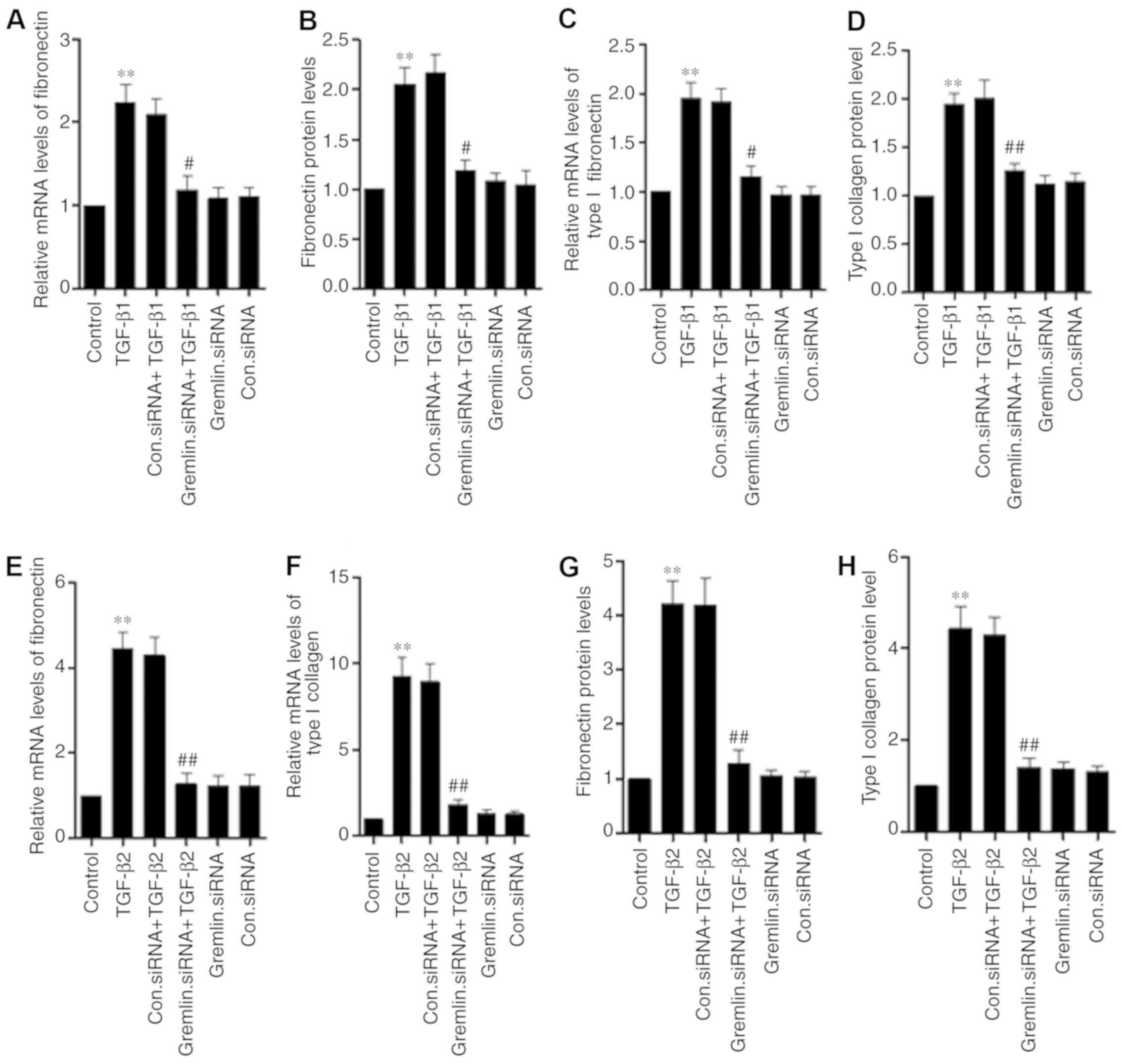 | Figure 4.Gremlin mediates the TGF-β-induced
induction of fibronectin and type I collagen expression and
secretion in human RPE cells. The expression and secretion of
fibronectin and type I collagen were measured using reverse
transcription-quantitative PCR and ELISA, respectively. (A)
Following treatment with TGF-β1 and/or gremlin knockdown,
fibronectin mRNA expression, (B) fibronectin secretion, (C) type I
collagen mRNA expression and (D) type I collagen secretion were
measured. (E) Following treatment with TGF-β2 and/or gremlin
knockdown, fibronectin mRNA expression, (F) fibronectin secretion,
(G) type I collagen mRNA expression and (H) type I collagen
secretion were measured. The data shown represent the mean ± SD of
three independent experiments. **P<0.01 vs. control;
#P<0.05, ##P<0.01 vs. respective TGF-β1
or TGF-β2 group. TGF-β, transforming growth factor-β; RPE, retinal
pigment epithelial cells. Con. represents Control. |
Discussion
Fibrotic membrane contractions can result in severe
visual impairment or blindness (20). RPE cell proliferation, migration and
ECM deposition are some of the main physiological changes that
occur during PVR, which is under the influence of a variety of
cytokines and growth factors (21–23). In
PVR, RPE cells undergo EMT and begin losing epithelial
characteristics whilst adopting a more mesenchymal, fibroblast-like
phenotype, resulting in increased proliferation and migration,
resistance to apoptosis and production of ECM proteins (24,25).
