Introduction
Cardiac fibrosis, characterized as excessive
deposition of extracellular matrix (ECM) in the perivascular and
interstitial region of the heart, is a common pathological
manifestation involved in multiple cardiac diseases, including
hypertension, myocardial infarction and valvular heart disease
(1-3).
Cardiac fibroblasts (CFs) are the predominant cell type in the
myocardium, and are the primary producers of ECM (2,4). During
cardiac remodeling, the important effect of CFs on cardiac fibrosis
is the differentiation to a myofibroblast phenotype in response to
various stimuli, which results in a markedly decreased
proliferation, migration and secretory ability, and notably
distinguished expression of α-smooth muscle actin (α-SMA) (5,6). Cardiac
fibrosis contributes to the excessive activation of myofibroblasts,
leading to myocardial stiffness, cardiac hypertrophy, and the
destruction of physiological cardiac tissue and impaired cardiac
functions, which causes ventricular remodeling, heart failure and
even cardiac death (3,7).
MicroRNAs (miRNAs) are endogenous and highly
conserved RNA sequences, that are 20-23 nucleotide in length, which
act as negative regulators of gene expression by promoting the
degradation or inhibiting the translation of target mRNAs (8-10).
Notably, miRNAs act as important modulators in the development and
progression of cardiac fibrosis, particularly regarding cell
proliferation, migration of CFs and the transformation to
myofibroblasts (11-13).
Angiotensin II (Ang II) plays a critical role in cardiac fibrosis
through inducing the production of collagens, other ECM proteins
and activation of the pro-fibrogenic cascade (14,15).
Notably, Ang II is involved in the pathogenesis of cardiac fibrosis
with an array of gene expression changes (16,17).
Among them, miRNAs may provide new insights. Previous studies have
demonstrated that miR-144 was involved in multiple types of human
malignancy through comprehensive meta-analyses of miRNA expression
microarrays (18-21).
There is a accumulating evidence to suggest that miR-144 plays an
important role in attenuating the proliferation and migration of
various cancer cells (22-24).
Recently, miR-144 has been reported to mediate the effect of ROCK1
inhibition on the decreased lung endothelial hyperpermeability
induced by LPS (25). miR-144
deficiency interrupts ECM remodeling characterized by increased
cardiac collagen content associated with changes in Zeb1/LOX1 axis
(26) and decreases left ventricular
remodeling following myocardial infarction leading to worsened
cardiac function (27). However, the
potential role and precise molecular mechanism underlying miR-144
in the process of cardiac fibrosis remains unknown.
In the present study, it was observed that miR-144
is downregulated both in the heart of TAC-induced mouse models and
in the CFs administered Ang II. The proliferation and migration
ability of CFs was attenuated by miR-144 overexpression.
Furthermore, miR-144 inhibited the differentiation of CFs into
myofibroblasts, which was characterized by the decreased expression
of collagen-I, collagen-III, CTGF, fibronectin and α-SMA. The
results from the present study further demonstrated that miR-144
directly targeted CREB and decreased its expression levels. By
contrast, such effects could be reversed by miR-144 knockdown. The
present study implied that overexpression of miR-144 could be a
novel therapeutic strategy for the treatment of cardiac
fibrosis.
Materials and methods
Mouse model of TAC
Cardiac hypertrophy and fibrosis was induced by
pressure overload via TAC in accordance with previously described
methods (28). Briefly, male C57BL/6
mice aged 8-10 weeks were anesthetized with pentobarbital sodium
(50 mg/kg) via intraperitoneal injection. After the chest was
opened and the thoracic aorta was exposed, TAC was performed by
inducing a 7-0 silk suture placed around the thoracic aorta and
tied around a 26-gauge blunt needle. Subsequently, the needle was
immediately removed and the thoracic cavity was closed. Sham
surgery animals underwent the same procedure but without the tying
of the suture around the aorta. The total number of mice used in
our study is 15, 6 for sham group and 9 for the surgery procedure;
meanwhile, the number of rat for CFs isolation is 12. All the
procedures involving animals experiments were performed according
to the Guide for the Care and Use of Laboratory Animals published
by the U.S. National Institutes of Health and were approved by the
Animal Care and Use Committee of the Dezhou People's Hospital.
Tissue collection and Histological
analysis
Hearts from mice subjected to sham or TAC for 4
weeks were excised after anesthetization with pentobarbital sodium
(50 mg/kg) via intraperitoneal injection and arrested in diastole
with 10% potassium chloride solution, fixed by 10% paraformaldehyde
and embedded in paraffin. Subsequently, these hearts were sectioned
transversely close to the apex to visualize the left and right
ventricles at 5 µm. Sections of each heart at the mid-papillary
muscle level were stained with hematoxylin-eosin (HE) for
histopathology or with picrosirius red (PSR) for the collagen
deposition.
CF isolation and culture
Neonatal rat CFs were isolated from 1-3-day-old
Sprague-Dawley (SD) rats. The rats were anesthetized with
pentobarbital sodium via intraperitoneal injection (50 mg/kg) and
the hearts were removed and quickly dissected. The heart samples
were washed in phosphate-buffered saline to remove blood and
impurities and then digested with pancreatic enzyme and collagenase
type II. Subsequently, pooled cell suspensions were centrifuged and
resuspended in Dulbecco's modified Eagle's medium (DMEM;
Sigma-Aldrich; Merck KGaA) containing 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc) for 60 min at 37˚C in
humidified air with 5% CO2, which allowed for
preferential attachment of fibroblasts to the bottom of the culture
flasks. Flasks were washed twice with PBS in order to remove the
weakly attached and non-adherent cells, whereas the CFs had
attached onto the culture plates. CFs were passaged when the cell
confluence achieved 70-80%, and the second or third passages were
used in the present study. In addition, CFs were starved for 24 h
in serum-free medium prior to treatment.
Cell transfection
The synthetic miR-144 analogs and antagomir were
synthesized by GenePharma. CF transfection was performed using a
riboFEC CP Transfection kit (Guangzhou RiboBio Co., Ltd.),
according to the manufacturer's protocol. CFs were transfected with
miR-144 mimic and antagomir, as well as the corresponding controls
at a concentration of 50 nM. After 6 h, the transfected CFs were
treated with Ang II (100 nM; Sigma-Aldrich; Merck KGaA) in
serum-free medium for 24 h in order to induce cardiac fibrosis
in vitro.
