Introduction
Ovarian cancer is the third most common
gynecological cancer worldwide, and has the highest mortality rate
of all gynecological malignancies (1,2). The
5-year survival rate for patients with early ovarian cancer (stage
I or II) is 80-90%, compared with the survival rate of patients
with stage III and IV disease, which is only 25% (3,4). Only
30% of American patients with ovarian cancer are diagnosed in the
early stages in 2016 (3,4). In patients with recurrent ovarian
cancer, treatment is less effective than in patients with a first
time diagnosis of ovarian cancer (5). As the number of relapses increases for
a particular individual, the disease remission period is reduced
(5). At present, there is no
effective preventive strategy for ovarian cancer (5). Interventions that reduce the risk of
ovarian cancer are particularly important given the limited
screening options and high mortality rates, which have not
significantly changed over the past several decades (5). Therefore, ovarian cancer is currently a
major public health concern. The molecular mechanisms underlying
the proliferation, migration and invasion of ovarian cancer cells
require further investigation to identify new biomarkers and
develop targeted drugs.
MicroRNAs (miRNAs/miRs) are non-coding RNAs ~22
nucleotides in length, which are involved in cell homeostasis and
tumorigenesis (6). The abnormal
expression of miRNAs in ovarian cancer is closely associated with
the progression and clinical prognosis of the disease (7,8). A
number of carcinogenic pathways may influence the same miRNA, and
equally, a number of miRNAs may control a post-transcriptional
program that affects numerous target genes (6,7). miRNAs
are often downstream effectors of tumor relevant protein kinases or
mutations, and therefore, targeting miRNAs may be a strategy to
increase drug specificity and overcome drug resistance (6,8).
The function and expression of miR-411 has been
studied in several types of cancer (9-12).
A number of previous studies have reported that miR-411 may
function as an oncogene in osteosarcoma and lung and hepatocellular
carcinoma (9-12).
Contrastingly, a number of studies have reported that miR-411
functions as a tumor suppressor in oral, cervical, breast and
colorectal cancer, as well as renal cell carcinoma and glioma
(13-21).
At present, the role of hsa-mir-411-5p (also known as miR-411-5p)
in the development of ovarian cancer is not completely understood.
However, it has been reported that the expression of hsa-mir-411-3p
in the stable state is low, which is unstable and easy to degrade
(22). The present study explored
the mechanisms of hsa-mir-411-5p in ovarian cancer cells in
vitro.
Materials and methods
Cell culture and transfection
The ovarian cancer cell lines OVCAR-8 and SKOV3 were
purchased from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences and American Type Culture Collection,
respectively. Cells were maintained in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.), at 5% CO2 and 37˚C. 293
cells (The Cell Bank of Type Culture Collection of the Chinese
Academy of Sciences) were cultured in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 4,500 mg/l glucose and 10% FBS.
The OVCAR-8 and SKOV3 cells were transfected with 25 nM negative
control mimic (MISSION® miRNA, Negative Control 1; cat.
no. HMC002; Sigma Aldrich; Merck KGaA) or miR-411-5p mimic (MISSION
microRNA Mimic hsa-miR-411-5p; cat. no. HMI0558; Sigma-Aldrich;
Merck KGaA) at 24 h after seeding, and cells were seeded into
6-well plates at a density of 1x106 cells/well. The
sequences used were as follows: Negative control mimic forward,
5'-[AmC6]UGAACAGUGUUACGUACGAUACC[dT][dT]-3', and reverse
5'-GGUUCGUACGUACACUGUUCA-3'. hsa-miR-411-5p mimic forward
5'-CGUACGCUAUACGGUCUAUCUA[dT][dT]-3', and reverse
5'-UAGUAGACCGUAUAGCGUACG-3'. Transfection was carried out using
Lipofectamine® RNAiMAX Transfection reagent (Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions.
IncuCyte proliferation assay
OVCAR-8 and SKOV3 cells 2,000 cells/well were seeded
into 96-well plates and allowed to adhere at 37˚C and 5%
CO2. After transfection for 12 h with miRNA mimics, cell
proliferation was monitored by analyzing the occupied area (%
confluence) of cell images at 0, 24, 48 and 72 h time points using
the IncuCyte Live Cell Analysis system (Essen BioScience) according
to the manufacturer's instructions.
Cell wound healing assay
OVCAR-8 and SKOV3 cells (1.5x104/well)
were seeded into 96-well ImageLock plates for 24 h in RPMI-1640
medium supplemented with 10% FBS at 37˚C and 5% CO2.
Following serum starvation (RPMI-1640 medium only) for 12 h at 37˚C
and 5% CO2, scratch wounds were made by the IncuCyte™
Cell Migration kit (Essen BioScience), using the 96-pin wound
making tool (WoundMaker). Subsequently, cells were cultured in
RPMI-1640 medium supplemented with 4% FBS for 24 h at 37˚C and 5%
CO2. Images were automatically monitored and analyzed
every 4 h for 24 h by the IncuCyte Live Cell Analysis system (Essen
BioScience; magnification, x10). Data were processed and analyzed
using IncuCyte™ Cell Migration Software Application Module (Essen
BioScience) for 24 h. Data are presented as the wound width.
Cell invasion assay
OVCAR-8 and SKOV3 cells
(~5x104-1x105) were resuspended in serum-free
RPMI-1640 medium and seeded in the top portion of a BioCoat
Matrigel Invasion Chamber (Corning, Inc.). The lower compartment of
the chamber contained RPMI-1640 medium supplemented with 10% FBS as
a chemoattractant. After a 28-h incubation at 37˚C and 5%
CO2, cells on the upper side of the membrane were
removed, washed with PBS and fixed in 50% ethanol for 15 min at
room temperature. Subsequently, cells were stained with Coomassie
brilliant blue for 15 min at room temperature and counted under 10
different microscopic fields using an inverted light microscope
(Leica Microsystems; magnification, x100).
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
To measure miRNA expression, total RNA was isolated
from cultured OVCAR-8 and SKOV3 cells using the mirVana™ miRNA
Isolation kit with phenol (Ambion; Thermo Fisher Scientific, Inc.).
