Introduction
As the predominant pathogen in both community- and
hospital-acquired wound infections, Staphylococcus aureus
(S. aureus) has attracted widespread attention and has been
extensively studied (1). Bacterial
colonization, which is defined as the presence of replicating
microorganisms adherent to the wound in the absence of tissue
damage, is the initial step in the development of wound infection
(2). A variety of surface-associated
factors are involved in the attachment and colonization of S.
aureus to host cells and extracellular matrix, such as wall
teichoic acid (WTA), fibronectin-binding protein A/B (FnBPA/B),
iron-regulated surface determinant A (IsdA), clumping factor A/B
and collagen-binding protein (2,3). In
particular, WTA (a surface-exposed polyanionic polymer of the cell
wall), which is biosynthesized by the enzyme UDP-phosphate
N-acetylglucosaminyl 1-phosphate transferase (tagO), serves
a crucial role in the initiation of S. aureus colonization
(4). FnBPA and IsdA, which are
members of the microbial surface components recognizing adhesive
matrix molecules protein family and encoded by fnbA and
isdA, respectively, serve a leading role in prolonged
persistence in wounds (2,5).
Bacterial colonization alone was reported to exhibit
little harm to host cells and the wound healing process (6,7).
However, colonized bacteria can form complex microbial communities,
which are called biofilms, and embed themselves in extracellular
polymeric substances (EPS), serving as a shield against host immune
cells and antibacterial agents (8).
Polysaccharide intercellular adhesion (synthesized by the enzyme
encoded by the ica operon) and extracellular DNA
[cooperatively regulated by the holin-like protein CidA
(cidA)/antiholin-like protein LrgA (lrgA) network and
LysR family regulatory protein CidR (cidR)] are the main
components of EPS, exhibiting an important effect on intercellular
adhesion and biofilm formation in vitro and in vivo
(8,9). With the protection of the biofilm
matrix, S. aureus proliferates continually to a critical
cell density, at which point the agr quorum-sensing system
is activated (10). This
interbacterial chemical communication causes the upregulation of
RNAIII, the main effector molecule of the agr system,
which results in the production of a variety of virulence factors
and subsequent tissue necrosis (11). Wound infection is defined as the
presence of replicating organisms within a wound with subsequent
host injury (6). The transition from
the bacterial colonization form to the sessile biofilm form makes
the infection more complex, further damaging wound healing and
reepithelialization (12,13).
In recent years, there has been increasing interest
in the use of negative pressure wound therapy (NPWT) for the
treatment of infected wounds due to its efficacy (14-16).
Despite growing research concerning the low incidence of
biofilm-associated infections, the mechanism of NPWT in infection
control remains to be elucidated, particularly with regard to the
changes of bacteria secondary to NPWT (17,18).
S. aureus responds to physical stimulations by regulating
gene expression (19-22).
Therefore, further studies are urgently required to determine the
existent form and gene expression pattern of S. aureus in
the microenvironment established by NPWT.
The aim of the present study was to investigate and
evaluate the response of S. aureus to NPWT during acute
wound infection. An established rabbit model was used to examine
the distribution and existing form of S. aureus in tissues.
Bacterial burden was also determined to confirm the survival and
persistence of bacteria. The adaptive expression of bacterial genes
associated with colonization and biofilm regulation secondary to
NPWT was further monitored. These experiments aimed to develop an
improved understanding of the role of NPWT in bacterial existent
form and infection control, setting a foundation for further
elucidation of its mechanism.
Materials and methods
Animal ethics
A total of 18 1-year-old female Japanese large-ear
white rabbits (~3 kg, purchased from the Laboratory Animal Centre
of the Academy of Military Medical Sciences) were used for the
present study. The animals were acclimated to standard housing and
fed ad libitum under a constant temperature (22̊C) and
humidity (45%) with a 12 h light/dark cycle. All experiments were
approved by the Medical Ethics Committee of the Chinese People's
Liberation Army (PLA) General Hospital (approval no. 2014-X9-8) in
compliance with the Guidelines for Care and Use of Animals in
Research (23).
Bacterial strain and culture
S. aureus strain RN6390 with constitutive
green fluorescent protein (GFP) expression (obtained from the
Chinese PLA Institute for Disease Control and Prevention) was
cultured in Luria-Bertani (LB) broth in a rotary shaker at 200 rpm
and 37̊C. S. aureus was grown overnight and subcultured
until the log phase. To prepare the inoculum, bacteria were
collected by centrifugation (5,000 x g, 4̊C, 10 min), washed thrice
with PBS and suspended to an optical density value of 1.0 at 600 nm
measured using an ultraviolet spectrophotometer (GeneQuant 1300;
Biochrom, Ltd.) in PBS, equivalent to 1x105
colony-forming units (CFUs)/µl empirically (24).
Rabbit model of acute wound
infection
The wounding and bacterial inoculation protocol was
based on a previously published wound model with minor
modifications (25,26). In brief, rabbits were anesthetized by
intramuscular injection of ketamine (45 mg/kg) and xylazine (5
mg/kg) mixture. The back of each animal was shaved and residual
hair was removed with depilatory cream. The surgical site was
sterilized twice with 70% ethanol. A standardized 3 cm-diameter
full-thickness dermal wound was created below the dorsal muscle on
each side of the back. Following hemostasis and confirmation of no
active bleeding, each wound was inoculated with a total of
1x107 CFUs of S. aureus at a volume of 0.1 ml.
The wounds were bandaged with sterile gauze dressings. Bacteria
were allowed to proliferate in vivo for a minimum of 6 h to
ensure bacterial adhesion and colonization (27). All procedures in the present study
were performed by the same surgeon.
Study design and treatment
protocol
All rabbits were used to create acute wound
infection models (two wounds/rabbit). For each animal, the two
wounds were respectively and randomly assigned to the ‘untreated
control side’ and the ‘NPWT side’. The rabbits were divided into
two groups: Group A (bacterial count, gene expression analysis and
healing condition, n=10) and group B (histological study, n=8).
The wounds were treated based on a previously
published protocol with minor modifications (25,26). In
brief, at 6 h post inoculation, the wound on the ‘NPWT side’ of
each rabbit was dressed with the standard NPWT dressing (consisting
of polyvinyl alcohol foam, semiocclusive transparent dressing and
suction tube; Wuhan VSD Medical Science and Technology Co., Ltd.)
trimmed to the appropriate size in advance. The suction tube was
then connected to the vacuum pump device (kindly provided by
Professor Lei Hu, Beihang University, Beijing, China). The rabbits
were single-caged and the vacuum pump device was placed outside the
cage. The tubing and the wound area were swathed in bandages to
prevent the rabbits from biting it. Wounds treated with NPWT were
subjected to continuous negative pressure at -125 mmHg. Dressings
were checked daily and changed every 48 h at most, as recommended
for infected wounds by the manufacturer. If the wound required
imaging on that day, dressings were also changed. After a period of
8 days of treatment, the rabbits in group B were euthanized. The
healing conditions of the rabbits in group A were observed until
the 28th day after operation. The wound on the contralateral side
of each rabbit was bandaged and followed the same protocol as the
treated wound, acting as the untreated control. All animals in
group A were euthanized at the end of the present study.
