Introduction
Chronic obstructive pulmonary disease (COPD) is
characterized by persistent airflow limitation, which is usually
progressive, and is associated with an enhanced chronic airway and
pleural inflammatory response to noxious particles or gases
(1,2). COPD is on course to become a global
health challenge in the near future. It is currently the fourth
leading cause of death worldwide, but is soon expected to become
the third (2).
The most important risk factor associated with COPD
is cigarette smoking (1). Tobacco
smoke and air pollutants are sources of oxidative stress, which is
induced by an increased burden of inhaled oxidants and elevated
levels of reactive oxygen species (ROS) released from inflammatory
cells (3). Increases in levels of
ROS, either directly or via the formation of lipid peroxidation
products, contribute to an enhanced inflammatory response in COPD
(3).
Corticosteroids have an anti-inflammatory effect in
chronic inflammatory diseases such as asthma and diffuse
panbronchiolitis (4). However, in
COPD patients, the anti-inflammatory effects of glucocorticoids are
limited (4). High-dose inhaled and
oral glucocorticoids have a limited effect in reducing the
production and release of inflammatory cells, cytokines and
proteases, including tumor necrosis factor-α (TNF-α), interleukin-8
(IL-8) and matrix metallopeptidase 9 (MMP-9) in COPD patients
(5,6). This suggests corticosteroid resistance
in these patients. Dexamethasone (Dex) is a long-acting
glucocorticoid that has routine application in clinical and
experimental work (7-9).
Histone acetyltransferases (HATs) and histone
deacetylases (HDACs) are families of enzymes that regulate and
affect inflammatory gene expression (7,8). HATs
can open chromatin structure by acetylation of the molecular
modification protein to ensure the transcription factor and RNA
polymerase bind to DNA for prompt gene transcription. However,
HDACs ensure that the chromatin conformation remains closed, and
thus block gene transcription (10,11).
Increasing evidence has shown that the acetylation-deacetylation
imbalance may be the determining factor in corticosteroid
resistance in COPD and, consequently, in the inability of
conventional treatments to prevent inflammation-induced lung
destruction (12,13). HDAC2, a member of the class I HDAC
family, has been shown to play a role in the regulation of cell
inflammatory responses (11,14). It has been reported that the impact
of oxidative stress on chromatin regulation by a reduction in the
activity of HDAC-2 leads to corticosteroid resistance in COPD
(3,15).
PI3Ks generate lipid second messengers that control
an array of intracellular signaling pathways, which have important
roles in inflammation (16).
Oxidative stress not only reduces HDAC-2 activity and, thereby,
glucocorticoid immunosuppression, but also activates the PI3K/Akt
pathway (17,18). PI3Ks are activated by cell-surface
receptors, such as receptor tyrosine kinases and G-protein-coupled
receptors, and serve to initiate intracellular signaling cascades
through the generation of the lipid secondary messenger
phosphatidylinositol-3,4,5-triphosphate (PIP3). PIP3 serves as a
docking site for the pleckstrin homology domain of proteins such as
the serine-threonine kinase Akt (19,20). The
class I PI3K isoforms, PI3K-δ and PI3K-γ, are predominantly
expressed in leukocytes and play a central role in inflammatory
cell function (20). The
oxidant-mediated induction of PI3K signaling is reportedly mediated
primarily through PI3K-δ, but not PI3K-γ (20).
Theophylline is a long-established respiratory
treatment that was originally used as a bronchodilator. At
relatively high concentrations, theophylline directly inhibits
phosphodiesterases and antagonizes adenosine receptors (21). In recent years, a combination of
low-dose theophylline with glucocorticoids has been used to good
anti-inflammatory effect, and shown to effectively reduce
glucocorticoid resistance in patients with COPD (9,22).
Several studies have shown that low-dose theophylline reverses the
reduction in HDAC2 expression, seen in macrophages, monocytes and
epithelial cells challenged with oxidative stress, and in mice,
rats, and human patients with COPD, to restore glucocorticoid
sensitivity (7,13,22,23).
In the current study, a combined theophylline and
Dex treatment was used in an in vitro experiment to
determine whether this drug combination has the potential to reduce
CSE-induced inflammation through rescue of HDAC2 expression and
inhibition of the PI3K/Akt pathway.
Materials and methods
Cell culture and treatments
U937 cells (human monocytic cell line; Type Culture
Collection of the Chinese Academy of Sciences), as utilized
previously (24), are routinely used
to explore the mechanisms underlying inflammation in COPD. U937
cells were maintained in continuous cell culture at 37˚C and 5%
CO2 in RPMI-1640 medium (GE Healthcare Life Sciences)
supplemented with 10% fetal bovine serum (GE Healthcare Life
Sciences). U937 cells in the combined theophylline (Sigma-Aldrich;
Merck KGaA) and Dex (Hubei Qianjiang Pharmaceutical Co., Ltd.)
treatment group, or theophylline only treatment group, were
incubated with theophylline and Dex (10-6 M) or
theophylline (10-3 M) alone for 5 h prior to CSE
exposure. U937 cells of the IC87114 group were treated with IC87114
(1 µM) before CSE exposure. U937 cells in CSE treatment groups were
stimulated with CSE for 12 h.
Preparation of CSE
CSE was prepared using the methods developed by
Mercado et al (25). Ten
full-strength burning cigarettes without filters were continuously
pumped with a syringe. The smoke was slowly dissolved into 10 ml
PBS and the pH value was adjusted to 7.4. This CSE solution was
twice filtered through a 0.22 µm filter membrane and used within 2
h of preparation. With PBS as the blank control, the optical
density was measured at a wavelength of 320 nm, and converted to
provide a percentage CSE concentration.
