Introduction
Breast cancer is one of the most common malignant
tumor types and is also one of the leading causes of
cancer-associated mortality worldwide (1). Over the past few decades, surgical
treatment, radiotherapy, chemotherapy and endocrine therapy have
been utilized for the clinical treatment of breast cancer. However,
the prognosis for patients with breast cancer remains poor due to
the recurrence and metastasis of breast cancer following
conventional treatment (2).
Bioactive peptides, which are composed of several
amino acids, have been indicated to be effective as a novel
therapeutic strategy in cancer therapies (3). Previous studies have revealed that a
novel anti-tumor peptide, SA12, inhibits proliferation and arrests
the cell cycle in MDA-MB-231 and MCF-7 breast cancer cells
(4,5). Tumor metastasis is critical in the
progression of breast cancer, which is regulated by a number of
genes. Tumor metastasis suppressor genes, including cadherin 1
(CDH1), non-metastasis 23-H1 (nm23-H1) and breast cancer metastasis
suppressor 1 (BRMS1) are important negative regulators during tumor
metastasis (6,7). E-cadherin, which is the gene encoding
the product of CDH1, is downregulated in a number of cancer types
during the epithelial-mesenchymal transition and its expression is
inversely correlated with metastasis. Overexpression of E-cadherin
suppresses the invasion of cancer cells (6). Non-metastatic 23A (NM23A), a potential
metastasis suppressor encoded by the nm23-H1 gene, was identified
as a metastasis suppressor in melanoma cells. NM23A expression has
been identified to be inversely correlated with clinical metastasis
of multiple tumors, mediating suppression of metastasis and
controlling cellular responses to the microenvironment (7). BRMS1 is currently the only metastasis
suppressor that has been identified in models of breast cancer and
has since been revealed to suppress metastasis in a broad spectrum
of cancer types, regulating the interactions between tumor cells
and the tumor microenvironment (6).
The present study aimed to explore whether the anti-cancer peptide
SA12 inhibited the metastasis of MDA-MB-231 and MCF-7 breast cancer
cells and to further investigate the roles of the tumor metastasis
suppressor genes, CDH1, nm23-H1 and BRMS1, as well as their gene
encoding products, E-cadherin, NM23A and BRMS1, in the inhibition
of SA12 in MDA-MB-231 and MCF-7 cells.
Materials and methods
Reagents
DMEM and FBS were purchased from Gibco; Thermo
Fisher Scientific, Inc. Bovine serum albumin and trypsin were
purchased from Invitrogen; Thermo Fisher Scientific, Inc. Cell
culture plates and Transwell chambers were purchased from Corning
Inc. RNAiso Plus reagent, PrimeScript RT reagent kit and SYBR
Premix Ex Taq II were purchased from Takara Biotechnology Co., Ltd.
RIPA lysis buffer was purchased from Sangon Biotech Co., Ltd. The
mouse anti-human E-cadherin monoclonal antibody (cat. no. ab1416),
the rabbit anti-human NM23A polyclonal antibody (cat. no. ab92327)
and the rabbit anti-human BRMS1 monoclonal antibody (cat. no.
ab134968) were purchased from Abcam. The mouse anti-human β-actin
monoclonal antibody (cat. no. TA328071), the horseradish
(HRP)-labelled goat anti-mouse secondary antibody (cat. no.
TA130003) and the HRP-labelled goat anti-rabbit secondary antibody
(cat. no. TA140003) were purchased from OriGene Technologies,
Inc.
Cell lines and culture
Human MDA-MB-231 and MCF-7 cell lines were purchased
from the American Type Culture Collection and cultured in DMEM
medium containing 10% FBS at 37˚C in an atmosphere of 5%
CO2.
Peptide preparation
The amino acid sequence of SA12 used in the present
study was Ser-Val-Pro-Leu-Phe-Asn-Phe-Ser-Val-Tyr-Leu-Ala. The
peptide was chemically synthesized and purified by GL Biochem
Shanghai, Ltd. The synthetic peptide was dissolved in DMSO using
ultrasound and vortexed for 30 sec at 200 x g, and then diluted to
100 µM in DMEM.
Wound healing assay
MDA-MB-231 and MCF-7 cells were seeded into 6-well
cell culture plates with a density of 5x105 cells/well
at 37˚C overnight. Upon reaching 80% confluence, the cells were
scratched using a micropipette tip and the floating cells were
removed through PBS solution washes. The cells were treated with
100 µM SA12 diluted in serum-free DMEM, while the serum-free DMEM
containing 0.1% DMSO was used as the blank control. Cells were
incubated at 37˚C for 48 h without changing the medium, and wound
healing was examined using a phase contrast microscope (Olympus
Corporation) at x200 magnification. The cell migration distance was
measured and the migration distance at 48 h was normalized to the
distance at 0 h in each sample.
Transwell assay
A total of 500 µl serum-free DMEM containing 50 µl
bovine serum albumin (10 g/l) was added to each Transwell chamber
and incubated at 37˚C for 30 min to hydrate the basement membrane.