Histological analysis
Samples (1x1x1 cm) from the center of the wounds
were harvested immediately after animal euthanasia on postoperative
day 8. Each sample was equally divided into three parts, which were
used for laser scanning confocal microscopy (LSCM), hematoxylin and
eosin (H&E) staining and scanning electron microscopy
(SEM).
To visualize the distribution of GFP-labeled S.
aureus in the tissue, the specimens were embedded in optimal
cutting temperature compound (Sakura Finetek USA, Inc.),
subsequently snap-frozen and stored in liquid nitrogen until
cryosectioning. Cryosections measuring 6 µm at the vertical section
were obtained using a Leica CM1950 freezing microtome (Leica
Microsystems GmbH). Observation of section slides was performed
using an FV1000 confocal laser scanning microscope (Olympus
Corporation; x200 magnification). For better evaluation,
transmitted light images were captured and merged with
GFP-fluorescent images (5).
For histopathological analysis, duplicate samples
from the wounds above were fixed in 10% neutral formalin for 24 h
at 25̊C, embedded in paraffin, sectioned vertically into 4 µm
slices and stained with H&E. Images were captured with an
Olympus BX51 microscope (Olympus Corporation) equipped with an
Olympus DP71 digital camera (Olympus Corporation).
Duplicate samples from the wounds above were fixed
in 2.5% glutaraldehyde (24 h) and 1% osmium tetroxide (12 h) at
25̊C, dehydrated through an ethanol series and
hexamethyldisilazane, mounted to specimen stubs using double-sided
tape and coated with gold in an auto sputter coater (TedPella,
Inc.). Imaging of the tissue samples was performed using a scanning
electron microscope (S-3400N; Hitachi, Ltd.). Digital images were
captured using the same parameters (15.0 kV, x5,000 magnification)
and analyzed by three blinded experienced observers (one
microbiologist and two pathologists). Wound sample and granulation
tissue was evaluated as described by Kamamoto et al
(28). Bacterial aggregates and
necrosis were observed in the region of interest, which were
regions with bacterial aggregates and necrosis below the wound
surface. Five sections were evaluated per sample.
Viable bacteria count measurement
Wound samples were harvested under anesthesia as
described by Morykwas et al (29) on postoperative days 1, 4 and 8. In
brief, each wound was equally divided into four quadrants. Three
samples (~0.1 g in total) were randomly obtained with biopsy
forceps in one of the quadrants at each time point (days 1, 4 and
8). Specimens were immediately weighed and homogenized into a 1 ml
suspension with sterile PBS at 4̊C. The homogenates were then
serially diluted, plated on Staphylococcus Isolation Agar (Hardy
Diagnostics) and incubated at 37̊C for 24 h. The standard
colony-counting method (25) was
performed and the results were expressed as the logarithm of CFUs
per g of tissue.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Wound samples (~0.1g for both groups) collected
under anesthesia on postoperative days 1, 2, 4 and 8 were
snap-frozen and stored in liquid nitrogen until RNA extraction and
subsequent RT-qPCR analysis as previously described (5). Briefly, frozen tissue was pulverized
using a hammer, homogenized in the presence of cell disruption
solution (cat. no. 4305895; Thermo Fisher Scientific, Inc.) and
lysozyme (AC142; Omega Bio-Tek, Inc.) and incubated at 37̊C for 10
min. Total RNA was extracted using a RNAprep Pure Cell/Bacteria kit
(Tiangen Biotech Co., Ltd.) according to the manufacturer's
instructions. Isolated RNA was treated with Recombinant DNase I
(Takara Bio, Inc.) at 37̊C for 20 min to remove contaminating DNA.
RNA quality and concentration were assessed using a
Qubit® 2.0 Fluorometer (Thermo Fisher Scientific, Inc.).
RNA was reverse transcribed into cDNA using a TIANScript RT kit
(Tiangen Biotech Co., Ltd.) according to the manufacturer's
instructions. The primer pairs used in the present study are listed
in Table I. Transcript levels of the
selected S. aureus genes were evaluated by RT-qPCR analysis
using a KAPA SYBR Fast qPCR kit (Roche Diagnostics GmbH) on a
7900HT Fast Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The following thermocycling conditions were used
for the PCR: Initial denaturation at 95̊C for 3 min; 40 cycles of
95̊C for 3 sec, 60̊C for 20 sec; and one dissociation step at 95̊C
for 15 sec, 60̊C for 15 sec and 95̊C for 15 sec. All samples were
analyzed in triplicate and Ct values were normalized against 16S
ribosomal RNA expression. The
2-ΔΔCq method (30) was used to determine the relative
expression levels of S. aureus genes in accordance with the
manufacturer's protocol. A total of 200 µl of the inoculum was
retained when establishing the animal model and snap-frozen in
liquid nitrogen. The inoculum was used as the calibrator with a
value of 1. The results were expressed as the mean fold-change of
gene expression level relative to that of the same gene found in
the inoculum.
 | Table IPrimer sequences used for reverse
transcription-quantitative PCR. |
Table I
Primer sequences used for reverse
transcription-quantitative PCR.
| | Primer |
|---|
| Gene | Description | Forward | Reverse |
|---|
| icaA |
Poly-beta-1,6-N-acetyl-D-glucosamine
synthase |
5'-TAAGCGAAGTCAGACACTTGCT-3' |
5'-GCTTCCAAAGACCTCCCAAT-3' |
| cidA | Holin-like protein
CidA |
5'-CTAACTTGGGTAGAAGACGGTG-3' |
5'-AACCTGAAGATAATGCAACGAT-3' |
| lrgA | Antiholin-like
protein LrgA |
5'-GTGCTGTTAAGTTAGGCGAAG-3' |
5'-GTGCTTGGCTAATGACACCTA-3' |
| cidR | LysR family
regulatory protein CidR |
5'-TTCTTGGTGCATTCCATCAG-3' |
5'-GCAAAGTGGTCACGCCTAT-3' |
| tagO | UDP-phosphate
N-acetylglucosaminyl 1-phosphate transferase |
5'-TATTTCGTTGCCAATGGGTA-3' |
5'-CAAACCATCGAGTCCATCAA-3' |
| fnbA | Fibronectin-binding
protein A |
5'-ACACAGCTATAGATGGTGGAGG-3' |
5'-CGCTATCCACAGCAGTATGG-3' |
| isdA | Heme transporter
IsdA |
5'-CAACAGCGAAATCTGAAAGC-3' |
5'-TTGTTTAGGCGTTTCGTTATG-3' |
| RNAIII | Quorum-sensing
regulator |
5'-AGATATCATTTCAACAATCGGTG-3' |
5'-TAGTGAATTTGTTCACTGTGTCGA-3' |
| 16S
rRNA | Ribosome
component |
5'-AGTGAAAGACGGTCTTGCTGTC-3' |
5'-ATTGCGGAAGATTCCCTACTG-3' |
Measurement of wound closure
Images of the wounds in group A were captured with a
digital camera (IXUSi; Canon, Inc.) every 3 days until
postoperative day 28. All images were independently analyzed by two
expert pathologists to evaluate and delimit the epithelialisation
boundary. The wound area was then calculated using ImagePro Plus
version 6.0 software (Media Cybernetics, Inc.) with the scale set
using the ruler in the image. The rate of wound closure was
expressed as the percentage of the initial wound area.
