Introduction
Osteoarthritis (OA) is a common degenerative joint
disease that usually occurs in elderly individuals (1). The pathogenesis of OA, which is caused
by the imbalance of joint tissue repair and destruction, is complex
(2). Chondrocytes are the cells of
the cartilage tissue that participate in the production of
cartilage extracellular matrix (ECM) (3), which serves an important role in the
maintenance of cartilage structure and function (4). With an increased global prevalence and
limited treatment options, the therapeutic strategies for OA are
still unsatisfactory (5,6); therefore, identifying novel biomarkers
for OA treatment is important.
MicroRNAs (miRNAs) are highly conserved short
non-coding RNAs 18-25 nucleotides long (7). Increasing evidence has indicated that
miRNAs serve vital roles during cartilage formation and remodeling
(8). Furthermore, a number of miRNAs
have been identified as regulators of OA progression, including
apoptosis and ECM degradation processes (9). For example, miRNA (miR)-103 restrained
chondrocyte proliferation by targeting Sox6 to trigger OA
progression (10). Additionally,
miR-9-5p overexpression hindered chondrocyte apoptosis by
inhibiting tenascin C expression in a mouse model of OA (11). In addition, miR-21-5p upregulated
collagen type II-α1 chain expression to facilitate cartilage
formation in interleukin (IL)-1β-induced chondrocytes (12). Therefore, miRNAs may serve as novel
therapeutic targets for OA. Of note, miR-17-5p has been
demonstrated to reverse HOX transcript antisense RNA and
fucosyltransferase 2-induced chondrocyte apoptosis (13); however, the molecular mechanism
underlying miR-17-5p in OA is not completely understood.
Enhancer of zeste homolog 2 (EZH2), a histone
methyltransferase, participates in the pathogenesis of different
types of cancer, including breast cancer, colon cancer and prostate
cancer (14). Emerging evidence has
demonstrated that EZH2 is often dysregulated at the transcriptional
and post-transcriptional level in a number of diseases, such as
prostate cancer and leukemia (15).
EZH2 dysregulation is a hallmark of disease progression in a number
of different types of cancer. For example, in gastric cancer
tissues, EZH2 expression was abnormally enhanced and EZH2
accelerated cell proliferation by targeting p21(16). Lui et al (17) reported that EZH2 was a regulator of
chondrocyte proliferation and hypertrophy; however, the
relationship between miR-17-5p and EZH2 has not been reported.
Therefore, the present study investigated the effect
of miR-17-5p on chondrocyte apoptosis and ECM degradation in
IL-1β-treated chondrocytes, as well as the molecular mechanisms
underlying miR-17-5p and miR-19b-3p activity in OA.
Materials and methods
Specimen collection
A total of 35 OA cartilage specimens from patients
with OA (age, 61.77±4.66 years; 23 female, 12 male) who underwent
joint replacement and 35 normal cartilage tissues from patients
(age, 41.51±4.01 years; 19 female, 16 male) with traumatic
emergency amputation without a history of OA or rheumatoid
arthritis were acquired from the People's Hospital of Rizhao
between July 2016 and August 2018. The present study was approved
by the Ethics Committee of the People's Hospital of Rizhao. All
participants provided written informed consent.
Cell culture
Cartilage samples were cut into small slices (<1
mm3) and digested with 0.2% trypsin for 30 min at 37˚C,
followed by 0.2% collagenase type II for 8 h at 37˚C. After
filtration and centrifugation at 1000 x g for 10 min, chondrocytes
were incubated in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) at 37˚C with 5% CO2.
Cell transfection and IL-1β
treatment
Vectors and oligonucleotides were synthesized by
Guangzhou RiboBio Co., Ltd. The following vectors and
oligonucleotides were used for transfection: miR-17-5p mimic
(miR-17-5p, 5'-CAAAGUGCUUACAGUGCAGGUAG-3'; 50 nM), mimic negative
control (miR-NC, 5'-UCGCUUGGUGCAGGUCGGGAA-3'; 50 nM), miR-17-5p
inhibitor (in-miR-17-5p, 5'-CUACCUGCACUGUAAGCACUUUG-3'; 100 nM),
inhibitor control (in-miR-NC, 5'-CAGUACUUUUGUGUAGUACAA-3'; 100 nM),
EZH2 overexpression vector (EZH2; 50 nM), empty overexpression
vector (pcDNA; 50 nM), small interfering RNA (si-RNA) targeting
EZH2 (si-EZH2, 5'-GAGGGAAAGUGUAUGAUAATT-3'; 100 nM), siRNA control
(si-NC, 5'-GGGAAAGAGUAUAUAGUGATT-3'; 100 nM), miR-19b-3p mimic
(miR-19b-3p, 5'-UGUGCAAAUCCAUGCAAAACUGA-3'; 50 nM) and miR-19b-3p
inhibitor (in-miR-19b-3p, 5'-UCAGUUUUGCAUGGAUUUGCACA-3'; 100 nM).
At 70% confluency, cells were transfected using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. After
transfection for 48 h at 37˚C, chondrocytes were treated with 10
ng/ml IL-1β (Beijing Solarbio Science and Technology Co., Ltd.) for
24 h at 37˚C.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cartilage tissues or
chondrocytes using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
Subsequently, RNA was reverse transcribed into cDNA using the
FastQuant RT kit (Tiangen Biotech Co., Ltd.) or miScript Reverse
Transcription kit (Qiagen GmbH), according to the manufacturer's
protocol. qPCR was performed using the SYBR Green PCR Master Mix
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. The reactions were incubated at 95˚C for 30 sec, followed
by 40 cycles of 95˚C for 5 sec, 60˚C for 10 sec, 95˚C for 5 sec and
60˚C for 10 sec. The following primer pairs were purchased from
(Guangzhou RiboBio Co., Ltd.) and used for qPCR: miR-17-5p forward,
5'-CGGCGGCAAAGTGCTTACAG-3' and reverse, 5'-GTGCAGGGTCCGAGGT-3';
miR-19b-3p forward, 5'-ACACTCCAGCTGGGTGTGCAAATCCATGCAA-3' and
reverse, 5'-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCAGTTT-3'; EZH2
forward, 5'-AATCAGAGTACATGCGACTGAGA-3' and reverse,
5'-AATCAGAGTACATGCGACTGAGA-3'; GAPDH forward,
5'-GCTGAGTATGTCGTGGAGTC-3' and reverse, 5'-AGTTGGTGGTGCAGGATGC-3';
and U6 forward, 5'-CTCGCTTCGGCAGCACA-3' and reverse,
5'-AACGCTTCACGAATTTGCGT-3'. mRNA and miRNA levels were normalized
to the internal reference genes GAPDH and U6, respectively.
Expression levels were quantified via the 2−ΔΔCq method
(18).
Western blot analysis
Total protein was extracted using RIPA buffer
(Sigma-Aldrich; Merck KGaA). Protein samples were quantified using
a BCA Protein Assay Kit (cat. no. ab102536; Abcam). Equal amounts
of protein samples (20 µg) were separated by 10% SDS-PAGE and
transferred to PVDF membranes (EMD Millipore). Subsequently, the
membrane was blocked with 5% skimmed milk for 2 h at room
temperature. The membrane was incubated with primary antibodies
overnight at 4˚C against matrix metalloproteinase-13 (MMP13; cat.
no. ab39012; 60 KDa; dilution, 1:4,000; Abcam), Collagen II (cat.
no. ab34712; 142 KDa; dilution, 1:2,000; Abcam), Aggrecan (cat. no.
ab36861; 110 KDa; dilution, 1:2,000; Abcam), EZH2 (cat. no.
ab186006; 85 KDa; dilution, 1:1,000; Abcam) and β-actin (cat. no.
ab8227; 42 KDa; dilution, 1:2,000; Abcam). Following primary
antibody incubation, the membranes were incubated for 2 h at room
temperature with a secondary anti-rabbit antibody marked by
horseradish peroxidase (cat. no. ab7090; dilution, 1:20,000;
Abcam). Protein bands were visualized using ECL reagents (EMD
Millipore) and quantified by densitometry analysis using ImageJ
software (version 1.6.0; National Institutes of Health, Inc.).