However, the relationship between gremlin and the expression of
proteins associated with the ECM in human RPE cells currently
remains unclear. In the present study, the role of gremlin in the
expression of fibronectin and type I collagen, and in RPE cell
proliferation, was investigated. It was found that gremlin
significantly increased the proliferation and expression of
fibronectin and type I collagen in RPE cells, suggesting that
gremlin may serve a role in the development of PVR.
TGF-β is a major cytokine involved in the regulation
of cell proliferation, migration, cell death, differentiation,
protein synthesis, and the pathological and physiological processes
associated with tissue repair and development (26). Elevated TGF-β levels have been
reported in the subretinal fluids, vitreous and epiretinal
membranes of patients with PVR (27–30).
TGF-β serves a vital role in PVR formation since it regulates cell
proliferation, enhances ECM protein synthesis and induces ECM
deposition (23,31). It has been previously demonstrated
that the inhibition of TGF-β expression may prevent PVR progression
(32). In addition, silencing of
gremlin expression has been shown to significantly reduce
TGF-β1-induced ECM expression in human tubular epithelial cells and
TGF-β2-induced ECM synthesis in human lens epithelial cells
(33,34). However, the relationship between
gremlin and TGF-β1 and TGF-β2 in human RPE cells remains currently
unclear. To clarify this, the results of the present study showed
that TGF-β1 and TGF-β2 increased gremlin expression in human RPE
cells, whilst gremlin knockdown significantly reversed TGF-β1- and
TGF-β2-induced ECM expression in human RPE cells. This observation
suggested that TGF-β1 and TGF-β2 can both induce the expression of
ECM proteins by increasing gremlin expression, further implicating
gremlin to be a mediator in PVR pathogenesis associated with
TGF-β.
The present study does have a number of limitations.
Apart from fibronectin and collagen type I, epithelial markers such
as E-cadherin and zonula occludens-1, and mesenchymal markers
including N-cadherin and α-smooth muscle actin, are also involved
in the development of PVR (35,36). In
the present study, the migration of RPE cells was not studied. In
addition, the levels of fibronectin and collagen I were measured
using ELISA and not by western blotting.
In conclusion, the data from the present study
suggested that gremlin can increase RPE cell proliferation and
induce fibronectin and collagen I expression in human RPE cells.
Silencing of gremlin expression significantly reduced TGF-β1- and
TGF-β2-induced fibronectin and collagen I expression in human RPE
cells. Thus, TGF-β induced fibronectin and collagen I expression
via regulation of gremlin. Therefore, gremlin may serve a role in
the development of PVR, which potentially extends our knowledge
regarding the role of gremlin in PVR pathogenesis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Department of
Science and Technology of Henan Province (grant no.
162300410112).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
DQ performed the experiments and wrote the paper. YJ
analysed the data and revised the paper. XJ designed the
experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sadaka A, Sisk RA, Osher JM, Toygar O,
Duncan MK and Riemann CD: Intravitreal methotrexate infusion for
proliferative vitreoretinopathy. Clin Ophthalmol. 10:1811–1817.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Claes C and Lafetá AP: Proliferative
vitreoretinopathy. Dev Ophthalmol. 54:188–195. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pena RA, Jerdan JA and Glaser BM: Effects
of TGF-beta and TGF-beta neutralizing antibodies on
fibroblast-induced collagen gel contraction: Implications for
proliferative vitreoretinopathy. Invest Ophthalmol Vis Sci.