Statistical analysis
Experimental data were analyzed using a mixed ANOVA
followed by Bonferroni's post hoc test to evaluate the difference
between groups at each time point. All analyses were performed
using SPSS 17.0 software (SPSS, Inc.) and data were presented as
the mean ± standard deviation. P<0.05 was considered to indicate
a statistically significant difference.
Results
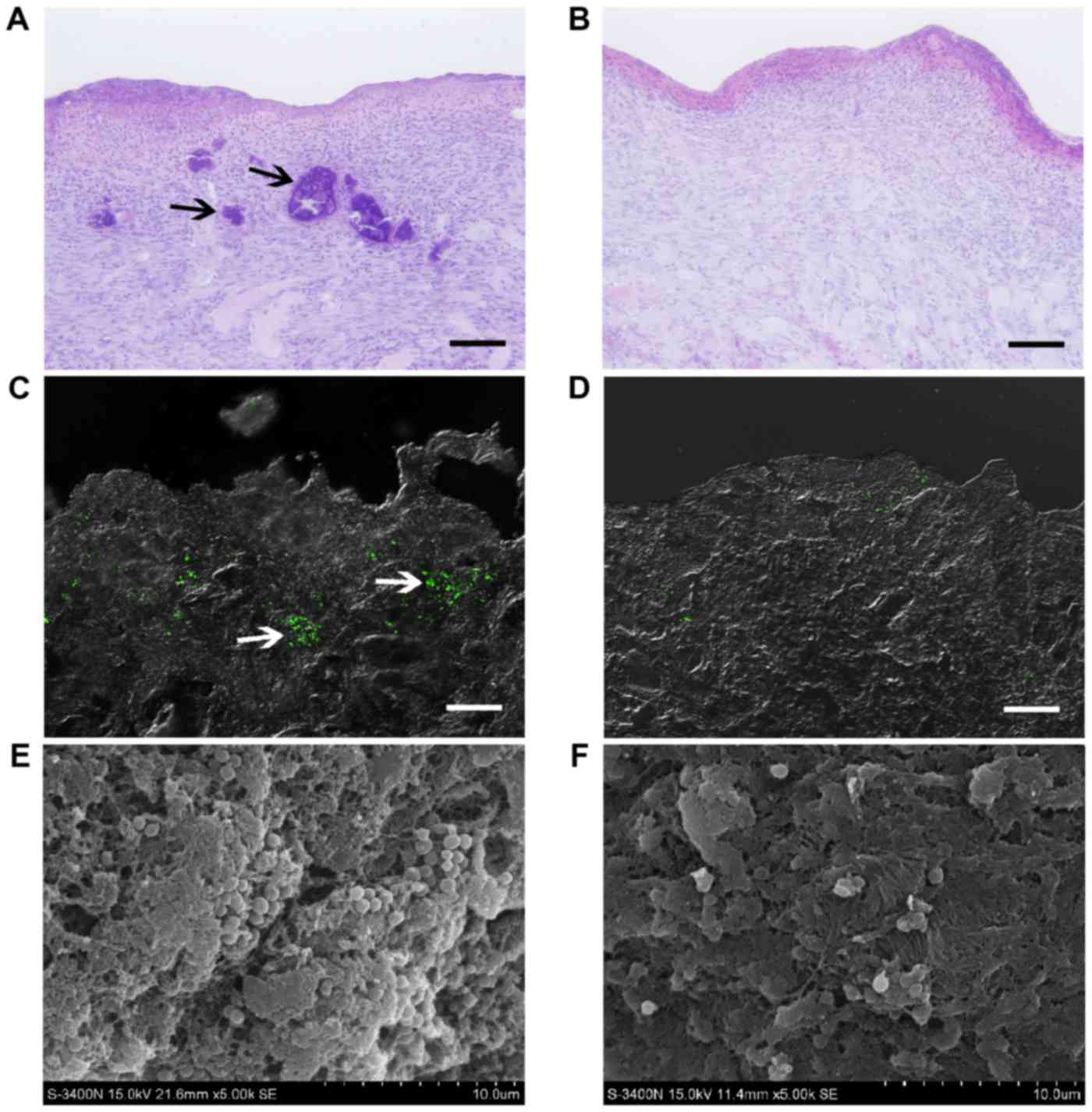
Histological examination and detection
of S. aureus in soft tissue
Sections of samples from the affected wounds at the
untreated control side presented with numerous discrete aggregates
and microcolonies of bacteria (indicated by black arrows; ~100 µm
in size; Fig. 1A) located in the
tissue, observed by conventional light microscopy on postoperative
day 8. These bacterial colonies caused local necrosis, leaving
cavities with distinct boundaries (indicated by black arrows).
However, bacterial aggregates were not detected at the NPWT side,
instead presenting with a healthier granulation bed without local
necrosis (Fig. 1B). LSCM utilizing
GFP-tagged S. aureus showed that bacteria appeared as
clusters and biofilms (~100 µm in size) in untreated wounds,
resulting in distinct regions in deep tissue layers (indicated by
white arrows; ~300 µm in depth; Fig.
1C). In comparison, bacteria in wounds treated with NPWT were
sparsely visible and dispersedly distributed in the tissue
(Fig. 1D). To confirm these results,
SEM was performed, presenting with numerous S. aureus
microcolonies (indicated by white arrows) embedded within the
lattice-like extracellular matrix in untreated wounds (Fig. 1E). Conversely, the bacteria in
tissues treated by NPWT appeared as single cells or diplococci,
lacking an extracellular matrix (Fig.
1F).