β-actin was used as the loading control.
Flow cytometry
Chondrocytes (1x105 cells/well) were
seeded into 6-well plates and washed twice with cold PBS. Apoptotic
cells were detected using the Annexin V-FITC/propidium iodide
Apoptosis Detection kit (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocol. Early apoptotic
cells were analyzed using an Attune NxT flow cytometer (Thermo
Fisher Scientific, Inc.) and calculated using Cell Quest
acquisition software (version 2.9; BD Biosciences, Inc.).
Dual-luciferase reporter assay
The binding sequences between miR-17-5p or
miR-19b-3p and EZH2 were predicted using MicroT-CDS software
(diana.imis.athena-innovation.gr/DianaTools/index.php?r=microT_CDS/index).
The EZH2 3'-untranslated region (3'-UTR) containing wild-type (WT)
or mutant (MUT) binding sites for miR-17-5p or miR-19b-3p
(Guangzhou RiboBio Co., Ltd.) were inserted into a pGL3 vector
(Promega Corporation). Subsequently, the luciferase reporter and
miR-17-5p, miR-19b-3p, miR-NC, in-miR-17-5p, in-miR-19b-3p or
in-miR-NC were co-transfected into chondrocytes (5x104
cells/well) using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Following 48 h incubation at 37˚C, luciferase activity
was detected using a Dual-Lucy assay kit (Beijing Solarbio Science
and Technology Co., Ltd.), according to the manufacturer's
protocol. Renilla luciferase activity was used for
normalization.
Radioimmunoprecipitation (RIP)
assay
The Magna RIP kit (EMD Millipore) was used to
perform the RIP assay, according to the manufacturer's protocol.
Briefly, chondrocytes were transfected with miR-17-5p, miR-19b-3p
or miR-NC. Subsequently, chondrocytes were harvested and lysed
using RIP lysis buffer. The cell lysates were collected and
incubated with RIP-argonaute RISC catalytic component 2 (Ago2)
antibody (EMD Millipore) or RIP-immunoglobulin G (IgG) antibody
(EMD Millipore) overnight at 4˚C. EZH2 expression was detected by
RT-qPCR.
Statistical analysis
Data are presented as the mean ± standard deviation.
GraphPad Prism 7.0 software (GraphPad Software, Inc.) was used to
perform statistical analyses. The correlation between miR-17-5p and
EZH2 levels in OA cartilage tissues was analyzed using Spearman's
correlation coefficient. Data were analyzed using the Student's
t-test or one-way ANOVA followed by Tukey's post hoc test. All
experiments were performed at least three times. P<0.05 was
considered to indicate a statistically significant difference.
Results
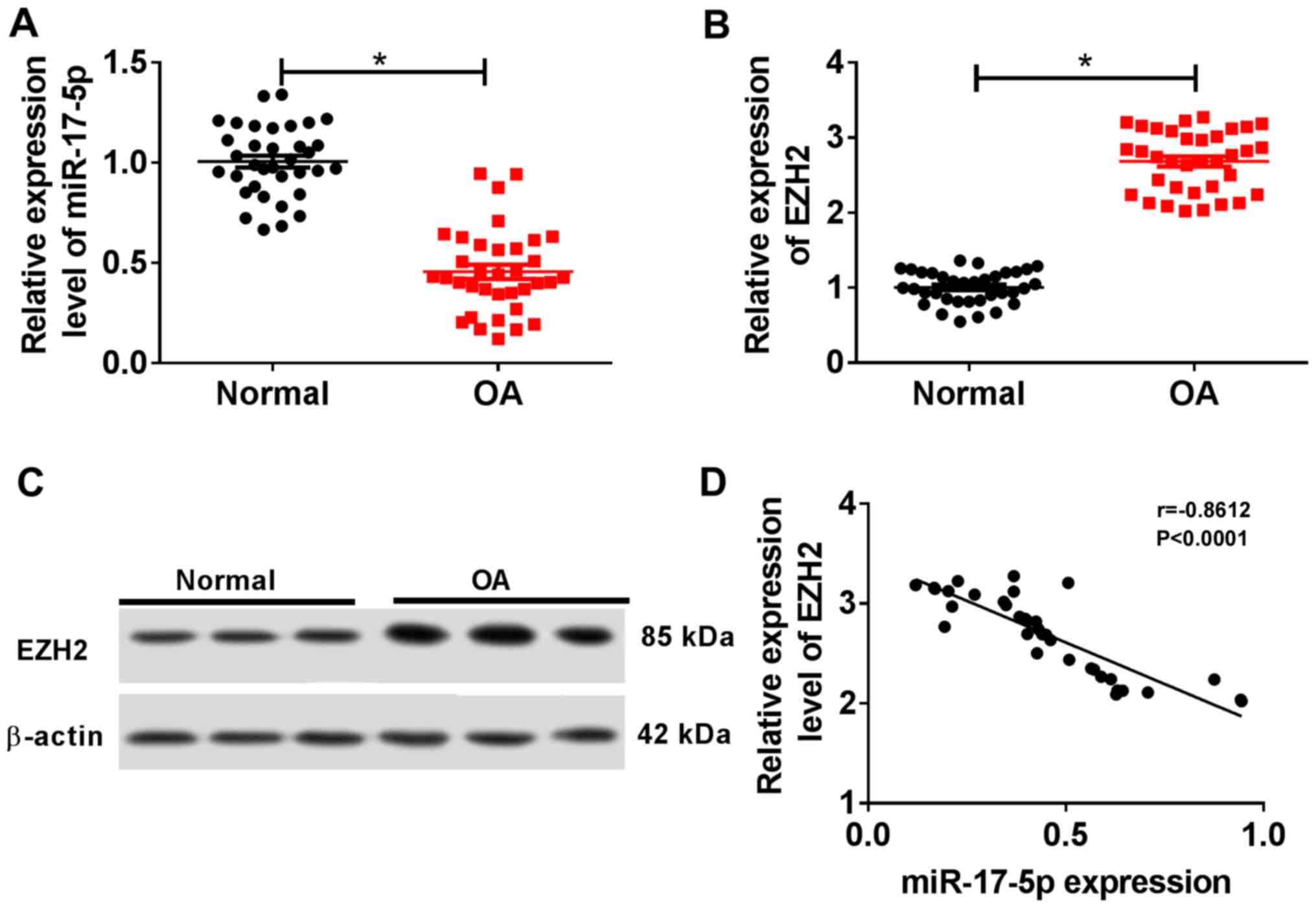
miR-17-5p is downregulated and EZH2 is
upregulated in OA cartilage tissues
The RT-qPCR results suggested that miR-17-5p
expression was significantly decreased in OA cartilage tissues
compared with normal cartilage tissues (Fig. 1A). By contrast, EZH2 expression was
significantly increased in OA cartilage tissues compared with
normal cartilage tissues (Fig. 1B
and C). Additionally, miR-17-5p
expression was negatively correlated with EZH2 expression in OA
cartilage tissues (Fig. 1D). These
results suggested that miR-17-5p may serve a role in OA
progression.
miR-17-5p inhibits IL-1β-induced
chondrocyte apoptosis and ECM degradation
To investigate the role of miR-17-5p during the
pathogenesis of OA, chondrocytes were transfected with miR-17-5p,
miR-NC, in-miR-17-5p or in-miR-NC and subsequently treated with 10
ng/ml IL-1β for 24 h. Transfection efficiency of the miR-17-5p
mimics and inhibitor was determined by RT-qPCR (Fig. S1A). The expression of miR-17-5p was
significantly decreased in the IL-1β group compared with that in
the control group, and the miR-17-5p mimic significantly reversed
the IL-1β-induced effect (Fig. 2A).
miR-17-5p expression was also significantly reduced in the IL-1β +
in-miR-17-5p group compared with the IL-1β + in-miR-NC group
(Fig. 2B). Furthermore, flow
cytometry demonstrated that the proportion of apoptotic cells was
significantly increased in the IL-1β group compared with that in
the control group. In addition, the miR-17-5p mimics inhibited the
IL-1β-induced apoptosis, and miR-17-5p inhibition significantly
increased the IL-1β-induced apoptosis (Fig. 2C and D). IL-1β treatment also resulted in a
significant decrease in the protein expression levels of the
cartilage formation proteins Collagen II and Aggrecan, and a
significant increase in the protein expression level of the
cartilage-degrading enzyme MMP13 compared with the control group.
