Introduction
Degenerative disc disease (DDD) refers to the
biological degeneration of the intervertebral disc under various
factors, which causes the changes in mechanical properties of the
intervertebral disc, causing corresponding damage in adjacent bone
joints and ligaments, affecting the spinal function and even
compressing the spinal cord, nerve roots, and vertebral artery
(1). It has gradually become the
main cause of neck and low back pain, which not only reduces the
quality of people's lives but also causes large Social and economic
burdens. The current treatments of DDD are mainly conservative or
surgical and not focusing on the cause of intervertebral disc
degeneration (IVDD) but only relieve symptoms and some surgery may
even lose adjacent segments (2).
Generally, the intervertebral disc begins to degenerate around the
age of 20, and IVDD becomes increasingly more serious as age
increases. Although the mechanism of IVDD has not been elucidated,
current studies suggest that oxidative stress-induced nucleus
pulposus (NP) cell apoptosis plays a crucial role in the
development of IVDD (3).
In recent years, many studies have found that
degeneration and aging of intervertebral discs are strictly related
to the reduction of NP cells caused by apoptosis (4). Although intervertebral disc cells
account for only 1% of the volume fraction of the intervertebral
disc, these cells can produce factors that affect the synthesis of
extracellular matrix (ECM), such as type I and II collagen,
proteoglycans (PGs), prostaglandins, and nitric oxide, which play
an essential role in maintaining the health of the intervertebral
disc (5). Reactive oxygen species
(ROS) is one of the most critical sources of oxidative stress,
including harmful intermediates such as 02-, OH- and
H2O-. Typically, the body has a series of
antioxidant defense systems: antioxidant enzymes, such as
superoxide dismutase (SOD), glutathione peroxidase (GSH-Px),
catalase; non-enzymatic molecules, such as thioredoxin, and
vitamins A, C, E. The production and elimination of ROS are in a
state of dynamic equilibrium (6).
When the balance between the generation of free
radicals and the antioxidant defense system is broken, ROS
accumulates resulting in cytotoxicity and causes tissue damage.
Excessive ROS acts as a second messenger to trigger apoptotic
signals, thereby modulating apoptosis by regulating the expression
of apoptosis-related genes and the activation of nuclear
transcription factors (7). High
levels of ROS in the intervertebral disc are also indicated to
induce apoptosis of NP cells and accelerate cell senescence to
accelerate IVDD. Activating transcription factor 4 (ATF4) is a key
transcription factor of the endoplasmic reticulum stress signaling
pathway (8). It is stated that ATF4
is involved in the metabolism of amino acid and glucose, as well as
intracellular antioxidant stress. ATF4 is also involved in the
transcriptional regulation of a range of inflammatory factors
(9). Overexpression of ATF4 can
trigger a cascade of apoptotic factors that trigger apoptosis.
Continuous overexpression of ATF4 promotes the upregulation of
apoptosis-inducible genes, such as the transcription factor C/EBP
homologous protein (CHOP), which ultimately leads to cell death
(10). However, the role of ATF4 in
IVDD and NP cell apoptosis caused by excess ROS is not clear. In
this study, degenerative intervertebral disc specimens were
collected, NP cells were isolated to explore the expression of
apoptosis-related proteins including ATF4 and downstream CHOP, in
order to provide a theoretical basis for elucidating the role of
ATF4 in oxidative stress-induced IVDD.
Patients and methods
Collection of patient tissue
samples
In total 12 patients (aged 41 to 72 years, average
age:53 years) who underwent lumbar disc herniation surgery in The
First People's Hospital of Fuyang District (Hangzhou, China) from
September to November 2018 were included in the study. The degree
of IVDD in each operation section was determined according to the
Pfirrmann classification score (11)
based on the Magnetic Resonance Imaging (MRI), the degenerated
degree was divided to grades I to V. The tissues samples from the
patients were grouped into i) Mild degenerated group (G2 and G3),
and ii) Severe degenerated group (G4 and G5). The tissues were
conserved in a sterile cell culture medium immediately after each
operation for the following experiments. The study was approved by
the Ethics Committee of The First People's Hospital of Fuyang
District and signed informed consents were obtained from the
patients and/or guardians. The study was conducted in accordance
with the Declaration of Helsinki.
Isolation and cell culture of NP
cells
The tissues in Pfirrmann classification grades 2
were washed with cold phosphate-buffered saline solution (PBS).
Then the samples were cut into 0.3x0.3 cm2 pieces and
incubated with 0.25% type I collagenase at 37.5˚C overnight for
digesting. Cell pellets were filtrated with a strainer and
re-suspended in Dulbecco's modified Eagle's medium (DMEM)
(containing 10% fetal bovine serum (FBS) and 1%
penicillin-streptomycin, Gibco; Thermo Fisher Scientific, Inc.).
For cell treatment 50 and 100 µM H2O2 was
used to induce ROS; siRNA transfection was used to silence ATF4 and
CHOP gene expression; 20 µM Z-LEHD-FMK (ZLF, a cell-permeable,
irreversible caspase-9 inhibitor; purchased from Selleck) was used
to inhibit the expression of caspase-9.
Malondialdehyde (MDA) analysis
The NP tissue was first lysed and homogenized using
RIPA lysis buffer (Beyotime Institute of Biotechnology). Formulated
TBA stock solution and MDA working fluid were prepared according to
the instructions. Lysate (0.1 ml) was used as a blank control, and
a 0.1 ml sample was added into a 0.2 ml MDA test solution for
measurement. Finally, the absorbance was measured at 532 nm using a
microplate reader.
Cell transfection
NP cells were seeded in 6-well plates at a density
of 2x105 to 4x105/well before transfection.
Transfection was performed using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) + Opti MEM I lipofection according
to the instructions. Negative control siRNA (null-siRNA), ATF4
siRNA, and CHOP siRNA were purchased from Invitrogen; Thermo Fisher
Scientific, Inc. The medium was changed 48 h after transfection and
cultured in a 37˚C, 5% CO2 incubator.