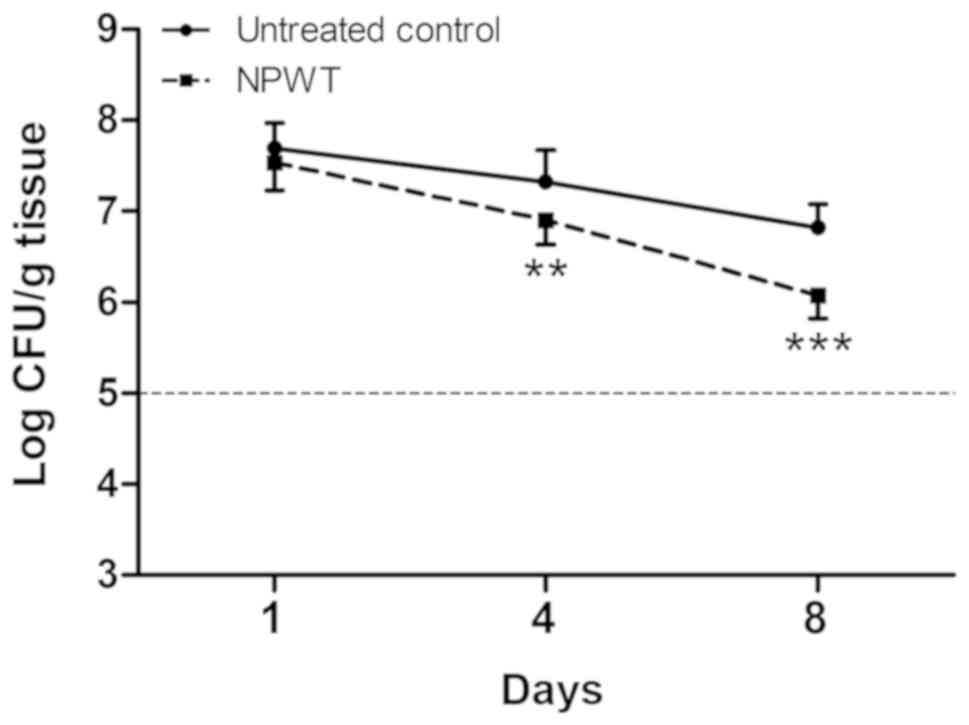
Viable bacteria count measurement
To investigate the effects of NPWT on the survival
of S. aureus, bacterial burden was determined at multiple
time points (Fig. 2). On
postoperative day 1, there was no statistical difference between
untreated and NPWT groups, with an average of 4.05x107
CFUs per g of tissue. Bacterial counts of untreated wounds showed a
consistent level of viable bacteria throughout the course of time.
In contrast, there was a significant decrease in viable bacteria
levels in the NPWT group compared with the untreated group,
verifying the consistency and reproducibility of the results in our
previous work (25). Nevertheless,
NPWT only led to a reduction in bacterial counts by one-log fold on
day 8, with an average of 1.16x106 CFUs/g, which was
still a considerable amount of bacteria in the tissue.
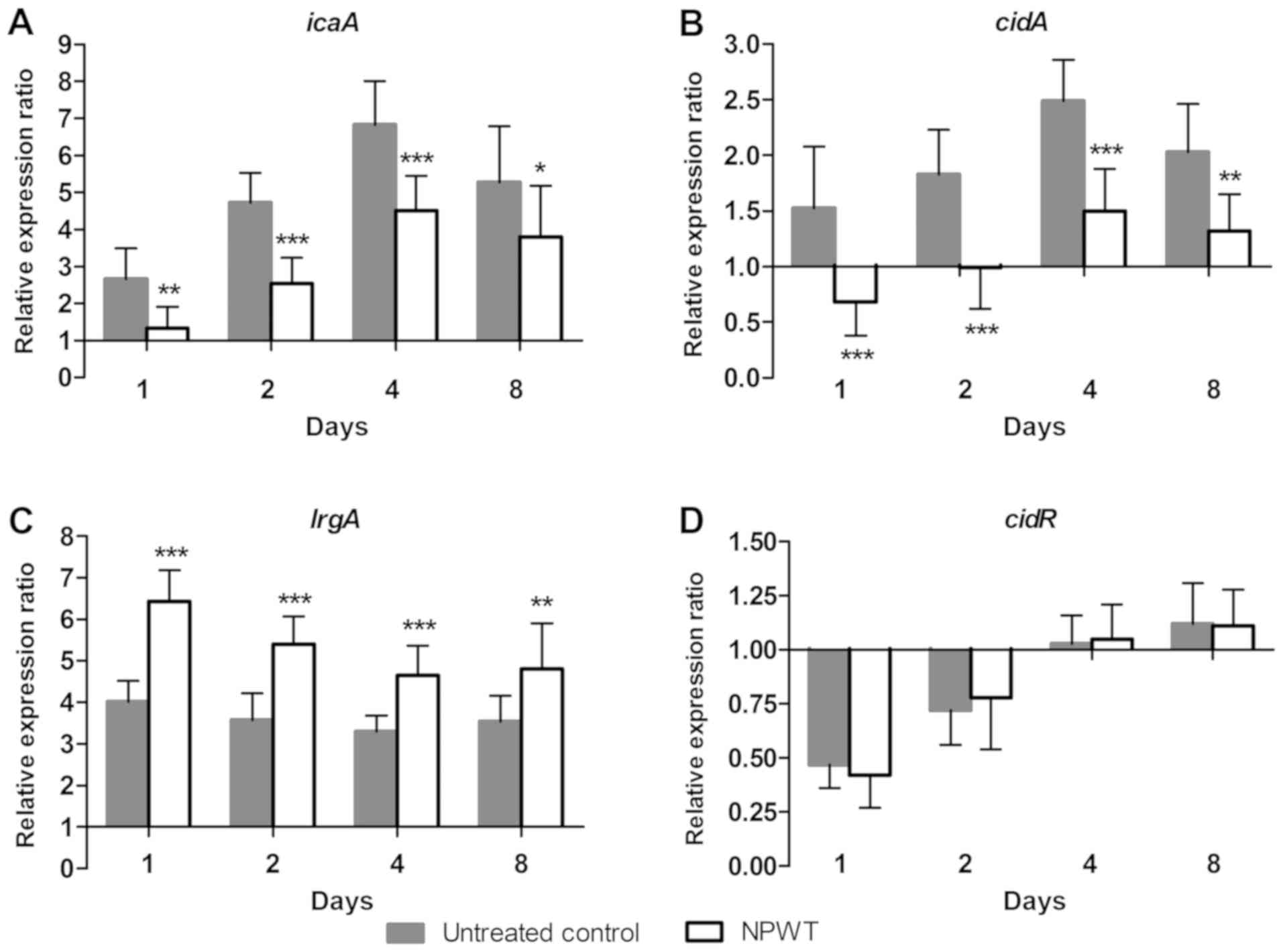
Effect of NPWT on the transcription of
S. aureus biofilm regulators
To investigate the mechanism of NPWT in affecting
the aggregation of S. aureus in tissue, the transcription of
biofilm regulators during the process of wound infection was
analyzed using RT-qPCR. The relative expression ratio of each gene
was expressed as the fold-change relative to the level found in the
inoculum. In the control group, a 2.6-fold increase for
poly-beta-1,6-N-acetyl-D-glucosamine synthase (icaA)
(Fig. 3A) was observed on post
operative day 1, after which the expression further increased and
reached a maximum of 6.8-fold on day 4. Expression then fell to
5.3-fold on day 8. Similar to icaA, the transcription of
holin-like protein CidA (cidA) (Fig. 3B) gradually increased and reached a
maximal 2.5-fold increase on day 4, then declined to 2.0-fold on
day 8. The expression of antiholin-like protein LrgA (lrgA)
(Fig. 3C) increased and remained
~4-fold higher compared with baseline levels over time. After an
initial decline on day 1 and 2, the expression of cidR
(Fig. 3D) returned to baseline
levels at day 4 and 8. In comparison, NPWT resulted in a
significant decrease in the expression of icaA and
cidA and a significant increase in lrgA levels
compared with that in the control group at days 1, 2, 4 and 8, but
did not have any significant effect on cidR expression
(Fig. 3).