Cell proliferation and migration
A Cell Counting Kit-8 (CCK-8) (Dojindo Molecular
Technologies, Inc.) was used to assay CF proliferation. CFs
transfected with the miR-144 mimic were serum-starved in serum-free
media for 24 h. The media was then placed with a mixture of fresh
serum-free DMEM and CCK-8 reagent. The CCK-8 assay was performed
after 12 h of incubation. The optical density was determined using
a microplate reader (Bio-Rad) at a wavelength of 450 nm. Cell
migration ability was analyzed using the Transwell chamber assay.
CFs were starved for 24 h and then the cells were placed in the
upper chamber of an insert (pore size, 8 µm). After 24 h of
incubation, the cells on the underside were fixed with 4%
paraformaldehyde for 20 min, and stained with 0.1% crystal violet
in 20% ethanol for 10 min. Images were captured using a phase
contrast microscope.
Reverse transcription-quantitative PCR
and western blotting
Total RNA from cultured CFs was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and then reverse-transcribed using a Transcriptor First
Strand cDNA synthesis kit (Roche Diagnostics) as previously
described (29). The expression
levels of target genes were quantified via real-time PCR using a
LightCycler 480 SYBR Green 1 Master Mix and a Light-Cycler 480 qPCR
system (Roche Diagnostics). The relative transcription levels of
the target genes were normalized GAPDH, while the level of miR-144
was normalized to U6 level. The primers for miR-144 (cat. no.
HmiRQP0190) were obtained from GeneCopoeia and the other primers
are listed in the Table I. Total
protein was extracted from the cultured CFs and the concentration
of the proteins was determined using a BCA assay. Equal
concentrations of proteins were separated via SDS-PAGE (10% gel)
and then transferred onto PVDF membranes. After blocking with 5%
non-fat dried milk in TBS for 1 h, the membranes were incubated
with primary antibodies. Membranes were then incubated with a
secondary antibody and treated with enhanced chemiluminescence
reagent. Images were captured using a Molecular Imager ChemiDoc™
XRS+ and quantified with Image Lab™ Software (version 5.1). The
expression levels of specific proteins were normalized against
GAPDH. The associated antibodies are listed in Table II.
 | Table IAntibody for immunofluorescence. |
Table I
Antibody for immunofluorescence.
| Antibody | Cat. no. | Manufacturer | Sources of
species |
|---|
| Primary | | | |
|
α-SMA | ab7817 | Abcam | Mouse |
| Secondary | | | |
|
Alexa Fluor
568-conjugated donkey anti-mouse IgG | A10037 | Invitrogen; Thermo
Fisher Scientific, Inc. | Mouse |
| Table IIAntibodies used for immunoblot
analysis. |
Table II
Antibodies used for immunoblot
analysis.
| Antibody | Cat. no. | Manufacturer | Sources of
species |
|---|
| Primary | | | |
|
CREB | ab32515 | Abcam | Rabbit |
|
GAPDH | 2118 | Cell Signaling
Technology, Inc. | Rabbit |
| Secondary | | | |
|
IRDye800CW
Conjugated Goat (polyclonal) Anti-Rabbit IgG (H+L) |
bs-40295G-IRDye8 | LI-COR
Biosciences | Rabbit |
Immunofluorescence staining
The cells were fixed with 3.7% formaldehyde in PBS
for 15 min at room temperature, and permeabilized with 0.1% Triton
X-100 in PBS for 40 min. Subsequently, the cell slides were
incubated overnight with the α-SMA (1:100 dilution) at 4˚C. After
rewarming at 37˚C for 1 h, the associated secondary antibody was
used. The nuclei were stained with DAPI and the cell size was
measured using Image Pro Plus software (version 6.0). The
antibodies used are listed in Table
III.
| Table IIIThe primers for reverse transcription
PCR. |
Table III
The primers for reverse transcription
PCR.
| Primer | Sequence
(5'-3') |
|---|
| α-SMA-F |
ACGATGGAAACTACCGTGGAG |
| α-SMA-R |
TTGAAGGCCAATGACGTGCT |
| Collagen
I-F |
CCTCAAGGGCTCCAACGAG |
| Collagen
I-R |
TCAATCACTGTCTTGCCCCA |
| Collagen
III-F |
ACGTAGATGAATTGGGATGCAG |
| Collagen
III-R |
GGGTTGGGGCAGTCTAGTC |
| CTGF-F |
TGACCCCTGCGACCCACA |
| CTGF-R |
TACACCGACCCACCGAAGACACAG |
|
Fibronectin-F |
CCGGTGGCTGTCAGTCAGA |
|
Fibronectin-R |
CCGTTCCCACTGCTGATTTATC |
| GAPDH-F |
GGTGGACCTCATGGCCTACA |
| GAPDH-R |
CTCTCTTGCTCTCAGTATCCTTGCT |
| U6-F |
CTCGCTTCGGCAGCACA |
| U6-R |
AACGCTTCACGAATTTGCGT |
Luciferase assay
For the luciferase assays, HEK293 cells purchased
from Type Culture Collection of the Chinese Academy of Sciences
were cultured in 24-well culture plates and co-transfected with a
pEZX-MT01 vector containing wild-type or mutant CREB 3'-UTR
reporters and 50 nm of miR-144 using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). Luciferase activities
were measured 48 h after transfection with the application of the
Dual luciferase Reporter Assay System (Promega Corporation)
according to the manufacturer's protocol.
