Introduction
Coronary artery disease (CAD) is defined as an
inflammatory disease developing mainly at branch points and bends
of arteries, in which arterial blood flow continuously causes
slight damage to the endothelial monolayer lining of the vessel
wall. This process seems to be the key to the progression of
atherosclerosis, because the maladaptation of endothelial cells
(ECs) to this physiological disorder increases the susceptibility
of branch points to pro-inflammatory states. Therefore, maladapted
ECs make it easier for arterial branch points to circulate in the
subendothelial space, lead to the inflow of oxidized low-density
lipoproteins (ox-LDLs), and stimulate monocyte recruitment by
releasing pro-inflammatory molecules (i.e., chemokines and adhesion
molecules) (1). Subendothelial
monocytes differentiate into macrophages, which locally proliferate
and absorb ox-LDLs. Then the deposition of lipid into plaques leads
to deterioration, thus triggering macrophages to become foam cells
due to insufficient lipoprotein uptake. The progression of advanced
lesions is featured with the accumulation of lipoproteins,
apoptosis and necrosis of macrophage-derived foam cells, the
formation of cholesterol crystals due to increased defective
phagocytes, and the formation of cap smooth muscle cells (SMCs)
(2). Hence, increased lesions in the
advanced stage lead to severe reduction of arterial lumen and blood
flow, decline of oxygen supply and rupture or dissolution of
plaques, thus causing thrombosis. Analysis of the early development
stage of atherosclerosis reveals that inflammation exerts a crucial
effect in the development of atherosclerosis. Surprisingly, the
emphasis of many current therapies applied to treat atherosclerosis
and thrombosis has been put on coping with the onset of acute
inflammatory states. It is noteworthy that statins can not only
reduce the cholesterol level, but also play an anti-inflammatory
role (3). This study, therefore,
aims to more specifically reduce inflammatory activities during
atherosclerosis using colchicine and interleukin (IL)-1β antibodies
or other modes targeting IL-6 or IL-1 receptors (4).
A genome-wide association study (GWAS) and a
single-cell sequencing database have revealed the existence of
complex regulatory networks triggering cardiovascular diseases
(5). In these networks, the
heterogeneity and correlation of vascular cell population as well
as their contributions to different stages of atherosclerosis are
especially prominent. In addition, the GWAS has shown
disease-related genetic variations in the non-protein coding
sequence space, which are actively transcribed into non-coding
ribonucleic acids (ncRNAs), i.e., micro RNAs (miRNAs) and long
ncRNAs (lncRNAs) (6). These new-type
ncRNAs exhibit differential expression in diseased tissues and
function as epigenetic regulators for gene expression. Among them,
the identification of molecules related to the interaction between
phenotypes and cells and epigenetic regulation are new and
selective therapeutic targets for the treatment of
atherosclerosis.
Circular antisense non-coding RNA in the INK4 locus
(circANRIL) is transcribed at the atherosclerotic CVD gene site on
chromosome 9p21 (Chr9p21), which may have anti-atherosclerosis
function. The high level of circANRIL expression in human vascular
tissues is related to lower CAD severity. In terms of mechanism,
circANRIL has been proven to bind to Pescadillo homolog 1 (PES1),
an indispensable preassembly factor for 60S-ribosomes, thus
possibly resulting in the damage of nucleic acid
exonuclease-mediated pre-ribosomal RNA (rRNA) processing and
ribosome biogenesis in VSMCs and macrophages (7). On the contrary, circANRIL induces p53
activation, leading to increased apoptosis, while the proliferation
of vascular SMCs (VSMCs) and macrophages is decreased. Therefore,
circANRIL might prevent atherosclerosis by inhibiting the excessive
proliferation of cells in atherosclerotic plaques.
This research explored the effects of circANRIL on
vascular endothelial injury, oxidative stress and inflammation in
rats with coronary heart disease, and investigated a new method of
treating CAD from the perspective of vascular biology.
Materials and methods
Materials
Sprague Dawley (SD) rats and feed were purchased
from HFK Bioscience, circANRIL overexpression plasmids and
silencers from JKJC Gene Technology Co., Ltd., rat IL-6 (cat. no.
k4143-100) and TNF-α (cat. no. k1052-100) enzyme-linked
immunosorbent assay (ELISA) kits from Jitai Yikai Biotechnology
Co., Ltd., rabbit anti-rat p38 mitogen-activated protein kinase
(p38MAPK, cat. no. ab170099) and phosphorylated (p)-p38MAPK
antibodies (cat. no. ab47363) were from Abcam, rabbit anti-rat
glyceraldehyde 3-phosphate dehydrogenase (GAPDH, cat. no.