By contrast, the miR-17-5p mimics counteracted the IL-1β-induced
effects, and miR-17-5p inhibition enhanced the IL-1β-induced
effects on cartilage-related protein expression (Fig. 2E and F). These results indicated that miR-17-5p
modulated apoptosis and ECM degradation in IL-1β-induced
chondrocytes.
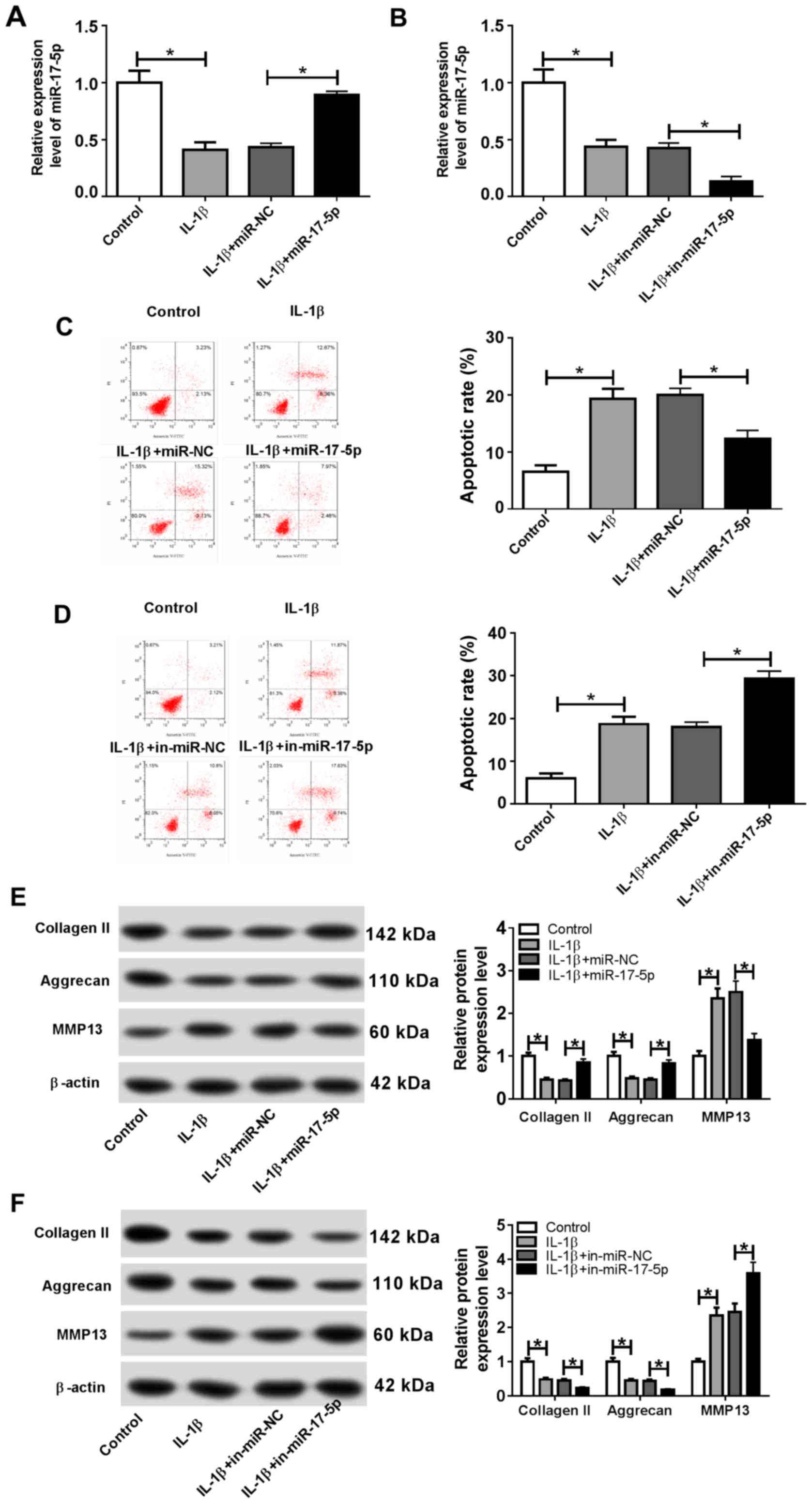 | Figure 2miR-17-5p decreases IL-1β-induced
cell apoptosis and ECM degradation in chondrocytes. Chondrocytes
were transfected with miR-17-5p mimic, miR-NC, in-miR-17-5p or
in-miR-NC prior to IL-1β treatment. (A and B) The expression of
miR-17-5p was measured by reverse transcription-quantitative PCR in
chondrocytes transfected with (A) the miR-17-5p mimics and (B)
in-miR-17-5p. (C and D) The proportion of apoptotic chondrocytes
was determined by flow cytometry following transfection with (C)
the miR-17-5p mimics and (D) in-miR-17-5p. (E and F) The protein
expression levels of ECM-associated proteins Collagen II, Aggrecan
and MMP13 were determined by western blotting in chondrocytes
transfected with (E) the miR-17-5p mimics and (D) in-miR-17-5p.
*P<0.05. miR, microRNA; IL-1β, interleukin-1β; ECM,
extracellular matrix; NC, negative control; in, inhibitor; MMP13,
matrix metalloproteinase 13; PI, propidium iodide. |
EZH2 is a target of miR-17-5p in
chondrocytes
To explore the mechanism underlying miR-17-5p
activity during OA progression, the MicroT-CDS online database was
used, which predicted that miR-17-5p and EZH2 3'UTR exhibited
putative binding sites (Fig. 3A).
The dual-luciferase reporter assay revealed that the miR-17-5p
mimics significantly reduced the luciferase activity of EZH2#1
3'UTR-WT, but displayed no effect on the luciferase activity of
EZH2#1 3'UTR-MUT compared with the control group (Fig. 3B). In addition, the luciferase
activity of EZH2#1 3'UTR-WT was significantly enhanced by the
miR-17-5p inhibitor, but was not altered for EZH2#1 3'UTR-MUT
compared with the control group (Fig.
3C). The RIP assay was performed to further investigate the
relationship between miR-17-5p and EZH2. The results demonstrated
that EZH2 was significantly enriched in the miR-17-5p group coated
with the Ago2 antibody compared with the control group (Fig. 3D). Furthermore, the protein
expression levels of EZH2 were measured in chondrocytes transfected
with miR-NC, miR-17-5p, in-miR-NC or in-miR-17-5p. The results
suggested that the miR-17-5p mimics significantly decreased the
expression of EZH2, and miR-17-5p inhibition significantly
increased the expression level of EZH2 in chondrocytes (Fig. 3E). These results suggested that EZH2
was a direct target of miR-17-5p in chondrocytes.