Western blot (WB) analysis
Protein extraction was performed on ice, and the
reaction solution was pre-cooled with ice. The tissues or cells
were washed with cold phosphate-buffered saline (PBS), lysed with
radioimmunoprecipitation assay (RIPA) (Beyotime Institute of
Biotechnology) lysis buffer, centrifuged (at 10,000 x g for 15 min
at 4̊C), discarded, and resuspended in a cell lysate containing
phosphatase inhibitor or protease inhibitor (Beyotime, Shanghai,
China). Bicinchoninic acid (BCA) protein concentration assay kit
(Beyotime Institute of Biotechnology) was used for the
determination of protein quality. Then, the protein was added to
the sodium dodecyl sulphate-polyacrylamide gel electrophoresis
(SDS-PAGE) and transferred onto the polyvinylidene fluoride (PVDF)
membranes (EMD Millipore). After 5% milk blocking, membranes were
coated with primary antibodies against: Collagen II (ab34712), ATF4
(ab31390), CHOP (ab240220), cleaved caspase-9 (ab2324), and β-actin
(as loading control, ab179467) overnight at 4˚C. All the antibodies
were purchased from Abcam. Membranes were then incubated with
secondary antibody for 1 h at room temperature. Chemiluminescent
ECL substrate (Beyotime Institute of Biotechnology) was used to
expose the band.
Immunofluorescence (IF)
NP cells were fixed with 4% paraformaldehyde (PFA)
for 15 min and then incubated with 0.1% Triton X-100 for 15 min at
room temperature and blocked with 5% bovine serum albumin (BSA) for
1 h at room temperature. The cells were washed and incubated with
primary antibodies against collagen II (ab34712; Abcam), caspase-9
(ab219590; Abcam) overnight at 4˚C. NP cells subsequently were
incubated with Alexa Fluor 488 conjugated secondary antibody
(Invitrogen; Thermo Fisher Scientific, Inc.) and
4',6-diamidino-2-phenylindole (DAPI) for 1 h in the dark. The
staining intensity was measured by Image-Pro Plus software (Version
X; Media Cybernetics).
Reverse transcription-polymerase chain
reaction (RT-PCR)
RT-PCR was performed to analyze the expression of
related gene mRNA in NP cells. Total cellular RNA was extracted
with RNAiso kit (Takara) according to the instructions, and reverse
PrimeScript RT reagent kit with gDNA Eraser (Takara) was used for
reverse transcription of RNA into cDNA. SYBR-Green Realtime PCR
Master Mix (Toyobo) reagent was used for the RT-PCR reaction, and
the system was carried out according to the instructions. Gene
expression was calculated by normalization to glyceraldehyde
3-phosphate dehydrogenase (GAPDH) according to the method of
2-ΔΔCt. The primers used for collagen II, ATF4, CHOP,
and caspase-9 are listed in Table
I.
 | Table IPrimer sequences of the genes for
RT-PCR. |
Table I
Primer sequences of the genes for
RT-PCR.
| Gene name | Forward
(5'-3') | Reverse
(5'-3') |
|---|
| Collagen II |
TGGACGATCAGGCGAAACC |
GCTGCGGATGCTCTCAATCT |
| ATF4 |
ATGACCGAAATGAGCTTCCTG |
GCTGGAGAACCCATGAGGT |
| CHOP |
GGAAACAGAGTGGTCATTCCC |
CTGCTTGAGCCGTTCATTCTC |
| Caspase-9 |
CTTCGTTTCTGCGAACTAACAGG |
GCACCACTGGGGTAAGGTTT |
| GAPDH |
ACAACTTTGGTATCGTGGAAGG |
GCCATCACGCCACAGTTTC |
Flow cytometry
NP cells were seeded in a 6-well plate. After
undergoing different treatments, the cells were collected at a
density of ~1x106/well. The cells were washed with cold
PBS and mixed with 5 µl PI and 5 µl Annexin V according to the
Annexin V-PI double staining test kit (Keygen). Following
incubating at room temperature for 15 min in the dark, the
apoptosis rate of each group was measured by flow cytometry. The
total apoptosis ratio was determined by the collection of both
early and late apoptotic cells.
Statistical analysis
All statistical analyses were performed using
Statistical Product and Service Solutions (SPSS) 13.0 statistical
software (SPSS Inc.). Data were expressed as the mean ± SD
(standard deviation). Differences between two groups were analyzed
by using the paired Student's t-test. A comparison between multiple
groups was made using a One-way analysis of variance (ANOVA) test
followed by the Post Hoc Test (Least Significant Difference).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Human intervertebral disc degenerates
combining with increased ROS and apoptosis
We collected degenerated intervertebral disc tissues
from the patients undergoing herniated disc surgery and divided
them into two groups according to the Pfirrmann score of disc
degeneration. Most patients with IDD who need surgery in the clinic
are highly degenerative. It is difficult for us to obtain a disc
without any degeneration. Therefore, we used the highly degenerated
specimens to compare with the mildly degenerated specimens. To
determine the level of oxygen radicals in the mild and severe
degenerated disc samples, MDA assay was performed, and the data
showed a higher degree of degeneration expressed the MDA level
(Fig. 1A). Collagen II is the main
protected element of ECM secreted by the NP cells that decreased
with the degeneration of NP cells (12). The result of western blot analysis
indicated the severely degenerated discs expressed a lower content
of collagen II and an increased level of ATF4, CHOP, and caspase-9
compared with the mild group (Fig.
1B and C). RT-PCR was also used
to analyze the mRNA expression of collagen II, ATF4, CHOP, and
caspase-9 of the intervertebral disc samples, and the results were
totally parallel to the protein levels (Fig. 1D). The data suggest that severely
degenerative disc is in a higher state of oxidative stress and a
severe level of apoptosis related to the activation of
ATF4/CHOP/caspase-9.
