Introduction
Colorectal cancer (CRC) is a common clinical cancer
that is associated with high morbidity and mortality (1). Gene mutations, inflammatory condition
and imbalances in the microbiome are associated with increased risk
of CRC development and progression (2-4).
Despite advances in radiotherapy, chemotherapy and surgery, severe
side effects compromise the CRC patient health (5,6). Thus,
there is an urgent interest and demand for identifying novel
therapeutic targets and prognosis predictors for CRC.
Long non-coding RNAs (lncRNAs) comprise a class of
transcripts that are >200 nucleotides in length but without any
protein-coding ability. Previous studies indicated that lncRNAs are
involved in various physiological and pathological processes
(7,8). lncRNAs exert their biological functions
by binding to DNA, RNA and proteins, thereby regulating protein
expression at the transcriptional or post-transcriptional level
(9,10). Numerous lncRNAs are dysregulated in
different types of cancer and are correlated with tumor cell
proliferation, apoptosis, migration and stem-like properties
(11-13).
lncRNA activated by transforming growth factor-β (ATB) has been
identified as a novel transforming growth factor-β (TGF-β)-induced
lncRNA that stimulates cancer cell proliferation, metastasis,
epithelial-mesenchymal transition (EMT) and angiogenesis in lung,
bladder, breast and gastric cancer (14-17).
In CRC, ATB promoted cancer cell proliferation and was found to be
a predictor of poor prognosis in patients (18,19).
However, the regulatory role of ATB in the maintenance of CRC cell
stemness remains unclear.
The present study aimed to determine the functional
role of ATB in maintaining the stemness of CRC cells. Colony
formation assays, sphere formation assays and xenograft models were
used to determine the functional role of ATB in maintaining CRC
stemness. The mechanisms underlying the effects of ATB were
additionally investigated by reverse transcription-quantitative PCR
(RT-qPCR), western blotting, and immunohistochemical staining. The
present study provided evidence for understanding ATB and
identifying a potential therapeutic target for CRC.
Materials and methods
Cell culture and treatment
The human colon cancer cell lines HCT116 and HT29
were purchased from the American Type Culture Collection and
cultured in DMEM containing 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.). Lentivirus-based short hairpin RNAs (shRNAs)
targeting ATB (shATB-1 and shATB-2, two different custom shRNA
sequences targeting ATB) and shRNA-negative control (shNC) were
obtained from Shanghai GenePharma Co., Ltd. HCT116 and HT29 cells
were seeded into 6 well plates (2x105 cells per well).
After culture for 24 h, 4x106 lentiviral particles were
added into the well and puromycin (2 mg/ml) was added into the
medium at 24 h post infection. The stably infected cells were
collected for further experiments. A specific activator of the
β-catenin pathway, CP21R7 (CP21; cat. no. S7954), was purchased
from Selleck Chemicals and used to treat HCT116 and HT29 cells for
48 h at 37˚C with a concentration of 2 mM. The equivalent volume of
DMSO was added as a control.
Colony formation assay
A total of 2x103 HCT116 or HT29 cells
were seeded into each well of a six-well plate containing 2 ml DMEM
supplemented with 10% FBS. A total of 14 days later, plates were
collected and washed with PBS three times at room temperature.
Next, 4% paraformaldehyde was added to fix the cells for 15 min at
room temperature. After washing cells with PBS three times at room
temperature, the cells were stained with 2% crystal violet
(Beyotime Institute of Biotechnology) for 15 min at room
temperature. The colonies in each well were counted under a light
microscope (Nikon Corporation) by two experimenters and analyzed.
Three independent experiments were performed in each group.
Sphere formation assay
A total of 2x104 HCT116 or HT29 cells
were seeded into each well of a six-well plate containing 2 ml
serum-free DMEM. Five days later, the images of spheres were
captured using an inverted microscope (x40; Nikon Corporation). The
number of spheres in each well was counted after dilution for 100
times with the inverted microscope (x40; Nikon Corporation) by two
experimenters and analyzed. Three independent experiments were
performed in each group.
RNA extraction and RT-qPCR
Total RNA was extracted using TRIzol™ reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Reverse transcription (RT) was
performed on isolated total RNA using a PrimeScript One Step RT-PCR
kit (cat. no. RR047A; Takara Bio, Inc.), and qPCR was performed
using a SYBR green Real Time PCR kit (Cat. No. RR085A; Takara Bio,
Inc.), according to the manufacturer's protocol. The following
thermocycling conditions were used for the qPCR: Initial
denaturation at 95˚C for 2 min; 40 cycles of 94˚C for 15 sec, 58˚C
for 30 sec and 70˚C for 30 sec; extension at 72˚C for 5 min. GAPDH
was used as an internal control. The primers were supplied as
follow: ATB-F, 5'-TCTGGCTGAGGCTGGTTGAC-3'; ATB-R,
5'-ATCTCTGGGTGCTGGTGAAGG-3'; GAPDH-F, 5'-TCAAGGCTGAGAACGGGAAG-3';
GAPDH-R, 5'-TCGCCCCACTTGATTTTGGA-3'. The relative gene expression
was calculated using the 2-ΔΔCq
method (20).