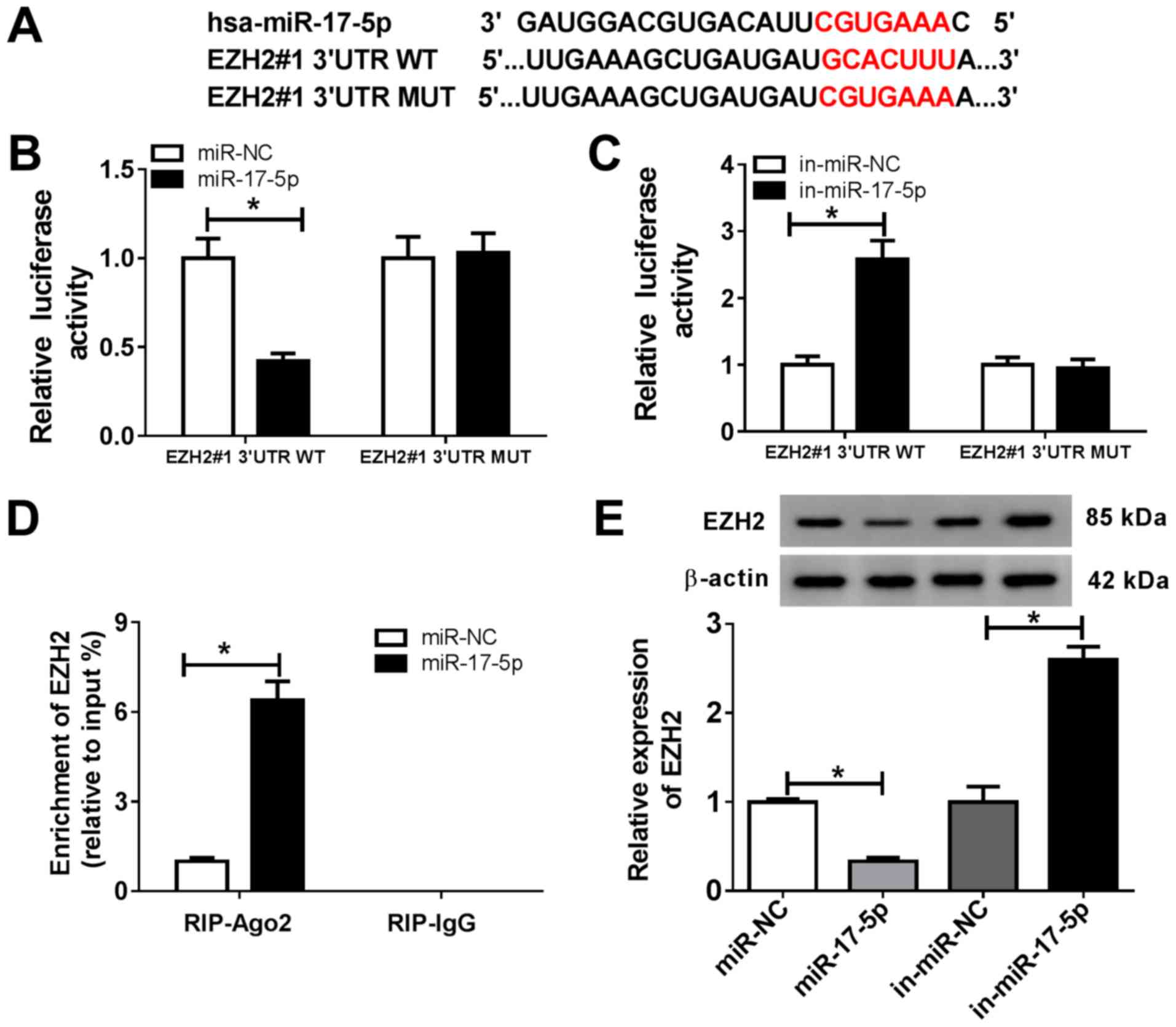 | Figure 3EZH2 is a target of miR-17-5p in
chondrocytes. (A) The predicted binding sites of miR-17-5p and EZH2
3'UTR. (B and C) The luciferase activity in chondrocytes
co-transfected with EZH2#1 3'UTR-WT or EZH2#1 3'UTR-MUT and (B)
miR-NC or the miR-17-5p mimic, or (C) in-miR-NC or in-miR-17-5p was
determined using a dual-luciferase reporter assay. (D) The RIP
assay was used to validate the relationship between miR-17-5p and
EZH2. (E) EZH2 protein expression was measured in chondrocytes
transfected with miR-NC, miR-17-5p mimic, in-miR-NC or
in-miR-17-5p. *P<0.05 vs. NC. EZH2, enhancer of zeste
homolog 2; miR, microRNA; 3'UTR, 3'-untranslated region; WT,
wild-type; MUT, mutant; NC, negative control; in, inhibitor; Ago2,
argonaute RISC catalytic component 2; IgG, immunoglobulin G. |
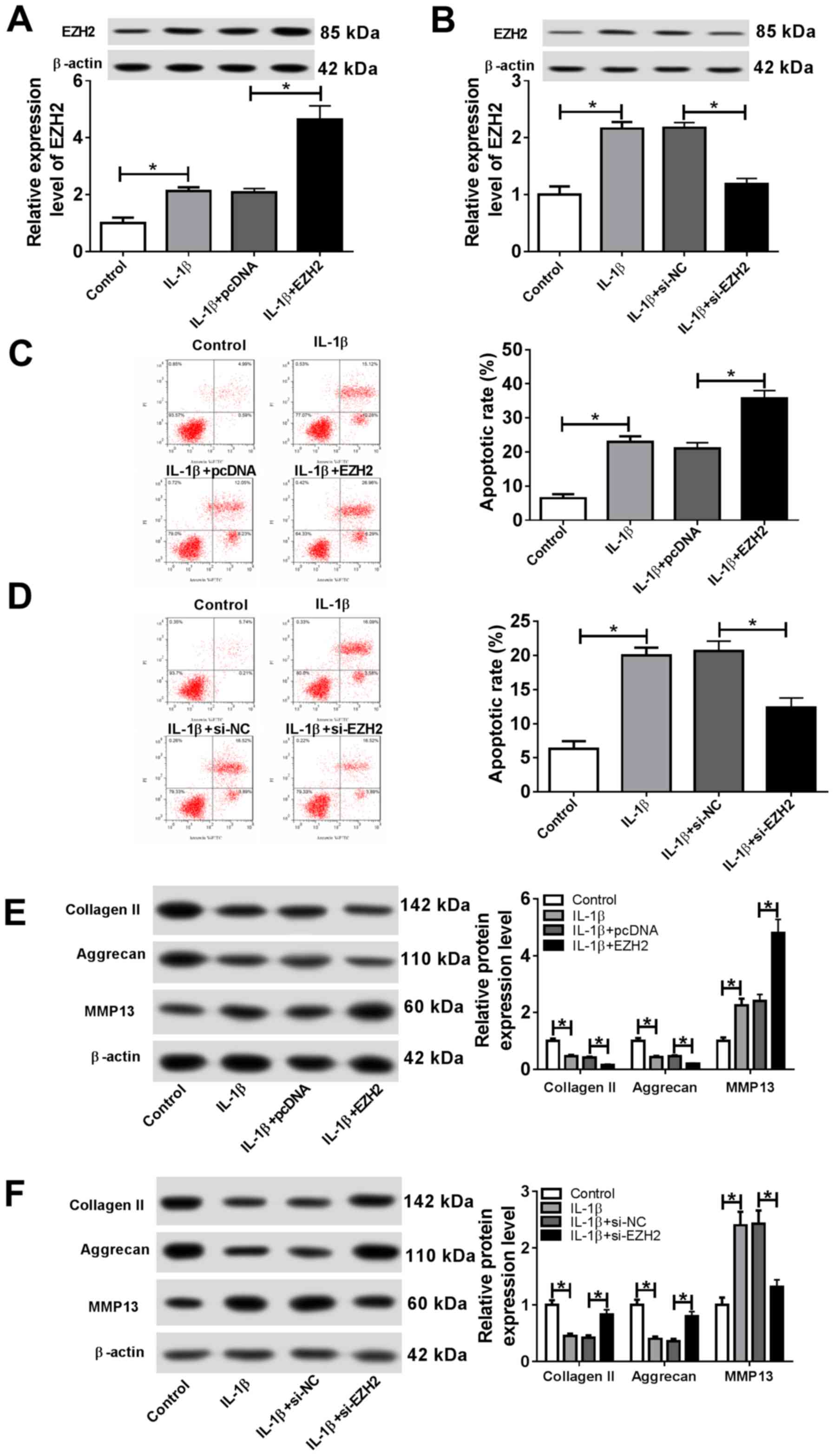
EZH2 facilitates IL-1β-induced
apoptosis and ECM degradation in chondrocytes
To investigate the effects of EZH2 on OA
progression, chondrocytes were transfected with EZH2, pcDNA,
si-EZH2 or si-NC. Subsequently, transfected cells were stimulated
with IL-1β. The transfection efficiency of EZH2 overexpression and
knockdown were detected by RT-qPCR (Fig. S1C). The results suggested that IL-1β
stimulation significantly increased EZH2 expression, whereas EZH2
overexpression significantly enhanced EZH2 expression, and EZH2
knockdown significantly decreased EZH2 expression in
IL-1β-stimulated chondrocytes (Fig.