Statistical analysis
All statistical data were analyzed using SPSS
software (version 19.0; IBM) and are presented as the mean ±
standard deviation. Differences between two groups were analyzed
using Student's t-tests, while differences among multiple groups
were analyzed by one-way ANOVA followed by a Bonferroni post hoc
test or Tamhane's T2 post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-144 is closely associated with
cardiac fibrosis
In order to investigate the potential role of
miR-144 in the pathogenesis of cardiac fibrosis, the mice were
subjected to TAC and cultured CFs administered with Ang II in the
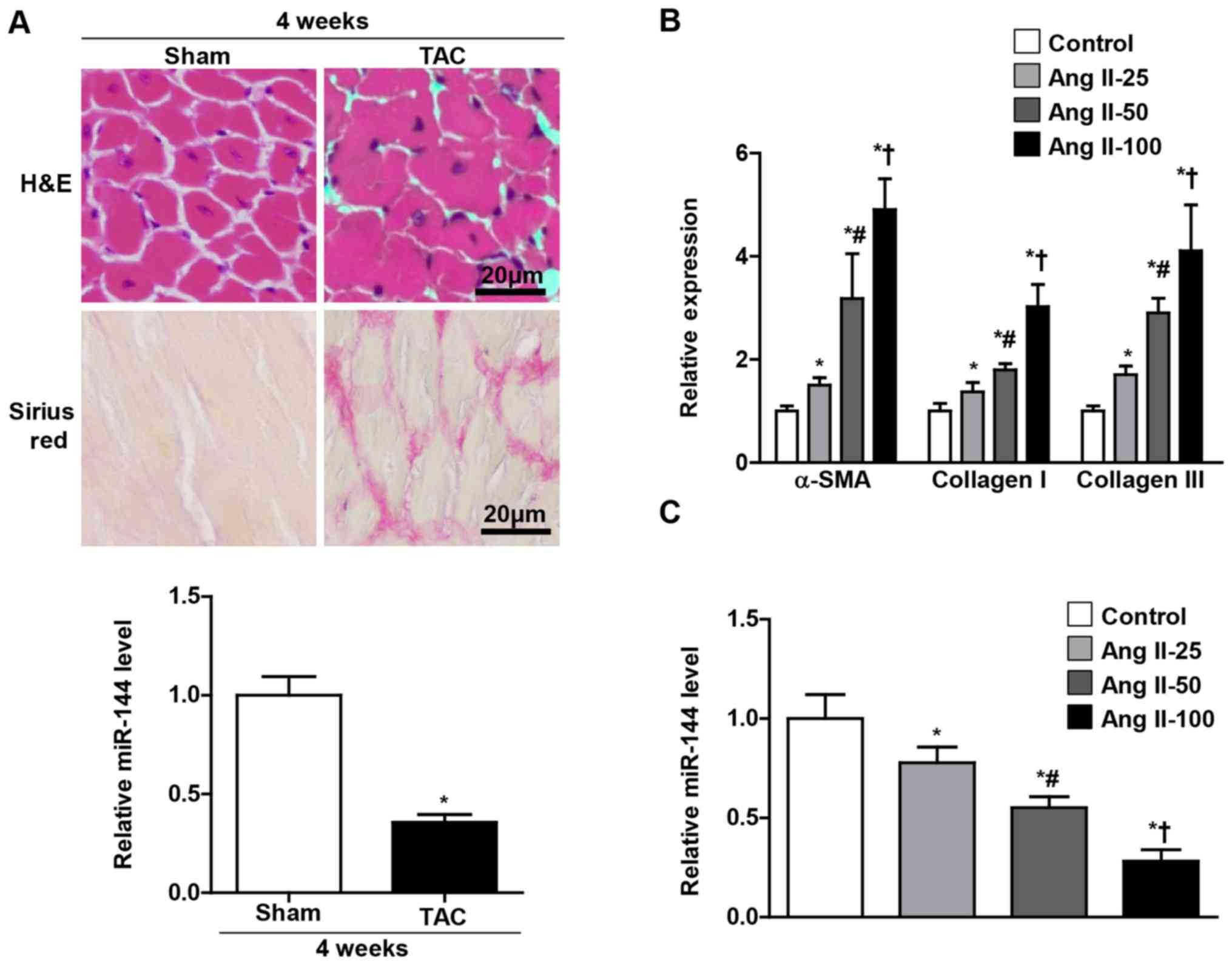
present study. As presented in Fig.
1A, the miR-144 expression levels were significantly decreased
in the heart of the TAC group, which exhibited a larger cell size
and accumulation of fibrosis than the sham group. Furthermore, it
was observed in the present study that the miR-144 expression in
the cultured CFs progressively decreased as the concentration of
Ang II increased (Fig. 1C), and the
level of α-SMA, collagen 1 and collagen III were reversely
increased (Fig. 1B).
Overexpression of miR-144 attenuates
the proliferation and migration of CFs
The CF activation underlying cardiac fibrosis is
characterized by enhanced proliferation and migration ability, as
well as the differentiation into myofibroblasts positive for α-SMA
expression (1,6). In order to verify the effect of
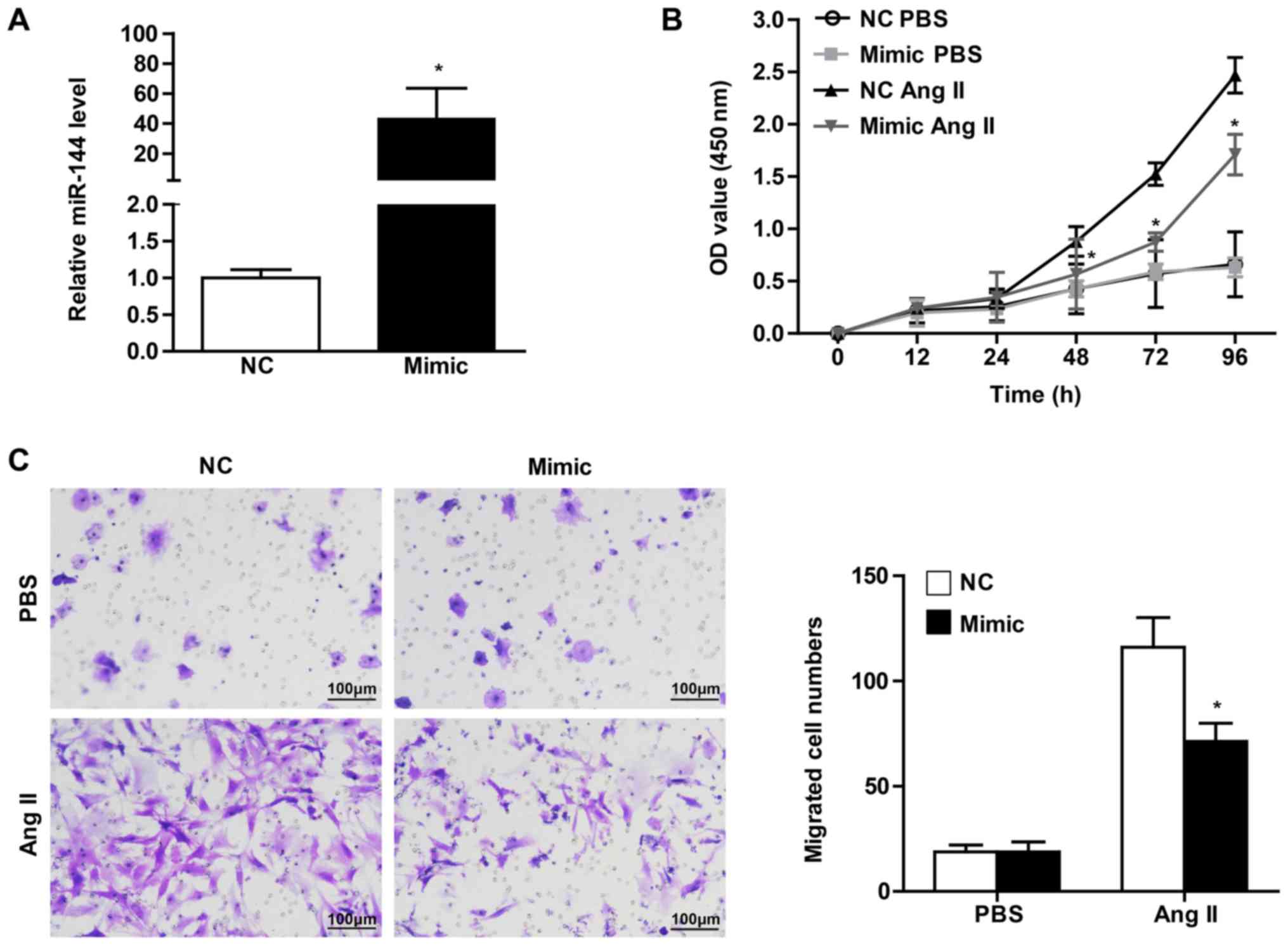
significant miR-144 change on CF activation, the present study
transfected the CFs with miR-144 mimic and the corresponding normal
control. It was observed that the CFs infected with miR-144 mimic
demonstrated a dramatically increased expression level, by ~40-fold
(Fig. 2A). CCK8 assay was performed
in order to evaluate the effect of miR-144 on the proliferation of
CFs. The results revealed that cell vitality of CFs infected with
miR-144 mimics was significantly attenuated upon Ang II
stimulation, whereas there was no difference in the groups treated
with PBS (Fig. 2B). Meanwhile, the
effect of miR-144 on the migration of CFs demonstrated results that
were consistent with Ang II administration (Fig. 2C).
