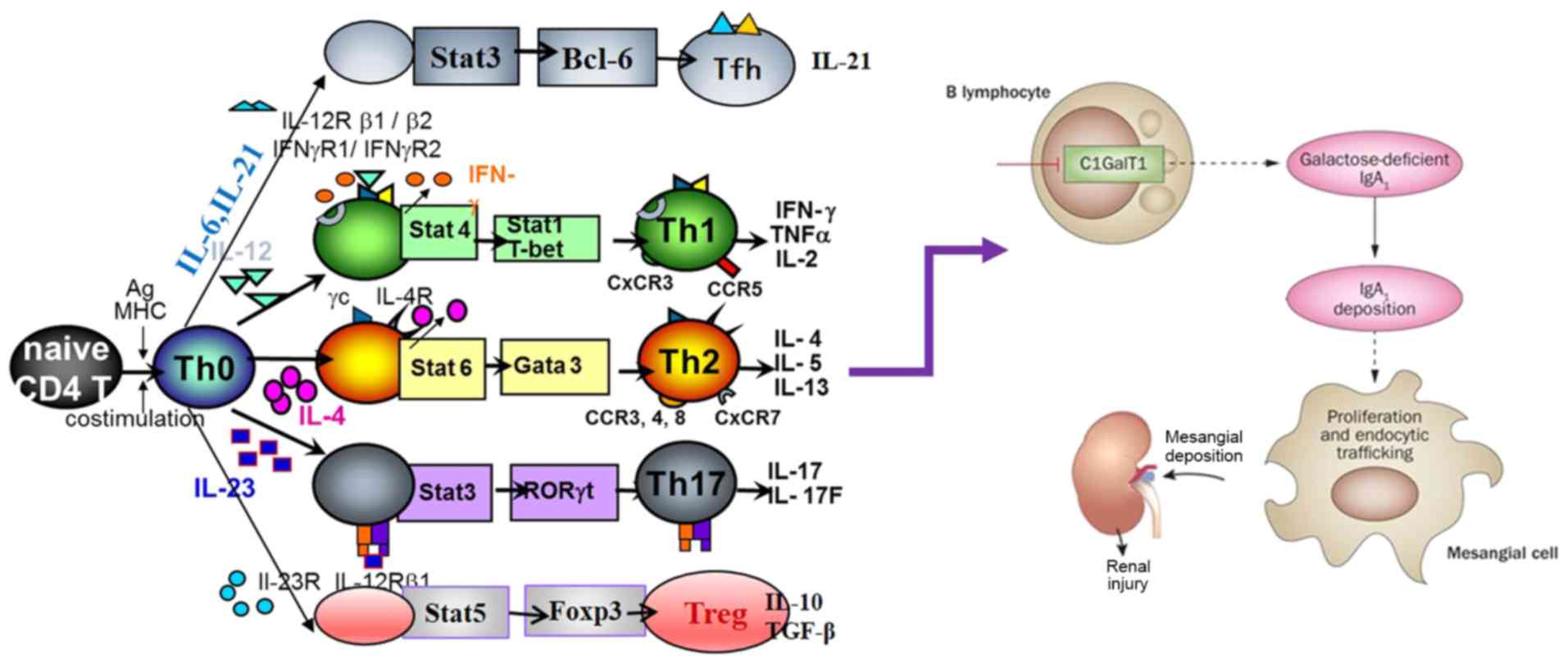
1. Introduction
Immunoglobulin A nephropathy (IgAN) is the most
common primary glomerulonephritis in the world, which normally
leads to end-stage renal disease. It is characterized by the
overexpression of IgA1 molecules, with low galactosylation in the
serum and mesangium of the kidney (1,2). There
are two subclasses of IgA in humans, IgA1 and IgA2. The main
difference between IgA1 and IgA2 is that there is a hinge region
(HR) in the IgA1 heavy chain (3).
Furthermore, in the HR of the heavy chain, there are 3 to 6
O-glycans connected to either a serine or threonine; the HR
is co-translated/post-translated by the addition of up to 6
O-glycan chains (4). These
chains include N-acetylgalactosamine (GalNAc), which is
linked by O-linkage with serine or threonine residues. The
galactose within GalNAc may be β-1, -3 using the enzyme core 1
β-1,3-galactosyltransferase (C1GalT1; the key enzyme of
galactosylation), which requires assistance from the molecular
chaperone, Cosmc; and is essential for correct folding and
stability. Galactose residues and GalNAc can be sialylated in a 2,
3- or a 2, 6-configuration, respectively.
Although the exact pathogenesis of IgAN remains
unclear, it is hypothesized to be associated with the immune
imbalance in the human body. Furthermore, the decreased expression
level of the molecular chaperone Cosmc in B cells of patients with
IgAN is associated with the number of B cells and the abnormal
glycosylation of IgA1 in IgAN (5-8).
However, the production of IgA1 with low galactosylation in IgAN
may be the result of B cell defect, and the activity of C1GalT1 is
decreased in B cells (9). This is
consistent with the observation that an increasing number of poly
IgA1 (pIgA1) plasma cells are found in the bone marrow and tonsils
of patients with IgAN with the inactivation of C1GalT1 of B cells,
and the increase in IgA1 synthesis of these plasma cells in
spontaneous culture is also observed (10,11).
Notably, pIgA1 is produced by polyclonal active B cells, and the
process of IgA production by B cells is regulated by T cells.
Lymphocytes induced by mucosal antigens produce abnormal amounts of
poorly glycosylated IgA1 and polymerized IgA-IgG immune complexes
(12,13). T lymphocytes play a key role in the
control of antigen-driven adaptive immune response; in particular
the polarization of T helper (Th) cells can affect IgAN (14-16).
It remains unclear where galactose-deficient IgA1
(Gd-IgA1) originates; however, an increasing number of studies have
revealed that Gd-IgA1 deposition could be derived from mucosally
primed plasma cells, which is related to the imbalance of a subset
of T cells (17). Furthermore,
previous studies have revealed that impaired mucosal IgA response
may lead to impaired mucosal antigen clearance. Mucosal immunity
depends on a delicate balance between antigen reactivity and
tolerance. CD4+ T cell subsets play a vital role in
maintaining or destroying this balance (18). Recent studies suggest that Th2 type
cytokines are the main cytokines secreted by circulating T cells in
IgAN. Th2 type cytokines could trigger over-secretion of abnormal
glycosylation of IgA1, which is easy to deposit in the mesangium
(19). However, additional reports
also revealed that the strong polarization of Th1 in IgAN, and the
advantage of Th1 is associated with the development of renal injury
in IgAN (20-23).
In a recent study, the upregulation of CX3CR1+
CD8+ T cells in the tonsils was found in patients with
IgAN (24). Taken together, these
findings revealed that T lymphocytes play an important role in the
development of IgAN. T cell dysregulation may induce B cells to
secrete excessive and abnormal levels of IgA1, which can lead to
IgA deposition in the glomerular and lead to injury.
In the present review, the roles of T lymphocytes in
IgAN are highlighted from the literature. Furthermore, the role of
T lymphocytes in the pathogenesis of IgAN and their potential as
therapeutic interventions and biomarkers for the treatment and
monitoring of IgAN will also be discussed.