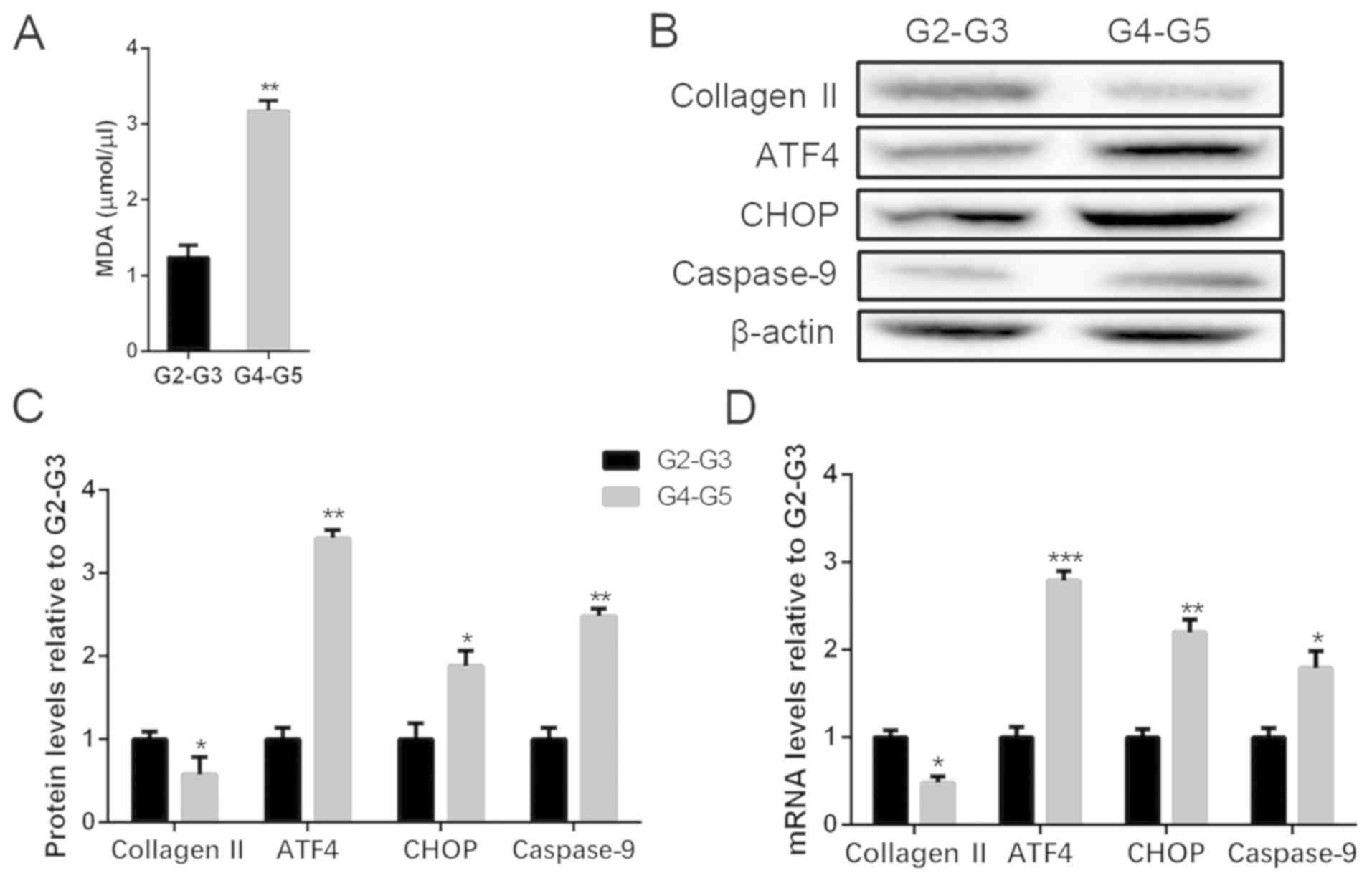 | Figure 1ROS and apoptosis levels in human
degenerated intervertebral disc tissues. Tissues were lysed and ROS
measured with MDA methods. Total protein and mRNA were extracted
from the tissues in mild (G2-G3) and severe (G4-G5) degeneration.
(A) MDA level of different degenerated degrees of disc tissues. (B
and C) The protein levels of collagen II, ATF4, CHOP and caspase-9
were determined by (B) Western blot and (C) quantification
analysis. (D) mRNA levels of collagen II, ATF4, CHOP and caspase-9
were determined by RT-PCR. The values are mean ± SD of three
independent experiments (n=3). (*P<0.05,
**P<0.01, ***P<0.001, compared with
G2-G3 group). ROS, reactive oxygen species; ATF4, activating
transcription factor 4; CHOP, C/EBP homologous protein; RT-PCR,
reverse transcription-polymerase chain reaction. |
H2O2 treatment
increases NP cell apoptosis by ATF4/CHOP and caspase-9
upregulation
To explore the effect of excess ROS in the progress
of NP cell degeneration, different concentrations of
H2O2 (50 or 100 µM) were used as a source of
oxygen radicals. As shown in Fig. 2A
and B, the expression of collagen II
was decreased, resulting from the H2O2
treatment; in addition, higher concentration caused more reduction
of collagen II. Besides, H2O2 promoted
caspase-9, the apoptosis maker, of NP cells as well. Moreover, the
mRNA and protein levels of collagen II, ATF4, CHOP and caspase-9
were also measured by RT-PCR and WB (Fig. 2C-E). The data indicated that collagen
II was reduced caused by H2O2 treatment with
a dose-dependence compared with the control; ATF4 and CHOP
expression were increased, which were more significant in the high
dose H2O2. In addition,
H2O2 also activated caspase-9 expression
compared with the control. Flow cytometry was also used to measure
apoptotic cells, and the result indicated a higher apoptotic
population after H2O2 treatment (Fig. 2F and G). The results suggested
H2O2 treatment promoted NP cell degeneration
and apoptosis, which might be related to the upregulation of
ATF4/CHOP and caspase-9.