The mature form of miRNAs were detected using the
TaqMan® Advanced miRNA cDNA Synthesis kit with TaqMan
Fast Advanced Master mix (Thermo Fisher Scientific, Inc.). The
hsa-miR-411-5p TaqMan Advanced miRNA assay kit (assay ID,
478086_mir) and the hsa-miR-26a-5p TaqMan Advanced miRNA assay kit
(assay ID, 477995_mir), which was an internal control, were
purchased from Thermo Fisher Scientific, Inc. RT-q PCR was
performed by Applied Biosystems 7900HT Fast Real-time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The relative
expression was calculated by 2-ΔΔCq method (23) and expression levels were normalized
to hsa-miR-26a-5p.
To investigate the effects of miRNA-411-5p
overexpression on the potential miRNA target gene HMMR, total RNA
from OVCAR-8 and SKOV3 cells was isolated using the RNeasy Plus
Mini kit (Qiagen Sciences, Inc.; cat. no. MD 20874) according to
the manufacturer's protocol. RNA was reverse transcribed into cDNA
using the High-Capacity cDNA Reverse Transcription kit (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
PCR was performed in triplicate using SYBR® Select
Master mix (Thermo Fisher Scientific, Inc.) on a 7900HT Fast
Real-time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The primer sequences used were as follows: HMMR forward,
5'-CCAGGTGCTTATGATGTTAAAACT-3' and reverse,
5'-TGAGATTCCTTCTTTGATTCCGA-3'; and β-actin forward,
5'-ATTGGCAATGAGCGGTTCCG-3' and reverse, 5'-CGTGGATGCCACAGGACTCC-3'.
The following thermocycling conditions were used: 95˚C for 5 min;
40 cycles of 95˚C for 10 sec, 60˚C for 20 sec and 72˚C for 20 sec.
Relative expression was calculated using the 2-ΔΔCq
method (23) and expression levels
were normalized to β-actin.
Bioinformatics analysis and Luciferase
reporter assay
starBase (version 2.0; starbase.sysu.edu.cn) online database was used to
predict that HMMR was a target of miRNA-411-5p. The human HMMR
3'UTR fragment (337 bp) was PCR-amplified from the genomic DNA and
subcloned into the pMIR-REPORT luciferase construct (Thermo Fisher
Scientific, Inc.) using the following cloning primers: Forward,
5'-TTGGTCCTACCTATTATCCTTCTA-3'; reverse,
5'-AATGACTTACTGTGTAATTTTATTTC-3'. The mutant HMMR 3'UTR was made
using a QuikChange Site-directed Mutagenesis kit (Agilent