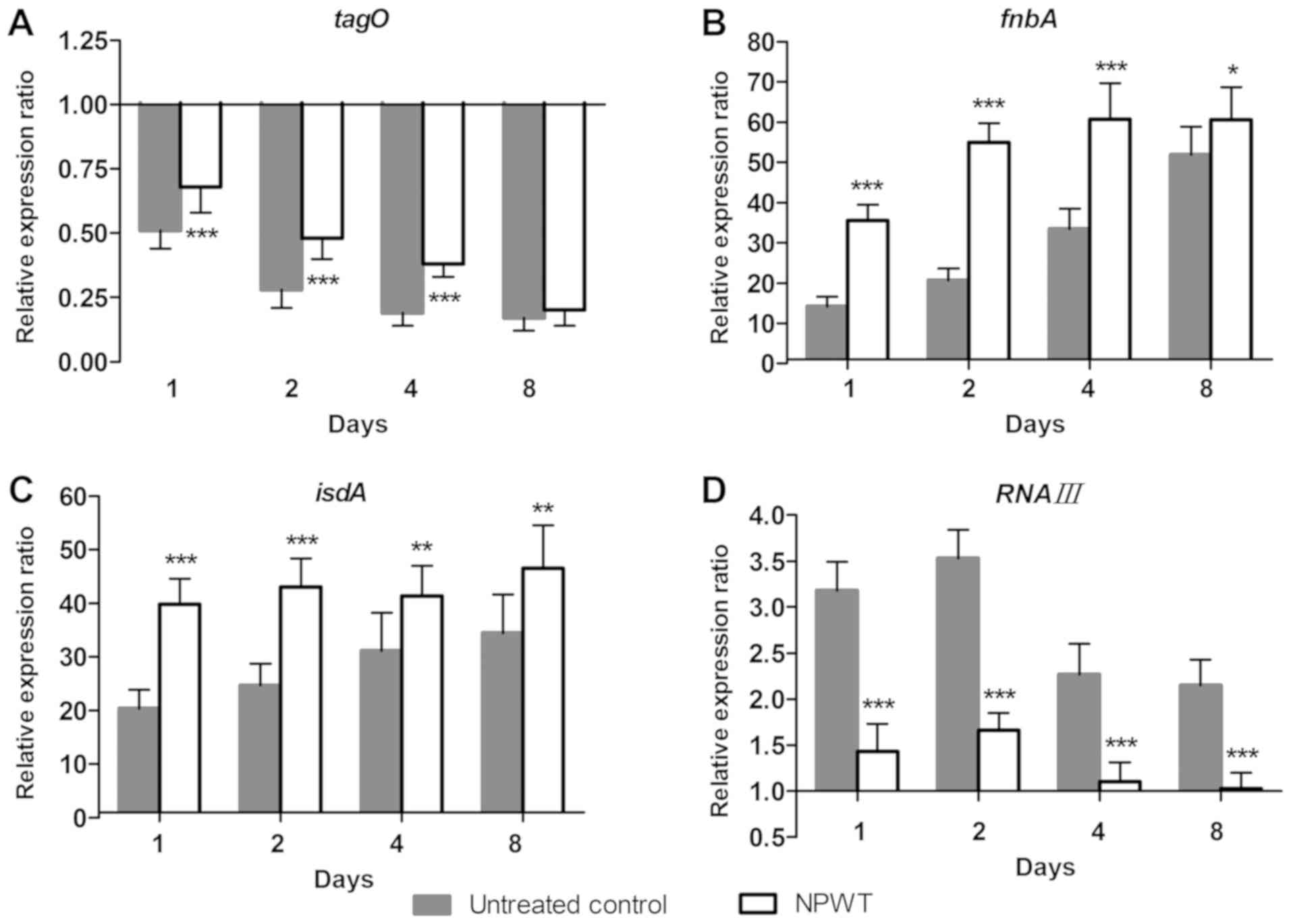
Changes in S
aureus colonization under NPWT treatment. To further
understand the living state of S. aureus in tissue, the adaptive
expression of bacterial genes associated with colonization during
wound infection was investigated. In the control group, the
expression of tagO (Fig. 4A)
continued to decrease in a time-dependent manner and reached 17% of
inoculum levels on day 8. Conversely, a 14-fold increase of fnbA
levels (Fig. 4B) was observed
compared with inoculum levels on day 1, after which the expression
gradually increased and reached 52-fold of baseline levels on day
8. The expression of isdA (Fig. 4C)
followed a similar pattern, with a 20-fold increase at day 1 and a
gradual increase to 34-fold at day 8. RNAIII expression (Fig. 4D) increased more slowly compared with
fnbA and isdA levels and reached a maximum of 3.5-fold increased
compared with baseline levels on day 2. Expression started to
decrease at days 4 and 8, but remained elevated with a 2.2-fold
increase above baseline levels on day 8. In contrast, NPWT led to
significant upregulation of tagO, fnbA and isdA levels, and
significant downregulation of RNAIII levels compared with the
controls at days 1, 2, 4 and 8 (Fig.
4).