10494-1-AP) from Proteintech, and factor VIII immunohistochemistry
kit was from ZSGB-Bio Co., Ltd.
Modeling of coronary heart
disease
A total of 40 healthy SD male rats aged 3-4 months,
with an average body weight of 250.24±62.27 g were included and
then, fed with normal diet and normal drinking water. At 7 days
after feeding, the rats were randomly divided into research group
(n=32) and control group (n=8). The rats in control group received
only basic feed and tap water with no special treatment, while
those in research group were fed with high-fat diet. At the
beginning of the experiment, vitamin D3 powder was injected into
the right lower limb of rats at a dose of 3x106 U/kg
every 30 days (8). The basic diet of
the rats accounted for 94.30%, and was supplemented with 2%
cholesterol, 3% lard, 0.5% sodium cholate and 0.2%
propylthiouracil. The experiment lasted three months. Then the rats
were sacrificed and aortic specimens were collected. The rats
received euthanasia via cervical dislocation (after being
anesthetized using peritoneal administration of pentobarbital
sodium at a dose of 40 mg/kg). If there was calcification of the
intima, the atherosclerosis model of the rat was successful. The
success rate of the model was 70%, and the death rate was 10%. The
cause of death may be related to the loss of appetite, poor
resistance and diarrhea. The study was approved by the Ethics
Committee of the People's Hospital of Zhangqiu Area (Jinan,
China).
Detection of blood calcium
(Ca2) and lipid
The rats were anesthetized using pentobarbital
sodium at a dose of 40 mg/kg intraperitoneally. Blood (2 ml) was
collected from the abdominal aorta of rats. After centrifugation at
4˚C, 10,500 x g for 5 min, plasma was separated, and 1 ml of blood
Ca2+ and 1 ml of lipid were measured. The expression of
total cholesterol (TC), triglyceride (TG), high-density lipoprotein
cholesterol (HDLC), low-density lipoprotein cholesterol (LDLC) and
blood Ca2+ was detected using a biochemical
analyzer.
Cell culture method and verification
of cell purity
In a sterile environment, the proximal coronary
artery of the rats was taken out and washed in phosphate buffer
containing double antibodies (1% penicillin and 1% streptomycin).
After centrifugation at 4˚C, 300 x g for 10 min, the precipitation
was collected and cultured in 20% fetal bovine serum (FBS),
Dulbecco's modified Eagle's medium-F12 (DEME-F12) (Gibco) in an
environment with moderate humidity and 5% CO2 at 37˚C.
It was observed that ECs crawled out from the edge of the tissue
mass and gradually extended outward within ~7 days. At this time,
with the tissue mass removed, the cells were washed 3 times with
phosphate-buffered saline (PBS) to remove the residual culture
medium. Then 0.25% trypsin was added to the culture dish for
digestion. When the cells contracted and became round (~10 min), 5
ml of 20% FBS DEME-F12 medium was added, and the cells were
inoculated into a 25 cm2 culture flask. Subsequently, 2
ml of cells were inoculated into a 6-well plate (with
pre-sterilized glass plates) at a density of 1.2x105
cells/ml. When the cells reached 70% fusion rate, the detection
started. ECs were flat and short fusiform or polygon, and they
contained only factor VIII. Through immunochemical staining as well
as the specific binding of antibodies and antigens, the nucleus
containing factor VIII was yellow, while that without factor VIII
was dark blue, which could be used to distinguish cells.
Cell transfection method
One day before transfection, the transfected cells
were seeded into a 12-well plate at a density of
2x105/cm2. Transfection was performed using
Lipofectamine™ 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
When the fusion rate reached ~70%, circANRIL transfection was
carried out according to the instructions of the transfection kit,
and corresponding negative control experiments were performed to
form blank group (untransfected cells), negative group (transfected
with blank vector), circANRIL group (transfected with circANRIL
overexpression plasmid, 20 µM), and circANRIL inhibitor group
(transfected with circANRIL silencer siRNA, 20 µM). After 18 h of
transfection, the fluorescent tag was added to detect the
transfection efficiency. Following incubation for 24 h, subsequent
experimental steps were conducted.