Technologies, Inc.) and confirmed using Sanger sequencing. 293
cells of 60% confluence in 24-well plates were co-transfected with
the pMIR-REPORT firefly luciferase reporter gene construct, the
pRL-TK Renilla luciferase construct (for normalization) and
25 nM miRNA mimics per well using Lipofectamine 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Cell extracts were
prepared 48 h after transfection and the luciferase activity was
measured using the Dual-Luciferase Reporter Assay system (Promega
Corporation).
Western blotting
Total protein from OVCAR-8 and SKOV3 cells was
extracted using RIPA buffer (Beyotime Institute of Biotechnology).
Total protein was quantified using a bicinchoninic acid assay kit
(Beyotime Institute of Biotechnology). 40 µg protein/lane was
separated on a 10% SDS-PAGE gel and then transferred to Hybond
membranes (GE Healthcare). Membranes were blocked for 1 h in 5%
milk in TBST at room temperature. For immunoblotting, membranes
were incubated overnight at 4˚C with antibodies targeted against
HMMR (cat. no. ab124729; 1:1,000; Abcam), phosphorylated-Erk1/2
(cat. no. 4377; 1:1,000; Cell Signaling Technology Europe, B.V.),
Erk1/2 (cat. no. 9102; 1:1,000; Cell Signaling Technology Europe,
B.V.) and GAPDH (cat. no. sc-25778; 1:1,000; Santa Cruz
Biotechnology, Inc.). Subsequently, membranes were incubated with
anti-rabbit or anti-mouse horseradish peroxidase conjugated IgG
(cat. no. 111-035-003 and 115-035-003; 1:20,000; Jackson
ImmunoResearch Laboratories, Inc.) for 1 h at room temperature.
Bands were visualized by ECL-Plus detection reagents (Santa Cruz
Biotechnology, Inc.).
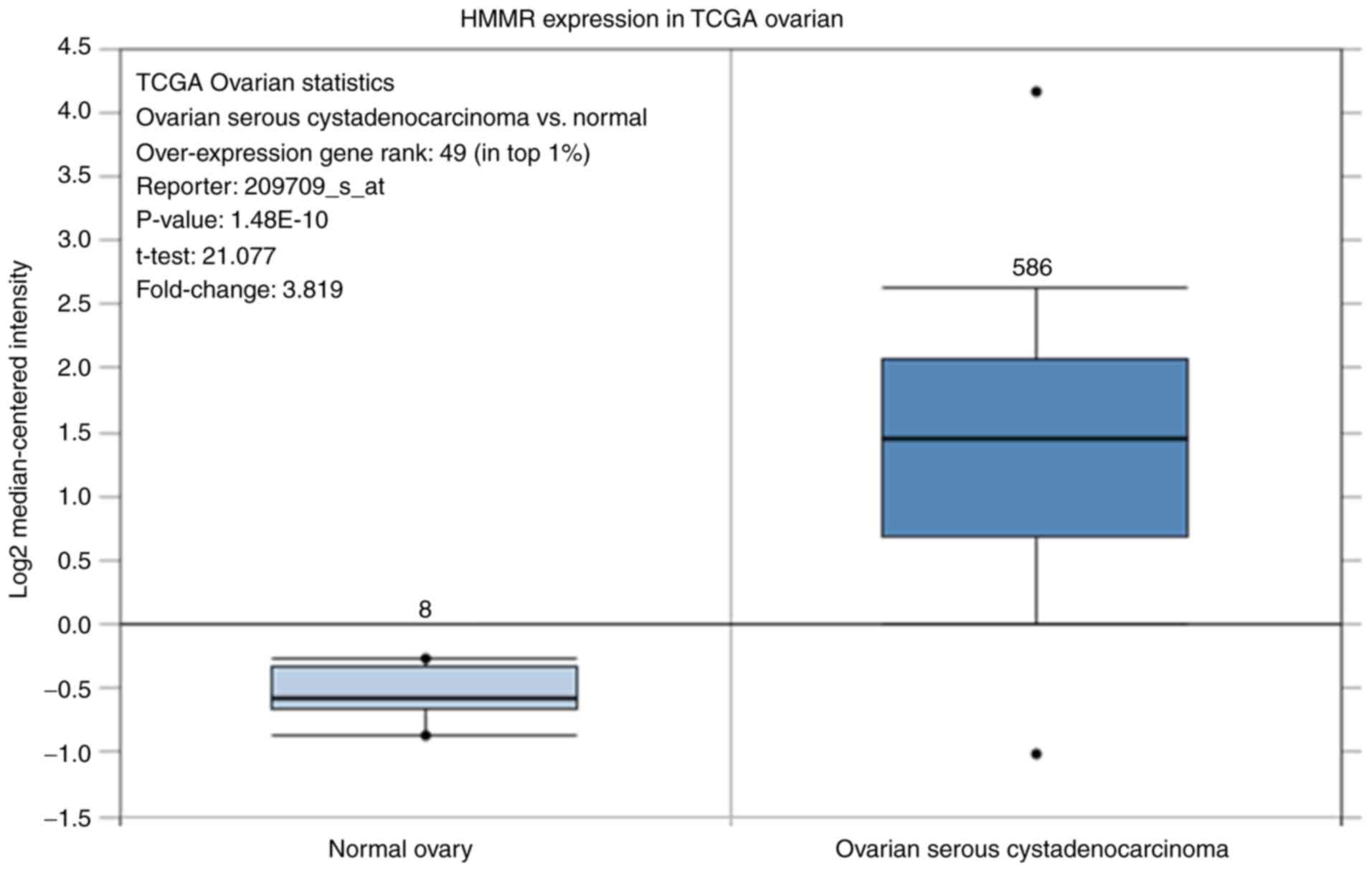
Oncomine gene differential analysis by
Oncomine
Differentially expressed HMMR mRNA between 586
ovarian serous cystadenocarcinoma and 8 normal ovarian tissues was
analyzed by Oncomine (www.oncomine.org). Filters were set up as: Gene, HMMR,
cancer type, ovarian cancer, dataset type, The Cancer Genome Atlas
(TCGA) datasets. The data were grouped by cancer and normal type,
and only samples included in the analysis are shown.
Statistical analysis
SPSS 20.2 software (IBM Corp.) was used to analyze
the data. Data are presented as the mean ± SEM. Differences between