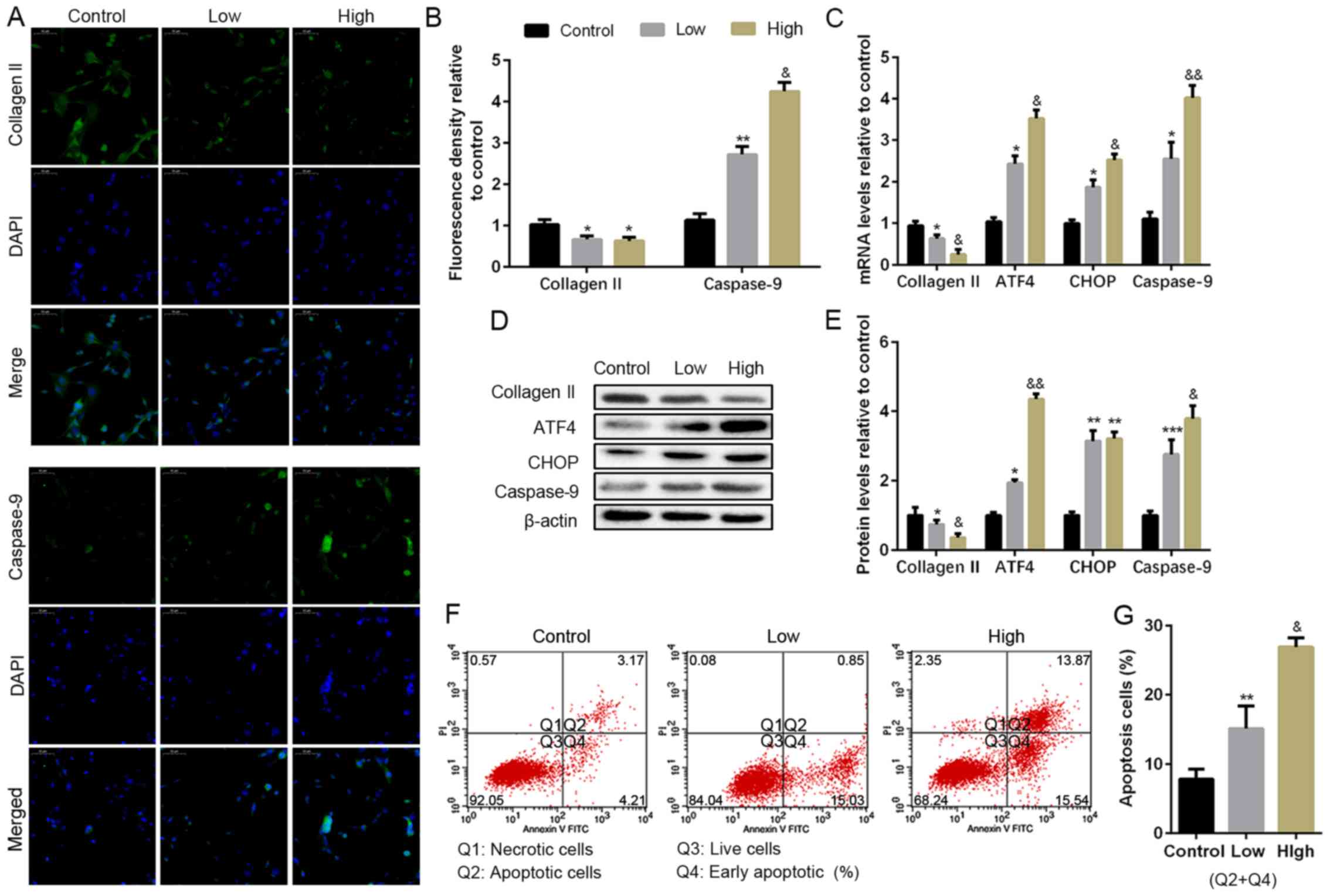 | Figure 2Apoptosis levels in
H2O2 induce NP cells in vitro. NP
cells of G2 tissues were cultured with a low or high concentration
of H2O2 (50 or 100 µM) for 24 h. (A and B)
The protein expression level of collagen II and caspase-9 were
determined by (A) IF (magnification, x400) and (B) quantification
analysis. (C) The mRNA expression levels of collagen II, ATF4,
CHOP, and caspase-9 were assayed by RT-PCR. (D and E) The protein
expression levels of collagen II, ATF4, CHOP and caspase-9 were
determined by (D) WB and (E) quantification analysis. (F and G) The
ratio of apoptotic cells was analyzed by (F) flow cytometry and (G)
quantification analysis. The values are mean ± SD of three
independent experiments (n=3). (*P<0.05,
**P<0.01, ***P<0.001, compared with
control; &P<0.05, &&P<0.01,
compared with 50 µM H2O2). ATF4, activating
transcription factor 4; CHOP, C/EBP homologous protein; WB, western
blot; NP, nucleus pulposus; RT-PCR, reverse
transcription-polymerase chain reaction; IF,
immunofluorescence. |
Silencing of ATF4 alleviates
H2O2-induced NP cells apoptosis
To determine the effect of ATF4 in
H2O2-induced NP cell apoptosis, the ATF4 gene
was silenced via transfecting with siRNA targeting ATF4, and
null-siRNA was used as a negative control. The non-transfected NP
cells and the siRNA transfected NP cells were cultured with 100 µM
H2O2 for 24 h. Silencing of ATF4 played a
protective effect on collagen II expression in the treatment of
H2O2. Furthermore, IF also showed ATP4
deficiency reduced the caspase-9 expression compared with the
H2O2 group, which was also confirmed in the
WB and RT-PCR (Fig. 3A and B). The silencing of ATF4 contributed to the
decrease of CHOP mRNA expression, which could also be responsible
for the reduction of caspase-9 (Fig.