miR-144 inhibits Ang II-induced
myofibroblast differentiation into CFs
As aforementioned, the important change in CFs
activation underlying cardiac fibrosis was the differentiation into
myofibroblasts, which exhibit characteristics of smooth muscle cell
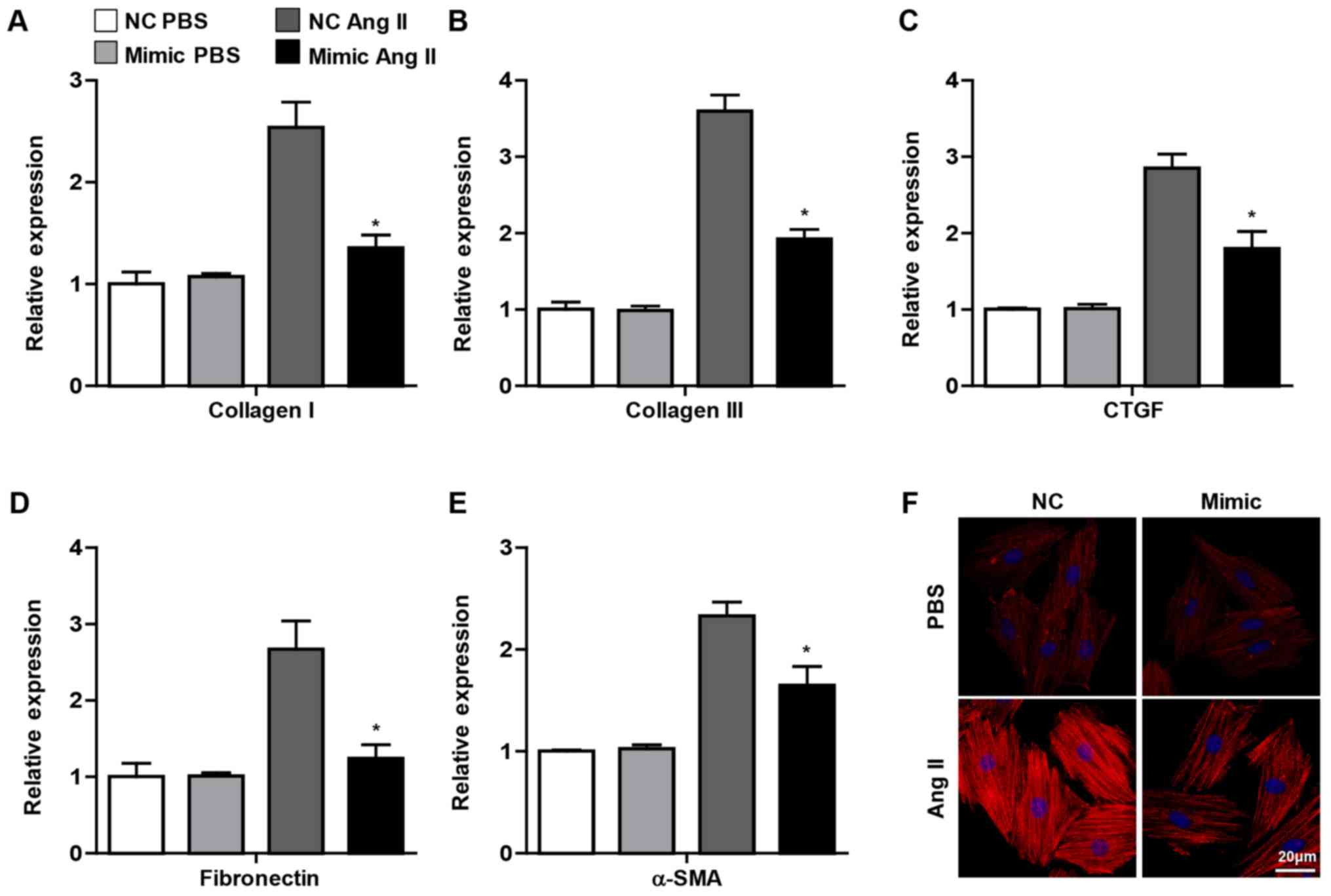
contraction and the ability to synthesize ECM (2). It was observed in the present study
that the pro-fibrotic genes, including collagen I, collagen III,
CTGF, fibronectin and α-SMA, were decreased in CFs transfected with
miR-144 mimic upon Ang II administration, whereas no significant
difference was observed in the PBS group (Fig. 3A-E). Furthermore, the
immunofluorescence staining demonstrated that overexpression of
miR-144 attenuated the α-SMA expression in CFs treated with Ang II
(Fig. 3F). Overall, these results
demonstrated that miR-144 ameliorated the activation of CFs.