groups were assessed using a paired Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
StarBase (version 2.0; http://starbase.sysu.edu.cn/) online database provided
the correlation analysis between miR-411-5p expression and HMMR
mRNA level. The correlation between miR-411-5p and HMMR was further
investigated using patient sample information from the starBase
online database.
Results
miR-411-5p is a potent suppressor of
ovarian cancer proliferation, migration and invasion
The effects of miR-411-5p on the human ovarian
cancer cell lines OVCAR-8 and SKOV3 were investigated using a
miR-411-5p mimic. OVCAR-8 and SKOV3 cells were cultured and
transfected with the negative control or the miR-411-5p mimic. At
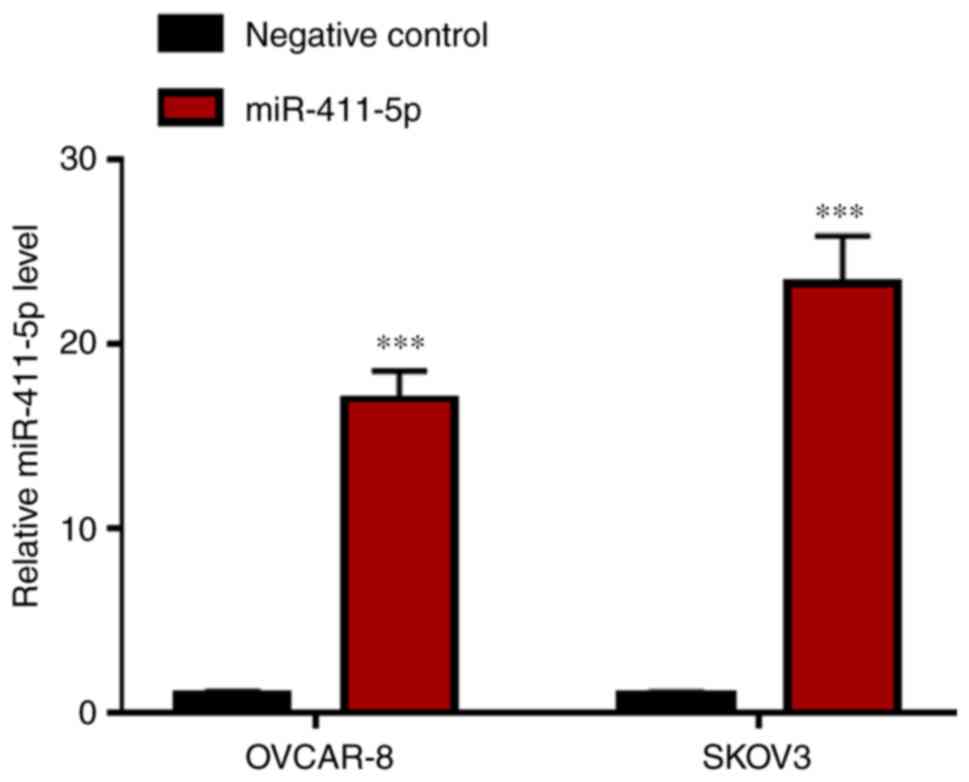
48 h after transfection, the miR-411-5p expression level was
significantly increased (P<0.001) in the miR-411-5p mimic
groups, compared with the negative control groups, measured by
RT-qPCR (Fig. 1). Furthermore,
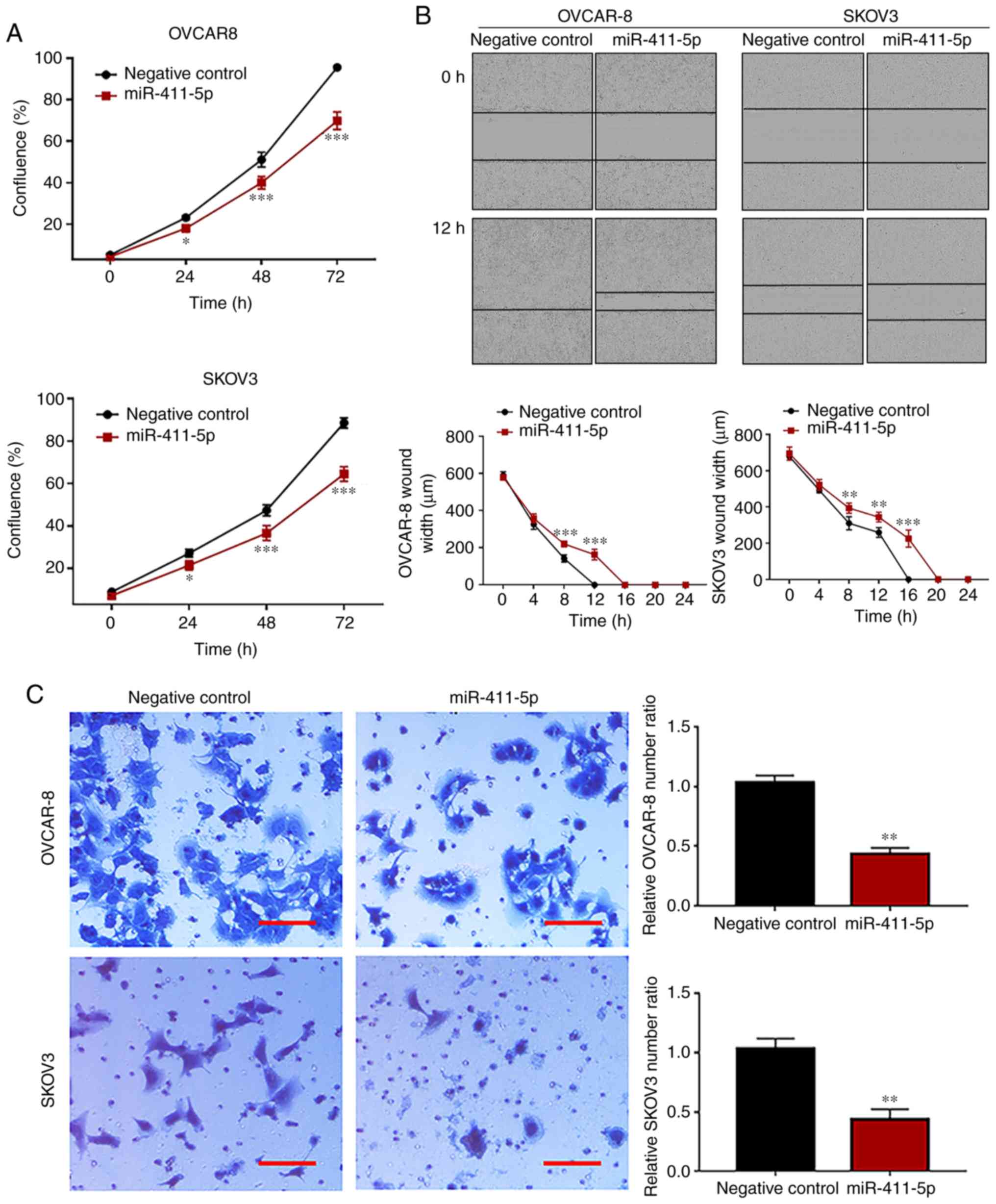
miR-411-5p significantly inhibited cell proliferation (P<0.001)
in both the OVCAR-8 and the SKOV3 cell lines compared with the
negative control, at all time points (Fig. 2A). Wound healing and Matrigel
invasion assays were carried out to evaluate the effect of the
overexpression of miR-411-5p on the migration and invasion of
ovarian cancer cells, respectively. The results of the cell wound
healing assay suggested that wound healing was decreased
(P<0.001) at 8 h (OVCAR-8 and SKOV3), 12 h (OVCAR-8 and SKOV3)
and 16 h (SKOV3) in the miR-411-5p group compared with the negative
control group (Fig. 2B), which
indicated that miR-411-5p suppressed ovarian cancer cell migration.
The results of the Matrigel invasion assay displayed that cell
invasion was significantly suppressed (P<0.01) in the miR-411-5p
group compared with the negative control group (Fig. 2C).
miR-411-5p directly targets HMMR
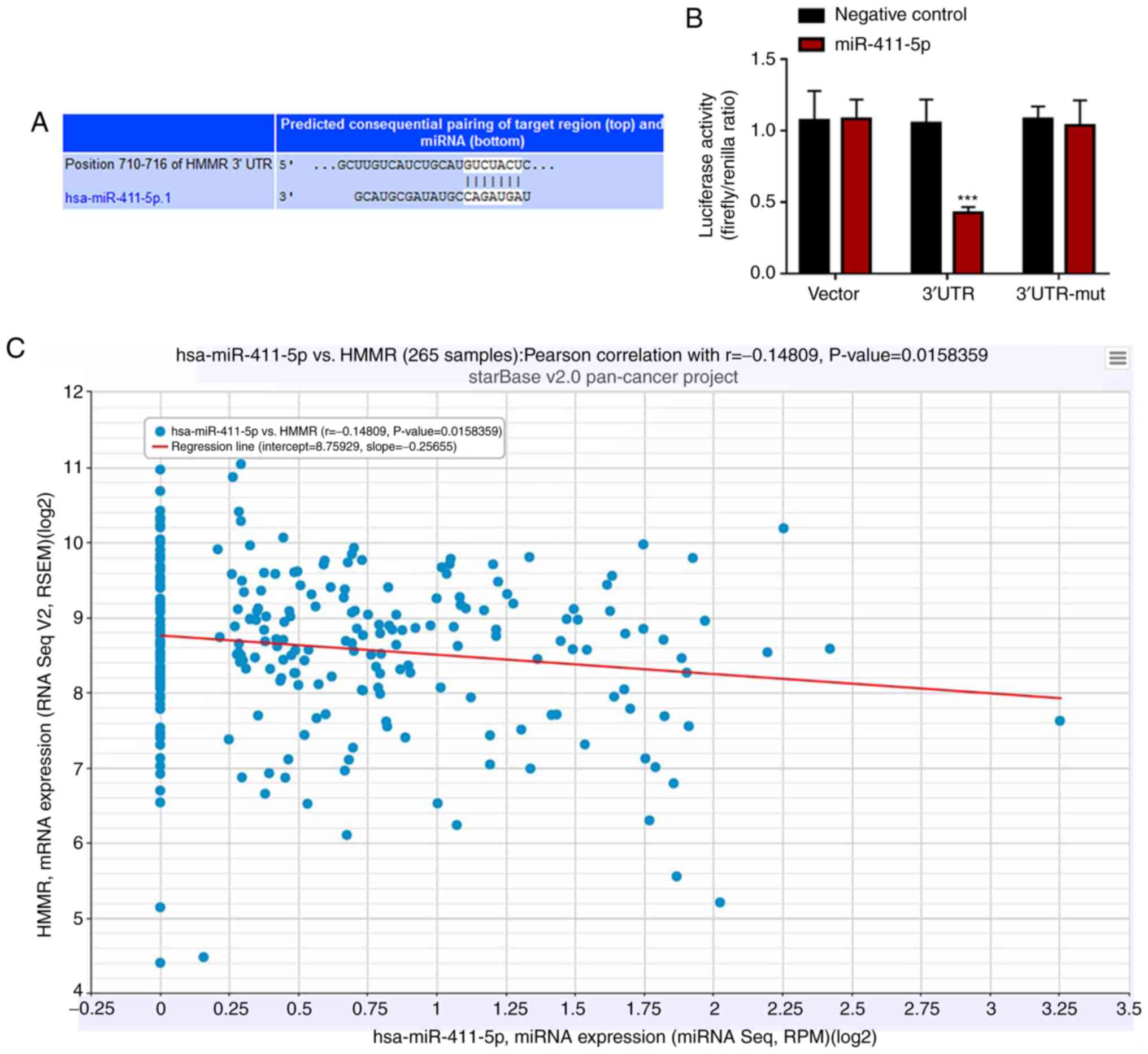
To identify the potential target of miR-411-5p in
ovarian cancer cell lines, starBase (version 2.0; starbase.sysu.edu.cn) online database was used to
predict the candidate downstream target genes. The results
suggested that the 3'-UTR of HMMR contained putative binding sites
