Introduction
Colorectal cancer (CRC) is one of the most common
malignant cancers worldwide, and is the third leading cause of
cancer-associated mortality in developed countries (1). A characteristic feature of CRC is the
rapid proliferation of cells in the colon and rectum. In China, it
was reported that the number of novel CRC cases was 376,300 and the
number of mortalities was 191,000 in 2015(2). In 2017 the incidence of CRC in the
general population worldwide was 5% and the 5-year survival rate
was between 40-60% (3). Despite
recent advances in the treatment of CRC, including surgical
resection, radiation therapy and chemotherapy, the survival rate of
patients is still low (4-6).
Therefore, the development of novel therapeutic options may confer
survival benefits on patients with CRC.
Previous studies have revealed (via integrated
genomic and transcriptome sequencing results) that >90% of DNA
sequences are actively transcribed, of which 98% are transcribed
into various non-coding RNAs, including microRNAs (miRNAs or miRs)
and long non-coding (lnc)RNAs (7,8). lncRNAs
are >200 nucleotides in length and do not code for proteins
(9). lncRNAs regulate gene
expression at different levels and serve a crucial role in multiple
biological processes (10,11). It has been reported that lncRNAs are
aberrantly expressed in mammalian and plant cells (12,13).
There is increasing evidence that lncRNAs may serve as novel
biomarkers for the diagnosis and prognosis of numerous types of
cancer, including hepatocellular carcinoma, gastric cancer,
non-small cell lung cancer and pancreatic cancer (14-19).
Colon cancer associated transcript 1 (CCAT1) is a
novel lncRNA that was identified via representative differential
analysis, cDNA cloning and the rapid amplification of cDNA ends.
The well-characterized cancer gene, c-Myc, is located in the
chromosomal 8q24 region, which is comprised of multiple genes
associated with CRC (20,21). Previous studies have revealed that
CCAT1 is activated and upregulated by c-Myc at the transcriptional
level, which results in the promotion of tumorigenesis,
progression, invasion and metastasis (22-24). miR-218 is a
tumor-suppressive miRNA that suppresses the proliferation of glioma
(25) and mesenchymal stem cells
(26). Additionally, miR-218
inhibits synovial osteogenic differentiation and inhibits the
angiogenesis of prostate cancer (27). However, the role of CCAT1 and miR-218
in CRC is yet to be fully elucidated. Therefore, the present study
aimed to investigate the expression and role of CCAT-1 and miR-218
in CRC.
Materials and methods
Tissue samples
A total of 30 CRC specimens and paired normal
adjacent cancer tissues (>2 cm from the tumor border) were
collected during colorectal resection from 30 patients with CRC (21
males, 9 females; age, 26-72 years old; median age, 51) during
gastrointestinal surgery in the Affiliated Suzhou Hospital of
Nanjing Medical University (Suzhou, China) between January 2016 and
January 2017. The exclusion criteria were: i) Patients that had
received pre-operative chemotherapy or radiotherapy treatments; ii)
patients that had other malignancies or infectious disease; iii)
patients with hereditary CRC syndromes. The present study was
approved by the Ethics Committee of the aforementioned Hospital and
all patients provided written informed consent.
Cell culture and transfection
Human CRC cells (SW480; ATCC) were cultured with
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) containing
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.)
at 37˚C in a 5% CO2 incubator. SW480 cells were
subsequently transfected with 1 µg short hairpin (sh)RNA
(CCAT1-shRNA; Guangzhou RiboBio Co., Ltd.), 1 µg negative control
shRNA (control-shRNA; Guangzhou RiboBio Co., Ltd.), 100 nM miR-218
inhibitors (5'-ACAUGGUUAGAUCAAGCACAA-3'; Guangzhou RiboBio Co.,
Ltd.), 100 nM miR-218 inhibitor controls (inhibitor controls;
5'-UCACAACCUCCUAGAAAGAGUAGA-3'; Guangzhou RiboBio Co., Ltd.), 1 µg
CCAT1-shRNA + 100 nM miR-218 inhibitors, 1 µg pcDNA3.1-CCAT1
(CCAT1-plasmids; Shanghai GenePharma Co., Ltd.), 1 µg control
pcDNA3.1 plasmids (control-plasmids; Shanghai GenePharma Co.,
Ltd.), 100 nM miR-218-mimics (5'-TTGTGCTTGATCTAACCATGT-3'; Shanghai
GenePharma Co., Ltd.), 100 nM miR-218 mimics negative controls
(mimic-controls: 5'-TTCTCCGAACGTGTCACGTTTC-3'; Shanghai GenePharma
Co., Ltd.) or 1 µg CCAT1-plasmid + 100 nM miR-218 mimics for 48 h
using Lipofectamine® 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
Transfection efficiency was determined 48 h post-transfection via
reverse transcription-quantitative (RT-q) PCR. Untreated cells were
used as controls.
MTT assay
SW480 cell viability was determined using an MTT
assay. Cells were seeded in 96-well plates at a density of
5x103 cells/well. After incubation at 37˚C for 48 h, 20
µl MTT reagent (Sigma-Aldrich; Merck KGaA) was added into each well
and incubated for a further 4 h at 37˚C. Subsequently, 150 µl
dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) was added into each
well and the suspension was agitated for 15 min. The optical
density was measured at 490 nm using a micro-plate reader.
Transwell assay
To investigate migration, SW480 cells
(2x104 cells per well) re-suspended in RPMI-1640 medium
were plated in the upper, uncoated Transwell chambers (pore size, 8
µm; Costar; Corning Inc.). RPMI-1640 medium with 10% FBS was plated
in the lower chamber to induce cell migration. After 24 h, the
migratory cells in the lower chamber were fixed with 4%
paraformaldehyde at room temperature for 30 min. Then, cells were
washed in triplicate with PBS and stained using 0.1% crystal violet
for 20 min at room temperature. The number of migrated cells were
counted under a light microscope at a magnification of x200 using
five random fields of view. For cell invasion assay, the upper
chamber was coated with 200 mg/ml of BD Matrigel™ Matrix (BD
Biosciences) at 37˚C for 30 min. The same protocol was followed as
described above for the migration assay.