3C). WB was also performed to analyze the protein expression of
CHOP, which was consistent with the result of RT-PCR (Fig. 3D and E). The data of flow cytometry also
confirmed silencing of ATF4 decreased the apoptotic cells compared
with the H2O2 group (Fig. 3F and G). In brief, the silencing of ATF4 blocked
the ATF4/CHOP pathway for the activation of downstream gene
caspase-9, which resulted in the reduction of NP cell
apoptosis.
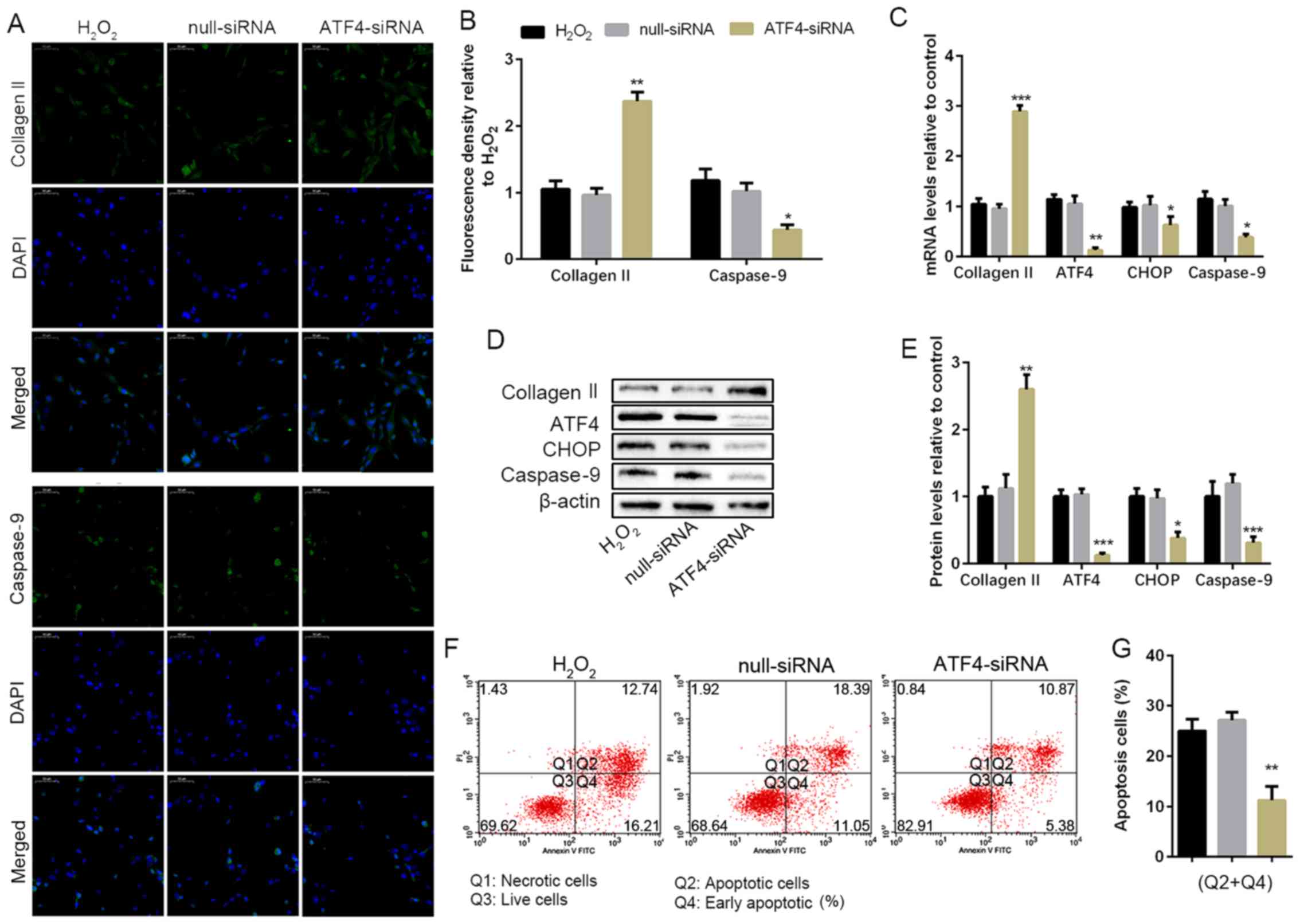 | Figure 3ATF4 deficiency weakens apoptosis and
protects human NP cell degeneration. NP cells of G2 tissues without
or with the ATF4-siRNA transfection were cultured with 100 µM of
H2O2 for 24 h. (A and B) The protein
expression level of collagen II and caspase-9 were determined by
(A) IF (magnification, x400) and (B) quantification analysis. (C)
The mRNA expression levels of collagen II, ATF4, CHOP and caspase-9
were assayed by RT-PCR. (D and E) The protein expression levels of
collagen II, ATF4, CHOP and caspase-9 were determined by (D) WB and
(E) quantification analysis. (F and G) The ratio of apoptotic cells
was analyzed by (F) flow cytometry and (G) quantification analysis.
The values are mean ± SD of three independent experiments (n=3).
(*P<0.05, **P<0.01,
***P<0.001, compared with 100 µM
H2O2). ATF4, activating transcription factor
4; CHOP, C/EBP homologous protein; WB, western blot; NP, nucleus
pulposus; RT-PCR, reverse transcription-polymerase chain reaction;
IF, immunofluorescence. |
Silencing of CHOP alleviates
H2O2-induced NP cell apoptosis
To determine the effect of CHOP, the downstream
target of ATF4, in H2O2-induced NP cell
apoptosis, the CHOP gene was blocked by transfecting with
CHOP-siRNA. The non-transfected NP cells and the siRNA transfected
NP cells were cultured with 100 µM H2O2 for
24 h. However, the silencing of CHOP significantly upregulated
collagen II protein expression and decreased the caspase-9
expression compared with the H2O2 group
(Fig. 4A and B). Similar results were obtained from the
RT-PCR and WB analysis (Fig. 4C-E).