2. T lymphocyte classification
CD4+ T cells, including Treg and effector
Th cells, play a key role in the host defense. However, the
imbalance of CD4+ T cell differentiation leads to
chronic infections as well as a variety of inflammatory allergic
and autoimmune diseases, such as chronic hepatitis B virus
infection (25), rhinallergosis
(26) and IgAN (27). The differentiation and functions of
CD4+ T cell subsets are dependent upon the cytokine
environment, which enables the differentiation into several
distinct subtypes to coordinate the immune response to clear the
pathogenic insult. At present, with respect to the lineage-specific
transcription factors, several different T cell subtypes have been
identified: T-bet for Th1 cells, GATA3 for Th2 cells, RORγt for
Th17, the master regulator Foxp3 for Treg cells, and Bcl6 for
follicular helper T cells (TFH cells) (28,29).
Each of these T cell subtypes have specific characteristic
cytokines and performs various immune functions according to the
type of cytokines (30). Th1 cells
produce pro-inflammatory cytokines, including interleukin (IL)-2
and interferon (IFN)-γ, which contribute to cellular immunity and
clearance of intracellular pathogens. Meanwhile, Th2 cells secrete
anti-inflammatory cytokines, including IL-4 and IL-10, which are
essential for humoral-mediated immunity of extracellular pathogens
(31-33).
In addition, Th17 cells include another subset of Th cells
characterized by the secretion of IL-17(34). The Th17 lineage has been revealed to
play an important role in the early response to bacterial pathogens
and the initiation of autoimmune diseases, such as rheumatoid
arthritis, IgAN and allergen-specific responses (35). Furthermore,
CD4+CD25+ Treg cells are a subgroup of
inhibitory T cells, including natural Treg (nTregs), inducible Treg
(iTregs) and thymus-derived Treg (tTregs) cells, and play a crucial
negative regulatory role in the immune system (36,37). In
a recent study, TFH cells have been revealed to be important
subsets of T cells involved in the regulation of B cell activation
and the formation of the germinal center, which can enhance the
humoral response (38). Similarly,
CD8+ T cells are part of the adaptive immune response.
With the help of CD4+ T cells, they can effectively
eliminate viruses (39,40). Novel T-lymphocyte subsets are
expected to be identified in the future.
3. Role of T lymphocytes in IgAN
T cells are important cells, which control
antigen-driven adaptive immune response. Inappropriate
differentiation of T cells leads to chronic infection, as well as
to various autoimmune diseases, such as glomerulonephritis
(41). Thus, it is important to
identify and understand the mechanisms underlying the preferential
induction of each of these T cell subsets. IgAN is hypothesized to
be associated with the immune abnormality of T cells, which leads
to abnormal signal transduction and changes in B cell response
(17,42) (Fig.
1). For example, in an animal model of IgAN, ddy mice showed
strong polarization to Th1 in the early stage of the disease
(43). In addition, the expression
of T-bet in urine sediment of newly diagnosed children with IgAN
was found to be higher compared with that in the control group,
which is associated with the positive staining of T-bet protein in
renal biopsy tissue (44).
Meanwhile, Th2 cytokine IL-4 may play an important role in
controlling the glycosylation of the IgA1 HR (45) and renal fibrosis (46). Furthermore, the Th1:Th2 ratio is
associated with proteinuria and renal pathologic changes in the
IgAN group (47). As specific
specialized signature cytokines, and the imbalance of T cells in
blood samples are relatively stable and easy to quantify, there is
potential for these to be used as biomarkers for the treatment and
monitoring of diseases. However, there is limited research on the
role of T cell subsets in the pathogenesis of IgAN and their
potential application as new therapeutic targets. Therefore,
further research is urgently needed to address this shortfall.
Here, the current literature on T cell subsets associated with IgAN
is reviewed.
4. Th1/Th2 lymphocytes
It is widely known that IgAN is the most common
immune complex associated with the cause of glomerulonephritis in
the world (48,49). There is emerging evidence which shows
that the imbalance of Th1/Th2 pro-inflammatory cytokines plays an
important role in the development and progression of IgAN (42,43). Th1
cells induce cytotoxic and cell-mediated immunity by the secretion
of IL-2, IFN-γ and tumor necrosis factor (TNF)-α, and antagonize
Th2 cell function. By contrast, Th2 cells adjust humoral immunity
via the predominant secretion of IL-4, IL-5, IL-6, and IL-13, and
antagonize Th1 cell function (18).
On the one hand, it was reported that in an animal model of IgAN,
ddy mice showed strong polarization to Th1 in the early stage of
the disease (43), while those with
quiescent disease were Th2 polarized. The level of IgA/IgG2a immune
complex in the serum was found to be significantly associated with
the degree of glomerular lesions (20). In another study, bioinformatic
analysis further revealed that three regions were aberrantly
methylated and affected the genes of CD4+ T cell
response and proliferation in patients with IgAN. This included a
total of two hypomethylated regions arranged in genes, including
TCR (T-cell receptor) signaling, DUSP3 (dual-specificity
phosphatase 3) and TRIM27 (tripartite motif-containing 27), and a
hypermethylated region involved in VTRNA2-1 (vault RNA 2-1)
non-coding RNA, also considered as the microRNA (miR)-886
precursor. Moreover, the hypermethylation of the miR-886 precursor
led to the decrease in CD4+ T cell proliferation and the
overexpression of TGF-β after TCR stimulation, which has been
proven to induce the transformation of IgA and IgG2, and increase
the secretion of IgA1 and IgA2 in human B cells (50-52).
In a study by Sallustio et al (50), the ratio of IL-2/IL-5 was
significantly increased in patients with IgAN and clearly indicated
a Th1 shift.
On the other hand, previous studies have suggested
that in severe renal insufficiency there is an increase in Th2
cytokines and IL-4 in patients with IgAN compared with that in the
controls (27,53). In addition, Th2 cytokines induce poor
glycosylation of IgA and involvement of these cytokines in
Th2-dependent modifications of the sugar chain in the
gastrointestinal mucosa and tonsils have also been demonstrated
(53-55).
Furthermore, the cytokine, IL-4, secreted by Th2 may play an
important role in controlling glycosylation of the IgA1 HR
(45) and renal fibrosis (46). A previous report demonstrated that
Th2 predominance in IgAN was associated with chronic tonsillitis.
In addition, α-hemolytic streptococcus (α-HS) promoted a Th2-type
immune response in tonsil mononuclear cells (TMCs) of IgAN
(47). Furthermore, the loss of the
encoding MAD homologue 4 (Smad4) gene in T cells leads to the
over-secretion of Th2 cytokines and the increase in the serum level
of IgA. Moreover,
Smad4co/co;Lck-cre mice
showed a large amount of glomerular IgA deposition, increased
albumin/creatinine ratio, abnormal glycosylation of IgA, complex of
IgA with IgG1 and IgG2a, and polymeric IgA, all of which are known
characteristics of human IgAN (56).
However, a previous report demonstrated that the mRNA level of IL-2
in Th1 cells in patients with IgAN was also significantly
associated with the mRNA level of IL-4 and IL-5 in Th2 cells
(57). Cumulatively, these findings
suggest that Th1/Th2 imbalance might play important roles in the
pathogenesis of IgAN due to the Th1/Th2 polarity in the systemic
immune response, which may induce the dysregulation of systemic
tolerance, followed by B-lymphocyte proliferation and the
production of abnormal IgA1. Notably, Thl cells may play a central
pathogenetic role in the early phase of IgAN. By contrast, Th2
cells could be important in the later stages of disease
progression. In addition, Thl cells and Th1 cytokines are
associated with glomerular lesions, whereas Th2 cells and Th2
cytokine expression were associated with tubulointerstitial
lesions. However, further validation studies are required to
investigate the expression of Th1/Th2 cells in different stages of
the disease.
5. Th17 lymphocytes
Th17 cells have been recently identified as a
subtype of Th cells that produce IL-17 and play a role in
nephritis, asthma and other autoimmune diseases (41,58-61).