MDA-MB-231 and MCF-7 cells were cultured until 90% confluent at
37˚C, and the serum was removed to starve the cells for 12 h. The
cells were then digested with trypsin and centrifuged at 200 x g
for 5 min at room temperature. The cell pellets were resuspended in
serum-free medium at a cell density of 5x105 cells/ml.
The Transwell chambers were placed in the 24-well cell culture
plates with 500 µl DMEM per well and 200 µl prepared cell
suspension was added into the Transwell chamber. The cells were
treated with 100 µM SA12 diluted in DMEM and the control group was
treated with serum-free cell culture medium containing 0.1% DMSO.
In order to prevent cell proliferation caused by the serum in the
lower chambers, the cells were collected following incubation for
24 h at 37˚C, without medium changes. The cells were fixed with 2%
glutaraldehyde for 30 min and stained with 0.2% gentian violet
(Baso Diagnostic, Inc.) for 10 min at temperature. The migrated
cells were photographed using a phase contrast microscope (Olympus
Corporation) at x200 magnification. The number of migrated cells in
5 visual fields was randomly counted using ImageJ 1.8.0 software
(National Institutes of Health) and the average number was
calculated to determine the cell mobility in cells following SA12
treatment.
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
The MDA-MB-231 and MCF-7 breast cancer cell lines
were seeded into 6-well plates at a density of 5x105
cells/well and cultured at 37˚C. Cells were treated with 100 µM
SA12 diluted in DMEM for 48 h at 37˚C, and cells were treated with
DMEM containing 0.1% DMSO as a control for 48 h at 37˚C. RNAiso
plus reagent (Takara Bio, Inc.) was used to extract total RNA and
subsequently, the obtained RNA was dried naturally at room
temperature and dissolved in 50 µl RNase-free water. PrimeScript RT
reagent kit (Takara Bio, Inc.) was used to reverse transcribe mRNA
to generate cDNA at 37˚C for 15 min and 85˚C for 5 sec. CDH1,
nm23-H1 and BRMS1 cDNA were amplified using SYBR Premix Ex Taq II
(Takara Bio, Inc.), with primers provided in Table I. β-actin was used as the internal
control. The following thermocycling conditions were used for the
qPCR: Initial denaturation at 95˚C for 3 min; 40 cycles of 95˚C for
15 sec, 54˚C for 20 sec and 72˚C for 20 sec; and a final extension
at 72˚C for 10 min. The relative mRNA expression levels were
calculated as fold changes using the
2-ΔΔCq method (8) to reflect the changes of CDH1, nm23-H1
and BRMS1 genes following treatment with SA12.
 | Table IPrimers designed for reverse
transcription-quantitative PCR. |
Table I
Primers designed for reverse
transcription-quantitative PCR.
| Gene | Primer |
|---|
| CDH1 | F:
5'-GAACGCATTGCCACATACAC-3' |
| R:
5'-GAATTCGGGCTTGTTGTCAT-3' |
| nm23-H1 | F:
5'-AGAAAGGATTCCGCCTTGTT-3' |
| R:
5'-GGCCCTGAGTGCATGTATTT-3' |
| BRMS1 | F:
5'-GCAGCGGAGCCTCAAGATTCG-3' |
| R:
5'-GCAGCGTGTCATAGAGCAGCAG-3' |
| β-actin | F:
5'-GACTTAGTTGCGTTACACCCTTTC-3' |
| R:
5'-TGCTGTCACCTTCACCGTTC-3' |
Western blot analysis
The MDA-MB-231 and MCF-7 breast cancer cell lines
were seeded into 6-well plates at a density of 5x105
cells/well and treated with 100 µM SA12 diluted in DMEM for 48 h at
37˚C. Cells treated with DMEM containing 0.1% DMSO were used as the
control. The cells were harvested following digestion with trypsin,
centrifuged at 200 x g for 5 min at room temperature, lysed with
200 µl RIPA lysis buffer (Beyotime Institute of Biotechnology)
containing 2 µl PMSF (100 mM) (Beyotime Institute of
Biotechnology), and incubated for 30 min on ice. The lysate was
centrifuged at 14,000 x g for 15 min at 4˚C, the supernatant was
extracted, and the concentration of total protein was calculated
using a bicinchoninic acid assay (Beyotime Institute of
Biotechnology). Subsequently, 80 µg protein/lane was separated via
12% SDS-PAGE (Beyotime Institute of Biotechnology). Subsequently,
the proteins were transferred onto nitrocellulose membranes and
blocked with 5% skimmed milk for 30 min at room temperature. The
membranes were then incubated with E-cadherin (1:50), NM23A
(1:1,000) and BRMS1 (1:1,000) primary antibodies overnight at 4˚C,
and subsequently with the goat anti-mouse or goat anti-rabbit
HRP-labeled secondary antibodies (1:2,000) for 2 h at room
temperature. The protein signals were visualized using ECL (EMD
Millipore). The density of bands in SA12 treated cells and controls
were compared and normalized to β-actin (1:1,000), which was used
as the internal control. The relative protein expression levels of
E-cadherin, NM23A and BRMS1 after SA12 treatment were calculated
using Gel-Pro Analyzer 4.0 software (Media Cybernetics, Inc.).