Western blotting
Total protein from HCT116 and HT29 cells was
extracted using RIPA buffer (Beyotime Institute of Biotechnology)
containing 1% protease inhibitor cocktail (EMD Millipore). The
protein concentration was determined by BCA kit (Beyotime Institute
of Biotechnology). Subsequently, 20 µg of total protein was loaded
in each lane of an SDS-PAGE (10% for β-catenin, LEF1 and GSK-3β or
12% for GAPDH). Following electrophoresis, proteins were
transferred onto PVDF membranes (EMD Millipore). Membranes were
blocked with 5% milk in TBS-Tween-20 buffer for 2 h at room
temperature. The membranes were then incubated with primary
antibodies supplied by Cell Signaling Technology, Inc. against
β-catenin (1:1,000; cat. no. 8480), lymphoid enhancer-binding
factor 1 (LEF1; 1:80; cat. no. 76010) and glycogen synthase
kinase-3β (GSK-3β; 1:1,000; cat. no. 12456) overnight at 4˚C.
Following incubation with HRP conjugated secondary antibody
(1:10,000; cat. nos. ZDR 5306 and ZDR 5307; Origene Technologies,
Inc.) at 22-25˚C for 2 h, an enhanced chemiluminescence kit (EMD
Millipore) was used to detect specific bands. The density of each
band was analyzed using ImageJ software (version: 1.52, National
Institutes of Health).
Animal studies
Animal studies were performed in accordance with the
institutional guidelines of Soochow University. Female BALB/c nude
mice (age, 6 weeks; weight, 18-20 g; 5 mice in each group; humidity
40-60%) were purchased from the Model Animal Research Center of
Nanjing University and allowed to acclimate for 1 week before use.
All of the mice were housed at 25˚C under a 12 h light/dark cycle
with ad libitum access to food and water. For xenograft
models, 5x106 HT29 cells were injected into the right
flank via the subcutaneous vein of mice. At 10 days post-injection,
the tumor length and width was measured using a vernier caliper
every 5 days. The tumor volume was calculated as tumor length x
tumor width2 x0.52(21).
At 25 days post-injection, the mice were sacrificed and tumors were
collected, weighed and used for histopathological studies.
Immunohistochemical staining and
RNA-fluorescence in situ hybridization
The tumor tissues were fixed with 4%
paraformaldehyde at 22-25˚C for 48 h and embedded in paraffin.
After heating at 65˚C for 2 h, the paraffin-embedded sections (5
µm) of colon tumor tissues were dewaxed with xylene and gradient
alcohol and then subjected to antigen retrieval with citrate buffer
(OriGene Technologies, Inc.). Following blocking with 5% normal
goat serum (Origene, Technologies, Inc.) at 37˚C for 15 min, the
sections were incubated with primary antibody against β-catenin
(1:200; cat. no. 8480, Cell Signaling Technology, Inc.) at 4˚C
overnight. After immunoperoxidase staining with a
Streptavidin-Peroxidase kit (OriGene Technologies, Inc.),
3,3'-diamino-benzidine (Fuzhou Maixin Biotechnology Development
Co., Ltd.) was used to detect the target protein. The cell nucleus
was stained with hematoxylin at 22-25˚C for 3 min (Beyotime
Institute of Biotechnology). For RNA-fluorescence in situ
hybridization, a custom probe for the specific detection of ATB was
synthesized by Guangzhou Ribobio Co., Ltd. The process of sample
preparation and hybridization was followed by the use of a
fluorescence in situ hybridization kit (cat. no. C10910;
Guangzhou Ribobio Co., Ltd). Images of sections were captured by
light microscope (x100; model BX51; Olympus Corporation). The
β-catenin positive cells and total cells in each image were counted
and the percent of positive cells were analyzed.
Statistical analysis
GraphPad Prism version 5.0 software (GraphPad
Software, Inc.) was used for statistical analysis. One-way ANOVA
followed with Dunnett's test was used to determine statistical
significance for more than two groups. All data are presented as
the mean ± SEM. P<0.05 indicated a statistically significant
difference.