4A and 4B). Additionally, the
proportion of apoptotic cells was significantly increased following
EZH2 overexpression, whereas si-EZH2 significantly reduced the
proportion of apoptotic IL-1β-treated chondrocytes (Fig. 4C and 4D). EZH2 overexpression significantly
decreased the expression level of the cartilage formation proteins
Collagen II and Aggrecan, and significantly increased the
expression level of the cartilage-degrading enzyme MMP13; however,
EZH2 knockdown displayed the opposite effect (Fig. 4E and 4F). The results suggested that EZH2
regulated cell apoptosis and ECM degradation in IL-1β-induced
chondrocytes.
EZH2 overexpression reverses the
effect of miR-17-5p overexpression on the progression of OA
To further investigate the role of miR-17-5p and
EZH2 during OA progression, chondrocytes were transfected with
miR-NC, miR-17-5p, miR-17-5p + pcDNA or miR-17-5p + EZH2, and
subsequently stimulated with IL-1β for 24 h. The results indicated
that EZH2 expression was significantly decreased by miR-17-5p
overexpression, which was reversed by EZH2 overexpression (Fig. 5A). Furthermore, the miR-17-5p mimics
significantly reduced the proportion of apoptotic chondrocytes, and
EZH2 overexpression reversed the miR-17-5p-induced effects on
chondrocyte apoptosis (Fig. 5B). In
addition, transfection with the miR-17-5p mimics resulted in a
significant increase in the expression levels of Collagen II and
Aggrecan and a significant decrease in MMP13 expression compared
with those in the control group. EZH2 overexpression reversed the
miR-17-5p mimic-induced effects on cartilage-associated protein
expression (Fig. 5C). These results
suggested that EZH2 overexpression counteracted the effects of the
miR-17-5p mimics on OA progression.
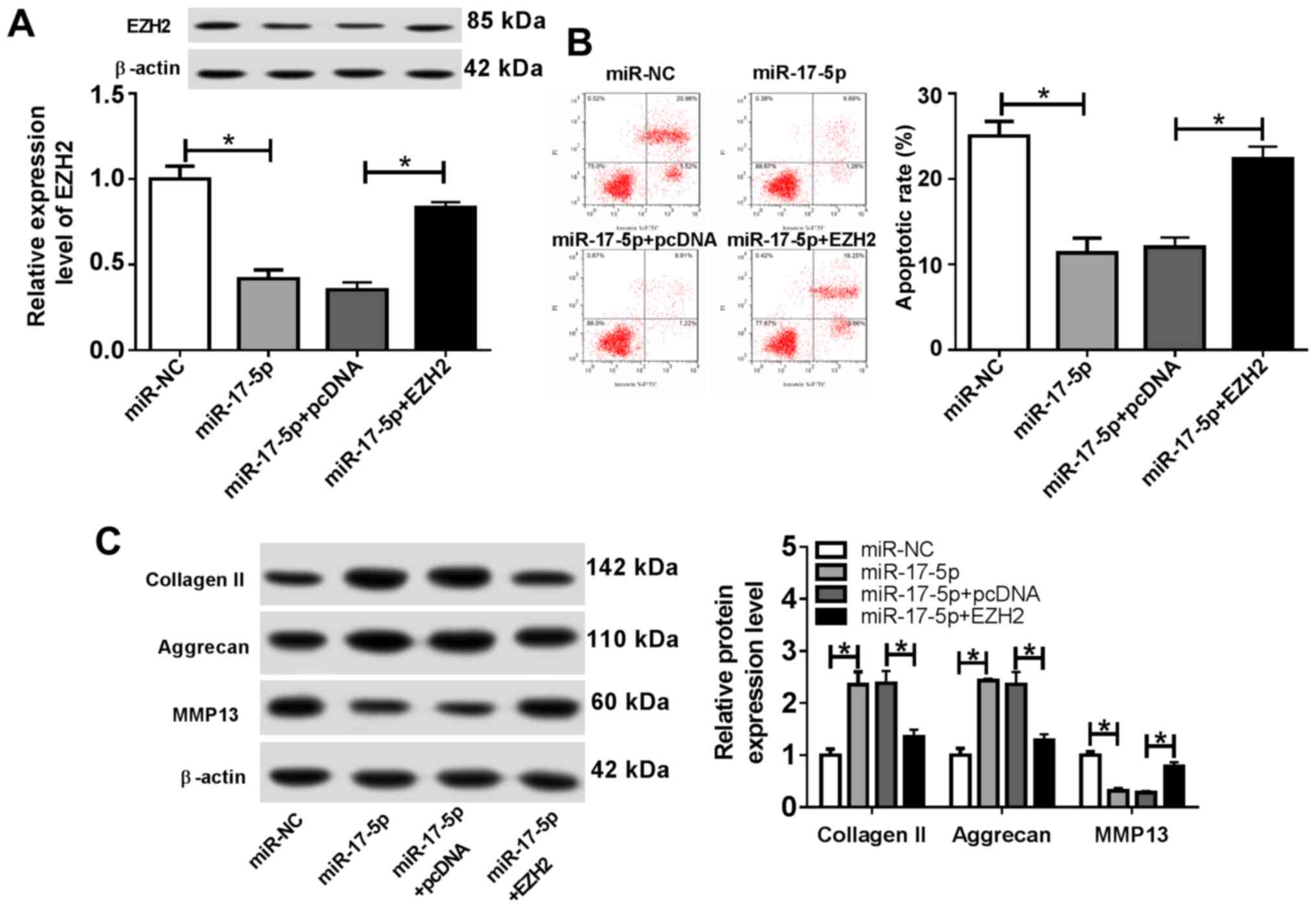 | Figure 5EZH2 overexpression reverses the
effects of the miR-17-5p mimics on osteoarthritis progression.
Chondrocytes were transfected with miR-NC, miR-17-5p mimic,
miR-17-5p mimic + pcDNA or miR-17-5p + EZH2 overexpression vector,
and treated with IL-1β for 24 h. (A) EZH2 protein expression was
determined using western blotting. (B) The proportion of apoptotic
cells was determined by flow cytometry. (C) The protein expression
levels of extracellular matrix-associated proteins were detected by
western blotting. *P<0.05, as indicated. EZH2,
enhancer of zeste homolog 2; miR, microRNA; NC, negative control;
IL-1β, interleukin-1β; PI, propidium iodide; MMP13, matrix
metalloproteinase 13. |
miR-19b-3p directly targets EZH2 in
chondrocytes. The MicroT-CDS online database also predicted
that miR-19b-3p and EZH2 3'UTR displayed putative binding sites
(Fig. 6A). The dual-luciferase
reporter assay suggested that the miR-19b-3p mimics significantly
decreased the luciferase activity of EZH2#2 3'UTR-WT, and the
miR-19b-3p inhibitor significantly enhanced the luciferase activity
of EZH2#2 3'UTR-WT (Fig. 6B and
C). The RIP assay results indicated
that EZH2 was significantly enriched in the miR-19b-3p group coated
with the Ago2 antibody compared with the control group (Fig. 6D). The transfection efficiency of
miR-19b-3p was verified by RT-qPCR (Fig. S1B). In addition, the protein
expression of EZH2 was significantly decreased in chondrocytes
transfected with the miR-19b-3p mimic compared with that in the
miR-NC group. EZH2 expression was significantly increased in
chondrocytes transfected with in-miR-19b-3p compared with that in
the in-miR-NC group (Fig. 6E). These
results suggested that miR-19b-3p directly targeted EZH2 and
negatively regulated EZH2 expression in chondrocytes.