miR-144 knockdown accelerates
myofibroblast differentiation into CFs, as well as the
proliferation and migration of CFs
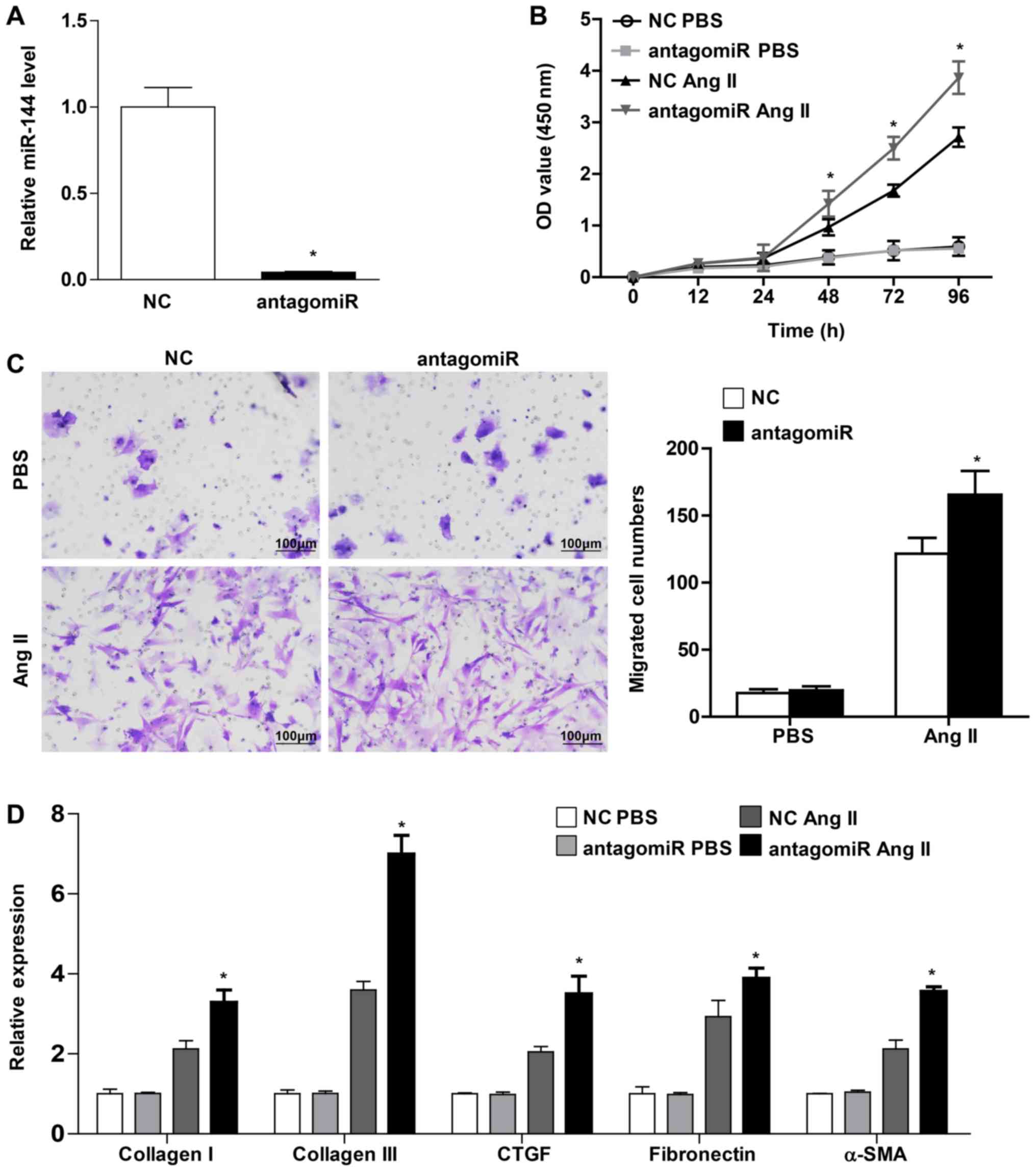
The present study transfected the CFs with miR-144
antagomiR in order to confirm the protective role of miR-144 on
cardiac fibroblast activation. miR-144 expression exhibited a
significant decrease, by 25-fold (Fig.
4A). The CFs exhibited a markedly enhanced proliferation and
migration ability in those that were infected with miR-144
antagomiR upon Ang II stimulation (Fig.
4B and C). Furthermore, the present study demonstrated that the
pro-fibrotic genes, including collagen I, collagen III, CTGF,
fibronectin and α-SMA, were increased in CFs transfected with
miR-144 antagomiR upon Ang II administration (Fig. 4D). Overall, these results revealed
that miR-144 knockdown accelerated the activation of CFs.