Statistical analysis
All the experiments were repeated at least 3 times
and the results are presented as the mean ± standard deviation.
Data were analyzed using unpaired or paired Student's t-tests.
P<0.05 was considered to indicate a statistically significant
difference. SPSS v14.0 software (SPSS, Inc.) was used for
statistical analysis.
Results
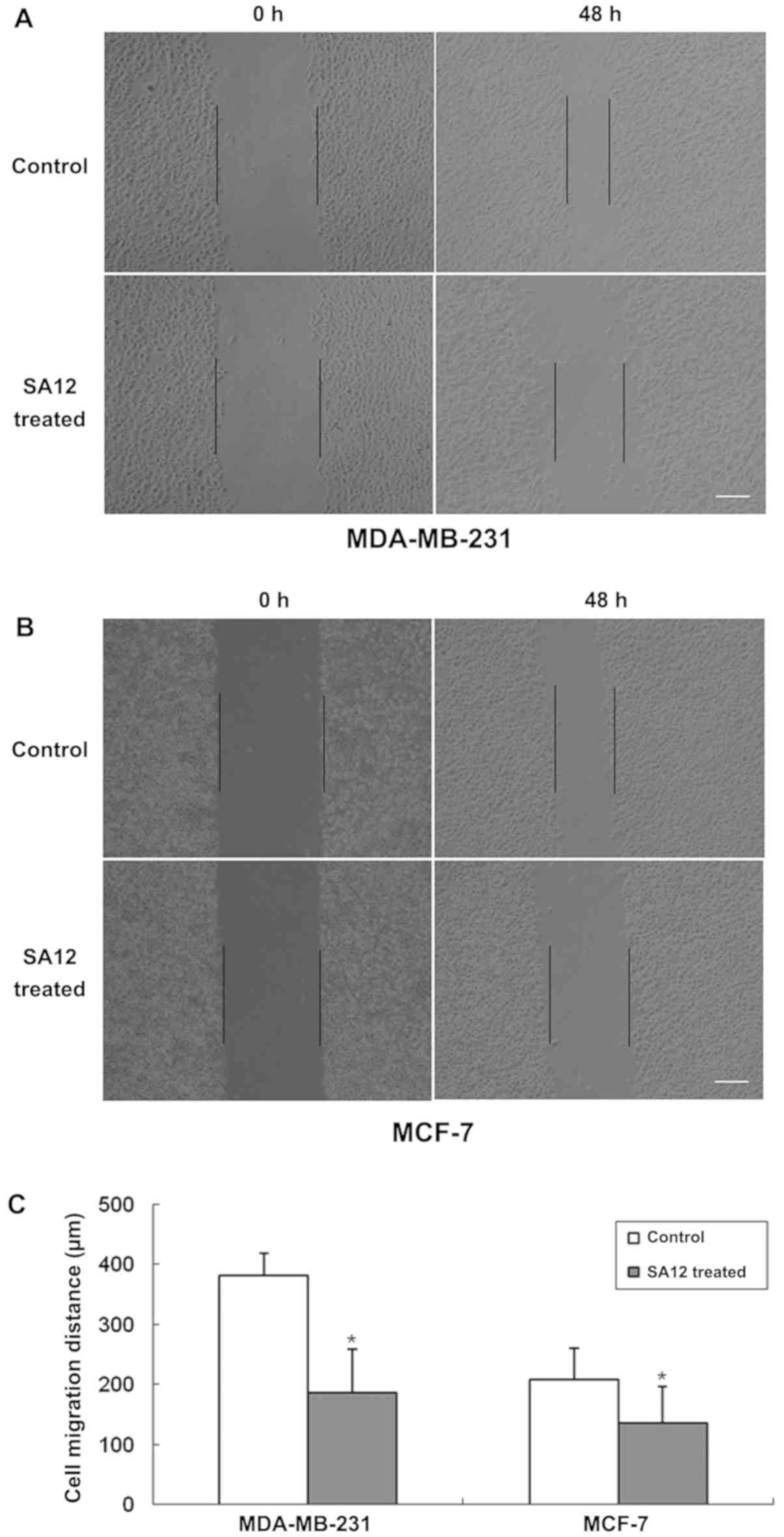
SA12 inhibits the migration of
MDA-MB-231 and MCF-7 cells in wound healing assays
To examine the inhibitory effect of SA12 on the
migration of MDA-MB-231 and MCF-7 breast cancer cells, wound
healing assays were performed. The cells were treated with 100 µM
SA12, while the control group was treated with 0.1% DMSO. The cells
in each group were cultured for 48 h and images were obtained. The
results revealed that SA12 inhibited the migration of MDA-MB-231
and MCF-7 breast cancer cells, and the cell migration distances
were reduced compared with the control group (P<0.05; Fig. 1). The percentage of migrated cells
was 51.3 and 34.6% in MDA-MB-231 and MCF-7 cells, compared with the
control group, respectively (data not shown).