Results
ATB knockdown impairs CRC stemness
maintenance in vitro
To investigate the potential role of ATB in CRC,
lentivirus-based shRNAs targeting ATB were used to infect HCT116
and HT29 cells. Following puromycin selection, stably-infected
cells were collected for determination of ATB expression by
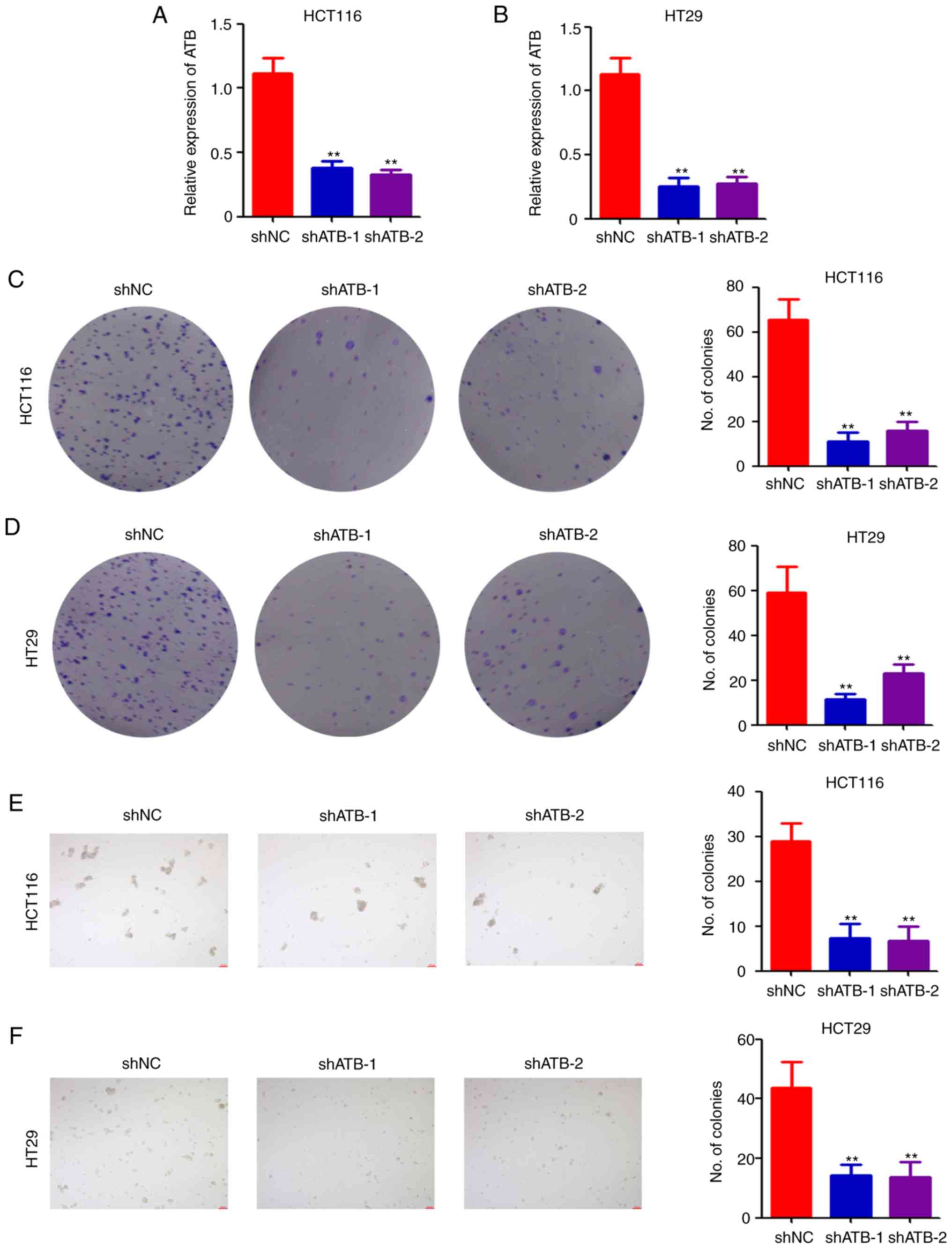
RT-qPCR. The results indicated that shATB significantly
downregulated ATB expression in both HCT116 (Fig. 1A) and HT29 (Fig. 1B) cells compared with shNC. The
stably infected cells were then analyzed using a colony formation
assay. Colony formation ability was significantly reduced in both
HCT116 (Fig. 1C) and HT29 (Fig. 1D) cells after ATB knockdown.
Furthermore, results of the sphere formation assay indicated that
ATB knockdown significantly impaired sphere formation in HCT116
(Fig. 1E) and HT29 (Fig. 1F) cells. Collectively, the above
results demonstrated the positive role of ATB in CRC stemness
maintenance.