Lactate dehydrogenase (LDH)
detection
When cells were damaged, LDH was released into the
supernatant. Since LDH in the supernatant did not easily react with
other components, the amount of LDH in the supernatant could be
measured to reflect the number of dead and damaged cells. Hundred
milliliters of the supernatant was absorbed by each well of a
96-well plate. Then the supernatant was aspirated carefully to
prevent cells from aspiration. Thereafter, 100 ml of working
solution was added to each well, and the well was coated with
aluminum foil to avoid light. After reacting at room temperature
for ~30 min, 50 ml of termination solution was added to each well,
and the absorption amount at 490 nm was immediately measured using
a microplate reader. Finally, the cell damage rate was calculated
according to the following formula: Cell damage rate = sample
well/high control well x100%.
Determination of superoxide dismutase
(SOD) and malon- dialdehyde (MDA)
The supernatant in each group was collected, and the
content of SOD in rat serum was determined by xanthine oxidase
method. After that, free radicals produced by reactions of xanthine
and xanthine oxidase were detected. Next, the content of MDA in
serum was determined by thiobarbituric acid colorimetry according
to the instructions of a kit provided by Shanghai Jinghua
Instruments Co., Ltd.
Detection of IL-6 and TNF-α by
ELISA
Cell supernatant was collected, and protein
concentration was determined by bicinchoninic acid assay (BCA)
(Pierce; Thermo Fisher Scientific, Inc.). According to the
instructions of the ELISA kit, the micropores of rat IL-1β, IL-6
and TNF-α capture antibodies were pre-coated, and then samples,
standard substances and horseradish peroxidase (HRP)-labeled
antibodies were sequentially added and thoroughly washed. When the
substrate TMB was used for color development, TMB turned blue under
the catalysis of peroxidase and yellow under the action of acid.
Its color was positively correlated with the content of IL-6 and
TNF-α in the supernatant of rat ECs. The optical density was
measured at 450 nm using the microplate reader, and the
concentrations of IL-6 and TNF-α were calculated.
Detection of the protein expression of
p-p38MAPK and p38MAPK in the supernatant via western blotting
Cell supernatant was collected from each group.
Based on the instructions of the BCA kit, the protein concentration
in ECs was detected. Then, 5X sample buffer was added and boiled,
and proteins (10 µg) were separated by polyacrylamide gel
electrophoresis and transferred onto a polyvinylidene fluoride
membrane. Next, the proteins were sealed with 10% skim milk,
blocked with 2% phosphoenzyme inhibitor p-Smad3, and incubated with
anti-p-p38MAPK antibody (1:2,000) and p38MAPK antibody (1:1,000)
after washing, followed by incubation overnight at room temperature
of 4˚C. Then the membranes were incubated with goat horseradish
peroxidase-conjugated goat anti-rabbit IgG H&L secondary
antibody (1:1,000; cat. no. ab7090; Abcam) at 25˚C for 1 h.
Thereafter, the exposure was performed by Bio-Rad gel imaging
system, and Image Lab 6.0 software was adopted for processing. The
gray value was calculated, and the ratio of the gray value of the
target band to the corresponding internal reference GAPDH reflected
the relative expressions of p-p38MAPK and p38MAPK.
Statistical analysis
Data in each group were collected using Statistical
Product and Service Solutions (SPSS) 19.0 software (SPSS Inc.) and
analyzed as (mean ± SD). Differences between two groups were
analyzed by using the Mann Whitney U test. Comparison between
multiple groups was done using the Kruskal-Wallis test with Dunn's
post hoc test.
Results
Comparison of the levels of blood
Ca2 and lipid in rats
Research group had higher levels of TC, TG and LDLC
(P<0.05 or P<0.01), and lower levels of HDLC and
Ca2+ than control group (P<0.05) (Table I).
 | Table IComparison of the levels of blood
Ca2+ and lipid in rats (mean ± SD, mmol/l). |
Table I
Comparison of the levels of blood
Ca2+ and lipid in rats (mean ± SD, mmol/l).