RT-qPCR
Tissue samples and SW480 cells were treated with
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.) and
total RNA was extracted according to the manufacturer's protocol.
cDNA was then synthesized using the miScript Reverse Transcription
kit (Qiagen GmbH) according to the manufacturer's protocol. The
temperature protocol for this step was as follows: 70˚C for 5 min,
37˚C for 5 min and 42˚C for 1 h. qPCR was then performed using
QuantiFast SYBR-Green PCR kit (Qiagen GmbH) in accordance with the
manufacturer's protocol. The thermocycling conditions were as
follows: 95˚C for 3 min, followed by 40 cycles of 95˚C for 30 sec,
56˚C for 30 sec and 72˚C for 30 sec. GAPDH or U6 were used as
internal controls for normalization. The primer sequences were as
follows: CCAT1 forward, 5'-AGAAACACTATCACCTACGC-3' and reverse,
5'-CTTAACAGGGCATTGCTAATCT-3'; GAPDH forward,
5'-TGTTGCCATCAATGACCCCTT-3' and reverse, 5'-CTCCACGACGTACTCAGCG-3';
U6 forward, 5'-GCTTCGGCAGCACATATACTAAAAT-3' and reverse
5'-CGCTTCACGAATTTGCGTGTCAT-3'; miR-218 forward,
5'-GTTGTGCTTGATCTAACCATGT-3' and reverse, 5'-CTCGCTTCGGCAGCACA-3';
vascular endothelial growth factor (VEGF) forward,
5'-GAGCCTTGCCTTGCTGCTCTAC-3' and reverse,
5'-CACCAGGGTCTCGATTGGATG-3'. Relative gene expression was
calculated using the 2-ΔΔCq method
(28).
Western blotting
Total protein from SW480 cells was extracted using
radioimmunoprecipitation assay (RIPA) buffer (Beyotime Institute of
Biotechnology) with phenylmethylsulfonyl fluoride (PMSF; Beyotime
Institute of Biotechnology). The protein concentration was
subsequently determined using a bicinchoninic acid assay kit
(Beyotime Institute of Biotechnology). A total of 30 µg of protein
was loaded per lane and separated via 12% SDS-PAGE. Samples were
then transferred to PVDF membranes (Bio-Rad Laboratories, Inc.),
which were subsequently were blocked using 5% skimmed milk in TBS
containing 0.1% Tween (Beyotime Institute of Biotechnology) at room
temperature for 1.5 h. Membranes were incubated with the following
primary antibodies: VEGF (1:1,000; cat no. ab53465; Abcam) and
β-actin (1:1,000; cat no. 4970; Cell Signaling Technology, Inc.) at
4˚C overnight. Subsequently, the membranes were incubated with
horseradish peroxidase-conjugated anti-rabbit IgG secondary
antibodies (1:2,000; cat no. 7074; Cell Signaling Technology, Inc.)
at room temperature for 1 h. Protein bands were visualized using an
enhanced chemiluminescence assay (EMD Millipore). β-actin served as
the loading control for normalization.
Dual-luciferase reporter assay
Starbase version 3.0 (http://starbase.sysu.edu.cn/) was used to identify
associations between CCAT1 and miR-218. The results indicated that
there were complementary binding sites between miR-218 and CCAT1.
The fragment of CCAT1 containing the target sequence of miR-218 was
amplified via RT-qPCR (as previously described) and then inserted
into a pmirGLO vector (Promega Corporation) to form the wild-type
CCAT1 reporter vector (CCAT1-WT). An additional expression vector
was also constructed by inserting a mutated binding site and was
termed CCAT1-mutated-type (CCAT1-MUT). SW480 cells were seeded into
24-well plates at a density of 5x104 cells per well.
When the confluency reached ~80%, SW480 cells were co-transfected
with CCAT1-WT or CCAT1-MUT and miR-218 mimics or the negative
control at 37˚C for 48 h using Lipofectamine 2000®
(Invitrogen; Thermo Fisher Scientific, Inc.), in accordance with
the manufacturer's protocol. The relative luciferase activity was
then determined using a Dual-Luciferase Reporter assay system
(Promega Corporation) according to the manufacturer's protocol.
Luciferase activity was normalized to the activity of
Renilla.
Statistical analysis
Experiments were repeated three times and data are
presented as the mean ± SD. Statistical analyses were performed
using SPSS version 18.0 software (SPSS, Inc.). Significant
differences between groups was calculated using a paired Student's
t-test or one-way ANOVA followed by Tukey's post-hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression of CCAT1 in patients with
CRC
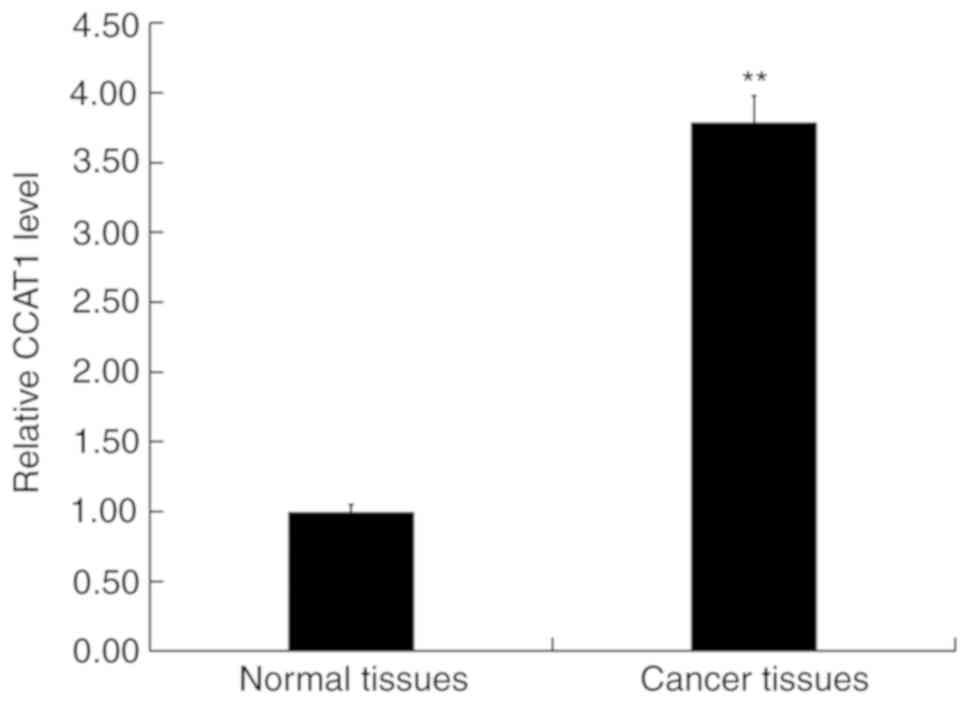
RT-qPCR was performed to detect the expression of
CCAT1 in CRC and adjacent normal tissues. The results revealed that
when compared with adjacent normal tissues, CCAT1 expression was
significantly upregulated in CRC (Fig.
1).