MTT assay
U937 cells were plated in three 96-well plates at a
density of 1.5x104 cell/ml and co-incubated with
theophylline (10-3-10-6 mol/l; Sigma-Aldrich;
Merck KGaA) or CSE (4, 8, 16, or 32%). The compounds were added to
a final volume of 100 µmol, and the cells were cultured for 24, 48,
or 72 h at 37˚C. MTT solution (10 µl of 5 mg/ml) was added to each
well, and the cultures were incubated for an additional 4 h. After
removing the culture solution, 100 µl DMSO solution was added and
the plates were shaken at low speed for 10 min. The absorbance at
570 nm in each well was determined using a 96-well plate reader.
The proliferation of treated cells was compared to that of control
cells.
TNF-α assay
TNF-α concentrations in the supernatant were
evaluated by ELISA (96-well plates; R&D Systems Inc.; cat. no.
DY210) in accordance with the manufacturer's instructions.
IL-8 assay and IC50-Dex
calculation
U937 cells in the CSE group were treated with CSE
(8%) for 12 h, and then incubated with Dex for 45 min before
stimulation with TNF-α (10 ng/ml; PreproTech) at 37˚C overnight.
U937 cells in drug treatment groups were incubated with
theophylline (10-3 M) and Dex (10-6 M),
theophylline alone, or IC87114 (1 µM), respectively, for 5 h prior
to their treatment with CSE for 12 h and subsequent treatment with
Dex (10-11, 10-10, 10-9,
10-8, 10-7 and 10-6 M) 45 min
before stimulation with TNF-α (10 ng/ml). IL-8 concentrations in
the supernatant were evaluated by ELISA (96-well plates; R&D
Systems Inc.) according to the manufacturer's instructions. In the
current study, the IC50 of dexamethasone
(IC50-Dex) was used as a marker for corticosteroid
sensitivity, and was determined based on the inhibition of IL-8
release at different Dex concentrations (10-11,
10-10, 10-9, 10-8, 10-7
and 10-6 M) (24).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR, SYBR Green Master Mix, Life
Technologies)
Total RNA was extracted from treated cells using
TRIzol® (Invitrogen, Thermo Fisher Scientific, Inc.).
The quality and quantity of total RNA was analyzed using a
spectrophotometer. The RNA samples were reverse transcribed into
cDNA using a reverse transcription kit (Transcriptor first strand
cdna synthesis kit; Sigma-Aldrich; Merck KGaA) using the following
temperature protocol: 95˚C for 5 sec followed by 60˚C for 30 sec.
To determine the mRNA expression levels, the following primers were
used: HDAC2 forward, 5'-CAATCTAACTGTCAAAGGTCATGC-3' and reverse,
5'-TGAAGTCTGGTCCAAAATACTCAA-3'; and β-actin forward,
5'-ACACTGTGCCCATCTACG-3' and reverse, 5'-TGTCACGCACGATTTCC-3'. The
2-ΔΔCq method (26) was used for relative quantification.
Amplification was performed using an ABI 9700 real-time PCR
instrument (Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: Pre-denaturation at 95˚C for 30 sec and
denaturation at 95˚C for 5 sec, followed by 40 cycles of annealing
at 60˚C for 5 sec, elongation 60˚C for 30 sec and extension at 60˚C
for 30 sec. Suitable cycle numbers were determined in accordance
with the standard plasmid amplification curve in RT-qPCR using
Rotor-Gene Real-time Analysis Software version 6.1 (Qiagen, Inc.).
The mRNA values of HDAC2 transcripts were normalized to that of
β-actin (23).
Western blotting
Proteins were extracted from the treated cells using
RIPA buffer (Pierce; Thermo Fisher Scientific, Inc.). Protein
quantification was undertaken using bicinchoninic acid (BCA;
Pierce, Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. For HDAC2, PI3K, phosphorylated
(p)-Akt and Akt, 20 µg of each sample was analyzed on an SDS-PAGE
(10% gel), then transferred onto nitrocellulose membranes, which
were then blocked with 5% bovine serum albumin (Pierce; Thermo
Fisher Scientific, Inc.) dissolved in Tris-buffered saline with
0.1% Tween-20 at 4˚C overnight. The membranes were incubated for 12
h at 4˚C with the following rabbit anti-human primary polyclonal
antibodies (each, 1:2,000; R&D Systems, Inc.): catalogue
numbers: HDAC2 (cat. no. MAB7679), PI3K (cat. no. MAB2687), Akt
(cat. no. MAB2055) and p-Akt (cat. no. AF887). Thereafter, the
membranes were incubated with secondary antibody (horseradish
peroxidase-conjugated goat anti-rabbit IgG, (1:10,000; R&D
Systems, Inc.; cat. no. AF789), and the blots were visualized with
enhanced chemiluminescence (ECL luminescent substrate; Pierce;
Thermo Fisher Scientific, Inc.). Mouse anti-human GAPDH antibody
(1:10,000; R&D Systems; cat. nos. AF5781) was used as a protein
loading control. Desnitometry was performed using AlphaView
software (NatureGene Corporation).
Statistical analysis
All data were analyzed using SPSS software (version
17.0; SPSS, Inc.). All quantitative data are presented as the mean
± SD of three independent repeats. Multiple comparisons were
performed using one-way ANOVA followed by a Tukey's post hoc test.
In data with a non-normal distribution, variance was determined by
Kruskal-Wallis analysis, followed by Dunn's multiple comparison
tests. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of CSE or theophylline on the
growth of U937 cells
Cell-proliferation assays were conducted, using a
range of concentrations of CSE and theophylline, in order to
determine whether exposure to CSE or theophylline was cytotoxic.