In addition, IL-17 is involved in the pathogenesis of IgAN. In a
study of 32 patients with IgAN [16 patients with non-IgA mesangial
proliferative glomerulonephritis (MsPGN) and 32 healthy subjects],
Th17 cells were significantly increased in patients with IgAN
compared with that in the healthy controls (62). Furthermore, Meng et al
(21) demonstrated that the number
of Th17 cells and the Th17:Treg ratio was increased in mice with
IgAN, who were also revealed to have proteinuria and microscopic
hematuria, mesangial hyperplasia, IgA deposition and high electron
density deposition in the mesangial area. Moreover, the levels of
the cytokines secreted by Th17 cells, including CCL20, IL-17A, IL-6
and IL-21 were all increased in the kidneys of mice with IgAN. In
addition, different experimental groups were investigated [mice
with IgAN; mice with IgAN infected using α-HS, mice with IgAN
treated with CCL20, and mice with IgAN infected using α-HS and
treated with CCL20) and it was revealed that the manifestations in
mice with α-HS-IgAN were more severe compared with that in mice
with IgAN, but was alleviated in the CCL20-treated groups. This
study by Meng et al (21)
suggests that α-HS may aggravate renal damage in IgAN through the
response to CCL20 secreted by Th17 cells. In an additional study of
60 biopsies from patients confirmed to have IgAN and 25 healthy
controls, flow cytometric analysis revealed that the percentage of
Th17 cells in the peripheral blood was markedly higher. Moreover,
ELISA results indicated that the serum level of cytokine IL-17 was
significantly higher in patients with IgAN compared with that in
the control group (63). In
addition, a previous study revealed that, compared with normal
controls, patients with IgAN showed an increased number of Th17
cells. The serum levels of IL-17A and IL-21, secreted from Th17
cells, were increased in patients with IgAN, and serum levels of
IL-17A was associated with 24-h proteinuria. Moreover, the
expression level of IL-17A was found in 34 out of 63 patients with
IgAN at the renal tubule site. Compared with 29 patients without
IL-17A expression, these patients had decreased renal function,
increased proteinuria and more severe tubulointerstitial damage
(64). Taken together, the
aforementioned results suggest that Th17 cells and the serum level
of IL-17 may be aggravating factors in IgAN; however, to the best
of our knowledge the mechanisms involved have not been determined.
Furthermore, the sample size of the aforementioned studies was
small, thus further studies are required with a much larger cohort
of patients.
6. TFH cells
TFH cells are a distinct type of CD4+ T
cell, which specifically regulate the antibody response. It is
characterized by increased expression levels of numerous molecules,
including Bcl-6, PD-1, ICOS, CD40-ligand (CD40L), and CXCR5, as
well as IL-21 (65-67).
With respect to the expression of CCR6 and CXCR3, 3 TFH subsets
have been defined: CCR6-CXCR3+ TFH1 cells,
CCR6-CXCR3- TFH2 cells, and
CCR6+CXCR3- TFH17 cells (68). TFH cells function as crucial
mediators of the humoral responses through direct interactions with
B lymphocytes. Therefore, they are important factors in the quality
of the antibody response after antigen stimulation. Over-secretion
of TFH cells can lead to autoimmunity (38,69,70),
particularly in systemic lupus erythematosus (SLE), rheumatoid
arthritis, and Sjögren's syndrome (66). In addition, a recent study revealed
that, compared with healthy controls, significantly higher
percentages of CD4+CXCR5+,
CD4+CXCR5+PD-1+ TFH and
CD4+CXCR5+ICOS+ cells, as well as
higher serum levels of IL-2, IL-4, IL-10, IFN-γ, IL-17A and IL-21,
were detected in patients with IgAN. Moreover, the percentages of
CD4+CXCR5+ TFH cells were negatively
correlated with the values of estimated glomerular filtration rate
(eGFR), and the percentage of
CD4+CXCR5+PD-1+ TFH cells was
positively correlated with serum levels of IL-21, Gd-IgA1 and 24-h
urinary proteins (23). Taken
together, the results from the aforementioned studies indicate that
TFH cells play an important role in IgAN, which is due to the
central role of TFH cells in antibody-associated immune response.
However, the mechanism remains unclear. It is hypothesized that TFH
cells provide a co-stimulation signal to activate antigen-specific
autoreactive B lymphocytes by regulating the clonal selection of
germinal center B cells and inducing high-affinity and long-lived
plasma cell development and survival, resulting in high levels of
autoantibody secretion (71,72). Moreover, TFH cells can promote B cell
activation and plasma cell differentiation by producing IL-6 and
IL-21(73). As a result, the
production of poorly galactosylated IgA1 autoantibodies interact
with antigens to form an immune complex, which deposits on the
renal mesangium, inducing several immune and pathological changes,
eventually culminating in the development of IgAN (74,75).
However, there is limited research regarding the role of TFH cells
in the pathogenesis of IgAN and their potential as biomarkers.
Thus, further validation studies are required.
7. Tregs
Tregs, defined as
CD4+CD25+FoxP3+ cells, are
well-known to be a subset of T lymphocytes that play a central role
in developing tolerance to autoantigens through direct cell contact
or by producing anti-inflammatory cytokines, such as TGF-β1 and
IL-10 (76,77). A recent study revealed that patients
with IgAN had decreased numbers of tonsillar Tregs, which was
negatively correlated with the number of cells producing dimeric
IgA (78). In addition, another
study found that, in patients with Henoch-Schönlein purpura and
primary IgAN, mRNA expression levels of TGF-β1 and FoxP3 were
significantly lower compared with that in healthy controls
(79). Furthermore, a recent study
has shown that, compared with normal controls, patients with IgAN
had a decreased number of the
CD45RA-FoxP3high aTreg cells and amount of
IL-10. Moreover, the numbers of aTreg cells were negatively
correlated with eGFR and 24-h proteinuria. Foxp3+
monocyte infiltration was observed in the renal interstitium of
patients with IgAN, particularly in patients with >25% tubular
atrophy/interstitial fibrosis (64).
In an additional study, it was revealed that the levels of proteins
in the urine decreased and the number of red blood cells were
improved after CD4+CD25+ T cells were infused
into the IgAN rat model. Moreover, renal pathological examination
revealed that there was an improvement in renal tissue.
Immunohistochemistry analysis of IgA revealed that the deposition
of IgA in the mesangial cells was lower compared with that in a rat
IgAN model (80). Yang et al
(81) demonstrated that the number
of iTregs in patients with IgAN was significantly lower compared
with that in healthy controls. However, the nTreg levels did not
significantly change. In addition, the expression levels of TGF-β
and IL-10 in patients with IgAN were lower, whereas those of IL-17
were higher compared with that in healthy controls (81). Furthermore, in a study of 35 patients
with IgAN and 35 patients without renal disease,
CD4+CD25+ Treg cells were significantly lower
in patients with IgAN compared with that in the controls before
tonsillectomy, and were also negatively correlated with blood urea
nitrogen, IL-4 levels and proteinuria, positively correlated with
eGFR, and gradually decreased as the severity of renal histology
increased. Furthermore, serum levels of IgA, IL-2, IL-4 and IL-6
were increased while the number of CD4+CD25+
Treg cells were decreased in patients with IgAN. After
tonsillectomy, serum levels of IgA, IL-2, IL-4, IL-6, urine protein
and urine erythrocytes were decreased, while the levels of CD4
+ CD25 + Treg cells were significantly
increased but were still lower compared with that in the control
groups (22). Taken together, the
aforementioned studies have shown that Treg cells are associated
with IgAN and increasing Treg cells in patients with IgAN may lead
to clinical improvement. Adoptive transfer of autologous or
donor-derived Tregs represents an exciting immunotherapeutic
strategy for patients with IgAN. However, to the best of our
knowledge experiments have only been performed in rat models of
IgAN. Further studies are urgently required to address this
limitation.