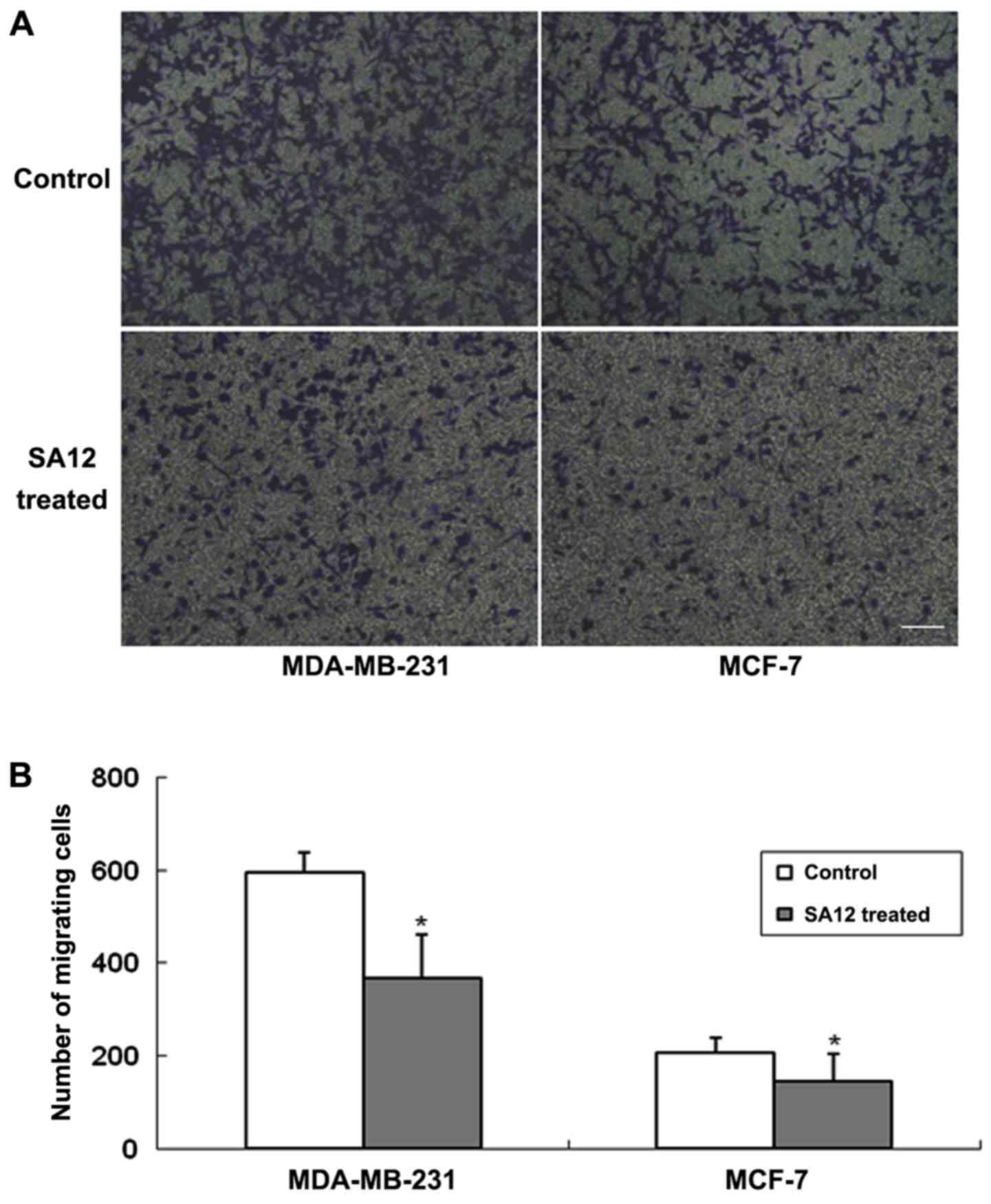
SA12 inhibits the migration of
MDA-MB-231 and MCF-7 cells in Transwell assays
To further determine the inhibitory effect of SA12
on the migration of MDA-MB-231 and MCF-7 breast cancer cells,
Transwell assays were performed. The cells were treated with 100 µM
SA12, with 0.1% DMSO-treated cells used as the control. The cells
were harvested following 24 h incubation and the number of cells
migrated from the basement membrane of the upper chambers in 5
visual fields was counted and the average number was calculated.
The results revealed that SA12 significantly inhibited the
migration of MDA-MB-231 and MCF-7 breast cancer cells following 24
h of treatment, and the number of migrated cells were decreased
compared with the control groups (P<0.05; Fig. 2). The percentage of migrated cells
was 38.2 and 29.8% in MDA-MB-231 and MCF-7 cells, respectively,
compared with the controls (data not shown). These results revealed
that SA12 inhibited the migration of breast cancer MDA-MB-231 and
MCF-7 cells.