Compared with the control group, the proliferation of
CSE-stimulated U937 cells (4 and 8% CSE) did not change
significantly over a period of 24, 48 and 72 h [CSE (4%),
0.689±0.055, 1.083±0.042 and 1.172±0.06; CSE (8%), 0.593±0.063,
0.928±0.061 and 0.996±0.046; control group, 0.716±0.051,
0.954±0.056 and 1.142±0.038]. However, concentrations of 16% and
32% CSE significantly slowed down cell growth [CSE (16%),
0.484±0.057, 0.518±0.044 and 0.528±0.075; CSE (32%), 0.266±0.032,
0.209±0.078 and 0.199±0.081; control group, 0.716±0.051,
0.954±0.056 and 1.142±0.038). There were no marked changes in cells
treated with theophylline (10-3-10-6 mol/l),
when compared with the control group, over a period of 24, 48 and
72 h [theophylline (10-6 mol/l) 0.702lline2, 0.9262lline
and 0.9942lline2; theophylline (10-5 mol/l),
0.668±0.036, 1.019±0.047 and 1.032±0.056; theophylline
(10-4 mol/l), 0.654±0.043, 0.947±0.068 and 1.066±0.072;
theophylline (10-3 mol/l), 0.672±0.066, 0.98±0.048 and
1.083±0.035; control group, 0.716±0.051, 0.954±0.056 and
1.142±0.038). Table I.
 | Table IAbsorbance values of U937 cells
exposed to different concentrations of CSE and theophylline (mean ±
SD; n=6). |
Table I
Absorbance values of U937 cells
exposed to different concentrations of CSE and theophylline (mean ±
SD; n=6).
| Groups | 24 h | 48 h | 72 h |
|---|
| Comtrol group | 0.716±0.051 | 0.954±0.056 | 1.142±0.038 |
| CSE (4%) | 0.689±0.055 | 1.083±0.042 | 1.172±0.06 |
| CSE (8%) | 0.593±0.063 | 0.928±0.061 | 0.996±0.046 |
| CSE (16%) |
0.484±0.057a |
0.518±0.044a |
0.528±0.075a |
| CSE (32%) |
0.266±0.032a |
0.209±0.078a |
0.199±0.081a |
| Theophylline
(10-6 mol/l) | 0.702±0.045 | 0.926±0.059 | 0.994±0.0.35 |
| Theophylline
(10-5 mol/l) | 0.668±0.036 | 1.019±0.047 | 1.032±0.056 |
| Theophylline
(10-4 mol/l) | 0.654±0.043 | 0.947±0.068 | 1.066±0.072 |
| Theophylline
(10-3 mol/l) | 0.672±0.066 | 0.98±0.048 | 1.083±0.035 |
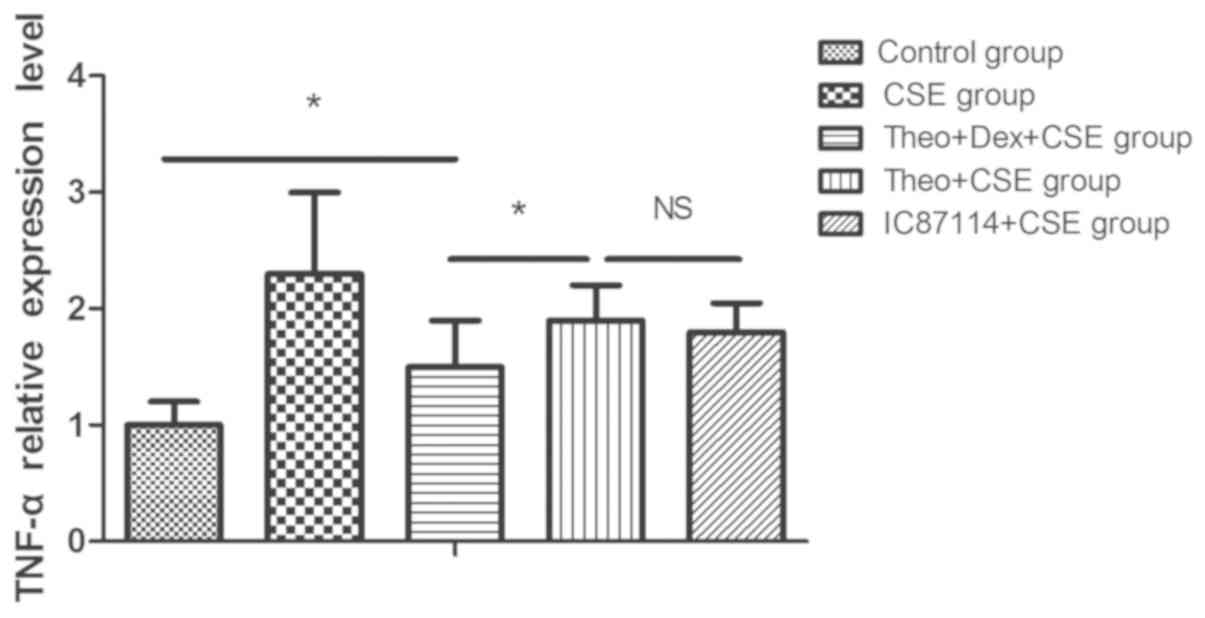
Theophylline-Dex pretreatment inhibits
TNF-α release in CSE-exposed U937 cells
U937 cells were incubated with theophylline and Dex,
theophylline alone, or IC87114 (1 µM) alone, respectively, for 5 h
prior to 12-h incubation with CSE. CSE-stimulation of U937 cells
markedly increased the level of TNFα released into the supernatant
when compared with the control group. Pretreatment with
theophylline significantly reduced the level of TNF-α released by
U937 cells exposed to CSE when compared with the CSE group.
However, a combination of theophylline and Dex pretreatment was
significantly more effective in reducing TNFα release than
theophylline alone. As a positive control, pretreatment with
IC87114 was used and shown to inhibit the increased release of TNFα
in response to CSE (Fig. 1).