8. Th22 cells
The newly discovered Th cell 22 (Th22) is a subset
of CD4+ T cells, which is distinguished by analyzing the
gene expression levels of C-C chemokine receptor type 4 (CCR4),
CCR6 and CCR10, which were first described in 2009 by Trifari et
al (82). These cells are
characterized by the secretion of different types of effector
cytokines, including IL-22, IL-13 and TNF-α (82). The abundance of Th22 cells and levels
of the IL-22 cytokine are increased and are positively correlated
with inflammatory and autoimmune disorders, such as psoriasis, SLE
and rheumatoid arthritis (83,84). A
recent study revealed that the numbers of Th22 cells and plasma
levels of IL-22 in patients with IgAN were significantly higher
compared with that in patients with non-IgA MsPGN and normal
controls. There was a positive correlation between the numbers of
Th22 cells and plasma levels IL-22 in patients with IgAN. In
addition, there was a significant positive correlation between Th22
and Th17 cells in patients with IgAN. Moreover, the number of Th22
cells in patients with IgAN with proteinuria was higher compared
with that in patients with IgAN without proteinuria (62). Meanwhile, in a recent exploratory
study of the number of Th22 cells in an IgAN mouse model and the
effect of losartan and dexamethasone on Th22 cells was
investigated. It was found that both drugs differentially reduced
the number of Th22 cells after 1 month, and mesangial cell
proliferation was also improved. Meanwhile, the expression of
IL-22, CCR10 and CCL27 was decreased following treatment with
either drug (85). In summary, based
on the aforementioned research, similar to Th17 cells, Th22 cells
and secreted serum IL-22 levels may be exacerbating factors in
patients with IgAN. However, there is limited research on the role
of Th22 cells in the pathogenesis of IgAN, and further studies are
required to clarify the role of Th22 cells in the pathogenesis and
progression of IgAN.
9. CD8+ cells
With the assistance of CD4+ T cells,
CD8+ T cells form part of the adaptive immune response,
functioning to effectively clear viruses (86). The role of CD4+ T cells in
the progression of IgAN has been aforementioned. Similarly, the
role of CD8+ T cells in the progression of IgAN was also
reported (24). Tomino et al
(87) reported that the most
prominent glomerular-infiltrating cell type was CD8+ T
cells in IgAN, and an immunoregulatory mechanism involving
CD8+ T cells might be one of the exacerbating factors in
patients with IgAN. Sabadini et al (88) reported that the degree of
interstitial CD8+ lymphocyte infiltration was correlated
with the severity of interstitial fibrosis and renal impairment.
Shimamine et al (89)
reported that anti-CD8 T-lymphocyte treatment produced a protective
effect against mesangial injury in ddY mice, an animal model of
spontaneous IgAN. In another study, 20 children with IgAN who were
followed for >5 years were divided into the progressive (n=5)
and non-progressive groups (n=15). It was revealed that the number
of CD8+T cells in the glomeruli and in the interstitium
was higher in the progressive group compared with that in the
non-progressive group. In addition, glomerular α-smooth muscle
actin staining, which is associated with mesangial cell
proliferation (90,91), was more intense in the progressive
group compared with that in the non-progressive group. Moreover,
urinary protein and the degree of histological changes were also
higher in the progressive group compared with that in the
non-progressive group. However, among these markers, the number of
glomerular CD8+ T cells was the most marked difference
between the two groups (92).
Meanwhile, a recent study revealed a greater distribution of
CX3CR1-positive cells in the interfollicular area of tonsils in
patients with IgAN compared with that in patients with non-IgAN.
CX3CR1-positive cells were also found in the affected renal
glomerulus of patients with IgAN. In addition, the expression level
of CX3CR1 on tonsillar CD8+ cells, cytotoxic effector
lymphocytes that includes natural killer and terminally
differentiated cytotoxic T cells (93,94),
which potentially express CD8 (94,95), was
significantly higher in patients with IgAN. Furthermore, in this
study, CpG-oligodeoxynucleotides enhanced the expression of
CD8+ cells in patients with IgAN. The chemotactic
response of tonsillar mononuclear cells to fractalkine was
significantly higher in patients with IgAN. The expression of
CX3CR1 in the peripheral blood CD8+ cells in patients
with IgAN was significantly higher and decreased after
tonsillectomy, along with the disappearance of hematuria (24). Taken together, the aforementioned
studies suggest that CD8+T cells might be one of the
exacerbating factors in patients with IgAN. In addition, the number
of glomerular CD8+ T cells may be a sensitive predictor
of disease progression in IgAN. However, to date, there is limited
research on the role of CD8+ T cells in the pathogenesis
of IgAN, and further studies are required to clarify the location
of CD8+ cells and pathophysiological role of
CD8+ T cells in IgAN.
10. Imbalance of T cell subsets as a target
for the treatment of IgAN
In addition to their general role in IgAN, which is
associated with the severity of renal damage and disease
progression, several studies have identified the imbalance of T
cell subsets as a target for the treatment of IgAN (96). Hyun et al (97) reported that, in ddY mice with high
levels of IgA (hyperserum IgA; HIGA), after injecting
adipose-derived stem cells (ASCs), which were reported to prevent
tissue damage through immunomodulating effects (98), 24-h proteinuria was markedly
decreased in all ASC-treated groups. In addition, mesangial
proliferation and glomerulosclerosis were markedly decreased in
most ASC-treatment groups. Furthermore, the beneficial effects of
ASC treatment in IgAN occurred via paracrine mechanisms that
modulate the Th1/Th2 cytokine balance, as ASC therapy significantly
decreased Th1 cytokine activity in the kidney and caused a shift to
Th2 responses in T cells from the spleen. In addition, conditioned
media from ASCs abrogated aggregated IgA-induced Th1 cytokine
production in cultured HIGA mesangial cells. Chen et al
(99) reported that, in rats with
IgAN, which were randomly divided into either the model group,
Tripterygium wilfordii glycoside (TWG) treatment group, or
the prednisone treatment group, treatment with TWG and prednisone
significantly reduced urine protein levels and urine red blood
cells at 24 h and reduced IgA deposition in renal tissue.
Furthermore, TWG showed therapeutic effects in IgAN rats and may
regulate the immune balance of Th17 and Tregs as they significantly
decreased the serum levels of IL-17 and increased the number of
Tregs in the blood. Xiao et al (85) reported that, in a mouse model of
IgAN, losartan and dexamethasone differentially decreased the
number of Th22 cells after 1 month, and mesangial cell
proliferation was also improved. Meanwhile, Zhang et al
(23) reported that treatment with
prednisone significantly reduced the number of
CD4+CXCR5+ and
CD4+CXCR5+ PD-1+ TFH cells and the
serum level of IL-21, but the treatment increased IL-4 and IL-10 in
patients with IgAN. Furthermore, in another study it was revealed
that, after CD4+CD25+ T cells were injected
into the IgAN rat model, levels of urine protein and red blood
cells were improved. Moreover, renal pathological examination
revealed that renal tissue was improved (77). Currently, patients with IgAN are
typically treated with corticosteroids and other immunosuppressants
clinically. Based on the aforementioned research, it is possible
that the immunosuppressants have therapeutic effects on IgAN by
regulating the imbalance of T cell subsets. Notably, Tregs may be
used for immunotherapy in IgAN by suppressing exuberant immune
system activation and promoting immunological tolerance. Adoptive
transfer of autologous or donor-derived Tregs represents a
promising immunotherapeutic strategy for IgAN (80,37).