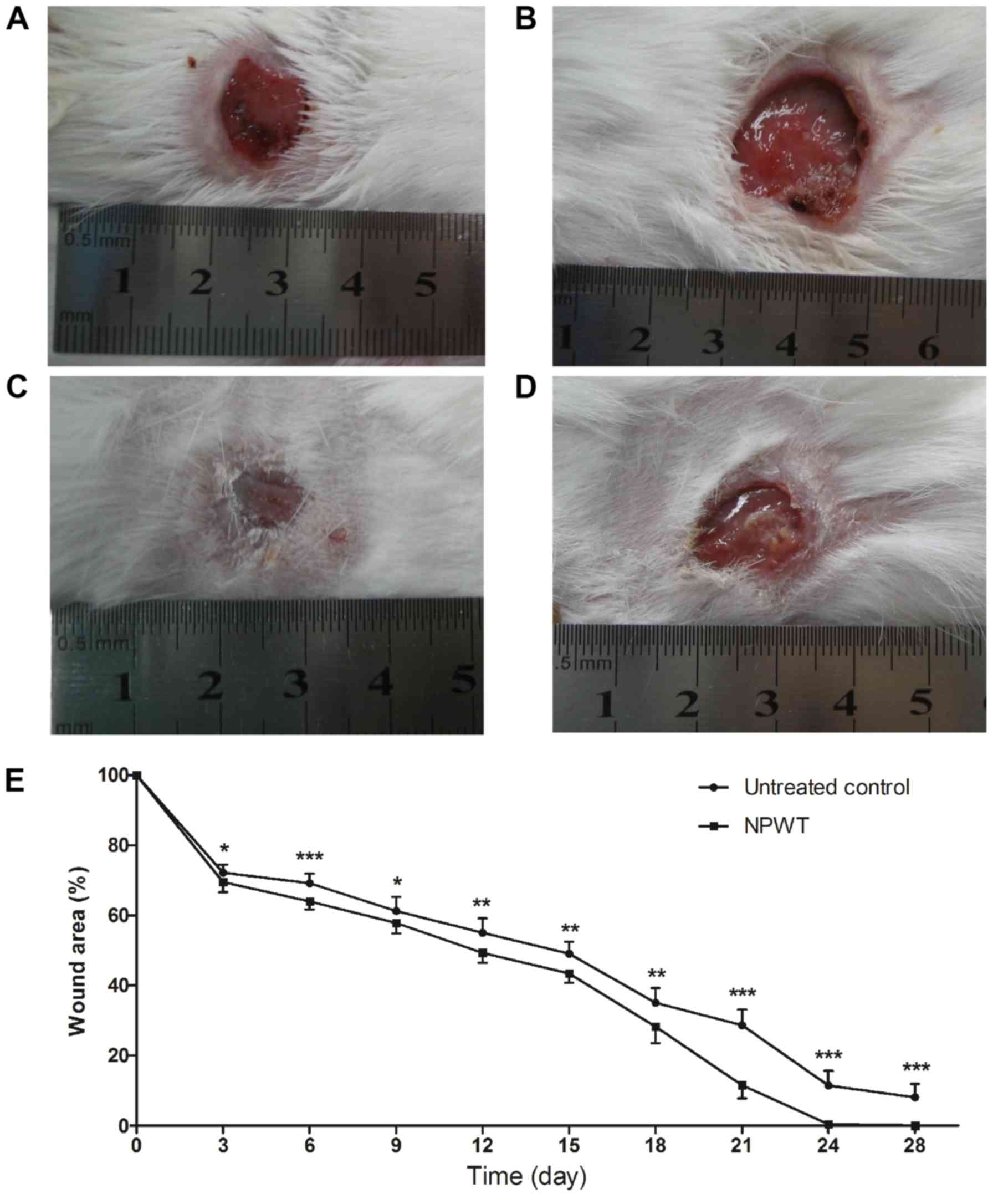
Wound healing condition
After 8 days of treatment, the wound healing
conditions of both groups were observed until postoperative day 28.
Gross appearance on the 18th day manifested with a small wound area
with clean granulation tissue bed and without necrosis or purulent
secretion in the NPWT group (Fig.
5A) compared with the control (Fig.
5B). On the 24th day, the wound treated with NPWT completely
healed (Fig. 5C), with a
neoepithelium covering the wound center. By contrast, a large skin
defect presented in the control group (Fig. 5D), with film-like exudates overlying
the wound bed. Although treatment was only administered for 8 days,
wound healing of the NPWT group was significantly improved compared
with the control group throughout time period (Fig. 5E).
Discussion
The pathogenesis of wound infection has long been
considered to be complex and multifactorial (31). Bacterial amount and invasiveness, as
well as the immune competencies of the host, affect the outcome of
wound infection (25,26). As the main cause behind wound healing
impairment, bacterial biofilm and quorum-sensing make the treatment
of infected wounds much more challenging (31-33).
With almost 1,000-fold higher antimicrobial resistance compared
with planktonic bacteria, biofilms develop rapidly and present
either at the surface or within the profound layers of wounds
(34). In an effort to identify
novel approaches to wound care and infection control, NPWT was
demonstrated to be safe and effective in avoiding
biofilm-associated infections as a physical therapy (14,16,35).
However, the mechanism involved remains to be elucidated,
particularly with regard to the response of bacteria secondary to
NPWT. It is well known that bacterial gene expression is highly
affected by the microenvironment (21). Transcriptomic research demonstrated
that quorum sensing is downregulated in infections in vivo
compared with in vitro conditions, suggesting bacterial
physiology differs in clinical infections relative to in
vitro experiments (36). The
present study investigated the distribution and existing form of
S. aureus in the local microenvironment established by NPWT
and monitored the adaptive expression of bacterial genes during
acute wound infection in vivo.
The results of the present study indicated that NPWT
had an obvious influence on the aggregation and distribution of
S. aureus in the tissue. In the present study, bacterial
aggregates were identified in the control group by histological
evaluation, along with distinct regions and micronecrosis under the
wound surface. This verified the persistence of bacteria within the
tissue for at least 8 days, mainly in the form of microcolonies
with extracellular matrix. However, NPWT led to a scattering
distribution of bacteria in wounds, accompanied by a healthier
granulation bed. Although the dispersive distribution of bacteria
is associated with active infection in some cases, wound healing
was improved by NPWT in the present study, which was also proven by
extensive clinical research (14,15,37).
To investigate the effect of NPWT on the survival of
S. aureus in wounds and the relationship between the
dispersive distribution and bacterial burden, viable bacteria
counts were measured. The present data showed a decrease in viable
bacteria in the NPWT group compared with the untreated control.
Nevertheless, the bacterial load was not dramatically reduced in
comparison with the control even at day 8, with ~1x106
CFUs/g, which was still a considerable amount of bacteria in the
tissue. This indicated that NPWT did not directly kill bacteria in
wounds and the dispersive distribution of S. aureus should
not be attributed to a low bioburden (38).
S. aureus responded to physical stimulations
by altering gene transcription (19-22).
To better understand the effects of NPWT on the living state of
S. aureus in wounds, the adaptive expression of bacterial
biofilm regulators during wound infection was studied.
Transcriptional analysis showed that there were significant
inhibitions on the expression of genes involved in the production
of biofilm components, including icaA and cidA.