| Group | n | TC | TG | HDLC | LDLC | Ca2+ |
|---|
| Control group | 8 | 5.12±0.75 | 4.52±0.86 | 2.63±1.17 | 2.03±0.93 | 3.67±0.14 |
| Research group | 32 |
16.73±1.72a |
1.19±0.04b |
2.01±0.94b |
7.83±1.39b |
2.42±0.07b |
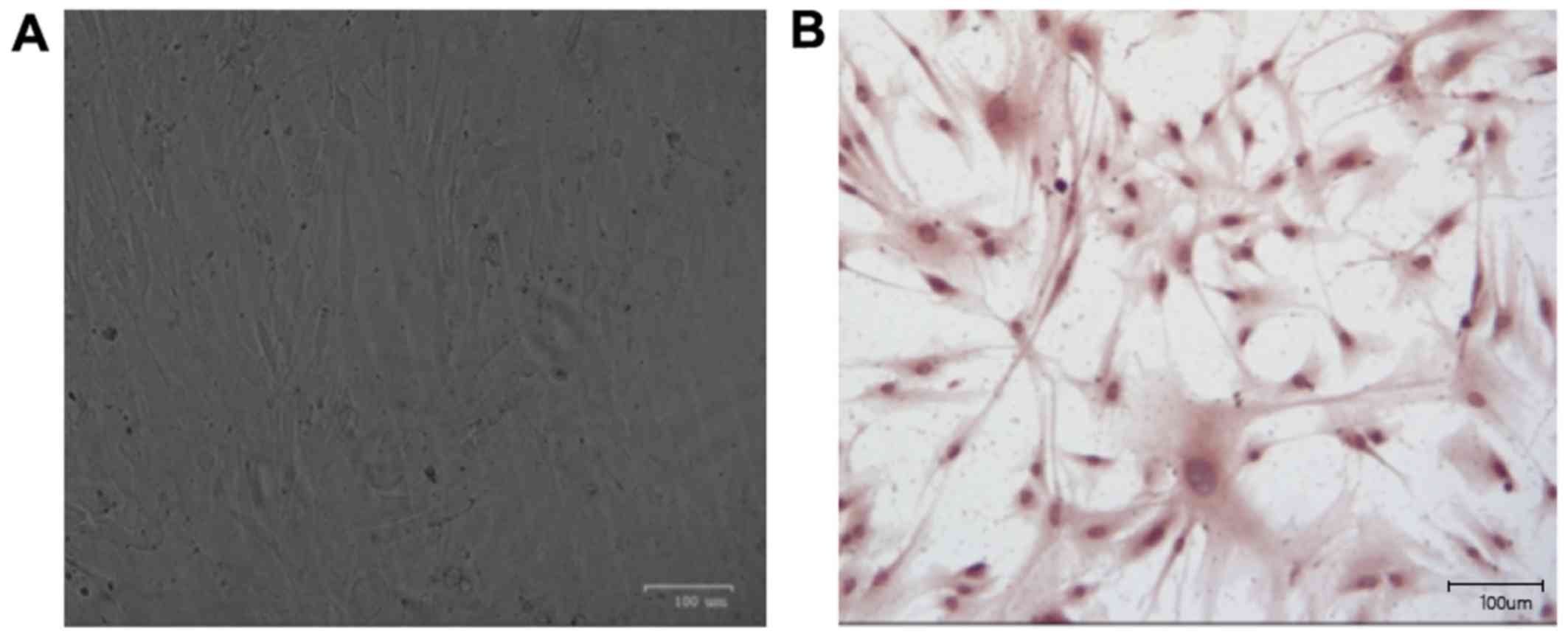
EC identification results
Within one week, cell adhesion was decreased and
growth became slow. After 7 days, a large number of cells grew out
of the clump in good growth conditions. Under an inverted
microscope, the cells adhered to the wall and grew in short spindle
or polygonal shape (Fig. 1A). Factor
VIII is released from ECs. Based on this principle, factor VIII was
detected by immunohistochemistry. As the basis for identifying ECs,
brown paving-like cells were ECs (Fig.
1B).