Overall, statistical evidence for the involvement of the imbalance
of T cell subsets as a target for the treatment with IgAN is
speculative, rather than concrete. Further validation studies in
this area are required.
11. Future perspectives
Over the past decade, evidence has emerged that
cellular immunity plays important roles in the pathogenesis of
IgAN. Notably, the identification of specific T cells and cell
cytokines involved in the pathogenesis and progression of IgAN
highlights the imbalance of T cell subsets as new therapeutic
targets and biomarkers for the treatment and monitoring of the
disease. Currently, corticosteroids and other immunosuppressants
have shown therapeutic effects on IgAN, possibly by regulating the
imbalance of T cell subsets. Furthermore, Tregs may be used for
immunotherapy of IgAN by adoptive transfer of
CD4+CD25+ cells. However, the potential
application of T cellular immunity quantification to the clinical
care of patients with IgAN remains in the early stages of
development, with available evidence limited to small-scale
studies.
Regarding the clinically feasible imbalance of T
cell subsets as new therapeutic targets and biomarkers for the
treatment and monitoring of disease to succeed, further studies are
urgently required in numerous areas. Notably, the diagnostic role
of specific T cells and cell cytokine levels have not been
confirmed in studies of adequate sample size, different patient
populations and in patients with other forms of glomerulonephritis
and CKD as controls. As a tool for risk stratification and disease
monitoring, the relationship between all T cell subsets and the
degree of histological damage has not been investigated, and data
on the association between different T cell subsets and the rate of
decline in renal function are limited. No research groups have
studied the association between specific T cells, cell cytokine
levels and ‘hard’ renal end points (such as end-stage renal disease
and doubling of serum creatinine). In addition, no data exist on
the possible role of the serial monitoring of specific T cells,
cell cytokine levels and the association between these T cell
subsets and disease changes at different stages. Nonetheless, the
investigation of specific T cells and cell cytokine levels on
disease progression, particularly in the early phase, should be
investigated further to validate its role in risk stratification
and monitoring of patients with IgAN.
12. Conclusions
Although a limited number of studies exist, T cell
subsets have been identified as potentially relevant to the
pathogenesis of IgAN. All of the studies discussed in the present
review have shed light on the pathogenesis of IgAN, nevertheless
they all share similar methodological deficiencies. Specifically,
all have small sample sizes, and their results have undergone
limited adjustment for multiple testing or potential clinical
confounding factors. A number of the studies did not have proper
disease controls to determine whether the findings are specific for
IgAN or are generic markers of kidney damage. In addition, no data
exist on all T cell subsets, comparisons of all T cell subsets
involved in the pathogenesis and progression of IgAN, and serial
monitoring of these T cell subset changes at different stages of
IgAN. Further validation studies, preferably in different stages of
the disease and using several different populations of patients,
are required before advancements can be made and the therapeutic
implication of these T-cell subsets can be used. In a word, the
imbalance of T cell differentiation leads to abnormal proliferation
of B lymphocytes, which leads to more secretion of Gd-IgA1 and
ultimately to the occurrence of IgA nephropathy.
Acknowledgements
Not applicable.
Funding
This study was supported by Shanghai Health and
Planning Commission Scientific Research Foundation (no. 20184Y0040)
and Research on Natural Science in Minhang District of Shanghai
(no. 2018MHZ069) and the National Natural Science Foundation of
China (no. 81774060).
Availability of data and materials
Not applicable.
Authors' contributions
YT, HD, PH and XX were all responsible for the
search and review of the cited literature and writing of the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ohyama Y, Yamaguchi H, Nakajima K, Mizuno
T, Fukamachi Y, Yokoi Y, Tsuboi N, Inaguma D, Hasegawa M, Renfrow
MB, et al: Analysis of O-glycoforms of the IgA1 hinge region by
sequential deglycosylation. Sci Rep. 10(671)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kiryluk K, Li Y, Moldoveanu Z, Suzuki H,
Reily C, Hou P, Xie J, Mladkova N, Prakash S, Fischman C, et al:
GWAS for serum galactose-deficient IgA1 implicates critical genes
of the O-glycosylation pathway. PLoS Genet.
13(e1006609)2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Wang X, Li T, Si R, Chen J, Qu Z and Jiang
Y: Increased frequency of PD-1hiCXCR5- T
cells and B cells in patients with newly diagnosed IgA nephropathy.
Sci Rep. 10(492)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Xing Y, Li L, Zhang Y, Wang F, He D, Liu
Y, Jia J, Yan T and Lin S: C1GALT1 expression is associated with
galactosylation of IgA1 in peripheral B lymphocyte in
immunoglobulin a nephropathy. BMC Nephrol. 21(18)2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yeo SC, Cheung CK and Barratt J: New
insights into the pathogenesis of IgA nephropathy. Pediatr Nephrol.
33:763–777. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Perše M and Večerić-Haler Ž: The role of
IgA in the pathogenesis of IgA nephropathy. Int J Mol Sci.
20(E6199)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Knoppova B, Reily C, Maillard N, Rizk DV,
Moldoveanu Z, Mestecky J, Raska M, Renfrow MB, Julian BA and Novak
J: The origin and activities of IgA1-containing immune complexes in
IgA nephropathy. Front Immunol. 7(117)2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Hu S, Bao H, Xu X, Zhou X, Qin W, Zeng C
and Liu Z: Increased miR-374b promotes cell proliferation and the
production of aberrant glycosylated IgA1 in B cells of IgA
nephropathy. FEBS Lett. 589:4019–4025. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Sallustio F, Curci C, Di Leo V, Gallone A,
Pesce F and Gesualdo L: A new vision of IgA nephropathy: The
missing link. Int J Mol Sci. 21(E189)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Ito S, Misaki T, Naka S, Wato K, Nagasawa
Y, Nomura R, Otsugu M, Matsumoto-Nakano M, Nakano K, Kumagai H and
Oshima N: Specific strains of Streptococcus mutans, a
pathogen of dental caries, in the tonsils, are associated with IgA
nephropathy. Sci Rep. 9(20130)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Makita Y, Suzuki H, Kano T, Takahata A,
Julian BA, Novak J and Suzuki Y: TLR9 activation induces aberrant
IgA glycosylation via APRIL- and IL-6-mediated pathways in IgA
nephropathy. Kidney Int. 97:340–349. 2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Suzuki H, Fan R, Zhang Z, Brown R, Hall S,
Julian BA, Chatham WW, Suzuki Y, Wyatt RJ, Moldoveanu Z, et al:
Aberrantly glycosylated IgA1 in IgA nephropathy patients is
recognized by IgG antibodies with restricted heterogeneity. J Clin
Invest. 119:1668–1677. 2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Xu BY, Meng SJ, Shi SF, Liu LJ, Lv JC, Zhu
L and Zhang H: MicroRNA-21-5p participates in IgA nephropathy by
driving T helper cell polarization. J Nephrol: Dec 20, 2019
doi.org/10.1007/s40620-019-00682-3 (Epub ahead of print).
|
|
14
|
Serino G, Sallustio F, Cox SN, Pesce F and
Schena FP: Abnormal miR-148b expression promotes aberrant
glycosylation of IgA1 in IgA nephropathy. J Am Soc Nephrol.