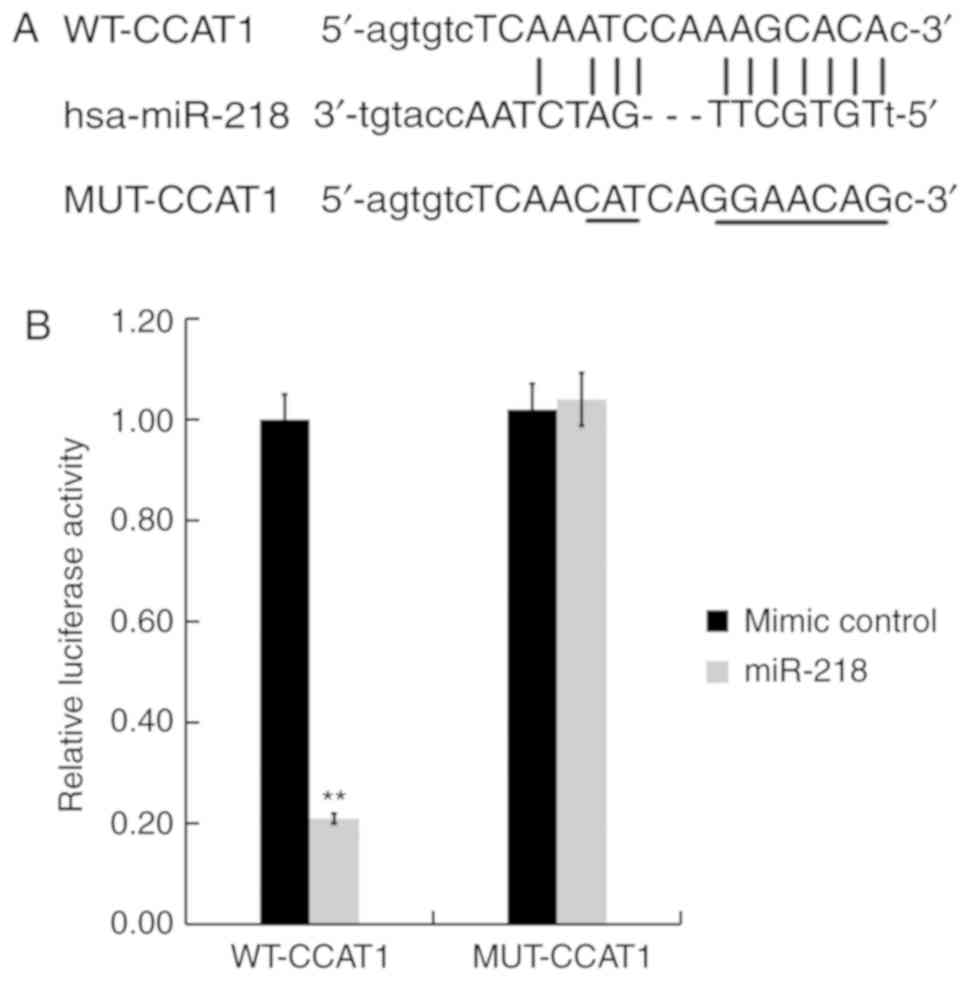
CCAT1 directly binds to miR-218
To verify the association between CCAT1 and miR-218,
bioinformatics analysis was performed using Starbase version 3.0 to
identify potential binding sites between CCAT1 and miR-218. The
results indicated that a complementary binding site existed between
CCAT1 and miR-218 (Fig. 2A). A
dual-luciferase reporter assay was then performed to validate the
predicted result. SW480 cells were co-transfected with a luciferase
plasmid containing the CCAT1 sequence (CCAT1-WT or CCAT1-MUT) and
the miR-218 mimic or the negative control. The results demonstrated
that the miR-218 mimic significantly inhibited the luciferase
activity of CCAT1-WT, while the miR-218 mimic did not exert an
inhibitory effect on CCAT1-MUT (Fig.
2B). Taken together, the results indicated that CCAT1 may bind
directly to miR-218.
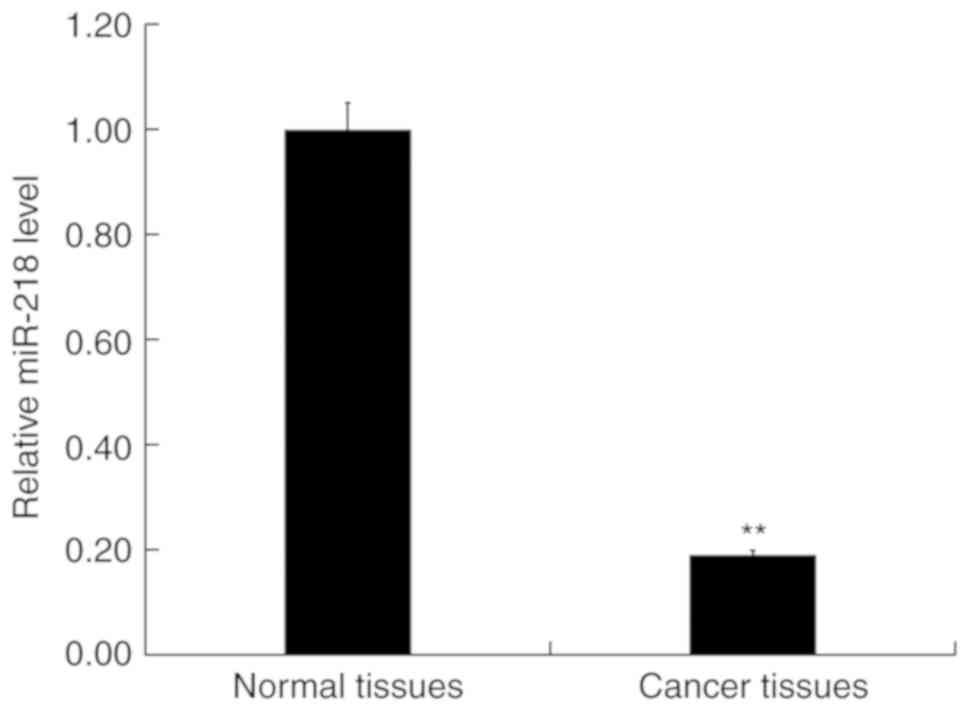
Expression of miR-218 in patients with
CRC
RT-qPCR was performed to detect the expression of
miR-218 in 30 CRC and paired adjacent normal tissues. The results
indicated that the expression of miR-218 was significantly
downregulated in CRC tissues compared with normal tissues (Fig. 3).