Furthermore, the transcription of lrgA, which encodes an
antiholin-like protein with an inhibitory effect on murein
hydrolase activity (9), was
increased in the NPWT group. These results suggested that NPWT
inhibited the biosynthesis of the biofilm matrix, leading to a
free-living growth pattern for bacteria in the tissue. To some
extent, this may explain the dispersive distribution of S.
aureus in the present study and the low incidence of
biofilm-associated infections in the clinical use of NPWT.
To further explore the events involved in the shift
of phenotypic growth pattern, the expression of adhesion factors
associated with bacterial colonization was analyzed. The present
data showed that NPWT led to increased expression of the adhesion
factors, suggesting an intensive colonization of S. aureus
in tissues. The colonization state alone was reported to do little
harm to host cells and does not delay the wound healing process
(6,7). The life styles of S. aureus in
colonization or spreading invasive infection were demonstrated to
be regulated by the agr quorum-sensing system, a cell
density-dependent regulatory mechanism (11). To determine whether the discrepancy
between bacterial dispersive distribution and enhanced colonization
in the NPWT group could be ascribed to the low activity of quorum
sensing, the transcript analysis of RNAIII, the main
effector molecule of the agr system, was included in the
present study. The results showed that RNAIII transcript
levels were significantly reduced in the NPWT group compared with
the untreated control group, suggesting a decreased activity of the
agr system. A possible explanation for this decrease may be
that the bacterial density in NPWT-treated wounds was under the
threshold for agr activation, thus not leading to active and
invasive infection (11,32). Bacterial dispersive distribution and
enhanced colonization appears to be a more uniform scenario, which
reflects the commensalism of S. aureus in the local niche
built by NPWT (39). This suggested
that bacteria benefitted from the host but does little harm to the
wound repair and healing process.
However, the present study exhibited a number of
limitations. Although the data of bacterial gene expression lend
support to the distribution and existing form of S. aureus
in tissues, the exact mechanism involved remains to be elucidated.
As a physical therapy based on continuous suction, NPWT might
affect the pathogenesis of S. aureus infection by multiple
ways and means, including stereotaxic drainage, pressure variation
and shear stress (40,41). In particular, multiple sampling and
further analysis on wound healing was performed to reduce the error
caused by individual differences. Biopsies were carefully performed
to ensure that other analyses are not affected. Despite concerns
about healing and disruption of the bacterial state by repeated
sampling, the actual impact was marginal (42). Only the transcription of genes
related to biofilm formation and bacterial colonization was
investigated in the present study. As the process of infection
involves numerous gene regulatory networks, it is necessary to
study global gene expression to validate the present findings in
future work.
In summary, the present findings indicated that
S. aureus responds to NPWT by regulating gene expression,
manifesting a decrease in biofilm formation and an increase in
bacterial adhesion. The transition from bacterial biofilm to less
invasive colonization may reflect a commensal relationship between
S. aureus and host cells, which potentially benefits the
wound repair and healing process. Further studies into the
processes influencing the divergence between biofilm infection and
harmless colonization may aid in the discovery of novel
therapeutics against infections in the field of wound care.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81472112).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
TL, GW, LZ and PT performed the animal studies. GW,
PY and ZL performed histological experiments. TL, PY and ZL
performed bacterial counting and RT-qPCR experiments. TL, GW, PY
and PT analyzed and interpreted the data. TL and PT prepared the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Medical Ethics
Committee of the Chinese PLA General Hospital (approval no.
2014-X9-8) in compliance with the Guidelines for Care and Use of
Animals in Research (8th edition).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Marimuthu K, Eisenring MC, Harbarth S and
Troillet N: Epidemiology of Staphylococcus aureus surgical
site infections. Surg Infect (Larchmt). 17:229–235. 2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Heilmann C: Adhesion mechanisms of
staphylococci. Adv Exp Med Biol. 715:105–123. 2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Speziale P, Pietrocola G, Rindi S,
Provenzano M, Provenza G, Di Poto A, Visai L and Arciola CR:
Structural and functional role of Staphylococcus aureus
surface components recognizing adhesive matrix molecules of the
host. Future Microbiol. 4:1337–1352. 2009.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Weidenmaier C, Kokai-Kun JF, Kulauzovic E,
Kohler T, Thumm G, Stoll H, Götz F and Peschel A: Differential
roles of sortase-anchored surface proteins and wall teichoic acid
in Staphylococcus aureus nasal colonization. Int J Med
Microbiol. 298:505–513. 2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Burian M, Rautenberg M, Kohler T, Fritz M,
Krismer B, Unger C, Hoffmann WH, Peschel A, Wolz C and Goerke C:
Temporal expression of adhesion factors and activity of global
regulators during establishment of Staphylococcus aureus
nasal colonization. J Infect Dis. 201:1414–1421. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Edwards R and Harding KG: Bacteria and
wound healing. Curr Opin Infectious Dis. 17:91–96. 2004.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Poutahidis T, Kearney SM, Levkovich T, Qi
P, Varian BJ, Lakritz JR, Ibrahim YM, Chatzigiagkos A, Alm EJ and
Erdman SE: Microbial symbionts accelerate wound healing via the
neuropeptide hormone oxytocin. PLoS One. 8(e78898)2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Lee K, Lee JH, Ryu SY, Cho MH and Lee J:
Stilbenes reduce Staphylococcus aureus hemolysis, biofilm
formation, and virulence. Foodborne Pathog Dis. 11:710–717.
2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Bayles KW: The biological role of death
and lysis in biofilm development. Nat Rev Microbiol. 5:721–726.
2007.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Otto M: Staphylococcal infections:
Mechanisms of biofilm maturation and detachment as critical
determinants of pathogenicity. Annu Rev Med. 64:175–188.
2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Thoendel M, Kavanaugh JS, Flack CE and
Horswill AR: Peptide signaling in the staphylococci. Chem Rev.
111:117–151. 2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Schierle CF, De la Garza M, Mustoe TA and
Galiano RD: Staphylococcal biofilms impair wound healing by
delaying reepithelialization in a murine cutaneous wound model.
Wound Repair Regen. 17:354–359. 2009.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Roche ED, Renick PJ, Tetens SP, Ramsay SJ,
Daniels EQ and Carson DL: Increasing the presence of biofilm and
healing delay in a porcine model of MRSA-infected wounds. Wound
Repair Regen. 20:537–543. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Anagnostakos K and Mosser P: Negative
pressure wound therapy in the management of postoperative
infections after musculoskeletal tumour surgery. J Wound Care.