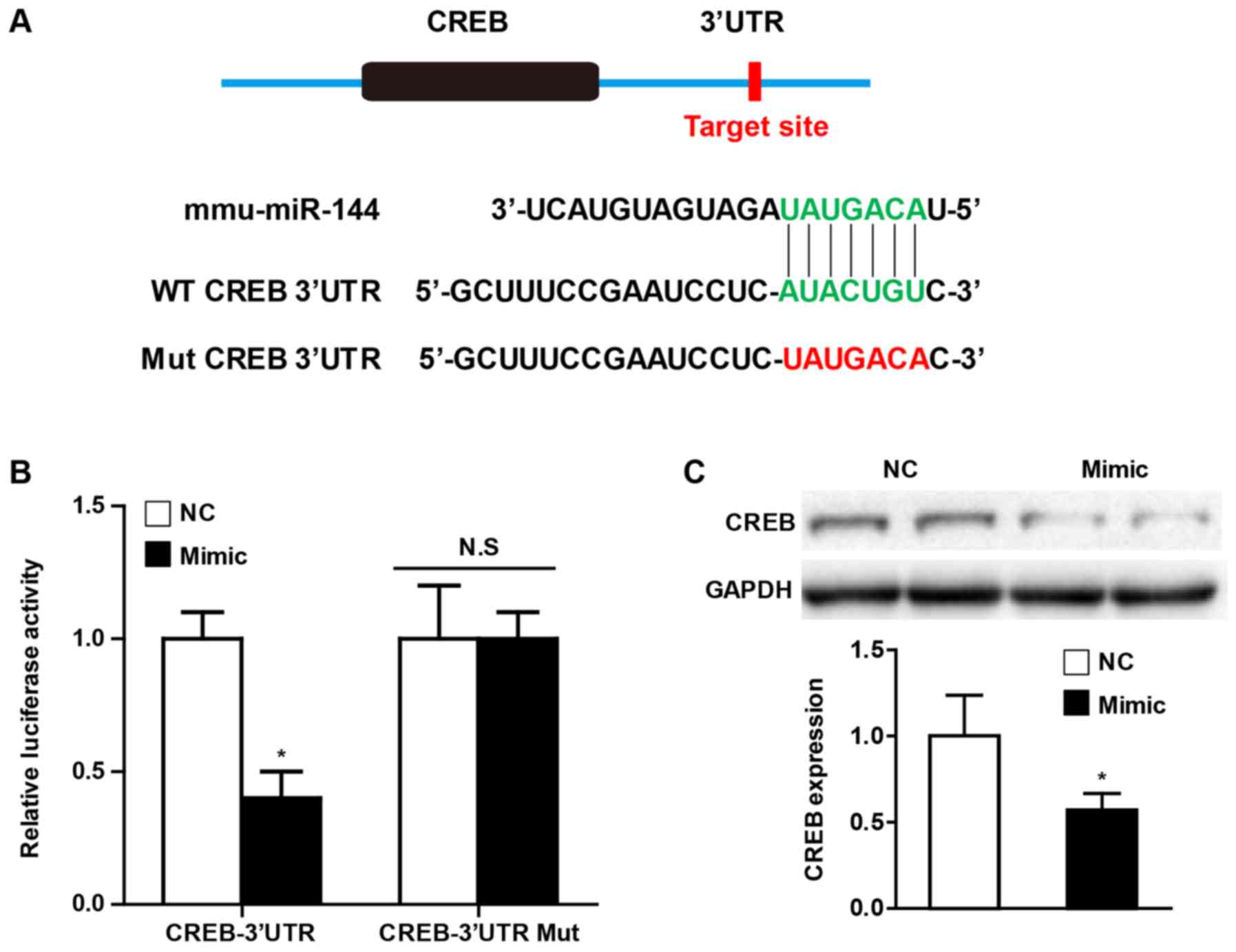
miR-144 directly targets CREB
Universally, the effects of miRNA on various
pathophysiological processes depend on the direct regulation of
their downstream targets. In order to further elucidate the
potential molecular mechanism by which miR-144 affects CF
activation, the present study performed a bioinformatics analysis
using the prediction algorithms TargetScan (version 7.0) in order
to identify the multiple target genes of miR-144. Of these
potential targets, CREB has been demonstrated to play an important
role in cardiac fibrosis, and the putative binding site for miR-144
in the 3'UTR is presented in Fig.
5A. Using luciferase reporter assays, the present study
observed that miR-144 mimic transfection decreased CREB luciferase
activity, and the effect was abolished when the predicted binding
sites within the CREB 3'UTR were mutated (Fig. 5B). Furthermore, the western blot
analysis demonstrated that overexpression of miR-144 significantly
attenuated the CREB protein level of CFs upon Ang II administration
(Fig. 5C).
Discussion
Accumulating evidence has demonstrated that cardiac
fibrosis is implicated in multiple cardiac diseases, including
hypertension, myocardial infarction and valvular heart disease
(2). Furthermore, the severity of
cardiac fibrosis is partially responsible for the progressive
morbidity, mortality, and healthcare expenditure caused by heart
failure (7,30). Cardiac fibrosis is characterized by
excessive extracellular matrix accumulation, which leads to the
destruction of normal heart tissue architecture, ventricular
remodeling and accelerated heart dysfunction (3). Despite improvements in the knowledge
surrounding cardiac fibrosis, there remains to be a lack of
effective treatment strategies. Aberrant miRNA expression has
emerged that links the pathophysiology of cardiac fibrosis with
heart failure, which may act as a potential novel therapeutic
target for the treatment of cardiac fibrosis (12,13).
miRNAs are endogenous, non-coding small RNA
molecules, which can be recruited to the RNA-induced silencing
complex and affect the target genes via diverse mechanisms
(8). Recently, the importance of
miRNAs in various cardiac disease has been well documented,
particularly the role of miRNAs in cardiac fibrosis (13). miR-26a has an effect on cardiac
fibrosis mediated by the NF-κB signaling pathway (31). miR-133a regulates collagen 1A and
connective tissue growth factor in myocardial fibrosis of cardiac
hypertension induced by Ang II stimulation (32). Let-7i negatively regulates cardiac
inflammation and fibrosis (33).
miR-122 suppresses fibrogenesis by targeting TGFβR1 in cardiac
fibroblasts. miR-21 promotes cardiac fibrosis following myocardial
infarction via targeting smad7(34).
miR-155 accelerates cardiac fibrosis in the process of Ang
II-induced cardiac remodeling (35).
miR-150 inhibits the activation of CFs by regulating c-Myb
(36). The previous miRNA arrays
demonstrated that miR-144 is associated with hypertrophic cardiac
tissues and cardiac diseases. However, the role and underlying
molecular mechanism of miRNAs in cardiac fibrosis remains
unclear.
miR-144 has been demonstrated to be involved in
multiple types of human malignancy and is downregulated in colon
cancer, lung cancer, prostate cancer and hepatocellular carcinoma,
as revealed by a comprehensive meta-analysis of miRNA expression
microarrays. Increasing evidence has verified that miR-144 acts as
a novel suppressor of the development of various tumors via
inhibition of the proliferation and migration of the cancer cells
(18-24).
Recently, it has been revealed that miR-144 plays an important role
in cardiac diseases. Specifically, miR-144 attenuates cardiac
ischemia/reperfusion injury by targeting FOXO1(37). miR-144 deficiency interrupts ECM
remodeling and decreases left ventricular remodeling following
myocardial infarction leading to worsened cardiac function
(26,27). Notably, bleomycin-induced pulmonary
fibrosis has been demonstrated to be associated with miR-144
expression (38), while miR-144
induces hepatic stellate cell activation in the human fibrotic
liver by targeting TGF-β1(39) and
regulates relaxin/insulin-like family peptide receptor 1 expression
in lung fibroblasts (40). Overall,
these results indicate that miR-144 may play an important role in
the fibrotic progression underlying various diseases. Previous
studies have demonstrated that cardiac fibrosis plays a critical
role in the regulation of heart function, ventricular remodeling
and the development of heart failure following cardiomyocyte
hypertrophy and myocardial infarction (1). In the present study, it was observed
that miR-144 expression was dramatically decreased in heart tissue
subjected to TAC surgery, and a gradual decrease in CFs was
observed as the concentration of Ang II increased. Furthermore, the
critical event in cardiac fibrosis is the transformation of CFs to
myofibroblasts, which is characterized by irreversible acquisition
of expression of α-SMA (6). The
present study clarified that miR-144 overexpression significantly
inhibited the proliferation and migration of CFs upon Ang II
administration. The multiple ECM proteins expressed in CFs were
attenuated by miR-144 which included minimal α-SMA positive
expression. By contrast, such effects could be reversed by
downregulation of miR-144. Therefore, the present study indicated
that miR-144 may play an important role in pathologies of cardiac
fibrosis.