Furthermore, both the mRNA and protein expression of CHOP were
significantly reduced resulting from siRNA transfection, but CHOP
silencing did not affect ATF4 expression (Fig. 4C-E). The number of apoptotic NP cells
was obviously reduced compared with the H2O2
group when the CHOP gene was silenced through flow cytometry assay
(Fig. 4F and G). The result indicated silencing of CHOP
broke the ATF4/CHOP pathway for the activation of caspase-9, which
contributed to the reduction of NP cell apoptosis in the
H2O2 situation.
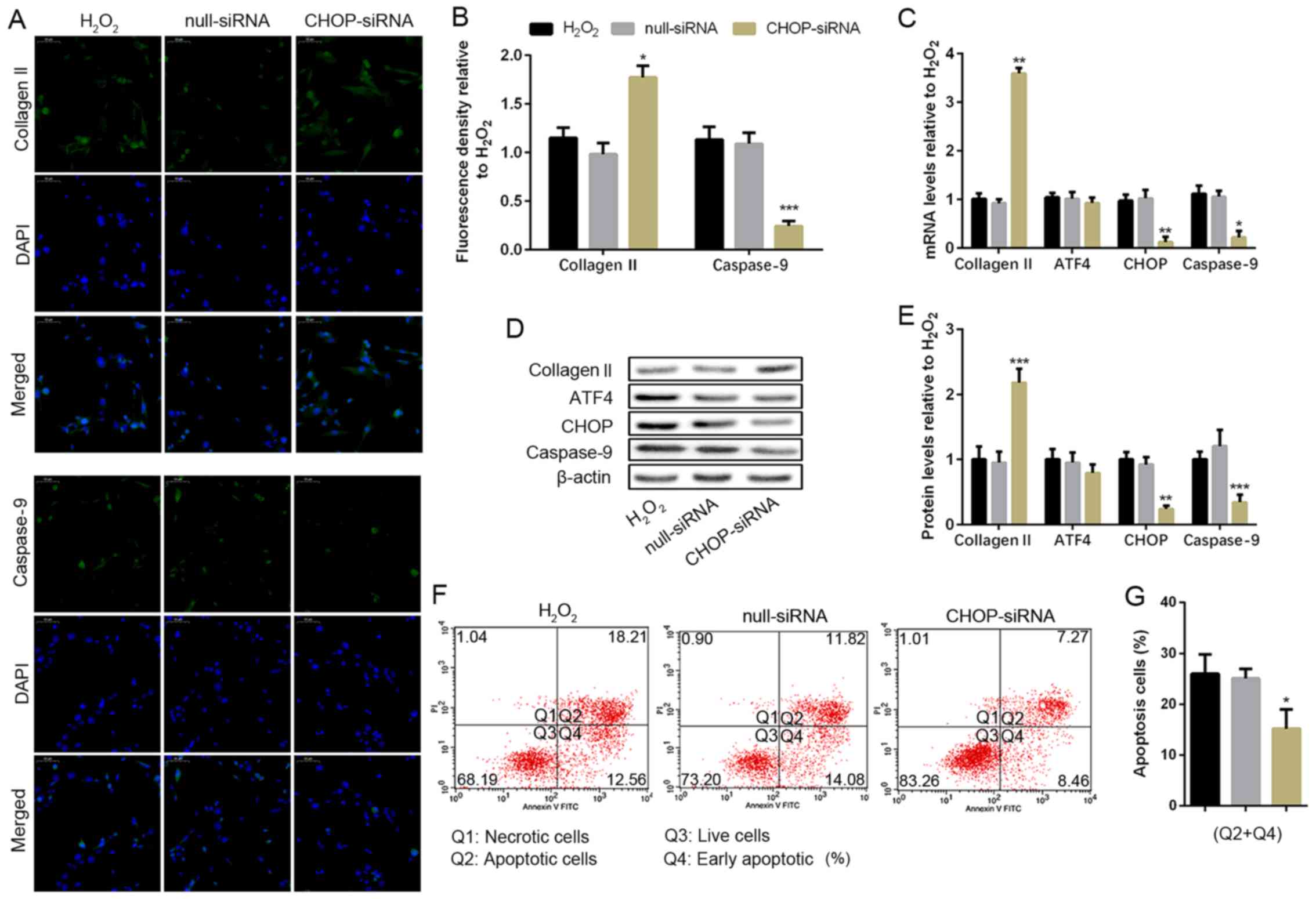 | Figure 4CHOP deficiency weakens apoptosis and
protects human NP cell degeneration. NP cells of G2 tissues without
or with the CHOP-siRNA transfection were cultured with 100 µM of
H2O2 for 24 h. (A and B) The protein
expression level of collagen II and caspase-9 were determined by
(A) IF (magnification, x400) and (B) quantification analysis. (C)
The mRNA expression levels of collagen II, ATF4, CHOP and caspase-9
were assayed by RT-PCR. (D and E) The protein expression levels of
collagen II, ATF4, CHOP, and caspase-9 were determined by (D) WB
and (E) quantification analysis. (F and G) The ratio of apoptotic
cells was analyzed by (F) flow cytometry and (G) quantification
analysis. The values are mean ± SD of three independent experiments
(n=3). (*P<0.05, **P<0.01,
***P<0.001, compared with 100 µM
H2O2). ATF4, activating transcription factor
4; CHOP, C/EBP homologous protein; WB, western blot; NP, nucleus
pulposus; RT-PCR, reverse transcription-polymerase chain reaction;
IF, immunofluorescence. |
Inhibiting caspase-9 alleviates
H2O2-induced NP cell apoptosis
Previous results of this study suggested caspase-9
was activated by ATF4/CHOP pathway. It was investigated whether the
change of caspase-9 levels would affect ATF4 or CHOP expression and
the degree of NP cell apoptosis caused by
H2O2. ZLF (13,14), a
widely used inhibitor of caspase-9 was used to pretreat NP cells
for 24 h. The chemical structural formula of ZLF is shown in
Fig. 5A. ZLF pretreated NP cells
showed a protected role of collagen II production compared with the
non-pretreated group (Fig. 5B-E).