35:2804–2808. 1994.PubMed/NCBI
|
|
4
|
Lappin DW, McMahon R, Murphy M and Brady
HR: Gremlin: An example of the re-emergence of developmental
programmes in diabetic nephropathy. Nephrol Dial Transplant. 17
(Suppl 9):S65–S67. 2002. View Article : Google Scholar
|
|
5
|
Grillo E, Ravelli C, Corsini M,
Ballmer-Hofer K, Zammataro L, Oreste P, Zoppetti G, Tobia C, Ronca
R, Presta M and Mitola S: Monomeric gremlin is a novel vascular
endothelial growth factor receptor-2 antagonist. Oncotarget.
7:35353–35368. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kišonaitė M, Wang X and Hyvönen M:
Structure of Gremlin-1 and analysis of its interaction with BMP-2.
Biochem J. 473:1593–1604. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yin Y, Yang Y, Yang L, Yang Y, Li C, Liu X
and Qu Y: Overexpression of Gremlin promotes non-small cell lung
cancer progression. Tumour Biol. 37:2597–2602. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wordinger RJ, Zode G and Clark AF: Focus
on molecules: Gremlin. Exp Eye Res. 87:78–79. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Müller I, Schönberger T, Schneider M,
Borst O, Ziegler M, Seizer P, Leder C, Müller K, Lang M,
Appenzeller F, et al: Gremlin-1 is an inhibitor of macrophage
migration inhibitory factor and attenuates atherosclerotic plaque
growth in ApoE−/− Mice. J Biol Chem. 288:31635–31645.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sethi A, Wordinger RJ and Clark AF:
Gremlin utilizes canonical and non-canonical TGFb signaling to
induce lysyl oxidase (LOX) genes in human trabecular meshwork
cells. Exp Eye Res. 113:117–127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cahill E, Costello CM, Rowan SC, Harkin S,
Howell K, Leonard MO, Southwood M, Cummins EP, Fitzpatrick SF,
Taylor CT, et al: Gremlin plays a key role in the pathogenesis of
pulmonary hypertension. Circulation. 125:920–930. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mitola S, Ravelli C, Moroni E, Salvi V,
Leali D, BallmerHofer K, Zammataro L and Presta M: Gremlin is a
novel agonist of the major proangiogenic receptor VEGFR2. Blood.
116:3677–3680. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kane R, Stevenson L, Godson C, Stitt AW
and O'Brien CO: Gremlin gene expression in bovine retinal pericytes
exposed to elevated glucose. Br J Ophthalmol. 89:1638–1642. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
McMahon R, Murphy M, Clarkson M, Taal M,
Mackenzie HS, Godson C, Martin F and Brady HR: IHG-2, a mesangial
cell gene induced by high glucose, is human gremlin. Regulation by
extracellular glucose concentration, cyclic mechanical strain, and
transforming growth factor-beta 1. J Biol Chem. 275:9901–9904.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee H, O'Meara SJ, O'Brien C and Kane R:
The role of Gremlin, a BMP antagonist, and
epithelial-to-mesenchymal transition in proliferative
vitreoretinopathy. Invest Ophthalmol Vis Sci. 48:4291–4299. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen J, Ma Y, Wang Z, Wang H, Wang L, Xiao
F, Wang H, Tan J and Guo Z: Thrombin promotes fibronectin secretion
by bone marrow mesenchymal stem cells via the protease-activated
receptor mediated signalling pathways. Stem Cell Res Ther.
5:362014. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang S, Shiels IA, Ambler JS and Taylor
SM: Pirfenidone reduces fibronectin synthesis by cultured human
retinal pigment epithelial cells. Aust N Z J Ophthalmol. 26 (Suppl
1):S74–S76. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Osusky R, Soriano D, Ye J and Ryan SJ:
Cytokine effect on fibronectin release by retinal pigment
epithelial cells. Curr Eye Res. 13:569–574. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kwon OW, Song JH and Roh MI: Retinal
detachment and proliferative vitreoretinopathy. Dev Ophthalmol.