miRNAs play important roles in the regulation of
cell development, differentiation, proliferation and apoptosis
through acting as negative regulators of target gene expression by
promoting the degradation or inhibiting the translation of
downstream mRNAs (10). In
silico target prediction analyses were performed in the present
study in order to investigate the underlying molecular mechanism by
which miR-144 regulated CF activation. Notably, CREB demonstrated a
conserved putative binding site for miR-144 in the 3'UTR in
different species, which is well recognized as being involved in
the multiple crucial signaling pathways implicated in cardiac
fibrosis and cardiac hypertrophy (41). Furthermore, several previous studies
have suggested that the activation of CREB was essential for
cardiomyocyte hypertrophy subjected to TAC surgery and Ang II
administration (42). Meanwhile,
during ventricular remodeling induced by pressure overload, cardiac
fibrosis was the hallmark pathological manifestation characterize
by excessive ECM deposition. In fact, the luciferase-reporter
assays demonstrated that the CREB luciferase activity was largely
attenuated by miR-144 mimic, and that this process was blocked when
the predicted binding sites within the CREB 3'UTR were mutated.
Notably, it was observed that CREB demonstrated a remarkable
decrease in expression of CFs upon Ang II administration. Overall,
it can be concluded that the detrimental role of miR-144 in
pathological CF activation is partially dependent on the negative
activation of CREB.
In conclusion, we first demonstrated that miR-144
showed a downregulation in heart tissue subjected to TAC and CFs
upon Ang II stimulation. The in vitro experiment further
clarified the miR-144 was largely implicated in the process of
cardiac fibrosis, which characterized by the decreased
proliferation and migration of CFs transfected with miR-144 induced
by Ang II treatment, as well as the attenuated myofibroblasts
transformation. Mechanistically, miR-144 directly targeted and
downregulated the CREB expression in CFs with Ang II stimulation.
Our current study suggests that miR-144-CREB axis could be an
important novel target for the treatment of cardiac fibrosis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FHL and QL designed the study and wrote the
manuscript. ZYL ML and JL performed the animal experiments. JL and
LLX performed the statistical analysis. ZNW and WWW performed the
RT-PCR and western blot analysis. YL and ZJS performed the cell
experiments.
Ethical approval and consent to
participate
All the procedures involving animal experiments were
performed according to the Guide for the Care and Use of Laboratory
Animals published by the U.S. National Institutes of Health and
were approved by the Animal Care and Use Committee of the Dezhou
People's Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Porter KE and Turner NA: Cardiac
fibroblasts: At the heart of myocardial remodeling. Pharmacol Ther.
123:255–278. 2009.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Fan D, Takawale A, Lee J and Kassiri Z:
Cardiac fibroblasts, fibrosis and extracellular matrix remodeling
in heart disease. Fibrogenesis Tissue Repair. 5(15)2012.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Leask A: Getting to the heart of the
matter: New insights into cardiac fibrosis. Circ Res.
116:1269–1276. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Leask A: TGFbeta, cardiac fibroblasts, and
the fibrotic response. Cardiovasc Res. 74:207–212. 2007.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Takeda N, Manabe I, Uchino Y, Eguchi K,
Matsumoto S, Nishimura S, Shindo T, Sano M, Otsu K, Snider P, et
al: Cardiac fibroblasts are essential for the adaptive response of
the murine heart to pressure overload. J Clin Invest. 120:254–265.
2010.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Weber KT, Sun Y, Bhattacharya SK, Ahokas
RA and Gerling IC: Myofibroblast-mediated mechanisms of
pathological remodelling of the heart. Nat Rev Cardiol. 10:15–26.
2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Thannickal VJ, Zhou Y, Gaggar A and Duncan
SR: Fibrosis: Ultimate and proximate causes. J Clin Invest.
124:4673–4677. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Trujillo RD, Yue SB, Tang Y, O'Gorman WE
and Chen CZ: The potential functions of primary microRNAs in target
recognition and repression. EMBO J. 29:3272–3285. 2010.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Shukla GC, Singh J and Barik S: MicroRNAs:
Processing, maturation, target recognition and regulatory
functions. Mol Cell Pharmacol. 3:83–92. 2011.PubMed/NCBI
|
|
11
|
Lima J Jr, Batty JA, Sinclair H and
Kunadian V: MicroRNAs in ischemic heart disease: From
pathophysiology to potential clinical applications. Cardiol Rev.
25:117–125. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Quiat D and Olson EN: MicroRNAs in
cardiovascular disease: From pathogenesis to prevention and
treatment. J Clin Invest. 123:11–18. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Thum T and Lorenzen JM: Cardiac fibrosis
revisited by microRNA therapeutics. Circulation. 126:800–802.
2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Schellings MW, Vanhoutte D, van Almen GC,
Swinnen M, Leenders JJ, Kubben N, van Leeuwen RE, Hofstra L,
Heymans S and Pinto YM: Syndecan-1 amplifies angiotensin II-induced
cardiac fibrosis. Hypertension. 55:249–256. 2010.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhang Y, Huang XR, Wei LH, Chung AC, Yu CM
and Lan HY: miR-29b as a therapeutic agent for angiotensin
II-induced cardiac fibrosis by targeting TGF-β/Smad3 signaling. Mol
Ther. 22:974–985. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Schneider MD: Serial killer: Angiotensin
drives cardiac hypertrophy via TGF-beta1. J Clin Invest.
109:715–716. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
17
|
Iwata M, Cowling RT, Yeo SJ and Greenberg
B: Targeting the ACE2-Ang-(1-7) pathway in cardiac fibroblasts to
treat cardiac remodeling and heart failure. J Mol Cell Cardiol.
51:542–547. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Iwaya T, Yokobori T, Nishida N, Kogo R,
Sudo T, Tanaka F, Shibata K, Sawada G, Takahashi Y, Ishibashi M, et
al: Downregulation of miR-144 is associated with colorectal cancer
progression via activation of mTOR signaling pathway.