for miR-411-5p (Fig. 3A). To further
confirm whether HMMR was the direct target of miR-411-5p, the
3'-UTR of HMMR was synthesized and cloned into a luciferase
reporter plasmid to construct dual luciferase reporter gene
plasmids including the pmiR-HMMR 3'-UTR wild-type and pmiR-HMMR
3'-UTR mutant. The dual luciferase reporter gene plasmids were
co-transfected into 293 cells with the negative control or
miR-411-5p mimic. The results indicated that 293 cells
co-transfected with the pmiR-HMMR 3'-UTR wild-type plasmid and the
miR-411-5p mimic significantly decreased the relative luciferase
activity compared with co-transfection with the negative control
(P<0.001; Fig. 3B). However,
co-transfection with the pmiR vector or pmiR-HMMR 3'-UTR mutant
plasmid plus the miR-411-5p or negative control mimic did not
significantly influence the relative luciferase activity of 293
cells (Fig. 3B). These results
illustrated that HMMR was a direct downstream target of miR-411-5p.
The correlation between miR-411-5p and HMMR was further
investigated using patient sample information from the starBase
online database. miR-411-5p expression from miRNA sequencing
displayed a significant negative correlation with HMMR mRNA level
from RNA sequencing (correlation coefficient r=-0.14809;
P=0.0158359) from 265 ovarian serous cystadenocarcinoma patient
samples (Fig. 3C) using the starBase
Pan-Cancer miRNA-Target Expression Profile (24).
HMMR downregulation mediated by
miR-411-5p overexpression may inhibit ovarian cancer cell
proliferation by downregulating the activity of ERK1/2
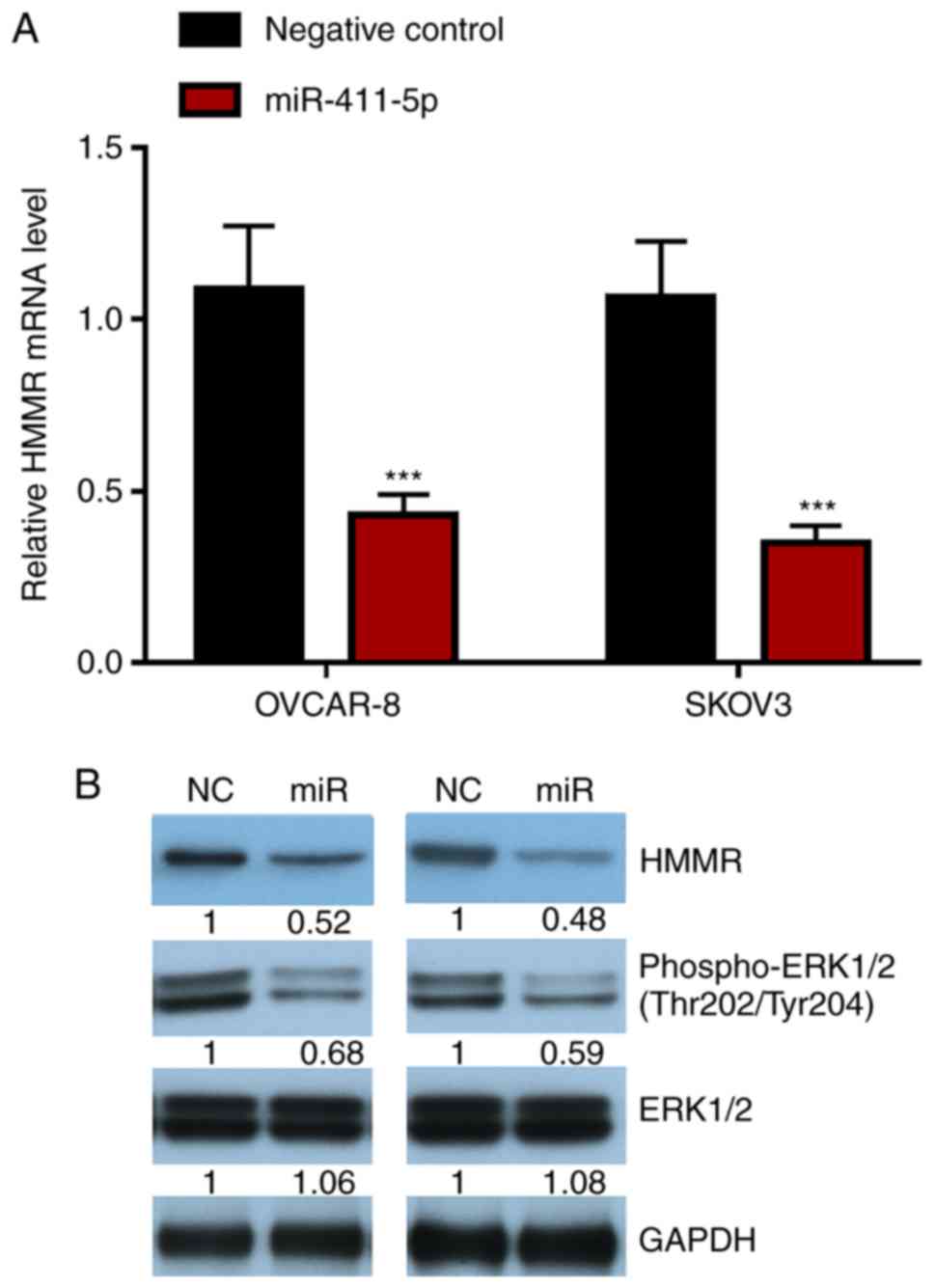
The role of miR-411-5p and HMMR in the proliferation
of ovarian cancer cells was explored. The results suggested that
ovarian cancer cells transfected with the miR-411-5p mimic
displayed reduced HMMR mRNA and protein expression levels, as well
as reduced ERK1/2 signaling pathway activity (Fig. 4A and B). Furthermore, differential HMMR mRNA
expression between paired ovarian serous cystadenocarcinoma and
normal tissue was analyzed by Oncomine. The results suggested that
HMMR mRNA expression in ovarian serous cystadenocarcinoma was
significantly higher (P<0.001) than that in normal ovarian
tissue (Fig. 5).