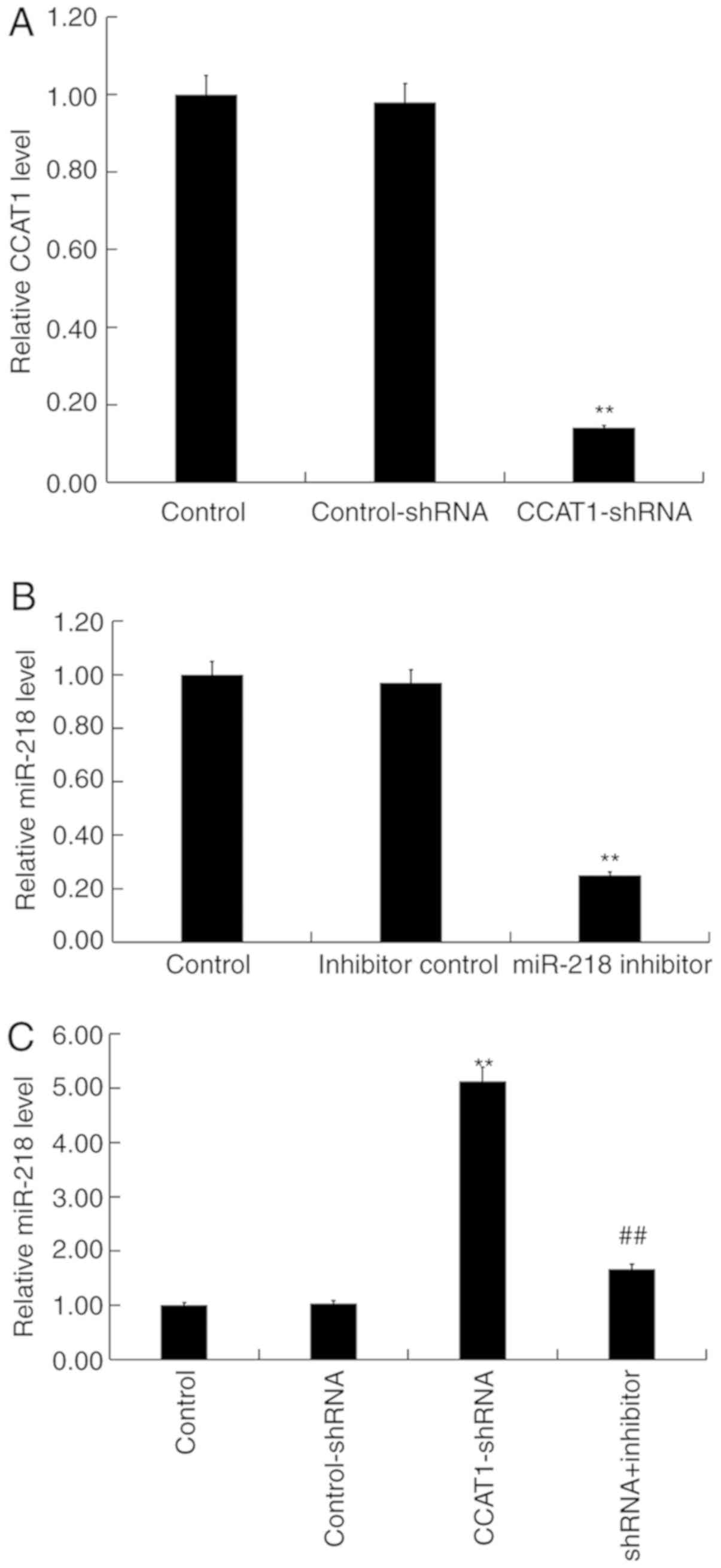
Downregulation of CCAT1 is associated
with an increase of miR-218 expression in SW480 cells
SW480 cells were transfected with either
CCAT1-shRNA, control-shRNA, miR-218 inhibitors, inhibitor controls
or CCAT1-shRNA + miR-218 inhibitor for 48 h, respectively.
Subsequently, RT-qPCR was performed to detect transfection
efficiency. When compared with the control group, CCAT1-shRNA
significantly reduced the expression of CCAT1 in SW480 cells
(Fig. 4A) whilst the miR-218
inhibitor significantly decreased the expression of miR-218
(Fig. 4B). Furthermore, the results
of the present study indicated that CCAT1-shRNA significantly
increased the expression of miR-218 in SW480 cells, which was
partially reversed following treatment with the miR-218 inhibitor
(Fig. 4C).
Effect of CCAT1 knockdown on SW480
cell viability, migration and invasion
To investigate the influence of CCAT1 on the
biological activity of SW480 cells, cells were transfected with
CCAT1-shRNA, control-shRNA or CCAT1-shRNA + miR-218 inhibitor for
48 h. The results of the MTT assay revealed that, when compared
with the control group, CCAT1-shRNA significantly inhibited the
viability of SW480 cells (Fig. 5A).
Furthermore, the results of the Transwell assay indicated that
CCAT1-shRNA significantly suppressed SW480 cell migration and
invasion (Fig. 5B-E). Western blotting demonstrated that
CCAT1-shRNA markedly reduced VEGF expression in SW480 cells at the
protein level (Fig. 5F).
Additionally, CCAT1-shRNA significantly reduced the mRNA expression
of VEGF in SW480 cells (Fig. 5G).
The aforementioned effects occurring after CCAT1-shRNA treatment
were all partially reversed by co-transfection with a
miR-218-inhibitor. The current results indicated that CCAT1-shRNA
inhibited CRC cell viability, migration and invasion by
downregulating VEGF expression via miR-218 modulation.