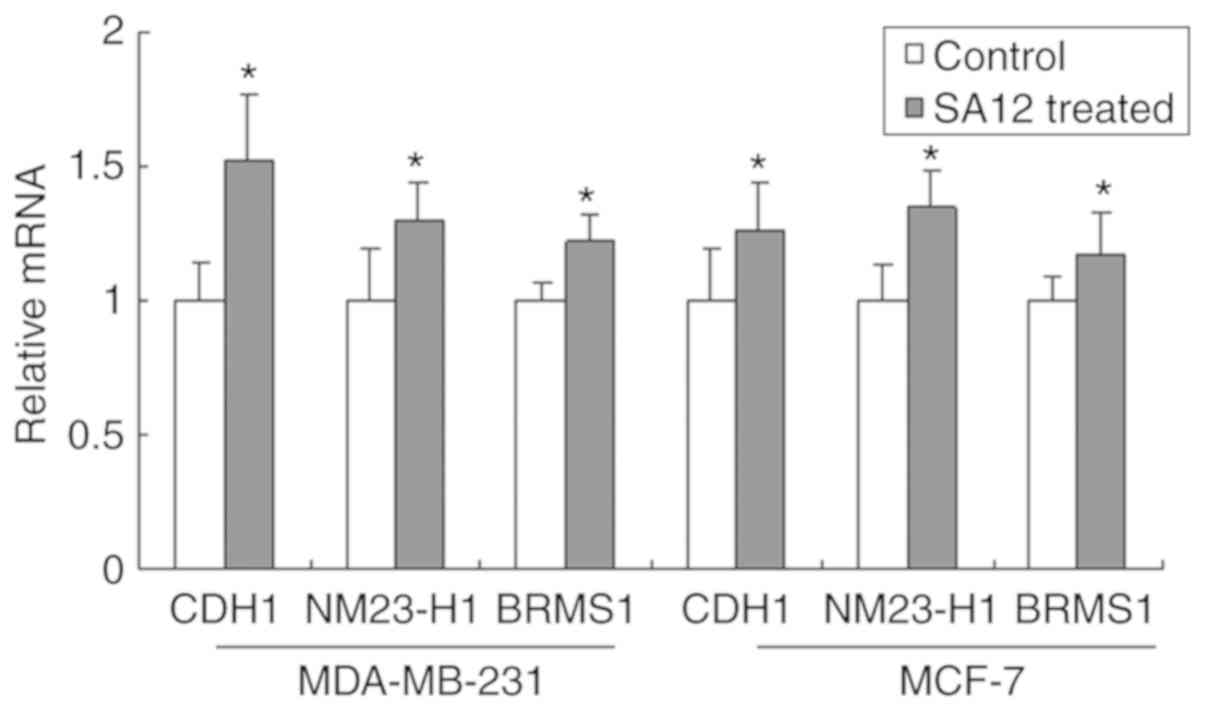
SA12 increases the mRNA expression
levels of CDH1, nm23-H1 and BRMS1
The tumor metastasis suppressor genes, CDH1, nm23-H1
and BRMS1 are important negative regulators in tumor metastasis
(6). To investigate the possible
mechanism of action behind how SA12 inhibited the metastasis of
MDA-MB-231 and MCF-7 breast cancer cells, the mRNA expression
levels of CDH1, nm23-H1 and BRMS1 were detected using RT-qPCR. As
presented in Fig. 3, the mRNA
expression levels of CDH1, nm23-H1 and BRMS1 were significantly
increased after treatment with 100 µM SA12 for 48 h (all
P<0.05). The percentage increase in the mRNA levels of the
aforementioned markers in MDA-MB cells compared with that in the
controls was 51.9, 30.2 and 22.3%, respectively, and 26.1, 34.4 and
17.2%, respectively, in MCF-7 cells. These results suggested that
treatment with SA12 significantly increased the mRNA expression
levels of CDH1, nm23-H1 and BRMS1 in MDA-MB-231 and MCF-7 breast
cancer cells.
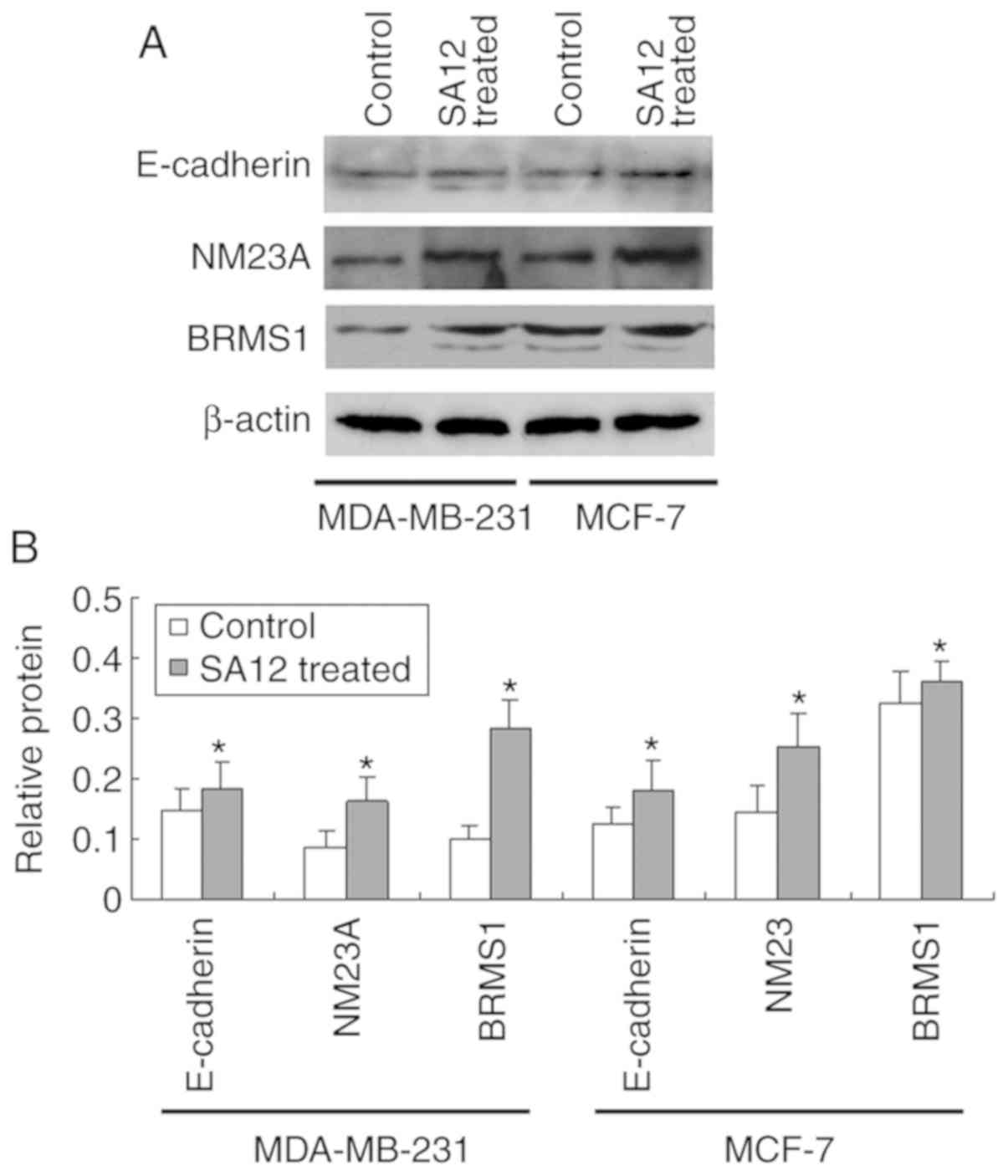
SA12 enhances the protein expression
levels of E-cadherin, NM23A and BRMS1
To further verify the aforementioned results, the
effect of SA12 on the protein expression levels of tumor metastasis
suppressors E-cadherin, NM23A and BRMS1 in MDA-MB-231 and MCF-7
breast cancer cells were examined using western blot analysis. As
presented in Fig. 4, following
treatment with 100 µM SA12 for 48 h, the protein expression levels
of E-cadherin, NM23A and BRMS1 in MDA-MB-231 and MCF-7 cells was
significantly increased (all P<0.05). The percentage increase in
the aforementioned protein levels in MDA-MB-231 cells compared with
that in the controls was 42.1, 76.3 and 41.9%, respectively, and
25.2, 32.8 and 23.0%, respectively, in MCF-7 cells. The results
indicated that the protein expression levels of E-cadherin, NM23A
and BRMS1 were also increased following SA12 treatment. SA12
increased the expression of the tumor metastasis suppressors
E-cadherin, NM23A and BRMS1 in MDA-MB-231 and MCF-7 cells. The