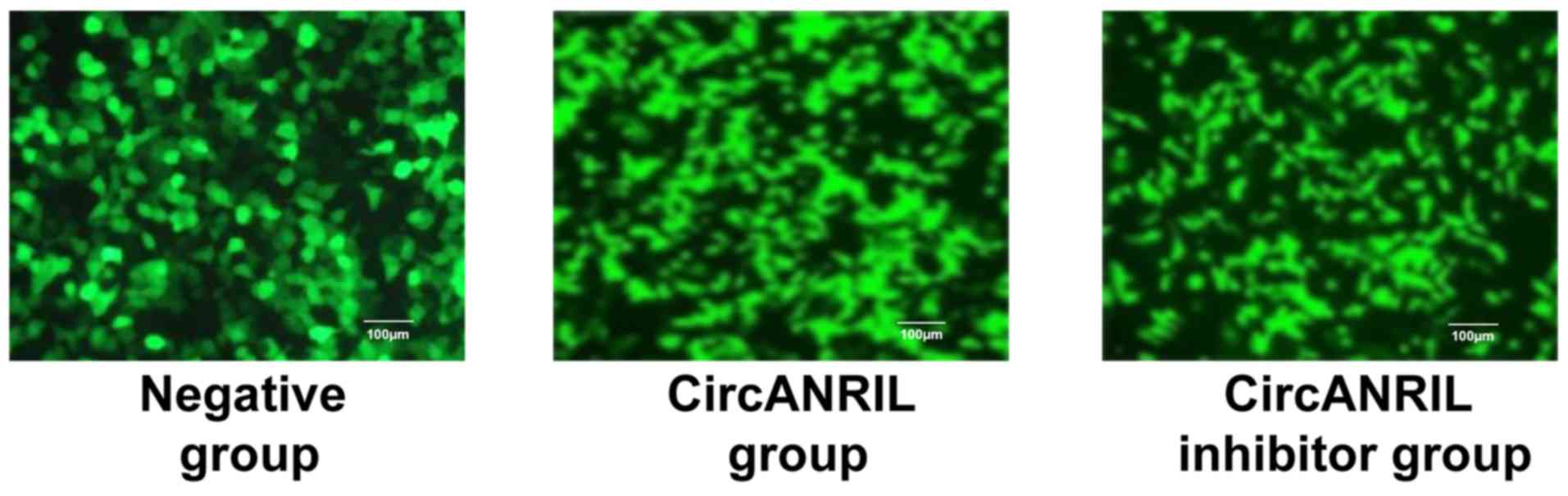
Cell transfection results
Negative control (empty vector), circANRIL
overexpression plasmid and cANRIL silencer were transfected into
ECs, separately. The transfection efficiency was >70% (Fig. 2), which met the requirements for the
next experiment.
Determination of EC damage rate in
each group
Compared with that in blank group, the damage rate
in negative group did not change significantly (P>0.05).
Besides, in comparison with that in the negative group, the cell
damage rate in the circANRIL group was decreased (P<0.01), but
it did not change in circANRIL inhibitor group (P>0.05)
(Table II).
| Table IIComparison of EC damage rate in each
group (mean ± SD, %). |
Table II
Comparison of EC damage rate in each
group (mean ± SD, %).
| Group | n | Damage rate |
|---|
| Blank group | 6 | 38.67±4.78 |
| Negative group | 6 |
40.62±4.65a |
| circANRIL group | 6 |
21.76±3.58b |
| circANRIL inhibitor
group | 6 |
39.72±4.97c |
Expression levels of SOD and MDA in
ECs in each group
Compared with those in blank group, there were no
significant changes in SOD and MDA releases in negative group
(P>0.05). Compared with those in negative group, SOD and MDA
releases in circANRIL group were remarkably decreased (P<0.01,
P<0.05). In addition, no obvious changes in SOD and MDA releases
were found between negative group and circANRIL inhibitor group
(P>0.05) (Table III).
| Table IIISOD and MDA in ECs in each group (mean
± SD, n=8). |
Table III
SOD and MDA in ECs in each group (mean
± SD, n=8).
| Group | SOD (U/ml) | MDA (mmol/l) |
|---|
| Blank group | 50.67±7.78 | 47.25±9.27 |
| Negative group |
52.67±8.03a |
46.85±8.75a |
| circANRIL group |
89.52±12.38b |
21.76±3.58c |
| circANRIL inhibitor
group |
53.62±11.17d |
45.82±8.67d |
Expression levels of IL-6 and TNF-α in
ECs in each group
Compared with those in blank group, there were no
evident changes in IL-6 and TNF-α releases in negative group
(P>0.05). Compared with those in negative group, IL-6 and TNF-α
releases in circANRIL group were reduced (P<0.05), but they did
not change in circANRIL inhibitor group (P>0.05) (Table IV).
| Table IVExpression levels of IL-6 and TNF-α
in ECs in each group (mean ± SD, n=8). |
Table IV
Expression levels of IL-6 and TNF-α
in ECs in each group (mean ± SD, n=8).