ATB promotes colon tumor growth in
vivo
Based on the in vitro study results, an in
vivo study was conducted to determine the functional role of
ATB in colon tumor growth. HT29 cells that were stably transfected
with shNC or shATB were injected into the right subcutaneous vein
to establish a xenograft model. HT29 tumor tissues were collected
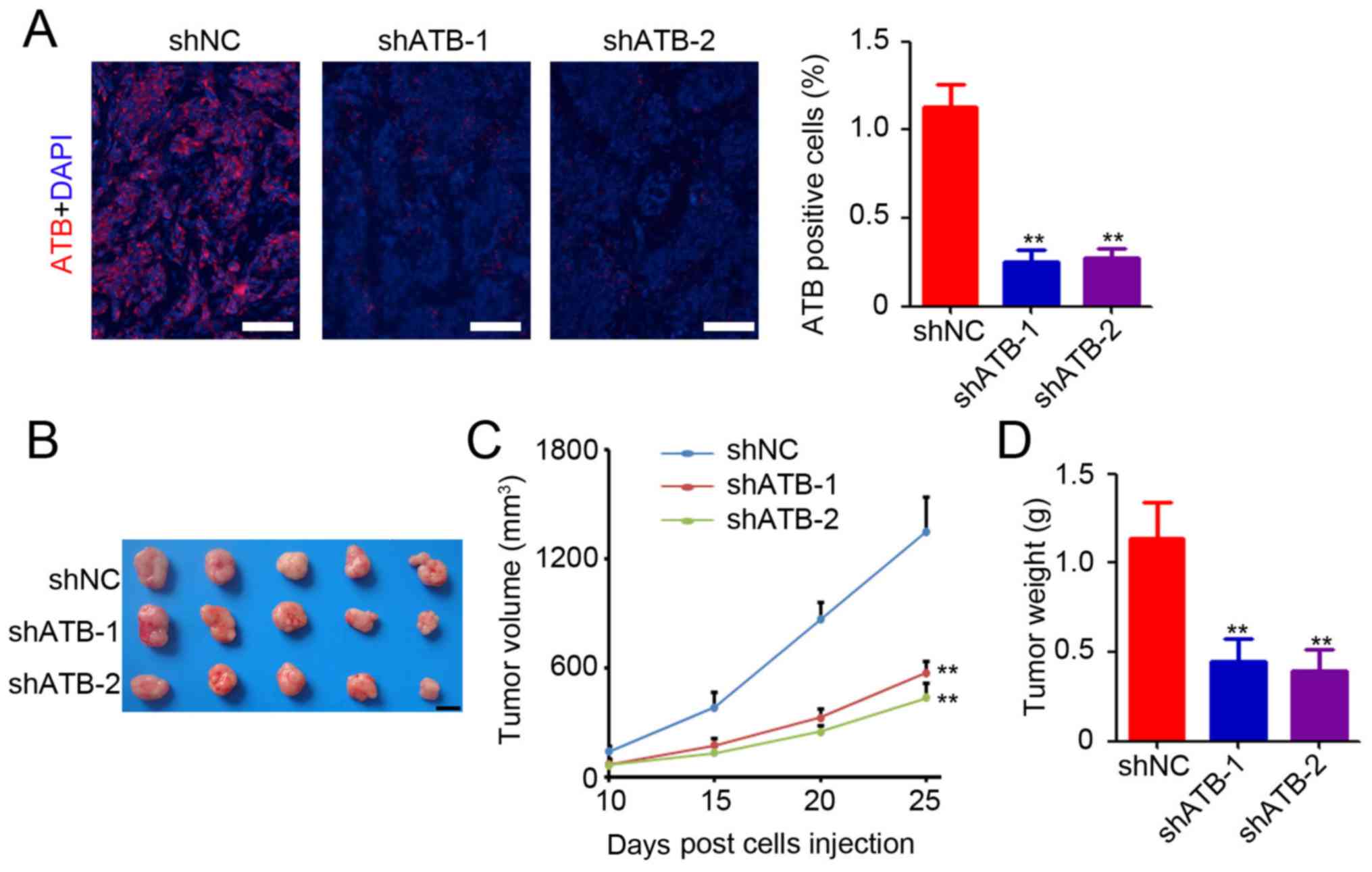
for determination of ATB expression by RNA-fluorescence in situ
hybridization. A small number of ATB-positive cells was observed in
the colon tumor tissues after ATB knockdown (Fig. 2A). Further statistical analysis
confirmed significant downregulation of ATB expression in
shATB-infected HT29 tumors (Fig.
2A). The results suggested that ATB knockdown in HT29 cells
significantly inhibited tumor growth (Fig. 2B), as evidenced by a mean reduction
of 76% in tumor volume (shATB-1 and sh-ATB-2 vs. shNC; Fig. 2C) and 68% decrease in tumor weight
(shATB-1 and sh-ATB-2 vs. shNC; Fig.
2D). The above results demonstrated the oncogenic role of ATB
in CRC.
ATB inhibits the expression and
activation of β-catenin
To determine the mechanisms underlying stemness
maintenance in CRC by ATB, stably-transfected HCT116 and HT29 cells
were collected for further molecular determination. Expression
levels of the downstream targets of the Wnt signaling pathway were
measured, and the results indicated that ATB knockdown
significantly inhibited the expression of G1/S-specific
cyclin-D1, transcription factor-1, c-Myc, matrix metalloproteinase
7, and twist-related protein (Twist) both in HCT116 and HT29 cells
compared with shNC (Fig. 3A).
Western blotting was performed to determine the expression levels
of β-catenin, LEF1, and GSK-3β in CRC cells. Significant
downregulation of β-catenin expression was observed in both
ATB-knockdown HCT116 and HT29 cells compared with shNC (Fig. 3B). However, ATB knockdown did not
significantly affect the expression of LEF1 and GSK-3β in CRC cells
(Fig. 3B). Reduced levels of
β-catenin-positive cells were found in shATB-infected HT29 tumor
tissues compared with shNC-infected HT29 tumor tissues (Fig. 3C). Taken together, the results
demonstrated the regulatory role of ATB on β-catenin expression in
CRC.