Discussion
The biological role of miR-411 in ovarian cancer
cells is not well understood (25),
however, it has been reported that miR-411 is downregulated in
ovarian cancer cells (25). To the
best of our knowledge, the present study is the first to
demonstrate the regulatory molecular mechanism of miR-411-5p in the
malignant biological behavior of ovarian cancer cells.
A number of previous studies have reported that
miR-411 plays an important role as an oncogene in tumorigenesis by
negatively regulating protein expression (10,26).
Further studies have reported that miR-411 may function as an
oncogene in osteosarcoma, lung and hepatocellular carcinoma
tumorigenesis and tumor development (9,11,12,26,27).
miR-411 promoted osteosarcoma cell proliferation and migration by
inhibiting the expression of the metastasis suppressor protein 1
(MTSS1) (9). Furthermore, high
miR-411 expression was an independent poor prognostic indicator for
patients with non-small cell lung carcinoma, and compared with
healthy controls, miR-411 expression levels were higher in the
serum of patients with non-small cell lung carcinoma (26). Additionally, other studies have
demonstrated that the oncogenic effect of miR-411 on lung cancer
cell proliferation was mediated by direct downregulation of
forkhead box O1(10). miR-411 also
increased cell migration in lung adenocarcinoma cells, was
upregulated in tumors from patients who relapsed systemically and
was associated with poor survival time following lung
adenocarcinoma resection (11).
Another study indicated that miR-411 promotes lung carcinogenesis
by directly targeting the tumor suppressor genes sprouty RTK
signaling antagonist 4 (SPRY4) and thioredocin interacting protein
(27). Furthermore, it has been
reported that miR-411 promoted hepatocellular carcinoma cell
proliferation by targeting itchy E3 ubiquitin protein ligase (ITCH)
(12).
Alternatively, numerous studies have reported that
miR-411 functions as a tumor suppressor in oral, cervical, breast
and colorectal cancer, as well as in renal cell carcinoma and
glioma (13,14,17,19,20).
Recent study has shown that miR-411-5p was downregulated in oral
cancer compared with non-cancerous tissue (13). Similarly, miR-411 was significantly
downregulated and played a role as a tumor suppressor in renal cell
carcinoma (14). In cervical cancer,
miR-411 acted as a potential tumor suppressor to inhibit cancer
progression by directly targeting STAT3(15). Additionally, miR-411-5p inhibited
proliferation and metastasis of breast cancer cells by directly
targeting growth factor receptor bound protein 2 (GRB2) (17). Zhao et al (18) reported that miRNA-411 inhibited
malignant biological behaviours of colorectal cancer cells by
targeting phosphoinositide-3-kinase regulatory subunit 3 (PIK3R3).
Additionally, miR-411-5p decreased glioma cell proliferation by
targeting topoisomerase II-α (TOPO2A) (19); overexpression of miR-411-5p inhibited
the proliferation of rhabdomyosarcoma cells in vitro and
tumorigenicity in vivo (20);
and miR-411-5p activated p38-mitogen activated protein kinase
phosphorylation by directly downregulating SPRY4 and promoted
apoptosis and myogenic differentiation (20). Furthermore, the miR-379/411 cluster
regulated IL-18, significantly suppressed the invasive capacity of
MESO1 cells (a tumor model) and contributed to drug resistance in
malignant pleural mesothelioma (21).
A number of targets of miR-411 have been reported
previously, including MTSS1(9),
TOP2A (19), ITCH (12), STAT3(15), specificity protein 1(16), GRB2(17), PIK3R3(18), SPRY4(20), IL-18(21), hypoxia inducible factor 1-α (28) and matrix metalloprotease-13(29). In the present study, HMMR was
identified as a novel direct target of miR-411-5p by a luciferase
reporter assay. HMMR is required for bipolar spindle assembly and
mitotic progression by integrating biochemical and structural
pathways (30). HMMR expression is
associated with poor prognosis and metastasis in non-small cell
lung carcinoma (31). Furthermore,
upregulation of HMMR in lung adenocarcinoma was associated with an
inflammatory molecular signature and poor prognosis (32). Cell surface HMMR is an hyaluronic
acid-binding protein that is not highly expressed in normal tissues
but is commonly upregulated in a number of advanced forms of cancer
(33,34). However, the underlying mechanism of
HMMR in ovarian cancer is not completely understood. Using TCGA,
the present study suggested that HMMR mRNA expression is higher in
ovarian serous cystadenocarcinoma than in normal ovarian
tissue.