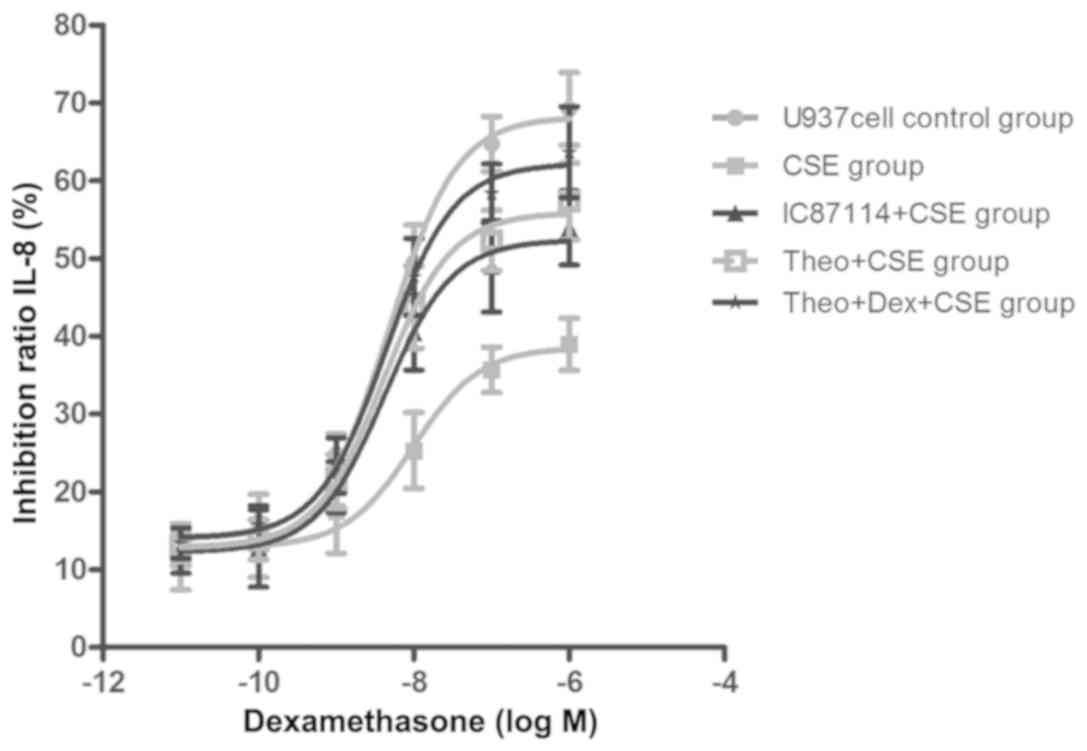
Theophylline-Dex pretreatment reduces
CSE-induced corticosteroid insensitivity
Treatment with CSE (8%) significantly increased
IC50-Dex (1.72±0.91x10-5 mol/l vs. control,
2.23±0.78x10-8 mol/l; P<0.05), which was reduced in
cells pretreated with theophylline (1.90±0.52x10-7
mol/l). A combination of theophylline and Dex was more effective
than theophylline alone (8.71±0.64x10-8 mol/l). A
positive control, CSE-treated U937 cells (7%) pretreated with
IC87114, could inhibit the increase of IC50
(2.31±0.94x10-7 mol/l) compared with the CSE group. The
results showed that U937 cells treated with CSE were less sensitive
to Dex compared with the control group, but that sensitivity was
increased in cells that had been pretreated with a combination
treatment or theophylline alone. Furthermore, combination therapy
showed an additive effect (Fig.
2).
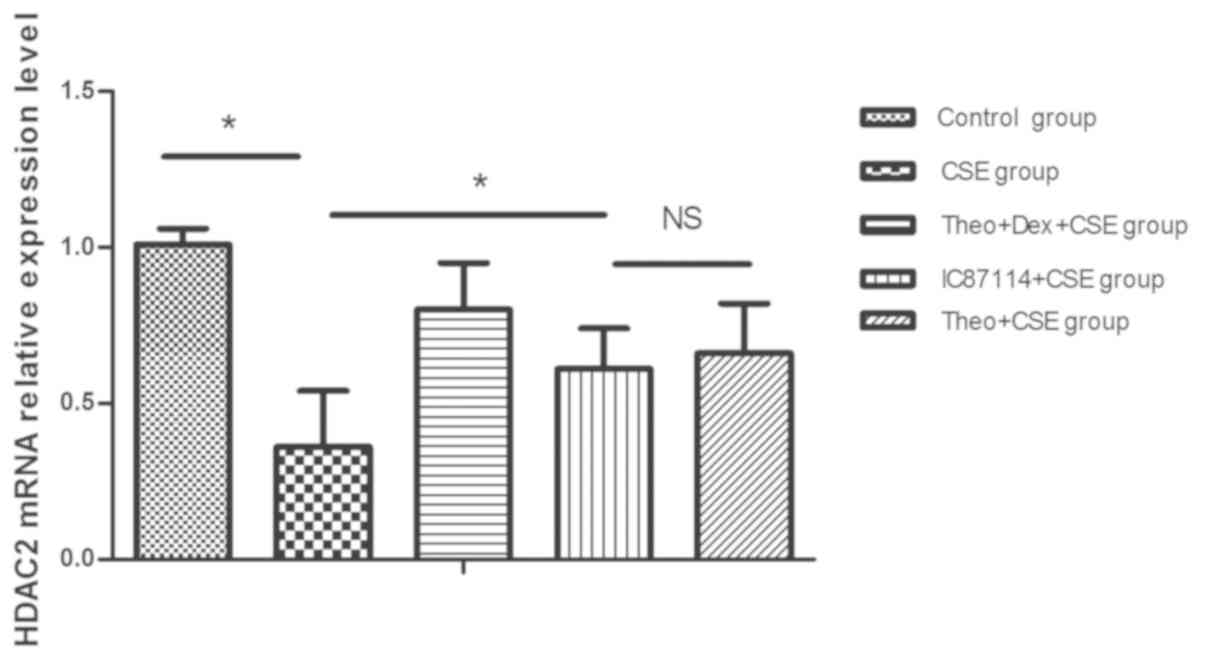
Theophylline-Dex pretreatment reduces
the decrease in HDAC2 mRNA expression seen in CSE-treated
cells
U937 cells treated with CSE expressed significantly
reduced levels of HDAC2 mRNA when compared with control cells, an
effect which was reduced in cells pre-incubated with a combination
of theophylline and Dex or theophylline alone. A combination
treatment was significantly more effective in preventing HDAC2
reduction than theophylline treatment alone. As a positive control,
pretreatment with IC87114 was also shown to significantly decrease
the level of reduction of HDAC2 mRNA expression when compared with
the CSE group. (P<0.05) (Fig.