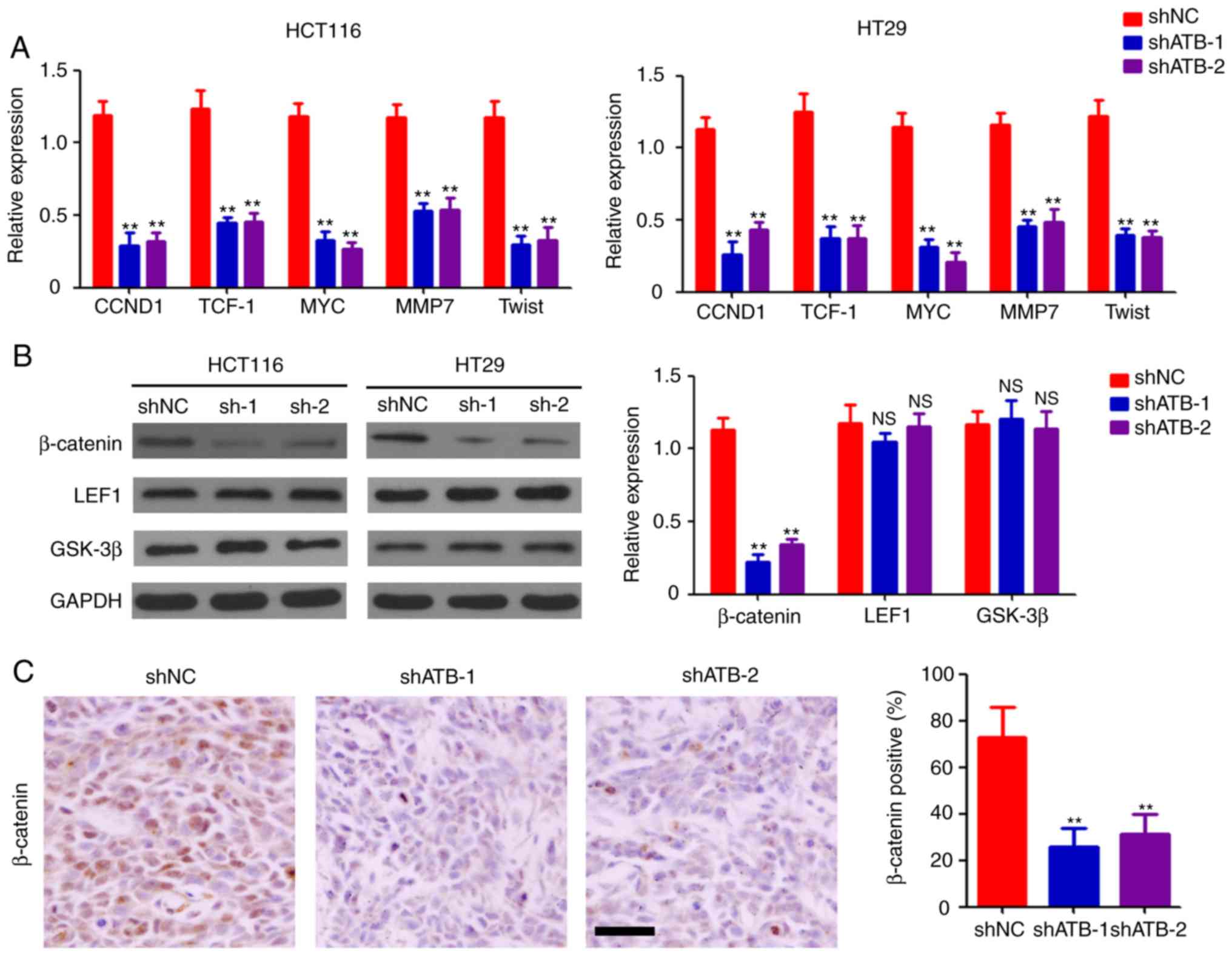 | Figure 3ATB regulates the activation of the
Wnt/β-catenin pathway. (A) Reverse transcription-quantitative PCR
analysis of CCND1, TCF-1, MYC, MMP7 and Twist expression in HCT116
and HT29 cells stably transfected with shNC, shATB-1 and shATB-2.