23:814–824. 2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Batra A, Smith AC, Feehally J and Barratt
J: T-cell homing receptor expression in IgA nephropathy. Nephrol
Dial Transplant. 22:2540–2548. 2007.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Enya T, Miyazawa T, Miyazaki K, Oshima R,
Morimoto Y, Okada M, Takemura T and Sugimoto K: Pathologic
tonsillar findings similar to IgA nephropathy and the role of
tonsillectomy in a patient with nephrotic syndrome. BMC Nephrol.
20(381)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Meng H, Ohtake H, Ishida A, Ohta N,
Kakehata S and Yamakawa M: IgA production and tonsillar focal
infection in IgA nephropathy. J Clin Exp Hematop. 52:161–170.
2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yang Y, Liu K, Chen Y, Gong Y and Liang Y:
Indoleamine 2,3-dioxygenase (IDO) regulates Th17/Treg immunity in
experimental IgA nephropathy. Folia Biol (Praha). 65:101–108.
2019.PubMed/NCBI
|
|
19
|
Yamada K, Kobayashi N, Ikeda T, Suzuki Y,
Tsuge T, Horikoshi S, Emancipator SN and Tomino Y: Down-regulation
of core 1 beta1,3-galactosyltransferase and Cosmc by Th2 cytokine
alters O-glycosylation of IgA1. Nephrol Dial Transplant.
25:3890–3897. 2010.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Suzuki H, Suzuki Y, Aizawa M, Yamanaka T,
Kihara M, Pang H, Horikoshi S and Tomino Y: Th1 polarization in
murine IgA nephropathy directed by bone marrow-derived cells.
Kidney Int. 72:319–327. 2007.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Meng T, Li X, Ao X, Zhong Y, Tang R, Peng
W, Yang J, Zou M and Zhou Q: Hemolytic Streptococcus may exacerbate
kidney damage in IgA nephropathy through CCL20 response to the
effect of Th17 cells. PLoS One. 9(e108723)2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Huang H, Sun W, Liang Y, Peng Y, Long XD,
Liu Z, Wen X, Jia M, Tian R, Bai C and Li C: CD4 (+)CD 25 (+)Treg
cells and IgA nephropathy patients with tonsillectomy: A clinical
and pathological study. Int Urol Nephrol. 46:2361–2369.
2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhang L, Wang Y, Shi X, Zou H and Jiang Y:
A higher frequency of CD4+CXCR5+ T follicular
helper cells in patients with newly diagnosed IgA nephropathy.
Immunol Lett. 158:101–108. 2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Otaka R, Takahara M, Ueda S, Nagato T,
Kishibe K, Nomura K, Katada A, Hayashi T and Harabuchi Y:
Up-regulation of CX3CR1 on tonsillar CD8-positive cells in patients
with IgA nephropathy. Hum Immunol. 78:375–383. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Chen X, Tang Y, Zhang Y, Zhuo M, Tang Z,
Yu Y and Zang G: Tapasin modification on the intracellular epitope
HBcAg18-27 enhances HBV-specific CTL immune response and inhibits
hepatitis B virus replication in vivo. Lab Invest. 94:478–490.
2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wu YJ, Song YN, Geng XR, Ma F, Mo LH,
Zhang XW, Liu DB, Liu ZG and Yang PC: Soluble CD83 alleviates
experimental allergic rhinitis through modulating antigen-specific
Th2 cell property. Int J Biol Sci. 16:216–227. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tortajada A, Gutierrez E, Pickering MC,
Praga Terente M and Medjeral-Thomas N: The role of complement in
IgA nephropathy. Mol Immunol. 114:123–132. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Shao F, Zheng P, Yu D, Zhou Z and Jia L:
Follicular helper T cells in type 1 diabetes. FASEB J. 34:30–40.
2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Gao Y, Jin H, Nan D, Yu W, Zhang J, Yang
Y, Hou R, Qin R, Hao H, Sun Y and Tian W: The role of T follicular
helper cells and T follicular regulatory cells in the pathogenesis
of autoimmune hemolytic anemia. Sci Rep. 9(19767)2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Oestreich KJ and Weinmann AS:
Transcriptional mechanisms that regulate T helper 1 cell
differentiation. Curr Opin Immunol. 24:191–195. 2012.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wang Q, Li J, Yu TS, Liu Y, Li K, Liu S,
Liu Y, Feng Q, Zhang L, Li GS, et al: Disrupted balance of
CD4+ T-cell subsets in bone marrow of patients with
primary immune thrombocytopenia. Int J Biol Sci. 15:2798–2814.
2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Magen A, Nie J, Ciucci T, Tamoutounour S,
Zhao Y, Mehta M, Tran B, McGavern DB, Hannenhalli S and Bosselut R:
Single-cell profiling defines transcriptomic signatures specific to
tumor-reactive versus virus-responsive CD4+ T cells.
Cell Rep. 29:3019–3032.e6. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Tang Y, Chen X, Zhang Y, Tang Z, Zhuo M,
Li D, Wang P, Zang G and Yu Y: Fusion protein of tapasin and
hepatitis B core antigen 18-27 enhances T helper cell type 1/2
cytokine ratio and antiviral immunity by inhibiting suppressors of
cytokine signaling family members 1/3 in hepatitis B virus
transgenic mice. Mol Med Rep. 9:1171–1178. 2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Edwards ESJ, Bosco JJ, Aui PM, Stirling
RG, Cameron PU, Chatelier J, Hore-Lacy F, O'Hehir RE and van Zelm
MC: Predominantly antibody-deficient patients with non-infectious
complications have reduced naive B, Treg, Th17, and Tfh17 cells.
Front Immunol. 10(2593)2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Shea-Donohue T, Fasano A, Smith A and Zhao
A: Enteric pathogens and gut function: Role of cytokines and STATs.
Gut Microbes. 1:316–324. 2010.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Blokland SLM, van Vliet-Moret FM, Hillen
MR, Pandit A, Goldschmeding R, Kruize AA, Bouma G, van Maurik A,
Olek S, Hoffmueller U, et al: Epigenetically quantified immune
cells in salivary glands of Sjögren's syndrome patients: A novel
tool that detects robust correlations of T follicular helper cells
with immunopathology. Rheumatology (Oxford). 59:335–343.
2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Singer BD, King LS and D'Alessio FR:
Regulatory T cells as immunotherapy. Front Immunol.
5(46)2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Read KA, Powell MD and Oestreich KJ: T
follicular helper cell programming by cytokine-mediated events.
Immunology. 149:253–261. 2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Phillips S, Chokshi S, Riva A, Evans A,
Williams R and Naoumov NV: CD8(+) T cell control of hepatitis B
virus replication: Direct comparison between cytolytic and
noncytolytic functions. J Immunol. 184:287–295. 2010.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Tang YY, Tang ZH, Zhang Y, Zhuo M, Zang
GQ, Chen XH and Yu YS: The fusion protein of CTP-HBcAg18-27-tapasin
mediates the apoptosis of CD8(+)T cells and CD8(+) T cell response
in HLA-A2 transgenic mice. Hepat Mon. 14(e16161)2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Krebs CF and Steinmetz OM: CD4+
T cell fate in glomerulonephritis: A tale of Th1, Th17, and novel
Treg subtypes. Mediators Inflamm. 2016(5393894)2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Habura I, Fiedorowicz K, Woźniak A,
Idasiak-Piechocka I, Kosikowski P and Oko A: IgA nephropathy
associated with coeliac disease. Cent Eur J Immunol. 44:106–108.