upregulation of E-cadherin, NM23A and BRMS1 may be responsible for
the SA12-induced inhibition of metastasis to the MDA-MB-231 and
MCF-7 cells.
Discussion
Malignant tumor metastasis is a complex biological
process, involving a number of cellular interactions between tumor
and stroma cells. Metastasis occurs when tumor cells break away
from the primary tumor and migrate through the extracellular matrix
(9). Tumor metastasis suppressor
genes, including CDH1, nm23-H1 and BRMS1 are important negative
regulators, which serve important roles in inhibiting tumor
metastasis (6,7,10). A
previous study identified an anti-tumor peptide SA12, which was
indicated to inhibit proliferation and arrest the cell cycle of
MDA-MB-231 and MCF-7 breast cancer cells (4,5). In the
present study, the anti-tumor peptide SA12 inhibited the migration
of MDA-MB-231 and MCF-7 breast cancer cells and enhanced the
expression of the tumor metastasis suppressor genes, CDH1, nm23-H1
and BRMS1. These genes were also demonstrated to be associated with
the SA12-induced inhibition of breast cancer cell migration.
Breast cancer metastasis is one of the leading
causes of mortality in women and is regulated by adhesion molecules
such as the cadherin superfamily (11). E-cadherin, which is the gene encoding
product of CDH1, is an important member of the cadherin
superfamily, which maintains the balance of cell-cell and
cell-extracellular matrix adhesion (12). E-cadherin is a transmembrane
glycoprotein expressed in human epithelial tissues, which connects
epithelial cells together and serve a role in cell adhesion, cell
morphology and cell movement (13).
Intercellular adhesion serves an important role in the invasion and
metastasis of tumor cells (14).
Dysregulation of E-cadherin in tumors is associated with the
epithelial-mesenchymal transition and subsequently induces tumor
cell dissemination as well as promoting cell migration and invasion
(15). E-cadherin supports the
function of cell adhesion molecules in the repression of tumor
metastasis in breast cancer (11,16).