Under the condition of caspase-9 suppression, we did not achieve a
significant difference in the protein expression of ATF4 and CHOP,
indicating that the level of caspase-9 cannot affect ATF4/CHOP
expression (Fig. 5D and E). Moreover, as a result of the suppression
of caspase-9, the apopotic NP cell population was lower than the
H2O2 group (Fig.
5F and G). In short, caspase-9
is a downstream target of the ATF4/CHOP pathway in the NP cells,
and ZLF is effective in suppressing apoptosis in
H2O2-treated NP cells.
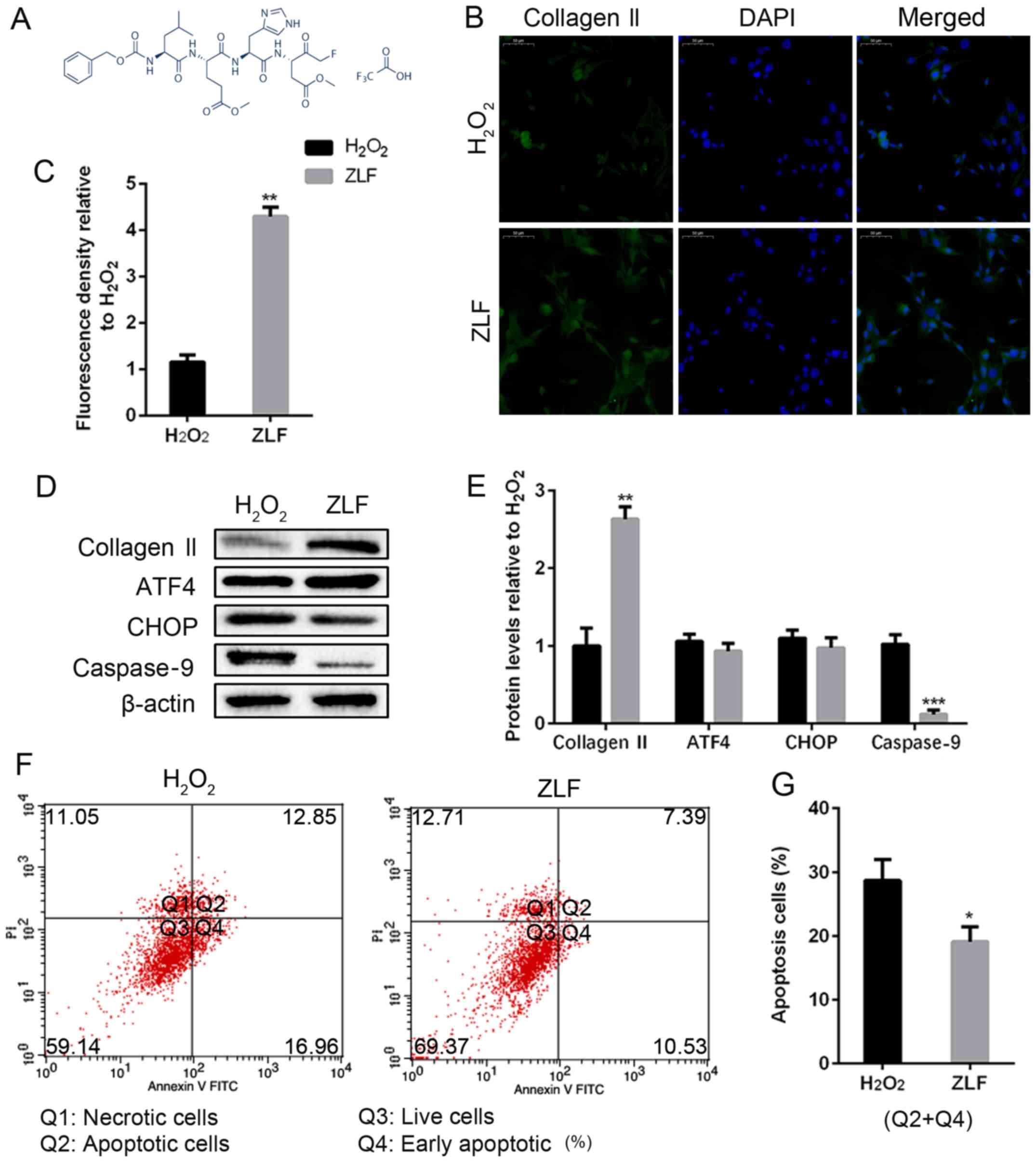 | Figure 5ZLF inhibits apoptosis and protects
human NP cell degeneration. NP cells of G2 tissues were pretreated
with ZLF (20 µM) for 24 h. Then, NP cells without or with ZLF
pretreatment were cultured with 100 µM of
H2O2 for 24 h. (A) The chemical structural
formula of ALF. (B and C) The protein expression level of collagen
II was determined by (B) IF (magnification, x400) and (C)
quantification analysis. (D and E) The protein expression levels of
collagen II, ATF4, CHOP and caspase-9 were determined by (D) WB and
(E) quantification analysis. (F and G) The ratio of apoptotic cells
was analyzed by (F) flow cytometry and (G) quantification analysis.
The values are mean ± SD of three independent experiments (n=3).