ERK1/2 kinases are ubiquitous and homologous mitogen
activated protein kinases that mediate ligand-stimulated signals
for the induction of proliferation, differentiation, survival,
apoptosis, angiogenesis and motility (35). Upregulation and elevated activation
of ERK1/2 kinases is common in human tumors (36-38).
Indeed, studies have reported that in a large number of different
types of cancer the ERK1/2 signaling pathway is constitutively
active and may be involved in the pathogenesis of these tumors
(36-38).
Of note, the ERK signaling pathway plays a critical role in ovarian
cancer pathogenesis and downregulation of this signaling pathway is
highly effective for the inhibition of ovarian tumor proliferation
(39). Furthermore, the nuclear
localization of phosphorylated ERK1/2 can serve as a marker for the
progression of ovarian cancer (39,40).
HMMR is a known modulator of ERK1/2 in certain types
of cancer (41,42). HMMR overexpression resulted in
activation of the ERK1/2 signaling pathway, which promotes cell
proliferation of osteoblastic cells (41). Cell surface HMMR and CD44 act
together and coordinate to sustain ERK1/2 signaling, leading to
high basal motility of invasive breast cancer cells (42). The results of the present study
suggested that the expression of phosphorylated ERK1/2 and HMMR is
decreased following the overexpression of miR-411-5p in ovarian
cancer cells, a result that is consistent with the aforementioned
studies. The present study suggested a potential interplay between
HMMR and ERK1/2, via phosphorylated ERK1/2, which may contribute to
cancer cell proliferation and motility modulation.
The present study indicated that the overexpression
of miR-411-5p could suppress ovarian cancer cell (OVCAR-8 and
SKOV3) proliferation and motility. In the present study, it was
also described that miR-411-5p could downregulate HMMR, which
resulted in the downregulation of the ERK1/2 signaling pathway
activity, and thus inhibited ovarian cancer cell proliferation and
motility. In different cancer types, miR-411-5p may have different
roles (12,19). In conclusion, the results of the
present study demonstrated that miR-411-5p may be potential
therapeutic target for ovarian cancer.
One limitation of the present study was that the
results were based on miRNA mimic-mediated overexpression
experiments in two ovarian cancer cell lines, where the miRNA level
may be much higher than the endogenous miRNA level. Additionally,
the wound healing should ideally be performed in 0% FBS, therefore
the use of 4% FBS is a minor limitation. Future investigations, in
particular in vivo experiments, should be performed to
confirm the functional mechanisms and investigate the clinical
application of miR-411-5p. Collectively, the present study provided
a rationale for further investigation into miR-411-5p-based
diagnosis, prognosis and targeted therapeutic strategies.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Liaoning
Province Science and Technology Foundation (grant no. 2019-ZD-0984)
and the Shenyang Science and Technology Grant (grant no.
17-230-9-41).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FH, DZ and XN conceived and designed the study and
drafted the manuscript. FH, DZ, CL, JN and XN performed the
experiments. All authors analyzed and interpreted the data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Suh DH, Kim M, Kim K, Kim HJ, Lee KH and
Kim JW: Major clinical research advances in gynecologic cancer in
2016: 10-year special edition. J Gynecol Oncol.
28(e45)2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Chornokur G, Amankwah EK, Schildkraut JM
and Phelan CM: Global ovarian cancer health disparities. Gynecol
Oncol. 129:258–264. 2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714.
2009.PubMed/NCBI View
Article : Google Scholar
|
|
7
|
Zaman MS, Maher DM, Khan S, Jaggi M and
Chauhan SC: Current status and implications of microRNAs in ovarian
cancer diagnosis and therapy. J Ovarian Res. 5(44)2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Langhe R: microRNA and ovarian cancer. Adv
Exp Med Biol. 889:119–151. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Xu N, Yang W, Liu Y, Yan F and Yu Z:
MicroRNA-411 promoted the osteosarcoma progression by suppressing
MTSS1 expression. Environ Sci Pollut Res Int. 25:12064–12071.
2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhao Z, Qin L and Li S: miR-411
contributes the cell proliferation of lung cancer by targeting
FOXO1. Tumour Biol. 37:5551–5560. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Nadal E, Zhong J, Lin J, Reddy RM, Ramnath
N, Orringer MB, Chang AC, Beer DG and Chen G: A MicroRNA cluster at
14q32 drives aggressive lung adenocarcinoma. Clin Cancer Res.
20:3107–3117. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Xia K, Zhang Y, Cao S, Wu Y, Guo W, Yuan W
and Zhang S: miR-411 regulated ITCH expression and promoted cell
proliferation in human hepatocellular carcinoma cells. Biomed
Pharmacother. 70:158–163. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zeljic K, Jovanovic I, Jovanovic J, Magic
Z, Stankovic A and Supic G: MicroRNA meta-signature of oral cancer:
Evidence from a meta-analysis. Ups J Med Sci. 123:43–49.
2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhang X, Zhang M, Cheng J, Lv Z, Wang F
and Cai Z: MiR-411 functions as a tumor suppressor in renal cell
cancer. Int J Biol Markers. 32:e454–e460. 2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Shan D, Shang Y and Hu T: MicroRNA-411
inhibits cervical cancer progression by directly targeting STAT3.
Oncol Res. 27:349–358. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Guo L, Yuan J, Xie N, Wu H, Chen W, Song S
and Wang X: miRNA-411 acts as a potential tumor suppressor miRNA
via the downregulation of specificity protein 1 in breast cancer.
Mol Med Rep. 14:2975–2982. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang Y, Xu G, Liu G, Ye Y, Zhang C, Fan
C, Wang H, Cai H, Xiao R, Huang Z and Luo Q: miR-411-5p inhibits
proliferation and metastasis of breast cancer cell via targeting
GRB2. Biochem Biophys Res Commun. 476:607–613. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhao J, Xu J and Zhang R: MicroRNA-411
inhibits malignant biological behaviours of colorectal cancer cells
by directly targeting PIK3R3. Oncol Rep. 39:633–642.