Studies from a meta-analysis revealed that reduced E-cadherin
expression levels are associated with a poorer prognosis in breast
cancer tissue; therefore, may be a potential therapeutic target for
treating breast cancer (17). In the
present study, the anti-tumor peptide SA12 was indicated to enhance
the expression of the tumor metastasis suppressor gene CDH1 in
MDA-MB-231 and MCF-7 cells. These results are consistent with a
previous report that revealed that E-cadherin influences the
imbalance of homogeneous adhesion between tumor cells and
heterogeneous adhesion between tumor cells and stromal cells
(18). This imbalance is crucial for
the detachment of tumor cells from the primary tumor and for the
invasion or metastasis of independent tumor cells (19-21).
Furthermore, the present study revealed that SA12
treatment increased the expression of nm23-H1 and BRMS1 in
MDA-MB-231 and MCF-7 cells, which are also tumor metastasis
suppressor genes. NM23A, which is a potential metastasis suppressor
encoded by the nm23-H1 gene (22,23), was
indicated to be highly expressed in differentiated tissues, but at
lower levels in breast cancer, gastric cancer, bladder cancer,
intestinal cancer and osteosarcoma tissues (24,25). The
expression levels of nm23-H1 are negatively correlated with lymph
node metastasis and reflect the metastatic ability of tumors
(26-28).
Phosphorylation of kinase suppressor of Ras1 (KSR1)-serine392 by
NM23A is known to reduce ERK activation and inhibit the ERK-MAPK
signaling pathway, which serves a crucial role in cancer metastasis
(29). Another important tumor
metastasis suppressor gene, BRMS1, which was discovered in breast
carcinoma cells, has since been identified to suppress metastasis
in a variety of cancer types (30).
A previous study demonstrated that BRMS1 regulates the interactions
between tumor cells and the tumor microenvironment, affecting the
key events of metastasis, including the inhibition of migration and
invasion, as well as the initiation of growth by single cells and
promotion of anoikis (31).
Increased expression of BRMS1 restores the homotypic and
heterotypic intercellular communication of gap junctions (30). Additionally, a role for BRMS1 in
transcriptional regulation has also been revealed since the
identification of BRMS1 in the mammalian Sin3 histone deacetylase
complex (30,31).
In conclusion, the anti-tumor peptide SA12 was
revealed to inhibit the metastasis of MDA-MB-231 and MCF-7 breast
cancer cells in the current study. SA12 enhanced the expression of
the tumor metastasis suppressor genes, CDH1, nm23-H1 and BRMS1, and
maybe responsible for SA12-induced inhibition of metastasis in
MDA-MB-231 and MCF-7 cells. To more conclusively demonstrate that
CDH1, nm-23-H1 and BRMS1 are involved in this suppression,
knockdown experiments should be performed, which is a limitation of
the present study and a future direction for research. A more
in-depth study should focus on the detailed molecular mechanisms of
action behind the reduction in angiogenesis induced by SA12.Further
studies to reveal the molecular mechanisms of action underlying the
SA12-induced inhibition of breast cancer cell metastasis and the
involvement of tumor metastasis suppressor genes in the cell signal
pathways are required to expand on the findings of the present
study.
Acknowledgements
Not applicable.
Funding
This study was supported by the Natural Science
Foundation of China (grant no. 81502671).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LY designed the study, acquired the data and drafted
the manuscript; FL performed the statistical analysis; ZG processed
the data; XC analyzed the results; KD and HZ interpreted the data
and revised the manuscript critically for important intellectual
content. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jamdade VS, Sethi N, Mundhe NA, Kumar P,
Lahkar M and Sinha N: Therapeutic targets of triple-negative breast
cancer: A review. Br J Pharmacol. 172:4228–4237. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Nagini S: Breast cancer: Current molecular
therapeutic targets and new players. Anticancer Agents Med Chem.
17:152–163. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Li ZJ and Cho CH: Development of peptides
as potential drugs for cancer therapy. Curr Pharm Des.
16:1180–1189. 2010.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Yang L, Cui Y, Shen J, Lin F, Wang X, Long
M, Wei J and Zhang H: Antitumor activity of SA12, a novel peptide,
on SKBr-3 breast cancer cells via the mitochondrial apoptosis
pathway. Drug Des Devel Ther. 9:1319–1330. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yang L, Liu H, Long M, Wang X, Lin F, Gao
Z and Zhang H: Peptide SA12 inhibits proliferation of breast cancer
cell lines MCF-7 and MDA-MB-231 through G0/G1 phase cell cycle
arrest. OncoTargets Ther. 11:2409–2417. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Bodenstine TM and Welch DR: Metastasis
suppressors and the tumor microenvironment. Cancer Microenviron.
1:1–11. 2008.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bohl CR, Harihar S, Denning WL, Sharma R
and Welch DR: Metastasis suppressors in breast cancers: Mechanistic
insights and clinical potential. J Mol Med (Berl). 92:13–30.
2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Robert J: Biology of cancer metastasis.
Bull Cancer. 100:333–342. 2013.PubMed/NCBI View Article : Google Scholar : (In French).
|
|
10
|
Khan I and Steeg PS: Metastasis
suppressors: Functional pathways. Lab Invest. 98:198–210.