Carcinogenesis. 33:2391–2397. 2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Guo Y, Ying L, Tian Y, Yang P, Zhu Y, Wang
Z, Qiu F and Lin J: miR-144 downregulation increases bladder cancer
cell proliferation by targeting EZH2 and regulating Wnt signaling.
FEBS J. 280:4531–4538. 2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zha W, Cao L, Shen Y and Huang M: Roles of
Mir-144-ZFX pathway in growth regulation of non-small-cell lung
cancer. PLoS One. 8(e74175)2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhao M, Huang J, Gui K, Xiong M, Cai G, Xu
J, Wang K, Liu D, Zhang X and Yin W: The downregulation of miR-144
is associated with the growth and invasion of osteosarcoma cells
through the regulation of TAGLN expression. Int J Mol Med.
34:1565–1572. 2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Cui SQ and Wang H: MicroRNA-144 inhibits
the proliferation, apoptosis, invasion, and migration of
osteosarcoma cell line F5M2. Tumour Biol. 36:6949–6958.
2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Bao H, Li X, Li H, Xing H, Xu B, Zhang X
and Liu Z: MicroRNA-144 inhibits hepatocellular carcinoma cell
proliferation, invasion and migration by targeting ZFX. J Biosci.
42:103–111. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Gu J, Liu X, Li J and He Y: MicroRNA-144
inhibits cell proliferation, migration and invasion in human
hepatocellular carcinoma by targeting CCNB1. Cancer Cell Int.
19(15)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Siddiqui MR, Akhtar S, Shahid M, Tauseef
M, McDonough K and Shanley TP: miR-144 mediated inhibition of ROCK1
protects against LPS induced lung endothelial hyperpermeability. Am
J Respir Cell Mol Biol. 61:257–265. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
He Q, Wang F, Honda T, James J, Li J and
Redington A: Loss of miR-144 signaling interrupts extracellular
matrix remodeling after myocardial infarction leading to worsened
cardiac function. Sci Rep. 8(16886)2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Li J, Cai SX, He Q, Zhang H, Friedberg D,
Wang F and Redington AN: Intravenous miR-144 reduces left
ventricular remodeling after myocardial infarction. Basic Res
Cardiol. 113(36)2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Jiang DS, Bian ZY, Zhang Y, Zhang SM, Liu
Y, Zhang R, Chen Y, Yang Q, Zhang XD, Fan GC and Li H: Role of
interferon regulatory factor 4 in the regulation of pathological
cardiac hypertrophy. Hypertension. 61:1193–1202. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Jiang DS, Wei X, Zhang XF, Liu Y, Zhang Y,
Chen K, Gao L, Zhou H, Zhu XH, Liu PP, et al: IRF8 suppresses
pathological cardiac remodelling by inhibiting calcineurin
signalling. Nat Commun. 5(3303)2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Brown RD, Ambler SK, Mitchell MD and Long
CS: The cardiac fibroblast: Therapeutic target in myocardial
remodeling and failure. Annu Rev Pharmacol Toxicol. 45:657–687.
2005.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wei C, Kim IK, Kumar S, Jayasinghe S, Hong
N, Castoldi G, Catalucci D, Jones WK and Gupta S: NF-κB mediated
miR-26a regulation in cardiac fibrosis. J Cell Physiol.
228:1433–1442. 2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Matkovich SJ, Wang W, Tu Y, Eschenbacher
WH, Dorn LE, Condorelli G, Diwan A, Nerbonne JM and Dorn GW II:
MicroRNA-133a protects against myocardial fibrosis and modulates
electrical repolarization without affecting hypertrophy in
pressure-overloaded adult hearts. Circ Res. 106:166–175.
2010.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang X, Wang HX, Li YL, Zhang CC, Zhou CY,
Wang L, Xia YL, Du J and Li HH: MicroRNA Let-7i negatively
regulates cardiac inflammation and fibrosis. Hypertension.
66:776–785. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Yuan J, Chen H, Ge D, Xu Y, Xu H, Yang Y,
Gu M, Zhou Y, Zhu J, Ge T, et al: Mir-21 promotes cardiac fibrosis
after myocardial infarction via targeting Smad7. Cell Physiol
Biochem. 42:2207–2219. 2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wei Y, Yan X, Yan L, Hu F, Ma W, Wang Y,
Lu S, Zeng Q and Wang Z: Inhibition of microRNA155 ameliorates
cardiac fibrosis in the process of angiotensin II induced cardiac
remodeling. Mol Med Rep. 16:7287–7296. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Deng P, Chen L, Liu Z, Ye P, Wang S, Wu J,
Yao Y, Sun Y, Huang X, Ren L, et al: MicroRNA-150 inhibits the
activation of cardiac fibroblasts by regulating c-Myb. Cell Physiol
Biochem. 38:2103–2122. 2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
E L, Jiang H and Lu Z: MicroRNA-144
attenuates cardiac ischemia/reperfusion injury by targeting FOXO1.
Exp Ther Med. 17:2152–2160. 2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Xie T, Liang J, Guo R, Liu N, Noble PW and
Jiang D: Comprehensive microRNA analysis in bleomycin-induced
pulmonary fibrosis identifies multiple sites of molecular
regulation. Physiol Genomics. 43:479–487. 2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Liu Z, Yi J, Ye R, Liu J, Duan Q, Xiao J
and Liu F: miR-144 regulates transforming growth factor-β1 iduced
hepatic stellate cell activation in human fibrotic liver. Int J
Clin Exp Pathol. 8:3994–4000. 2015.PubMed/NCBI
|
|
40
|
Bahudhanapati H, Tan J, Dutta JA, Strock
SB, Sembrat J, Alvarez D, Rojas M, Jäger B, Prasse A, Zhang Y and
Kass DJ: MicroRNA-144-3p targets relaxin/insulin-like family
peptide receptor 1 (RXFP1) expression in lung fibroblasts from
patients with idiopathic pulmonary fibrosis. J Biol Chem.
294:5008–5022. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Chan EC, Dusting GJ, Guo N, Peshavariya
HM, Taylor CJ, Dilley R, Narumiya S and Jiang F: Prostacyclin
receptor suppresses cardiac fibrosis: Role of CREB phosphorylation.
J Mol Cell Cardiol. 49:176–185. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
El Jamali A, Freund C, Rechner C,
Scheidereit C, Dietz R and Bergmann MW: Reoxygenation after severe
hypoxia induces cardiomyocyte hypertrophy in vitro: Activation of
CREB downstream of GSK3beta. FASEB J. 18:1096–1098. 2004.PubMed/NCBI View Article : Google Scholar
|