n=3. (B) Western blot analysis of β-catenin, LEF1 and GSK-3β
expression in HCT116 and HT29 cells stably infected with shNC,
shATB-1 and shATB-2. GAPDH was used as a loading control. The
relative expression of each band was analyzed. n=3. (C)
Immunohistochemical staining of β-catenin expression in HT29 tumors
(scale bar=100 µm). The percentage of β-catenin-positive cells was
analyzed. n=3. **P<0.01 vs. shNC. ATB, long
non-coding RNA activated by transforming growth factor-β; sh, short
hairpin RNA; NC, negative control; CCND1, cyclin-D1; TCF-1,
transcription factor-1; MMP7, matrix metalloproteinase 7; Twist,
twist-related protein; LEF1, lymphoid enhancer-binding factor 1;
GSK-3β, glycogen synthase kinase-3β; NS, not significant. |
β-catenin plays a crucial role in
ATB-mediated stemness maintenance in CRC
To determine whether β-catenin was important for
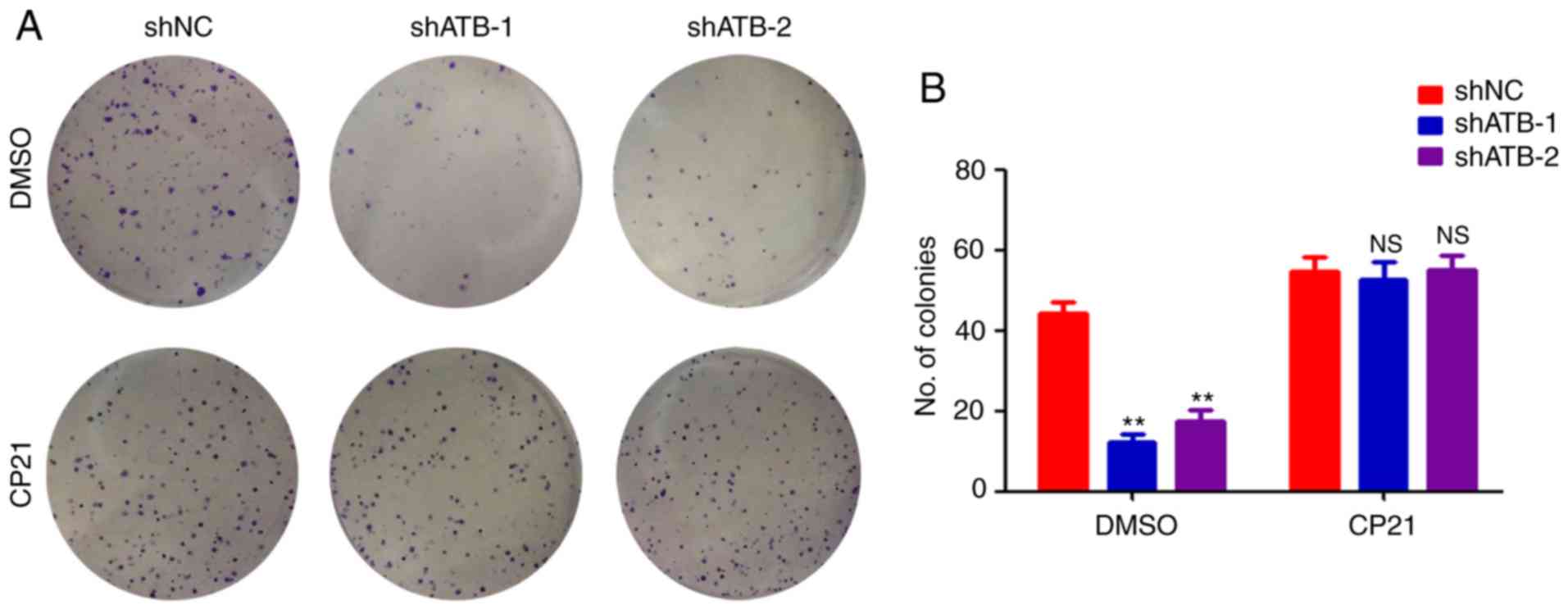
regulating stemness in CRC by ATB, HT29 cells were treated with
CP21, a specific activator of the β-catenin pathway. ATB knockdown
significantly inhibited the colony formation ability of HT29 cells
(Fig. 4A and B). However, CP21 treatment inhibited the
decrease of colony formation mediated by shATB in HT29 cells
(Fig. 4A and B). Collectively, the results confirmed that
β-catenin was required for ATB-mediated maintenance of stemness in
CRC.
Discussion
Considering their extensive biological functions,
lncRNAs are involved in a wide range of physiological and
pathological processes (22). In the
present study, ATB knockdown impaired colony and sphere formation
of CRC cells and significantly inhibited colon tumor growth.
Further results suggested that ATB regulated the transcriptional
activity of the β-catenin pathway by inhibiting β-catenin
expression. In addition, the results confirmed that β-catenin was
necessary for ATB-mediated regulation of stemness in CRC.
ATB is dysregulated in various types of cancer,
including CRC (19), lung cancer
(23), and liver cancer (24). ATB acts as an oncogene, and its
dysregulation is commonly associated with poor prognosis of
patients (25). In CRC, ATB is
upregulated in colon cancer tissues and correlated with clinical
cancer stage (19). In addition, ATB
is highly expressed in lung cancer tissues and associated with
tumor size and metastasis (23). A
functional study indicated that ATB promoted proliferation,
migration and invasion in osteosarcoma cells (26). Knockdown of ATB in endometrial cancer
cells was found to impair cell viability by inducing
caspase-3-related tumor apoptosis and G1/S arrest
(27). In addition, ATB was
responsible for the proliferation and apoptosis of CRC cells
(18). The results demonstrated that
ATB knockdown impaired stemness maintenance of CRC cells, indicated
by the inhibition of colony formation and sphere formation, and
colon tumor growth inhibition. The findings expand the current
understanding of ATB function.
Recent studies demonstrated that ATB acts as a
competing RNA by binding with microRNA (miR)-494 in lung cancer
cells (28), miR-141-3p in gastric
cancer cells (29), miR-590-5p in
melanoma (30), and miR-200 family
members in diverse types of cancer (18,26,31,32). ATB regulates
transcriptional coactivator YAP1 (YAP1), NF-κB, p38
mitogen-activated protein kinase (MAPK), Twist, and TGF-β in lung
cancer, glioma, breast cancer and malignant melanoma (23,30,33,34).