3).
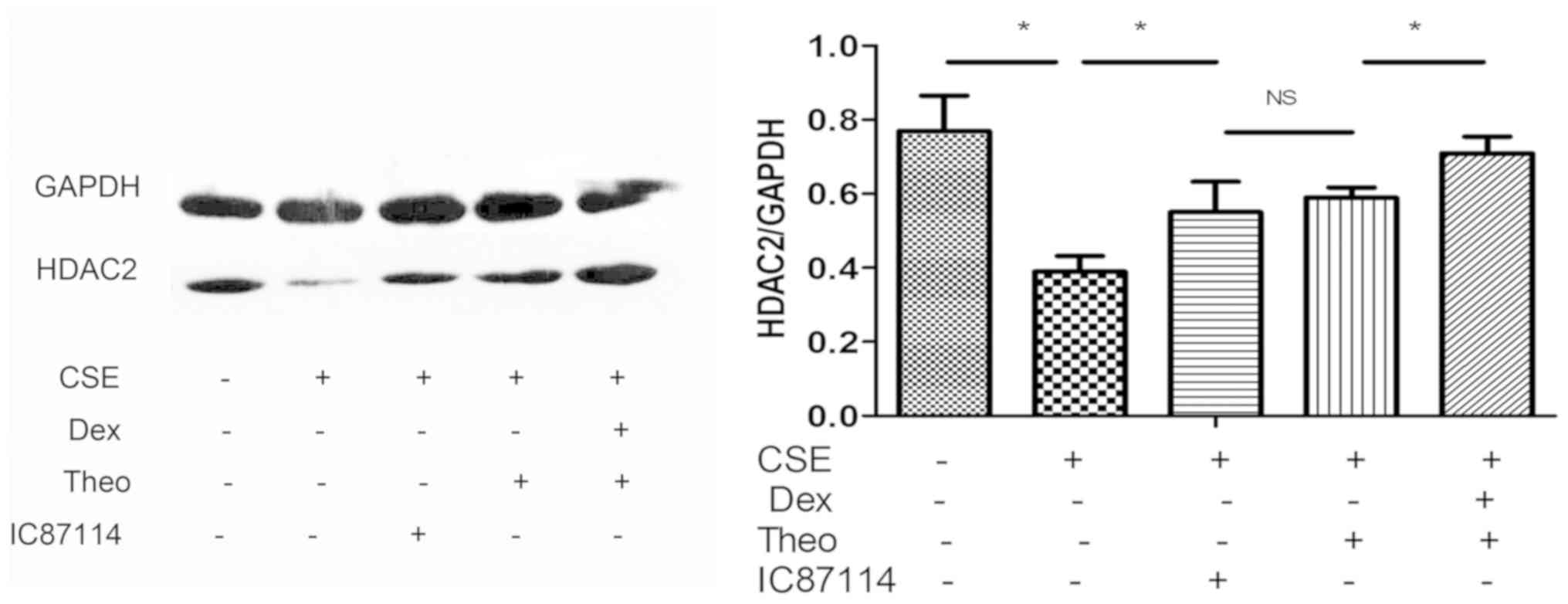
Theophylline-Dex pretreatment reduces
the decrease in HDAC2 protein expression seen in CSE-treated
cells
U937 cells treated with CSE showed a reduction in
HDAC2 protein expression, which was reversed in cells pretreated
with a combination treatment of theophylline and Dex or
theophylline alone; however, the combination treatment was
especially effective. As a positive control, pretreatment with
IC87114 was shown to decrease the reduction in HDAC2 protein
expression in U937 cells exposed to CSE (Fig. 4).
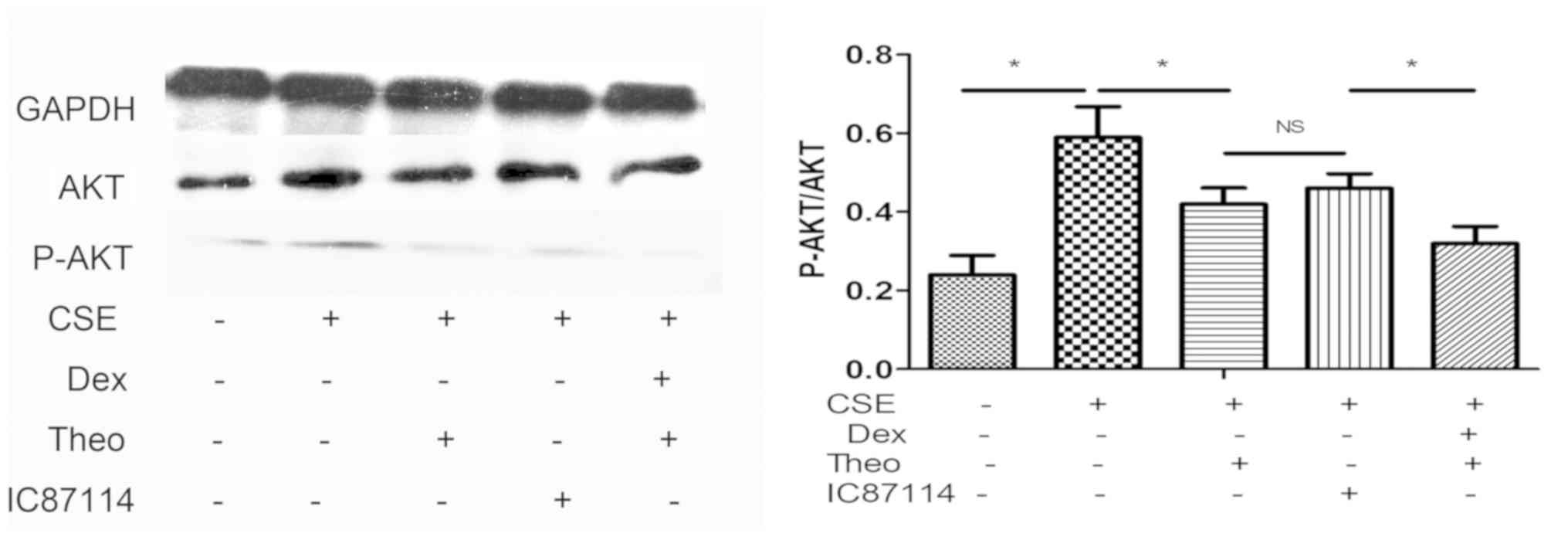
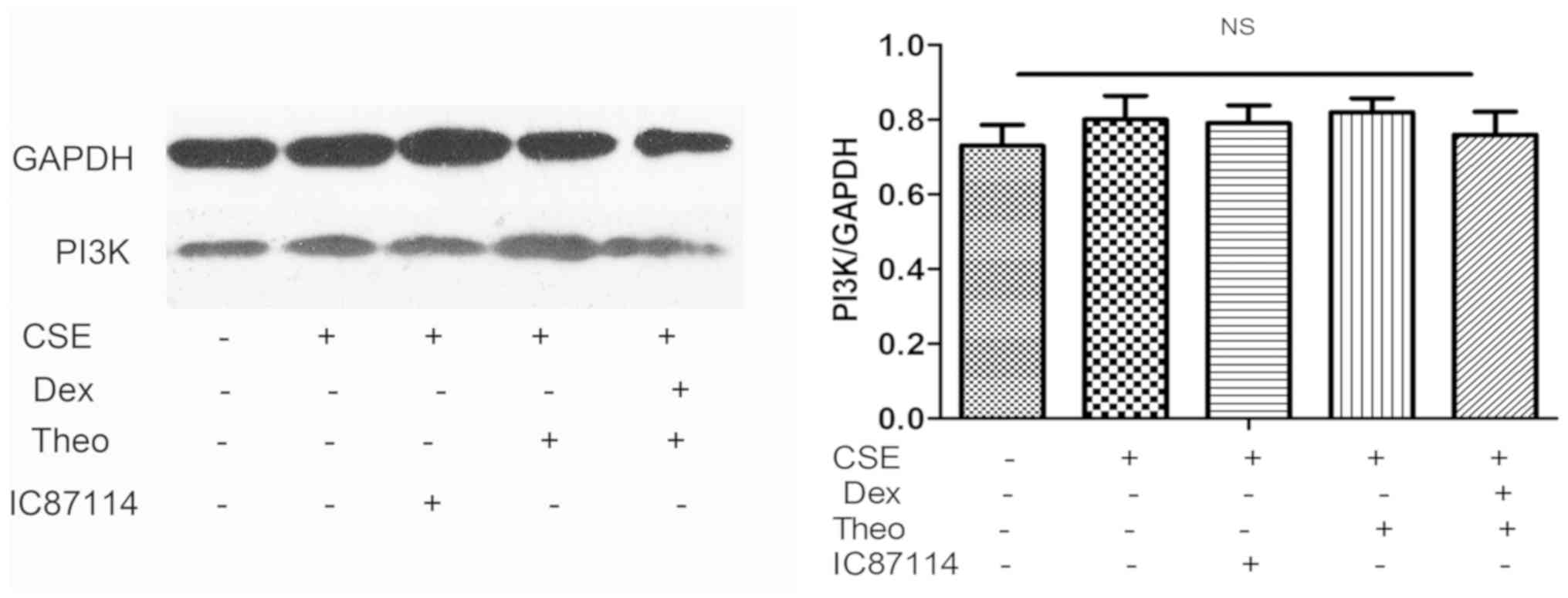
Theophylline-Dex combination decreases
Akt phosphorylation and does not affect Akt and PI3K expression
levels
CSE increased the phosphorylation of Akt protein in
U937 cells, and this was reversed by theophylline. Furthermore, a
combination of theophylline and Dex significantly decreased the
phosphorylation level of Akt protein in U937 cells exposed to CSE
compared with theophylline alone (Fig.
5). As the positive control, IC87114 decreased Akt
phosphorylation level in CSE-stimulated cells compared with the CSE
group. CSE, theophylline, IC87114, or combination of theophylline
and DEX did not affect PI3K or total Akt protein expression
(Fig. 6).
Discussion
Corticosteroids are the most effective
anti-inflammatory treatment for a number of inflammatory and immune
diseases, including asthma, rheumatoid arthritis, inflammatory