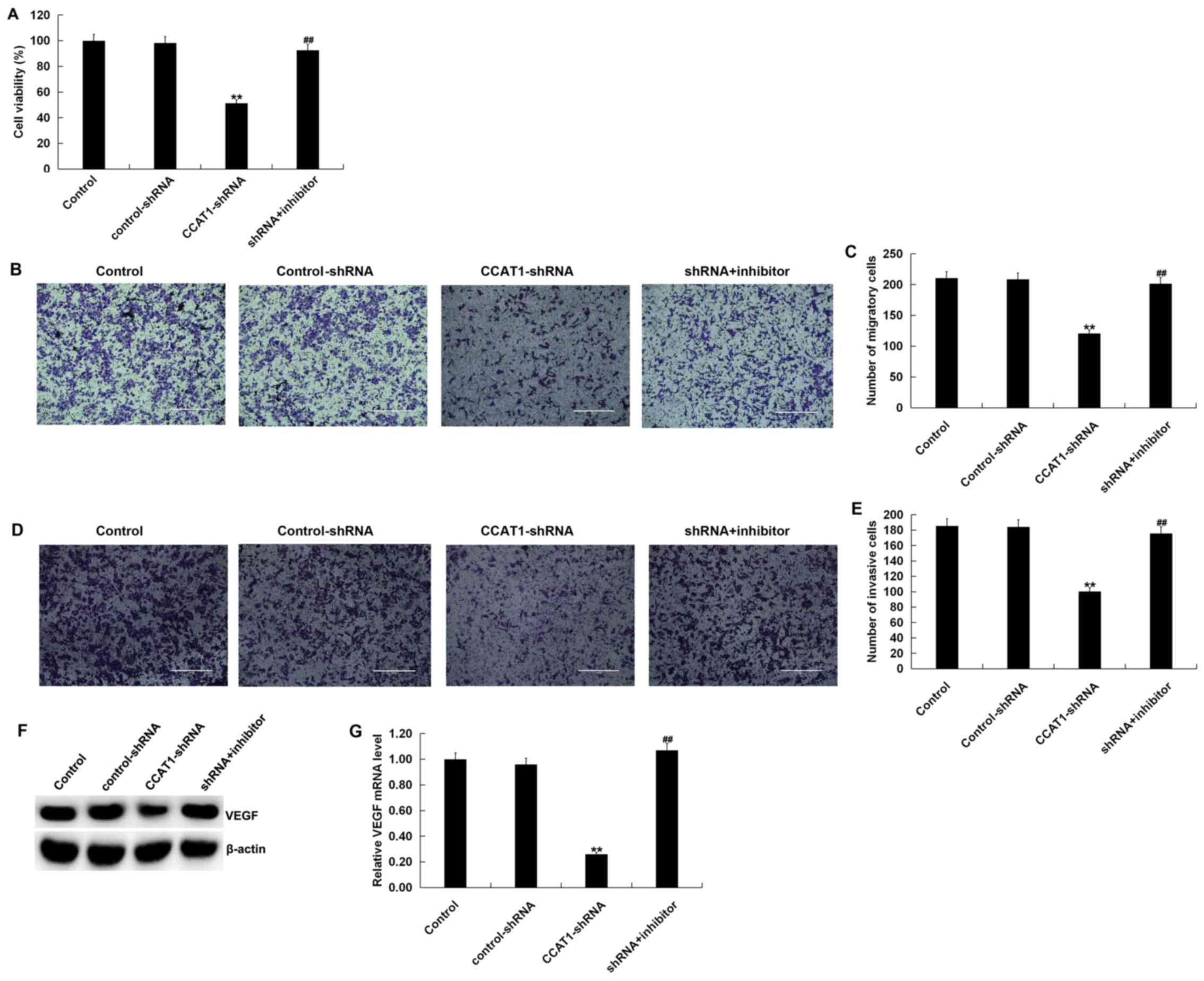 | Figure 5Knockdown of CCAT1 suppresses the
viability, migration and invasiveness of SW480 cells. SW480 cells
were transfected with CCAT1-shRNA, control-shRNA or
CCAT1-shRNA+miR-218 inhibitor for 48 h. (A) Cell viability was
detected using an MTT assay. Cell migration was subsequently
determined using a Transwell assay. (B) Cell images and (C) the
number of migratory cells are presented. Invasion was detected
using a Transwell assay with Matrigel. Results are presented as (D)
cell images and (E) the number of invasive cells. (F) Western
blotting was performed to measure the protein expression of VEGF in
SW480 cells transfected with CCAT1-shRNA, control-shRNA, or
CCAT1-shRNA + miR-218 inhibitor. (G) Reverse
transcription-quantitative PCR was used to detect the mRNA
expression of VEGF in SW480 cells transfected with CCAT1-shRNA,
control-shRNA or CCAT1-shRNA + miR-218 inhibitor. Data were
presented as the mean ± SD. **P<0.01 vs.
control; ##P<0.01 vs. CCAT1-shRNA. Scale bars, 400
µm. CCAT1, colon cancer associated transcript 1; shRNA, short
hairpin RNA; miR, microRNA; VEGF, vascular endothelial growth
factor. |
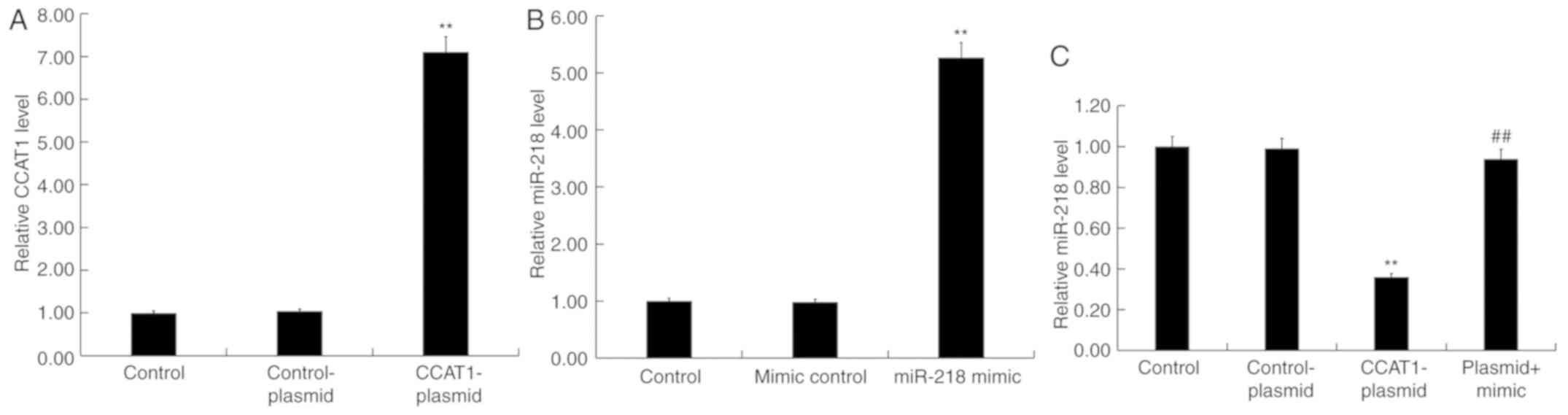
CCAT1 upregulation inhibits miR-218
expression in SW480 cells
To further investigate the role of CCAT1 and miR-218
in CRC, SW480 cells were transfected with CCAT1-plasmids,
control-plasmids, miR-218-mimics, mimic controls or CCAT1-plasmid +
miR-218 mimics for 48 h. Subsequently, an RT-qPCR assay was
performed to detect transfection efficiency. The results revealed
that the CCAT1-plasmid significantly increased the expression of
CCAT1 in SW480 cells compared with the control group (Fig. 6A). Furthermore, the miR-218 mimic
significantly increased the expression of miR-218 in SW480 cells
compared with the control group (Fig.