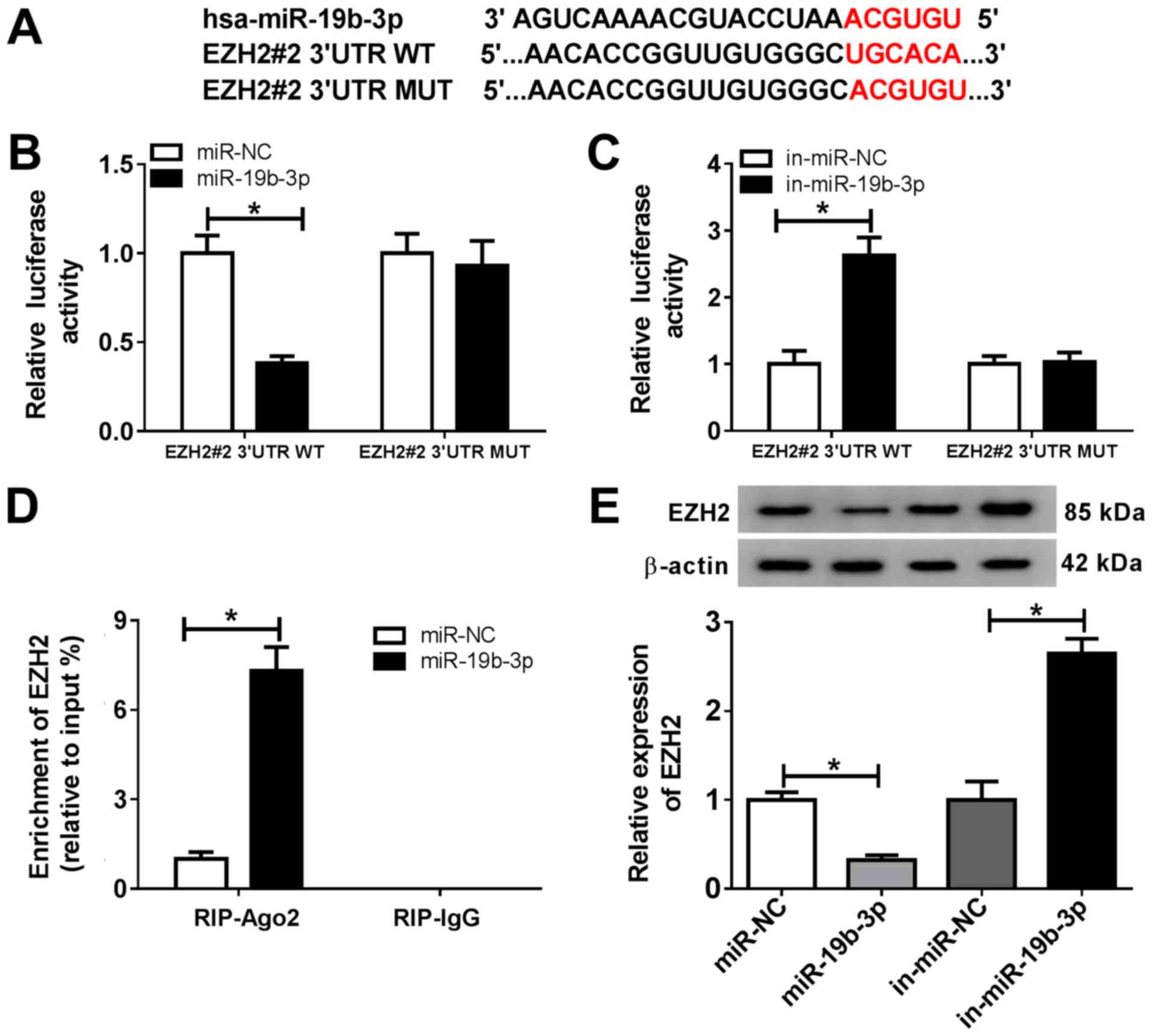 | Figure 6miR-19b-3p directly targets EZH2. (A)
The putative binding sites between miR-19b-3p and EZH2 3'UTR. (B
and C) EZH2#2 3'UTR-WT or EZH2#2 3'UTR-MUT and (B) miR-NC or the
miR-19b-3p mimics, or (C) in-miR-NC or in-miR-19b-3p were
co-transfected into chondrocytes, and a dual luciferase reporter
assay was performed to evaluate the luciferase activity. (D) The
RIP assay was performed to verify the relationship between
miR-19b-3p and EZH2. (E) EZH2 expression was detected in
chondrocytes transfected with miR-NC, the miR-19b-3p mimics,
in-miR-NC or in-miR-19b-3p. *P<0.05 vs. control. miR,
microRNA; EZH2, enhancer of zeste homolog 2; 3'UTR, 3'-untranslated
region; WT, wild-type; MUT, mutant; NC, negative control; in,
inhibitor; Ago, argonaute RISC catalytic component 2; IgG,
immunoglobulin G. |
EZH2 overexpression reverses the
effects of the miR-19b-3p mimics on OA progression
To further investigate the role of miR-19b-3p during
the development of OA, transfected chondrocytes were treated with
IL-1β for 24 h. The western blotting results indicated that EZH2
expression was significantly decreased in the miR-19b-3p mimics
group compared with that in the miR-NC group, whereas EZH2
overexpression reversed the miR-19b-3p-induced effects on EZH2
expression (Fig. 7A). The proportion
of apoptotic cells was significantly reduced in the miR-19b-3p
mimics group, and EZH2 overexpression reversed the effect of the
miR-19b-3p mimics on chondrocyte apoptosis (Fig. 7B). Furthermore, the levels of
Collagen II and Aggrecan expression were significantly increased
and MMP13 expression was significantly decreased in the miR-19b-3p
mimics group compared with the miR-NC group. Similarly, the
miR-19b-3p mimic-induced effects were reversed by EZH2
overexpression (Fig. 7C). These
results suggested that EZH2 overexpression reversed the effects of
miR-19b-3p overexpression on OA progression.