In hepatic fibrosis, ATB knockdown downregulated β-catenin
expression by upregulating the expression of endogenous miR-200a
(35). In the present study, the
results suggested that ATB regulated the transcriptional activity
of the β-catenin pathway by inhibiting β-catenin expression.
Further results confirmed the role of β-catenin during ATB-mediated
regulation of stemness in CRC. miR-200a, which is involved in
regulating CRC cell proliferation and β-catenin expression
(36,37), could serve as the direct target of
ATB in regulating stemness maintenance. However, further studies
are required to verify the above results.
The present study demonstrated the role of ATB in
maintaining stemness of CRC by regulating β-catenin expression.
However, further studies are required to investigate the direct
target of ATB for regulating β-catenin expression in CRC.
Collectively, the results indicated that ATB is a promising
therapeutic target for CRC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XY and HT were involved in the acquisition of the
data and drafting the manuscript. CW, WC and FH were involved in
the analysis and interpretation of the data. HQ was involved in the
conception and design of the present study. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This animal study was approved by the Ethics
Committee of Soochow University and complied with their
guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Dai L, Liu Y, Cheng L, Wang H, Lin Y, Shi
G, Dong Z, Li J, Fan P, Wang Q, et al: SARI attenuates colon
inflammation by promoting STAT1 degradation in intestinal
epithelial cells. Mucosal Immunol. 12:1130–1140. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Humphries A and Wright NA: Colonic crypt
organization and tumorigenesis. Nat Rev Cancer. 8:415–424.
2008.PubMed/NCBI View
Article : Google Scholar
|
|
5
|
Brungs D, Aghmesheh M, de Souza P, Carolan
M, Clingan P, Rose J and Ranson M: Safety and efficacy of
oxaliplatin doublet adjuvant chemotherapy in elderly patients with
stage III colon cancer. Clin Colorectal Cancer. 17:e549–e555.
2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Li J, Li XL, Yuan Y and Zhang SZ: Disputes
and exploration of neoadjuvant and adjuvant therapy for colon
cancer. Zhonghua Wei Chang Wai Ke Za Zhi. 22:329–335. 2019.(In
Chinese). PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zheng J, Mao Y, Dong P, Huang Z and Yu F:
Long noncoding RNA HOTTIP mediates SRF expression through sponging
miR-150 in hepatic stellate cells. J Cell Mol Med. 23:1572–1580.
2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wu S, Bono J and Tao YX: Long noncoding
RNA (lncRNA): A target in neuropathic pain. Expert Opin Ther
Targets. 23:15–20. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Liu H, Li C, Yang J, Sun Y, Zhang S, Yang
J, Yang L, Wang Y and Jiao B: Long noncoding RNA
CASC9/miR-519d/STAT3 positive feedback loop facilitate the glioma
tumourigenesis. J Cell Mol Med. 22:6338–6344. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yuan SX, Zhang J, Xu QG, Yang Y and Zhou
WP: Long noncoding RNA, the methylation of genomic elements and
their emerging crosstalk in hepatocellular carcinoma. Cancer Lett.
379:239–244. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Dai L, Li J, Dong Z, Liu Y, Chen Y, Chen
N, Cheng L, Fang C, Wang H, Ji Y, et al: Temporal expression and
functional analysis of long non-coding RNAs in colorectal cancer
initiation. J Cell Mol Med. 23:4127–4138. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wu Q, Guo L, Jiang F, Li L, Li Z and Chen
F: Analysis of the miRNA-mRNA-lncRNA networks in ER+ and ER-breast
cancer cell lines. J Cell Mol Med. 19:2874–2887. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lorenzi L, Avila Cobos F, Decock A,
Everaert C, Helsmoortel H, Lefever S, Verboom K, Volders PJ,
Speleman F, Vandesompele J and Mestdagh P: Long noncoding RNA
expression profiling in cancer: Challenges and opportunities. Genes
Chromosomes Cancer. 58:191–199. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chen Y, Wei G, Xia H, Tang Q and Bi F:
Long noncoding RNA-ATB promotes cell proliferation, migration and
invasion in gastric cancer. Mol Med Rep. 17:1940–1946.
2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Xu S, Yi XM, Tang CP, Ge JP, Zhang ZY and
Zhou WQ: Long non-coding RNA ATB promotes growth and
epithelial-mesenchymal transition and predicts poor prognosis in
human prostate carcinoma. Oncol Rep. 36:10–22. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhai X and Xu W: Long noncoding RNA ATB
promotes proliferation, migration, and invasion in bladder cancer
by suppressing MicroRNA-126. Oncol Res. 26:1063–1072.
2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang Y, Li J, Jia S, Wang Y, Kang Y and
Zhang W: Down-regulation of lncRNA-ATB inhibits
epithelial-mesenchymal transition of breast cancer cells by
increasing miR-141-3p expression. Biochem Cell Biol. 97:193–200.