| Group | IL-6 (pg/ml) | TNF-α (pg/ml) |
|---|
| Blank group | 100.87±10.78 | 47.83±5.62 |
| Negative group |
99.87±10.36a |
44.72±5.03a |
| circANRIL
group |
67.92±6.84b |
26.61±3.83b |
| circANRIL inhibitor
group |
107.56±11.62c |
45.62±5.93c |
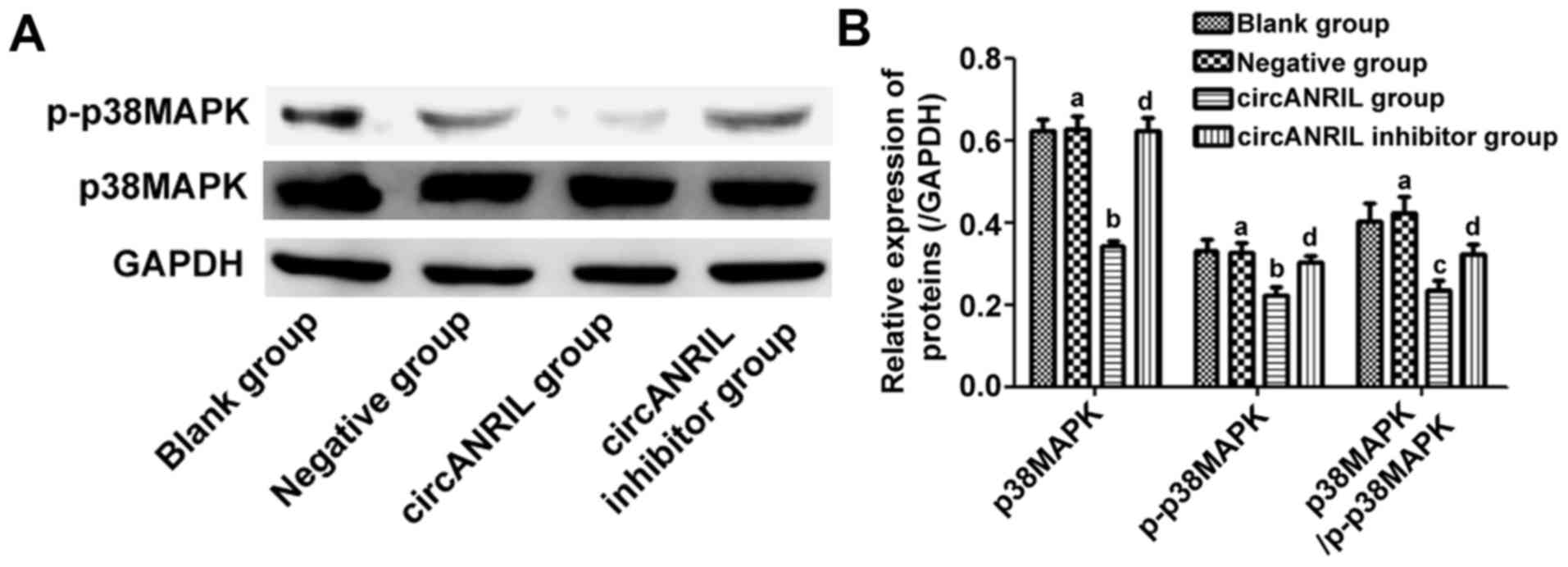
Expression of p-p38MAPK and p38MAPK
proteins in ECs
p-p38MAPK, p38MAPK and p38MAPK/p-p38MAP did not
markedly change in blank group compared with those in negative
group (P>0.05). In comparison with those in negative group, the
expression of p-p38MAPK and p38MAPK were decreased (P<0.05), and
p38MAPK/p-p38MAP distinctly declined in circANRIL group
(P<0.01), but they showed no changes in circANRIL inhibitor
group (P>0.05) (Fig. 3).
Discussion
The progressive lesions of human coronary
atherosclerosis can be reversed by calcium antagonists, while the
traditional high-fat diet fed animals have only cholesterol
deposition, but no obvious calcium deposition. It has been reported
that high dose of vitamin D3 can induce arterial wall
calcification, which can be reversed by calcium antagonists.
Therefore, rat model of coronary atherosclerosis induced by
high-fat diet combined with vitamin D3 conforms to human coronary
atherosclerosis better. In this study, high-fat diet combined with
vitamin D3 were used for modeling rat coronary atherosclerosis.
In the study of cardiovascular diseases, it is
urgent to find new methods for the diagnosis, treatment and
prognosis of coronary heart disease. At present, coronary heart
disease is still the major cause of death worldwide. People are
very interested in the discovery of molecular biomarkers, which can
supplement the traditional cardiovascular risk score in clinical
decision-making and help to stratify patients for personalized
treatment. With the development of next-generation sequencing and
bioinformatics techniques, the interest in circRNAs has gradually
emerged in recent years. It is noteworthy that molecules considered
as ‘junk RNAs’ for 20 years are now one of the most attractive
molecules (9). CircRNAs exert
crucial effects in physiological and pathological processes. They
are present not only in different tissues but also in body fluids,
such as blood, saliva and semen, confirming its potential role as
clinical biomarkers (10).