6B). Treatment with the CCAT1-plasmid significantly reduced the
expression of miR-218 in SW480 cells compared with the control
group, which was partially rescued by co-transfection with the
miR-218 mimic (Fig. 6C).
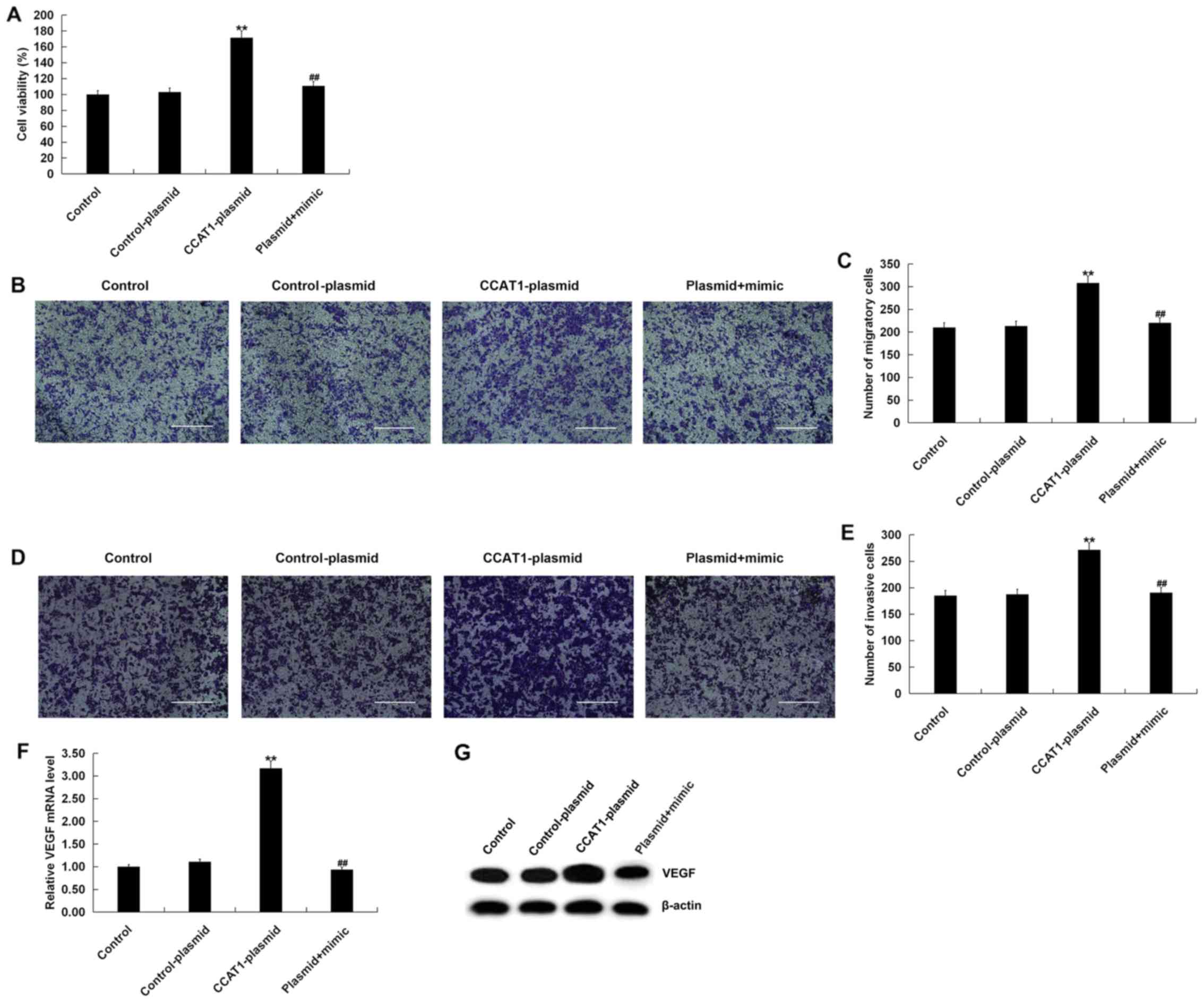
Increased expression of CCAT1
increases the viability, migration and invasiveness of SW480
cells
To investigate the effect of high CCAT1 levels on
the viability, migration and invasiveness of SW480 cells, MTT and
Transwell assays were performed. The MTT assay revealed that,
compared with the control group, transfection with the
CCAT1-plasmid significantly promoted the viability of SW480 cells
(Fig. 7A). The Transwell assay
indicated that transfection with a CCAT1-plasmid increased SW480
cell migration (Fig. 7B and C) and
invasiveness (Fig. 7D and E).
Furthermore, CCAT1-plasmid transfection increased VEGF expression
in SW480 cells at the mRNA and protein levels (Fig. 7F and G). All effects were partially
reversed following co-transfection with a miR-218 mimic. Taken
together, the results indicated that the CCAT1-plasmid increased
CRC cell viability, migration and invasiveness by promoting VEGF
expression via miR-218 modulation.
Discussion
A number of studies have suggested that the lncRNA
CCAT1 influences the progression of different types of cancer
(20-24,29). The present study focused on the investigation of the
role of lncRNA CCAT1 in CRC cells. Increasing evidence has revealed
that lncRNAs serve a crucial role in the occurrence, progression
and metastasis of various types of cancer (30-32).
It has been reported that lncRNAs affect the tumorigenesis of CRC
and are associated with clinical outcomes (33). Jiang et al (34) reported that lncRNA-gradually
increased during hepatocarcinogenesis (GIHCG) was highly expressed
in CRC tissues and cell lines, and that GIHCG deficiency suppressed
cell migration and invasion. Shi et al (35) also indicated that lncRNA-ZNFX1
antisense RNA 1 (ZNFX1-AS1) was upregulated in CRC tissues and
cells, and ZNFX1-AS1 knockdown inhibited cell proliferation. In
addition, previous studies have indicated that increased
lncRNA-CCAT2 expression may serve as a potential biomarker for CRC
diagnosis, and may also serve as an independent predictor of
prognosis (36,37). In the present study, CCAT1 expression
was significantly upregulated in CRC tissue.
Bioinformatics analysis indicated that CCAT1 and
miR-218 possess complementary binding sites. miR-218 has been
identified as a tumor-suppressive factor in cancers, including
cervical cancer, osteosarcoma and prostate cancer (38-40).
Zhu et al (38) demonstrated
that miR-218 is downregulated in cervical cancer and that miR-218
overexpression suppresses cervical cancer cell viability and
induces cell apoptosis. Additionally, Xuan et al (39) reported that miR-218 was significantly
downregulated in osteosarcoma cells and that treatment with miR-218
suppressed osteosarcoma formation in vivo. Zhang et
al (40) demonstrated that
lncRNA-prostate cancer antigen 3 (PCA3) regulated prostate cancer
by sponging miR-218-5p and modulating high mobility group box 1. In
the present study, it was demonstrated that miR-218 was
downregulated in CRC tissues.