bowel disease, autoimmune diseases and COPD (14). However, some patients with severe
COPD have a poor response to high-dose corticosteroid therapy
(14). Corticosteroids have been
shown to have almost no effect on alveolar macrophages from
patients with COPD (8,27). The IC50 represents the
concentration of a substance that is required to induce half of the
maximum inhibition of a biological factor or function and is
commonly used as a measure of inhibition effectiveness. In the
present study, U937 cells were stimulated by CSE, and
IC50-Dex was used as a marker to examine corticosteroid
sensitivity, as described previously (24). The results showed that CSE-stimulated
cells were less corticosteroid sensitive than control cells,
suggesting that CSE could cause corticosteroid resistance in U937
cells in vitro.
A number of studies have shown that low-dose
theophylline can reverse corticosteroid resistance in patients with
COPD and in smoke-induced rat models (12,21-23).
A study by To et al (9)
showed that theophylline inhibited the activity of PI3K-δ that had
been precipitated from H2O2-treated cells.
However, it is unknown whether a combination of theophylline and
Dex would be more effective in reversing corticosteroid resistance
and decreasing CSE-induced inflammation. The major aim of the
current study was to explore the effect of a combination treatment
of theophylline and Dex on the inflammation in CSE-induced U937
cells and to evaluate the underlying mechanism.