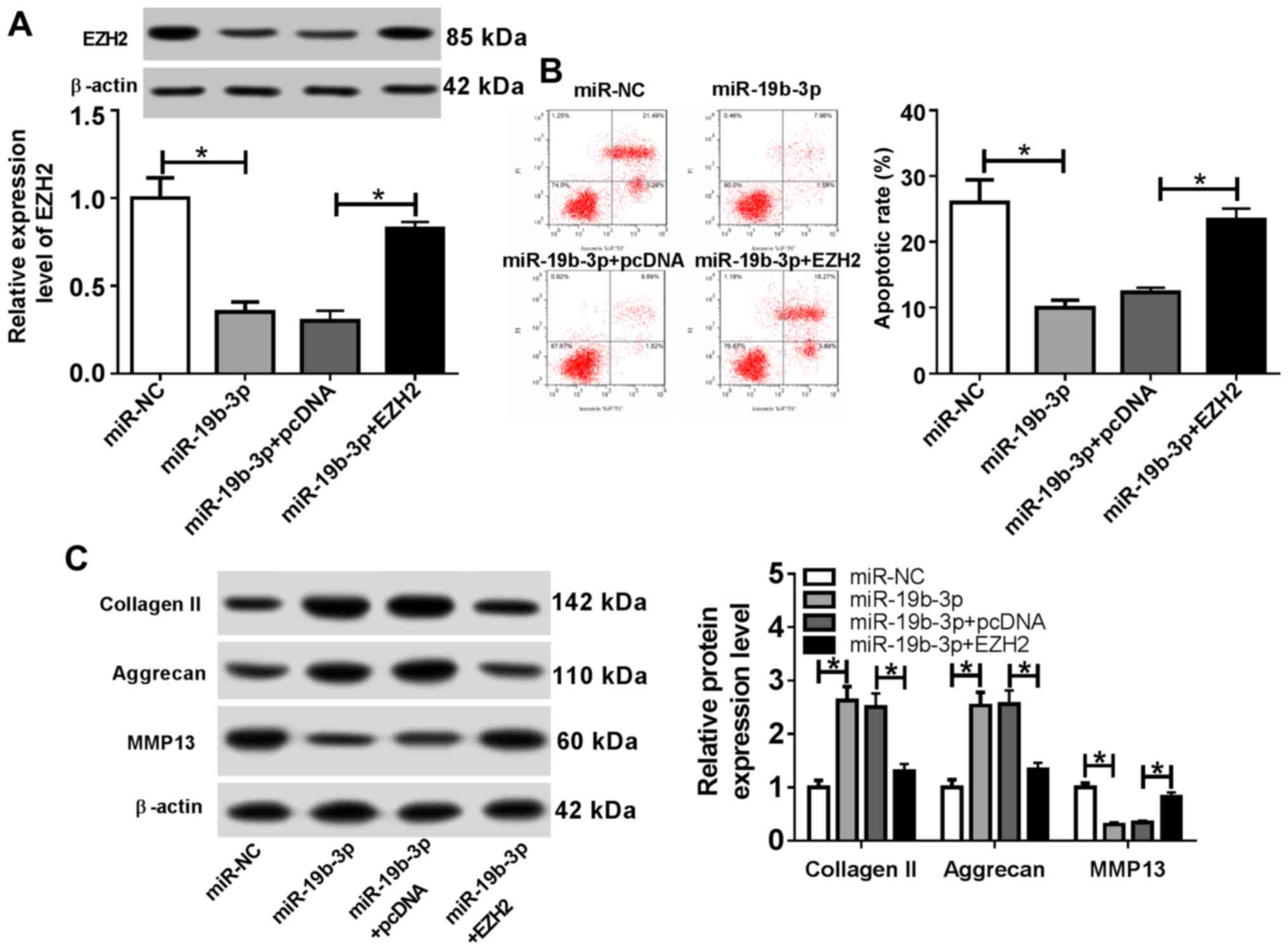 | Figure 7EZH2 overexpression reverses the
effects of the miR-19b-3p mimics on osteoarthritis progression.
Following transfection with miR-NC, miR-19b-3p mimic, miR-19b-3p
mimic + pcDNA or miR-19b-3p mimic + EZH2 overexpression vector,
chondrocytes were stimulated with IL-1β for 24 h. (A) EZH2 protein
expression was measured by western blotting. (B) The proportion of
apoptotic cells was assessed using flow cytometry. (C) The protein
expression levels of extracellular matrix-related proteins were
examined by western blotting. *P<0.05, as indicated.
EZH2, enhancer of zeste homolog 2; miR, microRNA; NC, negative
control; IL-1β, interleukin-1β; PI, propidium iodide; MMP13, matrix
metalloproteinase 13. |
Discussion
OA is characterized by cartilage destruction, and
the pathogenesis of the disease involves chondrocyte apoptosis and
ECM degradation (19). Musumeci
et al (20) reported that the
proportion of apoptotic chondrocytes was significantly increased
after injury. In a rat model of OA, adenylate cyclase-activating
polypeptide 1 prevented IL-1β-induced chondrocyte apoptosis in
vitro (21). Molecular markers that regulate chondrocyte
apoptosis are important during the development of OA (22). According to the literature, ECM
degradation participates in the pathogenesis of OA, leading to the
loss of cartilage tissue (23). To
reduce the incidence of degenerative joint diseases, a number of OA
treatment strategies have been developed in recent years, including
the use of fibrates or collagen cell carrier scaffolds (24,25).
Increasing evidence has suggested that a number of miRNAs are
associated with OA progression (26); therefore, the role of miRNAs in
chondrocyte apoptosis and ECM degradation was investigated in the
present study.
Previous studies have reported that miRNA-17-5p
modulates cell autophagy and apoptosis to affect cellular
senescence, aging and cancer (27).
For example, aberrant expression of miNA-17-5p regulates the
osteoblastic differentiation of mesenchymal stem cells by targeting
SMAD7(28). Moreover, increasing
evidence has indicated that miRNA-17-5p may serve as a tumor
inhibitor in breast cancer and an oncogene in pancreatic cancer
(29,30). miRNA-19b-3p blocks the progression of
breast cancer by mediating the PI3K/Akt signaling pathway (31). A previous study reported that
miRNA-17-5p was significantly downregulated in OA chondrocytes and
facilitated autophagy by decreasing p62 expression (32). Additionally, miRNA-19b-3p decreases
IL-1β-induced ECM degradation and inflammatory injury in
chondrocytes by regulating G protein-coupled receptor kinase 6
expression (33). Similar to
previous studies, the results of the present study indicated that
miR-17-5p expression was decreased in OA chondrocytes compared with
that in the control cells. The results also suggested that
miR-17-5p modulated OA progression by inhibiting chondrocyte
apoptosis and ECM degradation.
Histone methyltransferase EZH2 inhibits osteoblast
maturation and bone development (34). A number of studies have reported that
EZH2 upregulation in different tumors is associated with adverse
outcomes (35,36). Previous studies have also
demonstrated that the level of EZH2 in OA chondrocytes was
significantly elevated; therefore, the use of EZH2 inhibitors may
serve as a therapeutic strategy for OA (37,38).
Consistent with previous studies, EZH2 expression was significantly
increased in OA chondrocytes compared with control cells in the
present study. In addition, it has been reported that miRNAs serve
a vital role in a number of diseases, including prostate cancer,
osteoporosis and osteoarthritis (39,40), by
targeting the 3'UTR of target genes to downregulate protein
expression (40). In the present
study, the MicroT-CDS online database predicted that EZH2 and
miR-17-5p or miR-19b-3p displayed putative binding sites, which was
confirmed by the dual-luciferase reporter and RIP assays. In
addition, the rescue experiments indicated that miR-17-5p and
miR-19b-3p modulated OA progression by targeting EZH2.
In conclusion, the results of the present study
suggested that miR-17-5p and miR-19b-3p inhibited chondrocyte
apoptosis and ECM degradation by targeting EZH2, which indicated
that these miRNAs may serve as promising preventative and
therapeutic biomarkers for OA. However, animal experiments are
required to verify the results of the present study before the
miRNAs can be used in the clinic.
Supplementary Material
Efficiency of transfection determined
by reverse transcription-quantitative PCR. (A-C) Chondrocytes were
transfected with (A) miR-17-5p mimic, in-miR-17-5p, (B) miR-19b-3p
mimic, in-miR-19b-3p, (C) EZH2 overexpression vector and si-EZH2.
*P<0.05. miR, microRNA; in, inhibitor; si, small
interfering RNA; EZH2, enhancer of zeste homolog 2.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
FY and YS conceived the study and performed the
experiments. YS, XG and YL analyzed and interpreted the data. YL,
FY and YS drafted and critically revised the manuscript for
important intellectual content. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical Review
Committee of The People's Hospital of Rizhao (Rizhao, China).
Written informed consent was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Creamer P and Hochberg MC: Osteoarthritis.
Lancet. 350:503–508. 1997.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hunter DJ and Bierma-Zeinstra S:
Osteoarthritis. Lancet. 393:1745–1759. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Charlier E, Deroyer C, Ciregia F, Malaise
O, Neuville S, Plener Z, Malaise M and de Seny D: Chondrocyte
dedifferentiation and osteoarthritis (OA). Biochem Pharmacol.