VEGF, a potent angiogenic factor, serves a key role
in the formation of new blood vessels in CRC (41). VEGF is upregulated in CRC tissues and
anti-VEGF therapy has been applied to patients with metastatic CRC
(42,43). A previous study demonstrated that
CCAT1 downregulation decreased thyroid cancer cell viability,
proliferation, migration and invasion, and reduced VEGF expression
(44). A further study revealed that
miR-218 overexpression inhibited cell viability, promoted apoptosis
and decreased the expression of VEGF in cervical cancer cells
(40). The current study therefore
hypothesized that CCAT1 inhibition may influence CRC cell viability
and metastasis by downregulating VEGF expression via miR-218
sponging. Thus, the effect of CCAT1 on CRC cells and VEGF
expression was investigated. The results revealed that CCAT1 was
knocked down or overexpressed in CRC cells. The results further
indicated that CCAT1 knockdown inhibited CRC cell viability,
migration and invasion. Additionally, CCAT1 knockdown decreased the
expression of VEGF in CRC cells; however, these inhibitory effects
were partially reversed by co-transfection with a miR-218
inhibitor. CCAT1 overexpression exerted the opposite effect to
CCAT1 knockdown in CRC cells. Taken together, the current results
may provide novel insights into the roles and interactions of
lnc-CCAT1 and miR-218 in CRC. However, this study only assessed the
effects of CCAT1 on CRC cells in vitro, and in vivo
studies should be performed to reveal the present results. This is
a key limitation of this study.
In summary, CCAT1 influenced the expression of VEGF
in CRC cells by negatively regulating miR-218. In addition, CCAT1
was revealed to mediate multiple biological functions underpinning
the progression of CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Youth Science
and Technology Project of Promoting Health through Science and
Education in Suzhou (grant no. KJXW2016029), the National Natural
Science Foundation of China (grant no. 81804098) and the Jiangsu
Province Youth Medical Talent Project (grant no. QNRC2016255).
Availability of data and materials
All data sets used and/or generated during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
CG and SZ contributed to study design, data
collection, statistical analysis, data interpretation and
manuscript preparation. CH, JZ, RQ, QW, JQ and MZ contributed to
data collection and statistical analysis. SY and ZY contributed to
data collection, statistical analysis and manuscript preparation.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Human Ethics
Committee of the Affiliated Suzhou Hospital of Nanjing Medical
University (approval no. KL901058) and all patients provided
written informed consent.
Patient consent for publication
Patients provide written informed consent for the
publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. Ca
Cancer J Clin. 65:87–108. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kahouli I, Tomaro-Duchesneau C and Prakash
S: Probiotics in colorectal cancer (CRC) with emphasis on
mechanisms of action and current perspectives. J Med Microbiol.
62:1107–1123. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Su WB and Liu ZY: MiR-431 inhibits
colorectal cancer cell invasion via repressing CUL4B. Eur Rev Med
Pharmacol Sci. 22:3047–3052. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Goldstein DA, Zeichner SB, Bartnik CM,
Neustadter E and Flowers CR: Metastatic colorectal cancer: A
systematic review of the value of current therapies. Clin
Colorectal Cancer. 15:1–6. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Amaral PP, Dinger ME, Mercer TR and
Mattick JS: The eukaryotic genome as an RNA machine. Science.
319:1787–1789. 2008.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Guttman M, Amit I, Garber M, French C, Lin
MF, Feldser D, Huarte M, Zuk O, Carey BW, Cassady JP, et al:
Chromatin signature reveals over a thousand highly conserved large
non-coding RNAs in mammals. Nature. 458:223–227. 2009.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Nagano T and Fraser P: No-nonsense
functions for long noncoding RNAs. Cell. 145:178–181.
2011.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lekka E and Hall J: Noncoding RNAs in
disease. FEBS Lett. 592:2884–2900. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Kopp F and Mendell JT: Functional
classification and experimental dissection of long noncoding RNAs.
Cell. 172:393–407. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Liu Y, Ferguson JF, Xue C, Ballantyne RL,
Silverman IM, Gosai SJ, Serfecz J, Morley MP, Gregory BD, Li M and
Reilly MP: Tissue-specific RNA-Seq in human evoked inflammation
identifies blood and adipose LincRNA signatures of cardiometabolic
diseases. Arterioscler Thromb Vasc Biol. 34:902–912.
2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu YW, Sun M, Xia R, Zhang EB, Liu XH,
Zhang ZH, Xu TP, De W, Liu BR and Wang ZX: LincHOTAIR
epigenetically silences miR34a by binding to PRC2 to promote the
epithelial-to-mesenchymal transition in human gastric cancer. Cell
Death Dis. 6(e1802)2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chang S, Chen B, Wang X, Wu K and Sun Y:
Long non-coding RNA XIST regulates PTEN expression by sponging
miR-181a and promotes hepatocellular carcinoma progression. BMC
Cancer. 17(248)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Lu W, Zhang H, Niu Y, Wu Y, Sun W, Li H,
Kong J, Ding K, Shen HM, Wu H, et al: Long non-coding RNA linc00673
regulated non-small cell lung cancer proliferation, migration,
invasion and epithelial mesenchymal transition by sponging
miR-150-5p. Mol Cancer. 16(118)2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zheng J, Huang X, Tan W, Yu D, Du Z, Chang
J, Wei L, Han Y, Wang C, Che X, et al: Pancreatic cancer risk
variant in LINC00673 creates a miR-1231 binding site and interferes
with PTPN11 degradation. Nat Genet. 48:747–757. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
18
|
Sha M, Lin M, Wang J, Ye J, Xu J, Xu N and
Huang J: Long non-coding RNA MIAT promotes gastric cancer growth
and metastasis through regulation of miR-141/DDX5 pathway. J Exp
Clin Cancer Res. 37(58)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Lu Z, Li Y, Wang J, Che Y, Sun S, Huang J,
Chen Z and He J: Long non-coding RNA NKILA inhibits migration and
invasion of non-small cell lung cancer via NF-κB/Snail pathway. J
Exp Clin Cancer Res. 36(54)2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Nissan A, Stojadinovic A,
Mitrani-Rosenbaum S, Halle D, Grinbaum R, Roistacher M, Bochem A,
Dayanc BE, Ritter G, Gomceli I, et al: Colon cancer associated
transcript-1: A novel RNA expressed in malignant and pre-malignant
human tissues. Int J Cancer. 130:1598–1606. 2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ye Z, Zhou M, Tian B, Wu B and Li J:
Expression of lncRNA-CCAT1, E-cadherin and N-cadherin in colorectal
cancer and its clinical significance. Int J Clin Exp Med.