CircRNAs are produced by exon or intron sequences
and conserved among species, and they exhibit specific expression
in tissues. ANRIL is expressed in ECs, VSMCs, inflammatory cells
and tissues affected by atherosclerosis. It has been proved that
silencing ANRIL in human aortic VSMCs by siRNAs selectively
targeting exon1 or exon19 can differentially regulate apoptosis,
proliferation, inflammation and ECM remodeling, i.e. BCL2A1,
baculoviral IAP repeat-containing protein 3, cadherin 5 and heparin
binding EGF-like growth factor, show isotype-specific regulatory
characteristics (11). Lo Sardo
et al (12) recently induced
pluripotent stem cell-derived VSMCs from CAD risk and non-risk
individuals using TALEN technique, and deleted regions
corresponding to 60 kb risk haplotypes (depleted coding genes).
Transcription profile analysis demonstrated that individual CAD
risk VSMC displays altered gene expression patterns, similar to
previous findings in CAD risk individuals. Besides, the individual
VSMC also shows abnormal adhesion, contraction and proliferation.
Deleting risk haplotypes can save the normal phenotype of VSMCs. On
the contrary, risk phenotypes induced by lncRNA-ANRIL in non-risk
VSMCs are forced to be expressed, have no degradation mediated by
nucleic acid exonucleases and are more stable than most linear RNAs
(13) as circRNAs form a covalently
closed continuous loop, which is a great advantage in clinic.
CircRNAs modulate gene expression through various mechanisms. In
fact, circRNAs can act as miRNA sponges and play a competitive role
in binding miRNAs through post-transcriptional regulation (14). Additionally, circRNAs can also
regulate transcription by interacting with nuclear small RNAs or
RNA polymerase II (15). Moreover,
circRNAs are able to regulate RNA splicing by binding to
transcription factors (16). The
value of circRNAs in the diagnosis and prediction of heart diseases
needs exploration. Emerging clinical and experimental studies
verify that circRNA may be potential key regulatory factors for the
onset and development of CAD. Burd et al (17) found that the circANRIL expression is
related to INK4/ARF transcription and atherosclerotic disease risk.
It is interesting that the genetic variation of Chr9p21 is the most
widely recognized CAD genetic risk, which can regulate ANRIL
splicing and circANRIL production (18). circANRIL can protect atherosclerosis
by controlling the maturation of ribosomal RNAs and regulating the
formation pathway of atherosclerosis (19). Specifically, circANRIL binds to PES1,
which is an essential 60S-ribosome preassembly factor, thus
impairing nucleic acid exonuclease-mediated pro-rRNA processing and
ribosome biogenesis in VSMCs and macrophages. Therefore, circANRIL
induces nucleolar stress and p53 activation and causes induction of
apoptosis and inhibition of proliferation, which is the key to
atherosclerosis (20). Besides,
reducing the expression of circANRIL can prevent coronary
atherosclerosis by decreasing apoptosis of vascular ECs and the
expression of inflammatory factors (20). In the study of Song et al
(21) the expression levels of TC,
TG, LDL, IL-1, IL-6, matrix metalloproteinase 9 and C-reactive
protein in circANRIL group are low, and Bax, caspase-3 and the
apoptosis rate of ECs are reduced, while the expression levels of
HDL and Bcl-2 mRNAs and proteins are increased. Conversely, the
changes in the expression levels in circANRIL overexpression group
are the opposite to those in circANRIL low-expression group. The
results confirm the protective effect of circANRIL in
atherosclerosis, but when the dose exceeds a certain threshold, the
protective effect will resume.
Holdt et al (7) demonstrated that circANRIL is involved
in the maturation of VSMC and macrophage rRNAs. Specifically,
pre-rRNA treatment and ribosome biosynthesis are impaired,
resulting in nucleolar pressure, activation of p53, and
subsequently increased cell apoptosis and reduced proliferation
rate by binding circANRIL to PES1. Therefore, the protective effect
of atherosclerosis on the formation of atherosclerotic plaques is
proposed, which involves inhibiting cell proliferation in the early
stage of atherosclerotic plaque development. This indicates that
the genotype of Chr9p21 is essential for modulating the balance
between linearity and circANRIL levels in VSMCs and macrophages.
Therefore, changes in the percentage of ANRIL linear isotypes will
be beneficial to atherosclerosis. In fact, the expression of
exogenous circANRIL has been proven to be beneficial in the rat
model of coronary atherosclerosis (21).