23:191–194, 196-197. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Hahn HM, Lee IJ, Woo KJ and Park BY:
Silver-impregnated negative-pressure wound therapy for the
treatment of lower-extremity open wounds: A prospective randomized
clinical study. Adv Skin Wound Care. 32:370–377. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Singh DP, Gowda AU, Chopra K, Tholen M,
Chang S, Mavrophilipos V, Semsarzadeh N, Rasko Y and Holton Iii L:
The effect of negative pressure wound therapy with antiseptic
instillation on biofilm formation in a porcine model of infected
spinal instrumentation. Wounds. 28:175–180. 2017.PubMed/NCBI
|
|
17
|
Glass GE, Murphy GRF and Nanchahal J: Does
negative-pressure wound therapy influence subjacent bacterial
growth? A systematic review. J Plast Reconstr Aesthet Surg.
70:1028–1037. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ćirković I, Jocić D, Božić DD, Djukić S,
Konstantinović N and Radak D: The effect of vacuum-assisted closure
therapy on methicillin-resistant Staphylococcus aureus wound
biofilms. Adv Skin Wound Care. 31:361–364. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Islam N, Kim Y, Ross JM and Marten MR:
Proteomic analysis of Staphylococcus aureus biofilm cells
grown under physiologically relevant fluid shear stress conditions.
Proteome Sci. 12(21)2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Castro SL, Nelman-Gonzalez M, Nickerson CA
and Ott CM: Induction of attachment-independent biofilm formation
and repression of Hfq expression by low-fluid-shear culture of
Staphylococcus aureus. Appl Environ Microbiol. 77:6368–6378.
2011.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rosado H, Doyle M, Hinds J and Taylor PW:
Low-shear modelled microgravity alters expression of virulence
determinants of Staphylococcus aureus. Acta Astronautica.
66:408–413. 2010.
|
|
22
|
Li T, Wang G, Yin P, Li Z, Zhang L, Liu J,
Li M, Zhang L, Han L and Tang P: Effect of negative pressure on
growth, secretion and biofilm formation of Staphylococcus
aureus. Antonie van Leeuwenhoek. 108:907–917. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Institute of Laboratory Animal Resources
(US). Committee on Care, Use of Laboratory Animals, and National
Institutes of Health (US). Division of Research Resources: Guide
for the care and use of laboratory animals. 8th edition. National
Academies Press, Washington, DC, 2011.
|
|
24
|
Seth AK, Geringer MR, Nguyen KT, Agnew SP,
Dumanian Z, Galiano RD, Leung KP, Mustoe TA and Hong SJ:
Bacteriophage therapy for Staphylococcus aureus
biofilm-infected wounds: A new approach to chronic wound care.
Plast Reconstr Surg. 131:225–234. 2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Liu D, Zhang L, Li T, Wang G, Du H, Hou H,
Han L and Tang P: Negative-pressure wound therapy enhances local
inflammatory responses in acute infected soft-tissue wound. Cell
Biochem Biophys. 70:539–547. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Liu D, Li Z, Wang G, Li T, Zhang L and
Tang P: Virulence analysis of Staphylococcus aureus in a
rabbit model of infected full-thickness wound under negative
pressure wound therapy. Antonie van Leeuwenhoek. 111:161–170.
2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lalliss SJ, Stinner DJ, Waterman SM,
Branstetter JG, Masini BD and Wenke JC: Negative pressure wound
therapy reduces pseudomonas wound contamination more than
Staphylococcus aureus. J Orthop Trauma. 24:598–602.
2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kamamoto F, Lima ALM, Rezende MR,
Mattar-Junior R, Leonhardt MC, Kojima KE and Santos CCD: A new
low-cost negative-pressure wound therapy versus a commercially
available therapy device widely used to treat complex traumatic
injuries: A prospective, randomized, non-inferiority trial. Clinics
(Sao Paulo). 72:737–742. 2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Morykwas MJ, Argenta LC, Shelton-Brown EI
and McGuirt W: Vacuum-assisted closure: A new method for wound
control and treatment: Animal studies and basic foundation. Ann
Plast Surg. 38:553–562. 1997.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCt method. Methods. 25:402–408. 2001.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Omar A, Wright JB, Schultz G, Burrell R
and Nadworny P: Microbial Biofilms and Chronic Wounds.
Microorganisms. 5(pii:E9)2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Swanson EA, Freeman LJ, Seleem MN and
Snyder PW: Biofilm-infected wounds in a dog. J Am Vet Med Assoc.
244:699–707. 2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Boles BR and Horswill AR: Staphylococcal
biofilm disassembly. Trends Microbiol. 19:449–455. 2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Licker M, Moldovan R, Hogea E, Muntean D,
Horhat F, Bădiţoiu L, Rogobete A, Tirziu E and Zambori C: Microbial
biofilm in human health-An updated theoretical and practical
insight. Romanian J Laboratory Med. 25:9–26. 2017.
|
|
35
|
Li T, Zhang L, Han LI, Wang G, Yin P, Li
Z, Zhang L, Guo Q, Liu D and Tang P: Early application of negative
pressure wound therapy to acute wounds contaminated with
Staphylococcus aureus: An effective approach to preventing
biofilm formation. Exp Ther Med. 11:769–776. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Cornforth DM, Dees JL, Ibberson CB, Huse
HK, Mathiesen IH, Kirketerp-Møller K, Wolcott RD, Rumbaugh KP,
Bjarnsholt T and Whiteley M: Pseudomonas aeruginosa transcriptome
during human infection 115: E5125-E5134, 2018.
|
|
37
|
Copeland H, Newcombe J, Yamin F, Bhajri K,
Mille VA, Hasaniya N, Bailey L and Razzouk AJ: Role of negative
pressure wound care and hyperbaric oxygen therapy for sternal wound
infections after pediatric cardiac surgery. World J Pediatr
Congenit Heart Surg. 9:440–445. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Tahir S, Malone M, Hu H, Deva A and
Vickery K: The Effect of negative pressure wound therapy with and
without instillation on mature biofilms in vitro. Materials
(Basel). 11(pii: E811)2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Song J, Lays C, Vandenesch F, Benito Y,
Bes M, Chu Y, Lina G, Romby P, Geissmann T and Boisset S: The
expression of small regulatory RNAs in clinical samples reflects
the different life styles of Staphylococcus aureus in
colonization vs. infection. PLoS One. 7(e37294)2012.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Huang C, Leavitt T, Bayer LR and Orgill
DP: Effect of negative pressure wound therapy on wound healing.
Curr Probl Surg. 51:301–331. 2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Nie B and Yue B: Biological effects and
clinical application of negative pressure wound therapy: A review.
J Wound Care. 25:617–626. 2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Patil PS, Evancho-Chapman MM, Li H, Huang
H, George RL, Shriver LP and Leipzig ND: Fluorinated methacrylamide
chitosan hydrogel dressings enhance healing in an acute porcine
wound model. PLoS One. 13:e0203371. 2018.PubMed/NCBI View Article : Google Scholar
|