8:3707–3715. 2015.PubMed/NCBI
|
|
22
|
Xiang JF, Yin QF, Chen T, Zhang Y, Zhang
XO, Wu Z, Zhang S, Wang HB, Ge J, Lu X, et al: Human colorectal
cancer-specific CCAT1-L lncRNA regulates long-range chromatin
interactions at the MYC locus. Cell Res. 24:513–531.
2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Alaiyan B, Ilyayev N, Stojadinovic A,
Izadjoo M, Roistacher M, Pavlov V, Tzivin V, Halle D, Pan H, Trink
B, et al: Differential expression of colon cancer associated
transcript1 (CCAT1) along the colonic adenoma-carcinoma sequence.
BMC Cancer. 13(196)2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yang F, Xue X, Bi J, Zheng L, Zhi K, Gu Y
and Fang G: Long noncoding RNA CCAT1, which could be activated by
c-Myc, promotes the progression of gastric carcinoma. J Cancer Res
Clin Oncol. 139:437–445. 2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Gao Y, Sun L, Wu Z, Xuan C, Zhang J, You Y
and Chen X: miR218 inhibits the proliferation of human glioma cells
through downregulation of Yin Yang 1. Mol Med Rep. 17:1926–1932.
2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Cong R, Tao K, Fu P, Lou L, Zhu Y, Chen S,
Cai X and Mao L: MicroRNA218 promotes prostaglandin E2 to inhibit
osteogenic differentiation in synovial mesenchymal stem cells by
targeting 15-hydroxyprostaglandin dehydrogenase [NAD(+)]. Mol Med
Rep. 16:9347–9354. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Guan B, Wu K, Zeng J, Xu S, Mu L, Gao Y,
Wang K, Ma Z, Tian J, Shi Q, et al: Tumor-suppressive microRNA-218
inhibits tumor angiogenesis via targeting the mTOR component RICTOR
in prostate cancer. Oncotarget. 8:8162–8172. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
29
|
He X, Tan X, Wang X, Jin H, Liu L, Ma L,
Yu H and Fan Z: c-Myc-activated long noncoding RNA CCAT1 promotes
colon cancer cell proliferation and invasion. Tumour Biol.
35:12181–12188. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liang WC, Ren JL, Wong CW, Chan SO, Waye
MM, Fu WM and Zhang JF: LncRNA-NEF antagonized epithelial to
mesenchymal transition and cancer metastasis via cis-regulating
FOXA2 and inactivating Wnt/β-catenin signaling. Oncogene.
37:1445–1456. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wang ZY, Hu M, Dai MH, Xiong J, Zhang S,
Wu HJ, Zhang SS and Gong ZJ: Upregulation of the long non-coding
RNA AFAP1-AS1 affects the proliferation, invasion and survival of
tongue squamous cell carcinoma via the Wnt/β-catenin signaling
pathway. Mol Cancer. 17(3)2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Huang Y, Zhang J, Hou L, Wang G, Liu H,
Zhang R, Chen X and Zhu J: LncRNA AK023391 promotes tumorigenesis
and invasion of gastric cancer through activation of the PI3K/Akt
signaling pathway. J Exp Clin Cancer Res. 36(194)2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Yang Y, Junjie P, Sanjun C and Ma Y: Long
non-coding RNAs in colorectal cancer: Progression and future
directions. J Cancer. 8:3212–3225. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Jiang X, Li Q, Zhang S, Song C and Zheng
P: Long noncoding RNA GIHCG induces cancer progression and
chemoresistance and indicates poor prognosis in colorectal cancer.
Onco Targets Ther. 12:1059–1070. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Shi L, Hong X, Ba L, He X, Xiong Y, Ding
Q, Yang S and Peng G: Long non-coding RNA ZNFX1-AS1 promotes the
tumor progression and metastasis of colorectal cancer by acting as
a competing endogenous RNA of miR-144 to regulate EZH2 expression.
Cell Death Dis. 10(150)2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yu Y, Nangia-Makker P, Farhana L and
Majumdar APN: A novel mechanism of lncRNA and miRNA interaction:
CCAT2 regulates miR-145 expression by suppressing its maturation
process in colon cancer cells. Mol Cancer. 16(155)2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhang J, Jiang Y, Zhu J, Wu T, Ma J, Du C,
Chen S, Li T, Han J and Wang X: Overexpression of long non-coding
RNA colon cancer-associated transcript 2 is associated with
advanced tumor progression and poor prognosis in patients with
colorectal cancer. Oncol Lett. 14:6907–6914. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zhu L, Tu H, Liang Y and Tang D: MiR-218
produces anti-tumor effects on cervical cancer cells in vitro.
World J Surg Oncol. 16(204)2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Xuan C, Jin M, Gao Y, Xu S, Wang L, Wang
Y, Han R and An Q: miR-218 suppresses the proliferation of
osteosarcoma through downregulation of E2F2. Oncol Lett.
17:571–577. 2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhang G, He X, Ren C, Lin J and Wang Q:
Long noncoding RNA PCA3 regulates prostate cancer through sponging
miR-218-5p and modulating high mobility group box 1. J Cell
Physiol. 234:13097–13109. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Warren RS, Yuan H, Matli MR, Gillett NA
and Ferrara N: Regulation by vascular endothelial growth factor of
human colon cancer tumorigenesis in a mouse model of experimental
liver metastasis. J Clin Invest. 95:1789–1797. 1995.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Zhang D, Qiu X, Li J, Zheng S, Li L and
Zhao H: TGF-β secreted by tumor-associated macrophages promotes
proliferation and invasion of colorectal cancer via miR-34a-VEGF
axis. Cell Cycle. 17:2766–2778. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Schiffmann LM, Fritsch M, Gebauer F,
Günther SD, Stair NR, Seeger JM, Thangarajah F, Dieplinger G,
Bludau M, Alakus H, et al: Tumour-infiltrating neutrophils
counteract anti-VEGF therapy in metastatic colorectal cancer. Br J
Cancer. 120:69–78. 2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Yang T, Zhai H, Yan R, Zhou Z, Gao L and
Wang L: lncRNA CCAT1 promotes cell proliferation, migration, and
invasion by down-regulation of miR-143 in FTC-133 thyroid carcinoma
cell line. Braz J Med Biol Res. 51(e7046)2018.PubMed/NCBI View Article : Google Scholar
|