2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Suzuki H and Suzuki Y: Murine models of
human IgA nephropathy. Semin Nephrol. 38:513–520. 2018.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Tsuruga K, Oki E, Aizawa-Yashiro T,
Yoshida H, Imaizumi T and Tanaka H: Potential Th1⁄Th2
predominance in children with newly diagnosed IgA nephropathy. Acta
Paediatr. 99:1584–1586. 2010.PubMed/NCBI View Article : Google Scholar
|
|
45
|
He L, Peng Y, Liu H, Yin W, Chen X, Peng
X, Shao J, Liu Y and Liu F: Activation of the interleukin-4/signal
transducer and activator of transcription 6 signaling pathway and
homeodomain-interacting protein kinase 2 production by tonsillar
mononuclear cells in IgA nephropathy. Am J Nephrol. 38:321–332.
2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Liu L, Kou P, Zeng Q, Pei G, Li Y, Liang
H, Xu G and Chen S: CD4+ T Lymphocytes, especially Th2 cells,
contribute to the progress of renal fibrosis. Am J Nephrol.
36:386–396. 2012.PubMed/NCBI View Article : Google Scholar
|
|
47
|
He L, Peng Y, Liu H, Yin W, Chen X, Peng
X, Shao J, Liu Y and Liu F: Th1/Th2 polarization in tonsillar
lymphocyte form patients with IgA nephropathy. Ren Fail.
36:407–412. 2014.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Takahara M, Nagato T, Nozaki Y, Kumai T,
Katada A, Hayashi T and Harabuchi Y: A proliferation-inducing
ligand (APRIL) induced hyper-production of IgA from tonsillar
mononuclear cells in patients with IgA nephropathy. Cell Immunol.
341(103925)2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Glassock RJ: Mortality risk in IgA
nephropathy. J Am Soc Nephrol. 30:720–722. 2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Sallustio F, Serino G, Cox SN, Dalla Gassa
A, Curci C, De Palma G, Banelli B, Zaza G, Romani M and Schena FP:
Aberrantly methylated DNA regions lead to low activation of CD4+
T-cells in IgA nephropathy. Clin Sci (Lond). 130:733–746.
2016.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Jang YS, Seo GY, Lee JM, Seo HY, Han HJ,
Kim SJ, Jin BR, Kim HJ, Park SR, Rhee KJ, et al: Lactoferrin causes
IgA and IgG2b isotype switching through betaglycan binding and
activation of canonical TGF-β signaling. Mucosal Immunol.
8:906–917. 2015.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Seo GY, Jang YS, Kim HA, Lee MR, Park MH,
Park SR, Lee JM, Choe J and Kim PH: Retinoic acid, acting as a
highly specific IgA isotype switch factor, cooperates with TGF-β1
to enhance the overall IgA response. J Leukoc Biol. 94:325–335.
2013.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Bai L, Li H, Li J, Song J, Zhou Y, Liu B,
Lu R, Zhang P, Chen J, Chen D, et al: Immunosuppressive effect of
artemisinin and hydroxychloroquine combination therapy on IgA
nephropathy via regulating the differentiation of CD4+ T cell
subsets in rats. Int Immunopharmacol. 70:313–323. 2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Ruszkowski J, Lisowska KA, Pindel M,
Heleniak Z, Dębska-Ślizień A and Witkowski JM: T cells in IgA
nephropathy: Role in pathogenesis, clinical significance and
potential therapeutic target. Clin Exp Nephrol. 23:291–303.
2019.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Xiao J, Wang M, Xiong D, Wang Y, Li Q,
Zhou J and Chen Q: TGF-β1 mimics the effect of IL-4 on the
glycosylation of IgA1 by downregulating core 1 β1,
3-galactosyltransferase and Cosmc. Mol Med Rep. 15:969–974.
2017.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Inoshita H, Kim BG, Yamashita M, Choi SH,
Tomino Y, Letterio JJ and Emancipator SN: Disruption of Smad4
expression in T cells leads to IgA nephropathy-like manifestations.
PLoS One. 8(e78736)2013.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Lai KN, Ho RT, Lai CK, Chan CH and Li PK:
Increase of both circulating Th1 and Th2 T lymphocyte subsets in
IgA nephropathy. Clin Exp Immunol. 96:116–121. 1994.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Kagami S: IL-23 and Th17 cells in
infections and psoriasis. Nihon Rinsho Meneki Gakkai Kaishi.
34:13–19. 2011.(In Japanese). PubMed/NCBI View Article : Google Scholar
|
|
59
|
Thomi R, Schlapbach C, Yawalkar N, Simon
D, Yerly D and Hunger RE: Elevated levels of the antimicrobial
peptide LL-37 in hidradenitis suppurativa are associated with a
Th1/Th17 immune response. Exp Dermatol. 27:172–177. 2018.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Waite JC and Skokos D: Th17 response and
inflammatory autoimmune diseases. Int J Inflamm.
2012(819467)2012.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Fu Y, Liu S, Wang Y, Ren F, Fan X, Liang
J, Liu C, Li J, Ju Y and Chang Z: GdX/UBL4A-knockout mice resist
collagen-induced arthritis by balancing the population of
Th1/Th17 and regulatory T cells. FASEB J.
33:8375–8385. 2019.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Peng Z, Tian J, Cui X, Xian W, Sun H, Li
E, Geng L, Zhang L and Zhao P: Increased number of Th22 cells and
correlation with Th17 cells in peripheral blood of patients with
IgA nephropathy. Hum Immunol. 74:1586–1591. 2013.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Yang L, Zhang X, Peng W, Wei M and Qin W:
MicroRNA-155-induced T lymphocyte subgroup drifting in IgA
nephropathy. Int Urol Nephrol. 49:353–361. 2017.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Lin FJ, Jiang GR, Shan JP, Zhu C, Zou J
and Wu XR: Imbalance of regulatory T cells to Th17 cells in IgA
nephropathy. Scand J Clin Lab Invest. 72:221–229. 2012.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Jain S, Stock A, Macian F and Putterman C:
A distinct T follicular helper cell subset infiltrates the brain in
murine neuropsychiatric lupus. Front Immunol. 9(487)2018.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Gowthaman U, Chen JS, Zhang B, Flynn WF,
Lu Y, Song W, Joseph J, Gertie JA, Xu L, Collet MA, et al:
Identification of a T follicular helper cell subset that drives
anaphylactic IgE. Science. 365(eaaw6433)2019.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Nus M, Sage AP, Lu Y, Masters L, Lam BYH,
Newland S, Weller S, Tsiantoulas D, Raffort J, Marcus D, et al:
Marginal zone B cells control the response of follicular helper T
cells to a high-cholesterol diet. Nat Med. 23:601–610.
2017.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Grados A, Ebbo M, Piperoglou C, Groh M,
Regent A, Samson M, Terrier B, Loundou A, Morel N, Audia S, et al:
T cell polarization toward TH2/TFH2 and
TH17/TFH17 in patients with IgG4-related
disease. Front Immunol. 8(235)2017.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Webb LMC and Linterman MA: Signals that
drive T follicular helper cell formation. Immunology. 152:185–194.
2017.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Makiyama A, Chiba A, Noto D, Murayama G,
Yamaji K, Tamura N and Miyake S: Expanded circulating peripheral
helper T cells in systemic lupus erythematosus: Association with
disease activity and B cell differentiation. Rheumatology (Oxford).
58:1861–1869. 2019.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Zhang Y, Long X and Wang X: Primary T-cell
transduction to study follicular helper T-cell differentiation.
Methods Mol Biol. 2111:115–126. 2020.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Patakas A, Platt AM, Butcher JP, Maffia P,
McInnes IB, Brewer JM, Garside P and Benson RA: Putative existence
of reciprocal dialogue between Tfh and B cells and its impact on
infectious and autoimmune disease. Immunol Lett. 138:38–46.