The results showed that the expression levels of
PI3K and total Akt protein were not significantly different among
the CSE, IC87114, combination treatment, theophylline and control
groups. A previous report had indicated that PI3K expression in
peripheral lung macrophages from smokers with normal lung function
did not increase when compared with macrophages from nonsmokers
with normal lung function (19).
Those authors suggested that CSE could not affect PI3K or Akt
expression in cells. However, in the current study, the
phosphorylation level of Akt of CSE-stimulated cells was higher
than that of cells in the control group. The p-Akt protein level of
cells pretreated with theophylline and Dex or theophylline alone
was lower than that of cells stimulated by only CSE. Combination
treatment had a stronger effect. Moreover, IC87114, the positive
control, could decrease p-Akt levels following CSE stimulation.
These results suggested that CSE stimulation could activate the
PI3K/Akt pathway and reduce corticosteroid sensitivity. The
combination of theophylline and Dex significantly restored
corticosteroid sensitivity and decreased CSE-induced inflammation
through inhibition of PI3K/Akt pathway activation. In the current
study, p-PI3K protein was not assessed, as p-Akt was sufficient to
represent the activity of the PI3K/Akt pathway. This was congruent
with previous studies (8,9,24).
Previous studies have shown that HDAC2 expression
was lowest in COPD patients who were non-smokers for more than 1
year when compared with healthy people, healthy smokers and smokers
with COPD. Moreover, the HDAC2 expression level was gradually
reduced (23,24,28).
This suggested that oxidative stress induced by tobacco smoke
exposure reduced HDAC2 expression, which is an important cause of
corticosteroid resistance (23). It
was reported that low-dose theophylline could increase HDAC2
activity and restore glucocorticoid receptor sensitivity of
pulmonary macrophages (21). The
results of the present study showed that HDAC2 expression in
CSE-stimulated U937 cells was lower than that of control cells.
Furthermore, HDAC2 expression in cells pretreated with theophylline
and Dex was higher than that in cells pretreated with theophylline
alone. Therefore, this combination treatment was more effective in
restoring the HDAC2 expression of CSE-stimulated U937 cells in
vitro.
To the best of our knowledge, there is no known
evidence that PI3K/Akt can directly affect HDAC2 expression.
However, there may be some signaling molecules that provide a link
between the PI3K/Akt pathway and HDAC2 and indirectly modulate the
influence of PI3K/Akt on HDAC2. Ngkelo (29) showed that the activation of glycogen
synthase kinase 3β (GSK3β) could reverse corticosteroid resistance
under oxidative stress by increasing HDAC2 expression. Glycyrrhizic
acid inhibited inflammatory factors by mediation of the
PI3K/Akt/GSK3β pathway (30). Thus,
GSK3β may be the connection between PI3K/Akt and HDAC2. However,
whether other signaling molecules play an important role in the
relationship between PI3K/AKT and HDAC2 requires further
investigation (31).
In conclusion, a CSE-stimulated cell model was
developed to study the influence of theophylline and Dex on the
inflammatory effect and to explore the underlying mechanisms.
Theophylline and Dex in combination or theophylline alone were able
to reverse corticosteroid insensitivity and decrease inflammation
via inhibition of the PI3K/Akt pathway and rescue of HDAC2
expression. A combination of theophylline and Dex treatment was
more effective than theophylline treatment alone. This experiment
has potential clinical significance, as it suggested that the
combination of theophylline and Dex may be more beneficial in the
treatment of COPD and other inflammatory diseases associated with
smoke exposure than existing treatments.
Acknowledgements
The authors wish to thank Dr Zhiyi He (Department of
Respiratory and Critical Care Medicine, The First Affiliated
Hospital of Guangxi Medical University) for assistance with the
design of the present study.
Funding
This study was financially supported by the National
Nature Science Foundation of China (grant no. 81160009), Science
and Technology Department of Guangxi Province (grant no.
1598012-26) and the Liuzhou Science Research and Technology
Development Project (grant no. 2018BJ10507).
Availability of data and materials
All data generated or analyzed during this study are
included in this article.
Authors' contributions
XNZ designed the current study. XJS, ZHL, YZ, ZYH
performed the experiments. JHZ, SNC, YF analyzed the data. XJS and
ZHL wrote the manuscript. XJS and ZHL revised the manuscript. All
authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Osthoff M, Jenkins C and Leuppi J: Chronic
obstructive pulmonary disease-a treatable disease. Swiss Med Wkly.
143(w13777)2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Vestbo J, Hurd SS, Agusti AG, Jones PW,
Vogelmeier C, Anzueto A, Barns PJ, Fabbri LM, Martinez FJ,
Nishimura M, et al: Global strategy for the diagnosis, management,
and prevention of chronic obstructive pulmonary disease: GOLD
executive summary. Am J Respir Crit Care Med. 187:347–365.
2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kirkham P and Rahman I: Oxidative stress
in asthma and COPD: Antioxidants as a therapeutic strategy.
Pharmacol Ther. 111:476–494. 2006.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Barnes P: Inhaled corticosteroids are
beneficial in chronic obstructive pulmonary disease. Am J Respir
Crit Care Med. 161:341–342; discussion 344. 2000.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Culpitt SV, Maziak W, Loukidis S,
Nightingale JA, Matthews JL and Barnes PJ: Effect of high dose
inhaled steroid on cells, cytokines and proteases in induced sputum
in chronic obstructive pulmonary disease. Am J Respir Crit Care
Med. 160:1635–1639. 1999.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Hattotuwa KL, Gizycki MJ, Ansari TW,
Jeffery PK and Barnes NC: The effects of inhaled fluticasone on
airway inflammation in chronic obstructive pulmonary disease. Am J
Respir Crit Care Med. 165:1592–1596. 2002.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ford PA, Durham AL, Russell RE, Gordon F,
Adcock IM and Barnes PJ: Treatment effects of low-dose theophylline
combined with an inhaled corticosteroid in COPD. Chest.