2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Gao Z, Zhou H, Wang Y, Chen J and Ou Y:
Regulatory effects of lncRNA ATB targeting miR-200c on
proliferation and apoptosis of colorectal cancer cells. J Cell
Biochem. 121:332–343. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yue B, Qiu S, Zhao S, Liu C, Zhang D, Yu
F, Peng Z and Yan D: LncRNA-ATB mediated E-cadherin repression
promotes the progression of colon cancer and predicts poor
prognosis. J Gastroenterol Hepatol. 31:595–603. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Cheng L, Yang Q, Li C, Dai L, Yang Y, Wang
Q, Ding Y, Zhang J, Liu L, Zhang S, et al: DDA1, a novel oncogene,
promotes lung cancer progression through regulation of cell cycle.
J Cell Mol Med. 21:1532–1544. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Li CH and Chen Y: Insight into the role of
long noncoding RNA in cancer development and progression. Int Rev
Cell Mol Biol. 326:33–65. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wei L, Wu T, He P, Zhang JL and Wu W:
LncRNA ATB promotes the proliferation and metastasis of lung cancer
via activation of the p38 signaling pathway. Oncol Lett.
16:3907–3912. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lee YR, Kim G, Tak WY, Jang SY, Kweon YO,
Park JG, Lee HW, Han YS, Chun JM, Park SY and Hur K: Circulating
exosomal noncoding RNAs as prognostic biomarkers in human
hepatocellular carcinoma. Int J Cancer. 144:1444–1452.
2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Li J, Li Z, Zheng W, Li X, Wang Z, Cui Y
and Jiang X: LncRNA-ATB: An indispensable cancer-related long
noncoding RNA. Cell Prolif. 50:2017.doi: 10.1111/cpr.12381.
PubMed/NCBI View Article : Google Scholar
|
|
26
|
Han F, Wang C, Wang Y and Zhang L: Long
noncoding RNA ATB promotes osteosarcoma cell proliferation,
migration and invasion by suppressing miR-200s. Am J Cancer Res.
7:770–783. 2017.PubMed/NCBI
|
|
27
|
Zheng X, Liu M, Song Y and Feng C: Long
noncoding RNA-ATB impairs the function of tumor suppressor
miR-126-mediated signals in endometrial cancer for tumor growth and
metastasis. Cancer Biother Radiopharm. 34:47–55. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Cao Y, Luo X, Ding X, Cui S and Guo C:
LncRNA ATB promotes proliferation and metastasis in A549 cells by
down-regulation of microRNA-494. J Cell Biochem. 119:6935–6942.
2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Lei K, Liang X, Gao Y, Xu B, Xu Y, Li Y,
Tao Y, Shi W and Liu J: Lnc-ATB contributes to gastric cancer
growth through a MiR-141-3p/TGFβ2 feedback loop. Biochem Biophys
Res Commun. 484:514–521. 2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Mou K, Liu B, Ding M, Mu X, Han D, Zhou Y
and Wang LJ: lncRNA-ATB functions as a competing endogenous RNA to
promote YAP1 by sponging miR-590-5p in malignant melanoma. Int J
Oncol. 53:1094–1104. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li Z, Wu X, Gu L, Shen Q, Luo W, Deng C,
Zhou Q, Chen X, Li Y, Lim Z, et al: Long non-coding RNA ATB
promotes malignancy of esophageal squamous cell carcinoma by
regulating miR-200b/Kindlin-2 axis. Cell Death Dis.
8(e2888)2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
He T, Zhou H, Li C, Chen Y, Chen X, Li C,
Mao J, Lyu J and Meng QH: Methylglyoxal suppresses human colon
cancer cell lines and tumor growth in a mouse model by impairing
glycolytic metabolism of cancer cells associated with
down-regulation of c-Myc expression. Cancer Biol Ther. 17:955–965.
2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhou Y, Zheng X, Lu J, Chen W, Li X and
Zhao L: Ginsenoside 20(S)-Rg3 inhibits the warburg effect via
modulating DNMT3A/MiR-532-3p/HK2 pathway in ovarian cancer cells.
Cell Physiol Biochem. 45:2548–2559. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Tang F, Wang H, Chen E, Bian E, Xu Y, Ji
X, Yang Z, Hua X, Zhang Y and Zhao B: LncRNA-ATB promotes
TGF-β-induced glioma cells invasion through NF-κB and P38/MAPK
pathway. J Cell Physiol. 234:23302–23314. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Fu N, Zhao SX, Kong LB, Du JH, Ren WG, Han
F, Zhang QS, Li WC, Cui P, Wang RQ, et al:
LncRNA-ATB/microRNA-200a/β-catenin regulatory axis involved in the
progression of HCV-related hepatic fibrosis. Gene. 618:1–7.
2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Santasusagna S, Moreno I, Navarro A,
Martinez Rodenas F, Hernández R, Castellano JJ, Muñoz C and Monzo
M: Prognostic impact of miR-200 family members in plasma and
exosomes from tumor-draining versus peripheral veins of colon
cancer patients. Oncology. 95:309–318. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Yang W, Ning N and Jin X: The lncRNA H19
promotes cell proliferation by competitively binding to miR-200a
and derepressing β-catenin expression in colorectal cancer. Biomed
Res Int. 2017(2767484)2017.PubMed/NCBI View Article : Google Scholar
|