2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Deguchi S, Katsushima K, Hatanaka A,
Shinjo K, Ohka F, Wakabayashi T, Zong H, Natsume A and Kondo Y:
Oncogenic effects of evolutionarily conserved noncoding RNA
ECONEXIN on gliomagenesis. Oncogene. 36:4629–4640. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Sun M, Huang F, Yu D, Zhang Y, Xu H, Zhang
L, Li L, Dong L, Guo L and Wang S: Autoregulatory loop between
TGF-β1/miR-411-5p/SPRY4 and MAPK pathway in rhabdomyosarcoma
modulates proliferation and differentiation. Cell Death Dis.
6(e1859)2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yamamoto K, Seike M, Takeuchi S, Soeno C,
Miyanaga A, Noro R, Minegishi Y, Kubota K and Gemma A: MiR-379/411
cluster regulates IL-18 and contributes to drug resistance in
malignant pleural mesothelioma. Oncol Rep. 32:2365–2372.
2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kozomara A, Birgaoanu M and
Griffiths-Jones S: miRBase: From microRNA sequences to function.
Nucleic Acids Res. 47:D155–D162. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yang JH, Li JH, Shao P, Zhou H, Chen YQ
and Qu LH: starBase: A database for exploring microRNA-mRNA
interaction maps from Argonaute CLIP-Seq and Degradome-Seq data.
Nucleic Acids Res. 39:D202–D209. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kim YW, Kim EY, Jeon D, Liu JL, Kim HS,
Choi JW and Ahn WS: Differential microRNA expression signatures and
cell type-specific association with Taxol resistance in ovarian
cancer cells. Drug Des Devel Ther. 8:293–314. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang SY, Li Y, Jiang YS and Li RZ:
Investigation of serum miR-411 as a diagnosis and prognosis
biomarker for non-small cell lung cancer. Eur Rev Med Pharmacol
Sci. 21:4092–4097. 2017.PubMed/NCBI
|
|
27
|
Zhang C, Wang H, Liu X, Hu Y, Ding L,
Zhang X, Sun Q and Li Y: Oncogenic microRNA-411 promotes lung
carcinogenesis by directly targeting suppressor genes SPRY4 and
TXNIP. Oncogene. 38:1892–1904. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ai P, Shen B, Pan H, Chen K, Zheng J and
Liu F: MiR-411 suppressed vein wall fibrosis by downregulating
MMP-2 via targeting HIF-1α. J Thromb Thrombolysis. 45:264–273.
2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang G, Zhang Y, Zhao X, Meng C, Ma L and
Kong Y: MicroRNA-411 inhibited matrix metalloproteinase 13
expression in human chondrocytes. Am J Transl Res. 7:2000–2006.
2015.PubMed/NCBI
|
|
30
|
Chen H, Mohan P, Jiang J, Nemirovsky O, He
D, Fleisch MC, Niederacher D, Pilarski LM, Lim CJ and Maxwell CA:
Spatial regulation of Aurora A activity during mitotic spindle
assembly requires RHAMM to correctly localize TPX2. Cell Cycle.
13:2248–2261. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
31
|
Wang D, Narula N, Azzopardi S, Smith RS,
Nasar A, Altorki NK, Mittal V, Somwar R, Stiles BM and Du YN:
Expression of the receptor for hyaluronic acid mediated motility
(RHAMM) is associated with poor prognosis and metastasis in
non-small cell lung carcinoma. Oncotarget. 7:39957–39969.
2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Stevens LE, Cheung WKC, Adua SJ,
Arnal-Estapé A, Zhao M, Liu Z, Brewer K, Herbst RS and Nguyen DX:
Extracellular matrix receptor expression in subtypes of lung
adenocarcinoma potentiates outgrowth of micrometastases. Cancer
Res. 77:1905–1917. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Turley EA, Noble PW and Bourguignon LY:
Signaling properties of hyaluronan receptors. J Biol Chem.
277:4589–4592. 2002.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Adamia S, Maxwell CA and Pilarski LM:
Hyaluronan and hyaluronan synthases: Potential therapeutic targets
in cancer. Curr Drug Targets Cardiovasc Haematol Disord. 5:3–14.
2005.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Roskoski R Jr: ERK1/2 MAP kinases:
Structure, function, and regulation. Pharmacol Res. 66:105–143.
2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Holmström TH, Tran SE, Johnson VL, Ahn NG,
Chow SC and Eriksson JE: Inhibition of mitogen-activated kinase
signaling sensitizes HeLa cells to Fas receptor-mediated apoptosis.
Mol Cell Biol. 19:5991–6002. 1999.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Huang C, Jacobson K and Schaller MD: MAP
kinases and cell migration. J Cell Sci. 117:4619–4628.
2004.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Mansour SJ, Resing KA, Candi JM, Hermann
AS, Gloor JW, Herskind KR, Wartmann M, Davis RJ and Ahn NG:
Mitogen-activated protein (MAP) kinase phosphorylation of MAP
kinase kinase: Determination of phosphorylation sites by mass
spectrometry and site-directed mutagenesis. J Biochem. 116:304–314.
1994.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Steinmetz R, Wagoner HA, Zeng P, Hammond
JR, Hannon TS, Meyers JL and Pescovitz OH: Mechanisms regulating
the constitutive activation of the extracellular signal-regulated
kinase (ERK) signaling pathway in ovarian cancer and the effect of
ribonucleic acid interference for ERK1/2 on cancer cell
proliferation. Mol Endocrinol. 18:2570–2582. 2004.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Amsterdam A, Shezen E, Raanan C, Schreiber
L, Prus D, Slilat Y, Ben-Arie A and Seger R: Nuclear localization
of phosphorylated ERK1 and ERK2 as markers for the progression of
ovarian cancer. Int J Oncol. 39:649–656. 2011.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Hatano H, Shigeishi H, Kudo Y, Higashikawa
K, Tobiume K, Takata T and Kamata N: Overexpression of receptor for
hyaluronan-mediated motility (RHAMM) in MC3T3-E1 cells induces
proliferation and differentiation through phosphorylation of
ERK1/2. J Bone Miner Metab. 30:293–303. 2012.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hamilton SR, Fard SF, Paiwand FF, Tolg C,
Veiseh M, Wang C, McCarthy JB, Bissell MJ, Koropatnick J and Turley
EA: The hyaluronan receptors CD44 and Rhamm (CD168) form complexes
with ERK1,2 that sustain high basal motility in breast cancer
cells. J Biol Chem. 282:16667–16680. 2007.PubMed/NCBI View Article : Google Scholar
|