2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Shen T, Zhang K, Siegal GP and Wei S:
Prognostic value of E-cadherin and β-catenin in triple-negative
breast cancer. Am J Clin Pathol. 146:603–610. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hajra KM and Fearon ER: Cadherin and
catenin alterations in human cancer. Genes Chromosomes Cancer.
34:255–268. 2002.PubMed/NCBI View Article : Google Scholar
|
|
13
|
van Roy F and Berx G: The cell-cell
adhesion molecule E-cadherin. Cell Mol Life Sci. 65:3756–3788.
2008.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Behrens J and Birchmeier W: Cell-cell
adhesion in invasion and metastasis of carcinomas. Cancer Treat
Res. 71:251–266. 1994.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wong SHM, Fang CM, Chuah LH, Leong CO and
Ngai SC: E-cadherin: Its dysregulation in carcinogenesis and
clinical implications. Crit Rev Oncol Hematol. 121:11–22.
2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ashaie MA and Chowdhury EH: Cadherins: The
superfamily critically involved in breast cancer. Curr Pharm Des.
22:616–638. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Li Z, Yin S, Zhang L, Liu W and Chen B:
Prognostic value of reduced E-cadherin expression in breast cancer:
A meta-analysis. Oncotarget. 8:16445–16455. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Shiozaki H, Tahara H, Oka H, Miyata M,
Kobayashi K, Tamura S, Iihara K, Doki Y, Hirano S, Takeichi M, et
al: Expression of immunoreactive E-cadherin adhesion molecules in
human cancers. Am J Pathol. 139:17–23. 1991.PubMed/NCBI
|
|
19
|
Oka H, Shiozaki H, Kobayashi K, Inoue M,
Tahara H, Kobayashi T, Takatsuka Y, Matsuyoshi N, Hirano S,
Takeichi M, et al: Expression of E-cadherin cell adhesion molecules
in human breast cancer tissues and its relationship to metastasis.
Cancer Res. 53:1696–1701. 1993.PubMed/NCBI
|
|
20
|
Sawada K, Mitra AK, Radjabi AR, Bhaskar V,
Kistner EO, Tretiakova M, Jagadeeswaran S, Montag A, Becker A,
Kenny HA, et al: Loss of E-cadherin promotes ovarian cancer
metastasis via α 5-integrin, which is a therapeutic target. Cancer
Res. 68:2329–2339. 2008.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Bremnes RM, Veve R, Hirsch FR and Franklin
WA: The E-cadherin cell-cell adhesion complex and lung cancer
invasion, metastasis, and prognosis. Lung Cancer. 36:115–124.
2002.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Novak M, Jarrett SG, McCorkle JR, Mellon I
and Kaetzel DM: Multiple mechanisms underlie metastasis suppressor
function of NM23-H1 in melanoma. Naunyn Schmiedebergs Arch
Pharmacol. 384:433–438. 2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Steeg PS, Bevilacqua G, Pozzatti R, Liotta
LA and Sobel ME: Altered expression of NM23, a gene associated with
low tumor metastatic potential, during adenovirus 2 Ela inhibition
of experimental metastasis. Cancer Res. 48:6550–6554.
1988.PubMed/NCBI
|
|
24
|
Han W, Zhang C, Cao FY, Cao F, Jiang L and
Ding HZ: Prognostic and clinicopathological value of NM23
expression in patients with breast cancer: A systematic review and
meta-analysis. Curr Probl Cancer. 41:80–93. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Fang M, Tao Y, Liu Z, Huang H, Lao M,
Huang L and Zhu B: Meta-analysis of the relationship between NM23
expression to gastric cancer risk and clinical features. BioMed Res
Int. 2017(8047183)2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Ouatas T, Salerno M, Palmieri D and Steeg
PS: Basic and translational advances in cancer metastasis: Nm23. J
Bioenerg Biomembr. 35:73–79. 2003.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Steeg PS, de la Rosa A, Flatow U,
MacDonald NJ, Benedict M and Leone A: Nm23 and breast cancer
metastasis. Breast Cancer Res Treat. 25:175–187. 1993.PubMed/NCBI View Article : Google Scholar
|
|
28
|
MacDonald NJ, de la Rosa A and Steeg PS:
The potential roles of nm23 in cancer metastasis and cellular
differentiation. Eur J Cancer. 31A:1096–1100. 1995.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Salerno M, Palmieri D, Bouadis A,
Halverson D and Steeg PS: Nm23-H1 metastasis suppressor expression
level influences the binding properties, stability, and function of
the kinase suppressor of Ras1 (KSR1) Erk scaffold in breast
carcinoma cells. Mol Cell Biol. 25:1379–1388. 2005.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chen X, Xu Z and Wang Y: Recent advances
in breast cancer metastasis suppressor 1. Int J Biol Markers.
26:1–8. 2011.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kodura MA and Souchelnytskyi S: Breast
carcinoma metastasis suppressor gene 1 (BRMS1): Update on its role
as the suppressor of cancer metastases. Cancer Metastasis Rev.
34:611–618. 2015.PubMed/NCBI View Article : Google Scholar
|