137:1338–1644. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kobayashi Y, Wada H, Rossios C, Takagi D,
Charron C, Barnes PJ and Ito K: A novel macrolide/fluoroketolide,
solithromycin (CEM-101), reverses corticosteroid insensitivity via
phosphoinositide 3-kinase pathway inhibition. Br J Pharmacol.
169:1024–1034. 2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
To Y, Ito K, Kizawa Y, Failla M, Ito M,
Kusama T, Elliott WM, Hogg JC, Adcock IM and Barnes PJ: Targeting
phosphoinositide-3-kinase-delta with theophylline reverses
corticosteroid insensitivity in chronic obstructive pulmonary
disease. Am J Respir Crit Care Med. 182:897–904. 2010.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Barnes PJ, Adcock IM and Ito K: Histone
acetylation and deacetylation: Importance in inflammatory lung
diseases. Eur Respir J. 25:552–563. 2005.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Rahman I and Adcock IM: Oxidative stress
and redox regulation of lung inflammation in COPD. Eur Respir J.
28:219–242. 2006.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Sun X, Li Q, Gong Y, Ren L, Wan H and Deng
W: Low-dose theophylline restores corticosteroid responsiveness in
rats with smoke-induced airway inflammation. Can J Physiol
Pharmacol. 90:895–902. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
De Ruijter AJ, van Gennip AH, Caron HN,
Kemp S and van Kuilenburg AB: Histone deacetylases (HDACs):
Characterization of the classical HDAC family. Biochem J.
370:737–749. 2003.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Barnes PJ and Adcock IM: Glucocorticoid
resistance in inflammatory diseases. Lancet. 373:1905–1917.
2000.
|
|
15
|
Ito K, Caramori G and Adcock IM:
Therapeutic potential of phosphatidylinositol 3-kinase inhibitors
in inflammatory respiratory disease. J Pharmacol Exp Ther. 321:1–8.
2007.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ito K, Lim S, Caramori G, Chung KF, Barns
PJ and Adcock IM: Cigarette smoking reduces histone deacetylase 2
expression, enhances cytokine expression, and inhibits
glucocorticoid actions in alveolar macrophages. FASEB J.
15:1110–1112. 2001.PubMed/NCBI
|
|
17
|
Barned PJ: Cellular and molecular
mechanisms of asthma and COPD. Clin Sci (Lond). 131:1541–1558.
2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kok K, Geering B and Vanhaesebroeck B:
Regulation of phosphoinositide 3-kinase expression in health and
disease. Trends Biochem Sci. 34:115–127. 2009.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Marwick JA, Caramori G, Casolari P,
Mazzoni F, Kirkham PA, Adcock IM, Chung KF and Papi A: A role for
phosphoinositol 3-kinase delta in the impairment of glucocorticoid
responsiveness in patients with chronic obstructive pulmonary
disease. J Allergy Clin Immunol. 125:1146–1153. 2010.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ichiyama T, Hasegawa S, Matsubara T,
Hayashi T and Furukawa S: Theophylline inhibits NF-kappa B
activation and I kappa B alpha degradation in human pulmonary
epithelial cells. Naunyn Schmiedebergs Arch Pharmacol. 364:558–561.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Cosio BG, Tsaprouni L, Ito K, Jazrawi E,
Adcock IM and Barns PJ: Theophylline restores histone deacetylase
activity and steroid responses in COPD macrophages. J Exp Med.
200:689–695. 2004.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Cosio BG, Iglesias A, Rios A, Noguera A,
Sala E, Ito K, Barns PJ and Agusti A: Low-dose theophylline
enhances the anti-inflammatory effects of steroids during
exacerbations of COPD. Thorax. 64:424–429. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Li M, Zhong X, He Z, Wen M, Li J, Peng X,
Liu G, Deng J, Zhang J and Bai J: Effect of erythromycin on
cigarette-induced histone deacetylase protein expression and
nuclear factor-κB activity in human macrophages in vitro. Int
Immunopharmacol. 12:643–650. 2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Sun XJ, Li ZH, Zhang Y, Zhou G, Zhang JQ,
Deng JM, Bai J, Liu GN, Li MH, MacNee W, et al: Combination of
erythromycin and dexamethasone improves corticosteroid sensitivity
induced by cigarette smoke extract through inhibition of PI3K-δ/Akt
pathway and increased GR expression. Am J Physiol Lung Cell Mol
Physiol. 309:L139–L146. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Mercado N, To Y, Ito K and Barnes PJ:
Nortriptyline reverses corticosteroid insensitivity by inhibition
of phosphoinositide-3-kinase-δ. J Pharmacol Exp Ther. 337:465–470.
2011.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C (T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Culpitt SV, Rogers DF, Shah P, De Matos C,
Russell RE, Donnelly LE and Barnes PJ: Impaired inhibition by
dexamethasone of cytokine release by alveolar macrophages from
patients with chronic obstructive pulmonary disease. Am J Respir
Crit Care Med. 167:24–31. 2003.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Szulakowski P, Crowther AJ, Jimenez LA,
Donaldson K, Mayer R, Leonard TB, Macnee W and Drost EM: The effect
of smoking on the transcriptional regulation of lung inflammation
in patients with chronic obstructive pulmonary disease. Am J Respir
Crit Care Med. 174:41–50. 2006.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ngkelo A: The role of GSK3β in regulating
corticosteroid function under conditions of oxidative stress in
chronic obstructive pulmonary disease. Imperial College 2012.
|
|
30
|
Ortiz JL, Milara J, Lluch J, De Diego A,
Sanz C and Cortijo J: Phosphodiesterase-4 inhibition improves
corticosteroid insensitivity in pulmonary endothelial cells under
oxidative stress. Allergy. 68:64–73. 2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Hennessy BT, Smith DL, Ram PT, Lu Y and
Mills GB: Exploiting the PI3K/AKT pathway for cancer drug
discovery. Nat Rev Drug Discov. 4:988–1004. 2005.PubMed/NCBI View Article : Google Scholar
|