55:154–162. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Por ED, Greene WA, Burke TA and Wang HC:
Trichostatin a inhibits retinal pigmented epithelium activation in
an in vitro model of proliferative vitreoretinopathy. J Ocul
Pharmacol Ther. 32:415–524. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pennock S, Haddock LJ, Mukai S and
Kazlauskas A: Vascular endothelial growth factor acts primarily via
platelet-derived growth factor recptor α to promote proliferative
vitreoretinopathy. Am J Pathol. 184:3052–3068. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xiao W, Chen X, Liu X, Luo L, Ye S and Liu
Y: Trichostatin A, a histone deacetylase inhibotor, suppresses
proliferation and epithelial-mesenchymal transition in retinal
pigment epithelial cells. J Cell Mol Med. 18:646–655. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rouberol F and Chiquet C: Proliferative
vitreoretinopathy: Pathophysiology and clinical diagnosis. J Fr
Ophtalmol. 37:557–565. 2014.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Winkler J and Hoerauf H: TGF-β and
RPE-derived cells in taut subretinal strands from patients with
proliferative vitreoretinopathy. Eur J Ophthalmol. 21:422–426.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saika S: TGF beta pathobiology in the eye.
Lab Invest. 86:106–115. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen YS, Hackett SF, Schoenfeld CL,
Vinores MA, Vinores SA and Campochiaro PA: Localisation of vascular
endothelial growth factor and its receptors to cells of vascular
and avascular epiretinal membranes. Br J Ophthalmol. 81:919–926.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bochaton-Piallat ML, Kapetanios AD, Donati
G, Redard M, Gabbiani G and Pournaras CJ: TGF-beta1, TGF-beta
receptor II and ED-A fibronectin expression in myofibroblast of
vitreoretinopathy. Invest Ophthalmol Vis Sci. 41:2336–2342.
2000.PubMed/NCBI
|
|
29
|
Gaudric A, Glacet-Bernard A, Clement G,
Falquerho L, Barritault D and Coscas G: Transforming growth factor
beta in the vitreous of patients with epiretinal proliferation.
Ophthalmologe. 4:51–52. 1990.(In French).
|
|
30
|
Hirase K, Sugiyama T, Ikeda T, Sotozono C,
Yasuhara T, Koizumi K and Kinoshita S: Transforming growth factor
beta (2) increases in subretinal fluid in rhegmatogenous retinal
detachment with subretinal strands. Ophthalmologica. 219:222–225.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cai W, Wei Q, Liu Q, Ren C, Liu J, Zhang
R, He M, Wang Q, Du Y and Yu J: Effect of bradykinin on
TGF-β1-induced retinal pigment epithelial cell proliferation and
extracellular matrix secretion. BMC Ophthalmol. 16:1992016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nassar K and Grisanti S, Tura A, Lüke J,
Lüke M, Soliman M and Grisanti S: A TGF-b receptor 1 inhibitor for
prevention of proliferative vitreoretinopathy. Exp Eye Res.
123:72–86. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rodrigues-Diez R, Lavoz C, Carvajal G,
Rayego-Mateos S, Rodrigues Diez RR, Ortiz A, Egido J, Mezzano S and
Ruiz-Ortega M: Gremlin is a downstream profibrotic mediator of
transforming growth factor-beta in cultured renal cells. Nephron
Exp Nephrol. 122:62–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ma B, Kang Q, Qin L, Cui L and Pei C:
TGF-b2 induces transdifferentiation and fibrosis in human lens
epithelial cells via regulating gremlin and CTGF. Biochem Biophys
Res Commun. 447:689–695. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feng H, Zhao X, Guo Q, Feng Y, Ma M, Guo
W, Dong X, Deng C, Li C, Song X, et al: Autophagy resists EMT
progress to maintain retinal pigment epithelium homeostasis. Int J
Biol Sci. 15:507–521. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shanmuganathan S, Sumantran VN and
Angayarkanni N: Epigallocatechin gallate and curcumin prevent
transforming growth factor beta 1-induced epithelial to mesenchymal
transition in ARPE-19 cells. Indian J Med Res. 146 (Suppl
2):S85–S96. 2017. View Article : Google Scholar : PubMed/NCBI
|