In conclusion, inhibition of circANRIL expression in
coronary heart disease can reduce vascular endothelial injury,
oxidative stress and inflammatory responses, thus providing new
ideas and molecular biological methods for the diagnosis and
treatment of CAD.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
PS and JW designed the study and performed the
experiments. PS and HJ established the animal models. HZ was also
involved in the conception of the study. JY and RG analyzed the
data. PS and JW wrote the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the People's Hospital of Zhangqiu Area (Jinan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Li GM, Zhang CL, Rui RP, Sun B and Guo W:
Bioinformatics analysis of common differential genes of coronary
artery disease and ischemic cardiomyopathy. Eur Rev Med Pharmacol
Sci. 22:3553–3569. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Natarelli L and Weber C: Next-generation
therapeutic concepts for atherosclerosis: Focus on cell specificity
and noncoding RNAs. Thromb Haemost. 119:1199–1201. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Martini E, Stirparo GG and Kallikourdis M:
Immunotherapy for cardiovascular disease. J Leukoc Biol.
103:493–500. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Busygina K, Denzinger V, Bernlochner I,
Weber C, Lorenz R and Siess W: Btk Inhibitors as first oral
atherothrombosis-selective antiplatelet drugs? Thromb Haemost.
119:1212–1221. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ivanov A, Memczak S, Wyler E, Torti F,
Porath HT, Orejuela MR, Piechotta M, Levanon EY, Landthaler M,
Dieterich C, et al: Analysis of intron sequences reveals hallmarks
of circular RNA biogenesis in animals. Cell Rep. 10:170–177.
2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Holdt LM, Stahringer A, Sass K, Pichler G,
Kulak NA, Wilfert W, Kohlmaier A, Herbst A, Northoff BH, Nicolaou
A, et al: Circular non-coding RNA ANRIL modulates ribosomal RNA
maturation and atherosclerosis in humans. Nat Commun.
7(12429)2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Gou SH, Liu BJ, Han XF, Wang L, Zhong C,
Liang S, Liu H, Qiang Y, Zhang Y and Ni JM: Anti-atherosclerotic
effect of Fermentum rubrum and Gynostemma
pentaphyllum mixture in high-fat emulsion- and vitamin
D3-induced atherosclerotic rats. J Chin Med Assoc. 81:398–408.
2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Qu S, Zhong Y, Shang R, Zhang X, Song W,
Kjems J and Li H: The emerging landscape of circular RNA in life
processes. RNA Biol. 14:992–999. 2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7(e30733)2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Congrains A, Kamide K, Katsuya T, Yasuda
O, Oguro R, Yamamoto K, Ohishi M and Rakugi H: CVD-associated
non-coding RNA, ANRIL, modulates expression of atherogenic pathways
in VSMC. Biochem Biophys Res Commun. 419:612–616. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Lo Sardo V, Chubukov P, Ferguson W, Kumar
A, Teng EL, Duran M, Zhang L, Cost G, Engler AJ, Urnov F, et al:
Unveiling the role of the most impactful cardiovascular risk locus
through haplotype editing. Cell. 175:1796–1810.e20. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–157. 2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Han YN, Xia SQ, Zhang YY, Zheng JH and Li
W: Circular RNAs: A novel type of biomarker and genetic tools in
cancer. Oncotarget. 8:64551–64563. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zeng X, Lin W, Guo M and Zou Q: A
comprehensive overview and evaluation of circular RNA detection
tools. PLOS Comput Biol. 13(e1005420)2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Burd CE, Jeck WR, Liu Y, Sanoff HK, Wang Z
and Sharpless NE: Expression of linear and novel circular forms of
an INK4/ARF-associated non-coding RNA correlates with
atherosclerosis risk. PLoS Genet. 6(e1001233)2010.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li CY, Ma L and Yu B: Circular RNA
hsa_circ_0003575 regulates oxLDL induced vascular endothelial cells
proliferation and angiogenesis. Biomed Pharmacother. 95:1514–1519.
2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Geng HH, Li R, Su YM, Xiao J, Pan M, Cai
XX and Ji XP: The circular RNA Cdr1as promotes myocardial
infarction by mediating the regulation of miR-7a on its target
genes expression. PLoS One. 11(e0151753)2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhao Z, Li X, Gao C, Jian D, Hao P, Rao L
and Li M: Peripheral blood circular RNA hsa_circ_0124644 can be
used as a diagnostic biomarker of coronary artery disease. Sci Rep.
7(39918)2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Song CL, Wang JP, Xue X, Liu N, Zhang XH,
Zhao Z, Liu JG, Zhang CP, Piao ZH, Liu Y, et al: Effect of circular
ANRIL on the inflammatory response of vascular endothelial cells in
a rat model of coronary atherosclerosis. Cell Physiol Biochem.
42:1202–1212. 2017.PubMed/NCBI View Article : Google Scholar
|