2011.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Chen Y, Yu M, Zheng Y, Fu G, Xin G, Zhu W,
Luo L, Burns R, Li QZ, Dent AL, et al:
CXCR5+PD-1+ follicular helper CD8 T cells
control B cell tolerance. Nat Commun. 10(4415)2019.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Suzuki H, Kiryluk K, Novak J, Moldoveanu
Z, Herr AB, Renfrow MB, Wyatt RJ, Scolari F, Mestecky J, Gharavi AG
and Julian BA: The pathophysiology of IgA nephropathy. J Am Soc
Nephrol. 22:1795–1803. 2011.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Wyatt RJ and Julian BA: IgA nephropathy. N
Engl J Med. 368:2402–2414. 2013.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Charbonnier LM, Cui Y, Stephen-Victor E,
Harb H, Lopez D, Bleesing JJ, Garcia-Lloret MI, Chen K, Ozen A,
Carmeliet P, et al: Functional reprogramming of regulatory T cells
in the absence of Foxp3. Nat Immunol. 20:1208–1219. 2019.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Cormican S and Griffin MD: The complex
role of interleukin 6 in regulating T-cell responses during acute
glomerulonephritis. J Am Soc Nephrol. 30:1341–1344. 2019.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Huang H, Peng Y, Liu H, Yang X and Liu F:
Decreased CD4+CD25+ cells and increased
dimeric IgA-producing cells in tonsils in IgA nephropathy. J
Nephrol. 23:202–209. 2010.PubMed/NCBI
|
|
79
|
Donadio ME, Loiacono E, Peruzzi L, Amore
A, Camilla R, Chiale F, Vergano L, Boido A, Conrieri M, Bianciotto
M, et al: Toll-like receptors, immunoproteasome and regulatory T
cells in children with Henoch-Schönlein purpura and primary IgA
nephropathy. Pediatr Nephrol. 29:1545–1551. 2014.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Shen BL, Qu QS, Miao SZ, Liu BL, Liu RY
and Gu DF: Study on the effects of regulatory T cells on renal
function of IgAN rat model. Eur Rev Med Pharmacol Sci. 19:284–288.
2015.PubMed/NCBI
|
|
81
|
Yang S, Chen B, Shi J, Chen F, Zhang J and
Sun Z: Analysis of regulatory T cell subsets in the peripheral
blood of immunoglobulin A nephropathy (IgAN) patients. Genet Mol
Res. 14:14088–14092. 2015.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Trifari S, Kaplan CD, Tran EH, Crellin NK
and Spits H: Identification of a human helper T cell population
that has abundant production of interleukin 22 and is distinct from
T(H)-17, T(H)1 and T(H)2 cells. Nat Immunol. 10:864–871.
2009.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Azizi G, Rastegar Pouyani M, Navabi SS,
Yazdani R, Kiaee F and Mirshafiey A: The newly identified T helper
22 cells lodge in leukemia. Int J Hematol Oncol Stem Cell Res.
9:143–154. 2015.PubMed/NCBI
|
|
84
|
Xiao C, Xiao P, Li X, Huang G, Li H and
Chen Y: Streptococcus may aggravate inflammatory damage in
chronic nephritis via the chemotaxis of Th22 cells. Am J Transl
Res. 11:7432–7440. 2019.PubMed/NCBI
|
|
85
|
Xiao C, Zhou Q, Li X, Li H, Zhong Y, Meng
T, Zhu M, Sun H, Liu S, Tang R, et al: Losartan and dexamethasone
may inhibit chemotaxis to reduce the infiltration of Th22 cells in
IgA nephropathy. Int Immunopharmacol. 42:203–208. 2017.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Liu K, Yang Y, Chen Y, Li S, Gong Y and
Liang Y: The therapeutic effect of dendritic cells expressing
indoleamine 2,3-dioxygenase (IDO) on an IgA nephropathy mouse
model. Int Urol Nephrol. 52:399–407. 2020.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Tomino Y, Ozaki T, Koide H, Yagame M,
Eguchi K, Nomoto Y and Sakai H: Glomerular T cell and monocyte
populations in patients with IgA nephropathy. Nihon Jinzo Gakkai
Shi. 31:221–226. 1989.PubMed/NCBI
|
|
88
|
Sabadini E, Castiglione A, Colasanti G,
Ferrario F, Civardi R, Fellin G and D'Amico G: Characterization of
interstitial infiltrating cells in Berger's disease. Am J Kidney
Dis. 12:307–315. 1988.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Shimamine R, Shibata R, Ozono Y, Harada T,
Taguchi T, Hara K and Kono S: Anti-CD8 monoclonal antibody protects
against spontaneous IgA nephropathy in ddY mice. Nephron.
78:310–318. 1998.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Johnson RJ, Iida H, Alpers CE, Majesky MW,
Schwartz SM, Pritzi P, Gordon K and Gown AM: Expression of smooth
muscle cell phenotype by rat mesangial cells in immune complex
nephritis. Alpha-smooth muscle actin is a marker of mesangial cell
proliferation. J Clin Invest. 87:847–858. 1991.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Alpers CE, Hudkins KL, Gown AM and Johnson
RJ: Enhanced expression of ‘muscle-specific’ actin in
glomerulonephritis. Kidney Int. 41:1134–1142. 1992.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Watanabe T, Kawachi H, Ikezumi Y,
Yanagihara T, Oda Y and Shimizu F: Glomerular CD8+ cells predict
progression of childhood IgA nephropathy. Pediatr Nephrol.
16:561–567. 2001.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Segerer S, Hughes E, Hudkins KL, Mack M,
Goodpaster T and Alpers CE: Expression of the fractalkine receptor
(CX3CR1) in human kidney diseases. Kidney Int. 62:488–495.
2002.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Nishimura M, Umehara H, Nakayama T, Yoneda
O, Hieshima K, Kakizaki M, Dohmae N, Yoshie O and Imai T: Dual
functions of fractalkine/CX3C ligand 1 in trafficking of
perforin+/granzyme B+ cytotoxic effector
lymphocytes that are defined by CX3CR1 expression. J Immunol.
168:6173–6180. 2002.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Addison EG, North J, Bakhsh I, Marden C,
Haq S, Al-Sarraj S, Malayeri R, Wickremasinghe RG, Davies JK and
Lowdell MW: Ligation of CD8alpha on human natural killer cells
prevents activation-induced apoptosis and enhances cytolytic
activity. Immunology. 116:354–361. 2005.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Yamanaka T, Tamauchi H, Suzuki Y, Suzuki
H, Horikoshi S, Terashima M, Iwabuchi K, Habu S, Okumura K and
Tomino Y: Release from Th1-type immune tolerance in spleen and
enhanced production of IL-5 in Peyer's patch by cholera toxin B
induce the glomerular deposition of IgA. Immunobiology.
221:577–585. 2016.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Hyun YY, Kim IO, Kim MH, Nam DH, Lee MH,
Kim JE, Song HK, Cha JJ, Kang YS, Lee JE, et al: Adipose-derived
stem cells improve renal function in a mouse model of IgA
nephropathy. Cell Transplant. 21:2425–2439. 2012.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Hong SJ, Traktuev DO and March KL:
Therapeutic potential of adipose-derived stem cells in vascular
growth and tissue repair. Curr Opin Organ Transplant. 15:86–91.
2010.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Chen F, Ma YL, Ding H and Chen BP: Effects
of Tripterygium wilfordii glycosides on regulatory T cells
and Th17 in an IgA nephropathy rat model. Genet Mol Res.
14:14900–14907. 2015.PubMed/NCBI View Article : Google Scholar
|