165:49–65. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Şahin Ş, Tuncel SA, Salimi K, Bilgiç E,
Korkusuz P and Korkusuz F: Advanced injectable alternatives for
osteoarthritis. Adv Exp Med Biol. 1077:183–96. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ghouri A and Conaghan PG: Update on novel
pharmacological therapies for osteoarthritis. Ther Adv
Musculoskelet Dis. 11(1759720X19864492)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Vinatier C, Merceron C and Guicheux J:
Osteoarthritis: From pathogenic mechanisms and recent clinical
developments to novel prospective therapeutic options. Drug Discov
Today. 21:1932–1937. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yao Q, Chen Y and Zhou X: The roles of
microRNAs in epigenetic regulation. Curr Opin Chem Biol. 51:11–17.
2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wu C, Tian B, Qu X, Liu F, Tang T, Qin A,
Zhu Z and Dai K: MicroRNAs play a role in chondrogenesis and
osteoarthritis (review). Int J Mol Med. 34:13–23. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Malemud CJ: MicroRNAs and osteoarthritis.
Cells 7: pii. (E92)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen J and Wu X: MicroRNA-103 contributes
to osteoarthritis development by targeting Sox6. Biomed
Pharmacother. 118(109186)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen H, Yang J and Tan Z: Upregulation of
microRNA-9-5p inhibits apoptosis of chondrocytes through
downregulating Tnc in mice with osteoarthritis following tibial
plateau fracture. J Cell Physiol. 234:23326–23336. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhu H, Yan X, Zhang M, Ji F and Wang S:
miR-21-5p protects IL-1β-induced human chondrocytes from
degradation. J Orthop Surg Res. 14(118)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hu J, Wang Z, Shan Y, Pan Y, Ma J and Jia
L: Long non-coding RNA HOTAIR promotes osteoarthritis progression
via miR-17-5p/FUT2/β-catenin axis. Cell Death Dis.
9(711)2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Yan KS, Lin CY, Liao TW, Peng CM, Lee SC,
Liu YJ, Chan WP and Chou RH: EZH2 in cancer progression and
potential application in cancer therapy: A friend or foe? Int J Mol
Sci. 18: pii(E1172)2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yamagishi M and Uchimaru K: Targeting EZH2
in cancer therapy. Curr Opin Oncol. 29:375–381. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Xu J, Wang Z, Lu W, Jiang H, Lu J, Qiu J
and Ye G: EZH2 promotes gastric cancer cells proliferation by
repressing p21 expression. Pathol Res Pract.
215(152374)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lui JC, Garrison P, Nguyen Q, Ad M,
Keembiyehetty C, Chen W, Jee YH, Landman E, Nilsson O, Barnes KM
and Baron J: EZH1 and EZH2 promote skeletal growth by repressing
inhibitors of chondrocyte proliferation and hypertrophy. Nat
Commun. 7(13685)2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhang FJ, Luo W and Lei GH: Role of HIF-1α
and HIF-2α in osteoarthritis. Joint Bone Spine. 82:144–147.
2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Musumeci G, Castrogiovanni P, Loreto C,
Castorina S, Pichler K and Weinberg AM: Post-traumatic caspase-3
expression in the adjacent areas of growth plate injury site: A
morphological study. Int J Mol Sci. 14:15767–15784. 2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Giunta S, Castorina A, Marzagalli R,
Szychlinska MA, Pichler K, Mobasheri A and Musumeci G: Ameliorative
effects of PACAP against cartilage degeneration. Morphological,
immunohistochemical and biochemical evidence from in vivo and in
vitro models of rat osteoarthritis. Int J Mol Sci. 16:5922–5944.
2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Musumeci G, Castrogiovanni P, Trovato FM,
Weinberg AM, Al-Wasiyah MK, Alqahtani MH and Mobasheri A:
Biomarkers of chondrocyte apoptosis and autophagy in
osteoarthritis. Int J Mol Sci. 16:20560–20575. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Shi Y, Hu X, Cheng J, Zhang X, Zhao F, Shi
W, Ren B, Yu H, Yang P, Li Z, et al: A small molecule promotes
cartilage extracellular matrix generation and inhibits
osteoarthritis development. Nat Commun. 10(1914)2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Szychlinska MA, Ravalli S and Musumeci G:
Pleiotropic effect of fibrates on senescence and autophagy in
osteoarthritis. EBioMedicine. 45:11–12. 2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Szychlinska MA, Castrogiovanni P, Nsir H,
Di Rosa M, Guglielmino C, Parenti R, Calabrese G, Pricoco E,
Salvatorelli L, Magro G, et al: Engineered cartilage regeneration
from adipose tissue derived-mesenchymal stem cells: A
morphomolecular study on osteoblast, chondrocyte and apoptosis
evaluation. Exp Cell Res. 357:222–235. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Trachana V, Ntoumou E, Anastasopoulou L
and Tsezou A: Studying microRNAs in osteoarthritis: Critical
overview of different analytical approaches. Mech Ageing Dev.
171:15–23. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Dellago H, Bobbili MR and Grillari J:
MicroRNA-17-5p: At the crossroads of cancer and aging-a
mini-review. Gerontology. 63:20–28. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Jia J, Feng X, Xu W, Yang S, Zhang Q, Liu
X, Feng Y and Dai Z: MiR-17-5p modulates osteoblastic
differentiation and cell proliferation by targeting SMAD7 in
non-traumatic osteonecrosis. Exp Mol Med. 46(e107)2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Li J, Lai Y, Ma J, Liu Y, Bi J, Zhang L,
Chen L, Yao C, Lv W, Chang G, et al: miR-17-5p suppresses cell
proliferation and invasion by targeting ETV1 in triple-negative
breast cancer. BMC Cancer. 17(745)2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zhu Y, Gu J, Li Y, Peng C, Shi M, Wang X,
Wei G, Ge O, Wang D, Zhang B, et al: MiR-17-5p enhances pancreatic
cancer proliferation by altering cell cycle profiles via disruption
of RBL2/E2F4-repressing complexes. Cancer Lett. 412:59–68.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Jin J, Sun Z, Yang F, Tang L, Chen W and
Guan X: miR-19b-3p inhibits breast cancer cell proliferation and
reverses saracatinib-resistance by regulating PI3K/Akt pathway.
Arch Biochem Biophys. 645:54–60. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Li H, Miao D, Zhu Q, Huang J, Lu G and Xu
W: MicroRNA-17-5p contributes to osteoarthritis progression by
binding p62/SQSTM1. Exp Ther Med. 15:1789–1794. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Duan L, Duan D, Wei W, Sun Z, Xu H, Guo L
and Wu X: MiR-19b-3p attenuates IL-1β induced extracellular matrix
degradation and inflammatory injury in chondrocytes by targeting
GRK6. Mol Cell Biochem. 459:205–214. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Camilleri ET, Dudakovic A, Riester SM,
Galeano-Garces C, Paradise CR, Bradley EW, McGee-Lawrence ME, Im
HJ, Karperien M, Krych AJ, et al: Loss of histone methyltransferase
Ezh2 stimulates an osteogenic transcriptional program in
chondrocytes but does not affect cartilage development. J Biol
Chem. 293:19001–19011. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Stazi G, Zwergel C, Mai A and Valente S:
EZH2 inhibitors: A patent review (2014-2016). Expert Opin Ther Pat.
27:797–813. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Wang X, Hu B, Shen H, Zhou H, Xue X, Chen
Y, Chen S, Han Y, Yuan B, Zhao H, et al: Clinical and prognostic
relevance of EZH2 in breast cancer: A meta-analysis. Biomed
Pharmacother. 75:218–225. 2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Chen L, Wu Y, Wu Y, Wang Y, Sun L and Li
F: The inhibition of EZH2 ameliorates osteoarthritis development
through the Wnt/β-catenin pathway. Sci Rep. 6(29176)2016.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Trenkmann M, Brock M, Gay RE, Kolling C,
Speich R, Michel BA, Gay S and Huber LC: Expression and function of
EZH2 in synovial fibroblasts: epigenetic repression of the Wnt
inhibitor SFRP1 in rheumatoid arthritis. Ann Rheum Dis.
70:1482–1488. 2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115.
2011.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Moore BT and Xiao P: MiRNAs in bone
diseases. Microrna. 2:20–31. 2013.PubMed/NCBI View Article : Google Scholar
|