(*P<0.05, **P<0.01,
***P<0.001, compared with 100 µM
H2O2). ATF4, activating transcription factor
4; CHOP, C/EBP homologous protein; WB, western blot; NP, nucleus
pulposus; IF, immunofluorescence. |
Discussion
The expression of ROS has been confirmed to be
increased during the progress of IVDD, suggesting that ROS may play
an essential role in the pathological process of IVDD (15). In the present study, the
intervertebral disc with severe degenerated degree expressed a
higher level of MDA compared with the mild, suggesting the degree
of degeneration is positively related to the content of ROS.
Apoptosis is the natural physiological process of the body as well
as cell production. Both coexist and maintain dynamic balance. This
balance is a necessary condition for maintaining the morphological
stability of the body tissues and organs, the physiological
functions of the body, and the security of the internal and
external environment. If the cell apoptosis is too much or too
little, this balance damage will lead to disease. Excessive
apoptosis of NP cells is the direct cause of IVDD (16). Ha et al (17) found that the cell apoptosis rate of
patients with intervertebral disc prolapse was 74.3%, disc
herniation was 42.8%, while the regular control group was only 28%.
Zhao et al (18) confirmed
that various stimulating factors could induce apoptosis of
intervertebral disc cells and further cause the reduction of
intervertebral disc matrix and promote the IVDD process. Numbers of
studies have elucidated that low levels of oxygen free radicals can
induce apoptosis. At the same time, ROS, such as
O2-, H2O2, lipid
peroxide, and NO, is considered to be widely involved in apoptosis.
As shown in our results, apoptosis-related ATF4, CHOP and caspase-9
were significantly decreased in the severely degenerated disc
tissues. Therefore, ROS is suggested to be related to apoptosis in
the degenerated intervertebral disc.
Collagen Ⅱ, secreted by NP cells, is one of the most
important components of the extracellular matrix, and its content
is an important indicator of whether NP cells are functional. In
this experiment, H2O2 was used to upregulate
ROS in inducing apoptosis of NP cells (19). As the concentration of
H2O2 increased, the expression of collagen II
in NP cells decreased, but apoptosis level increased significantly,
along with the upregulation of ATF4, CHOP and caspase-9 expression.
A large number of reports have reported that ROS participate in
cell apoptosis by activating ATF4/CHOP pathway. Xian et al
(20) stated ROS activates apoptosis
in adriamycin-treated, bortezomib sensitized human osteosarcoma
cells via p-eIF2α/ATF4/CHOP axis. Zong et al (21) reported that upregulating ATF4
expression could increase cellular ROS and the sensitivity to
apoptosis. ATF4 is upregulated by stress signals containing
oxidative stress, hypoxia, and endoplasmic reticulum stress. ATF4
also affects the expression of genes involved in oxidative stress
(22). The ATF4 gene expression was
silenced resulting in the downregulation of both CHOP and
caspase-9, reversing the negative effect of
H2O2 on the NP cells. In addition, CHOP, a
transcription factor, is expressed at deficient levels under normal
conditions and is significantly elevated in endoplasmic reticulum
stress through the activation of ATF4(23). After suppressing the expression of
CHOP by gene silencing, caspase-9 and apoptotic cell population
were also obviously reduced without affecting ATF4. They
contributed to upregulation of the expression of collagen II under
conditions of oxidative stress, which confirms CHOP as the
downstream gene of ATF4 in the apoptosis-related progress.
Among the CHOP-mediated endoplasmic reticulum
stress-responsive apoptotic pathways, the upstream regulation
mechanism is basically precise, but its downstream mechanism
remains to be further studied. The present study indicates that the
target genes of CHOP may include Bcl-2, TRB3, GADD34 and DOCs
(24,25). Regardless of which targets downstream
of CHOP, activated caspase-9 will be initiated to trigger apoptosis
at the end (26,27). Regardless of silencing by ATF4 or
CHOP in H2O2 treated NP cells, apoptosis was
decreased significantly as well as the caspase-9 level in the
present study. As previously noted, ROS mediated caspase-9
activation increases apoptosis of EBV-transformed B cells (28); in addition, inhibiting caspase-9 is
efficacious to suppress apoptosis (29,30). ZLF
is a novel specific inhibitor of caspase-9(31). After ZLF treatment, NP cells
expressed a raised level of collagen II and the apoptotic cell
population decreased. Though ZLF did not affect the expression of
ATF4 and CHOP, it achieved the same result in reversing the
H2O2 induced apoptosis.
In conclusion, this study recovered, for the first
time, crosstalk between CHOP and ATF4 in the development of IDD,
especially under the H2O2 mediated high-ROS
situation, which involves caspase-9 related apoptosis. With the
aggravation of IVDD, the ATF4/CHOP pathway is activated, resulting
in apoptosis of NP cells. Blocking ATF4 or CHOP can partly suppress
caspase-9 and alleviate ROS related NP cell apoptosis, which also
protects NP cell degeneration. Suppressing the expression of
caspase-9 by ZLF can protect NP cells from apoptosis. In
conclusion, H2O2 induces NP cell apoptosis by
ATF4/CHOP signaling pathway, and blocking ATF4/CHOP or caspase-9
can protect NP cell degeneration via apoptosis suppression.
Additionally, to improve the limitation and verify the findings of
the present study, we plan to use siRNA to suppress the expression
of caspase-9 and establish animal model to set up a
non-degenerative NP tissue control group in the next project.
Additionally, human studies with a control group are also required
to verify the findings of this study.
In conclusion, in this study,
H2O2 was found to promote NP cell apoptosis
by activating the ATF4/CHOP signaling pathway resulting in the
upregulation of caspase-9. Interdict of ATF4, CHOP, or caspase-9
contributed to the reduction of apoptosis caused by
H2O2.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YL designed the study and performed the experiments,
acquired and analyzed the data, prepared the manuscript, and read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The First People's Hospital of Fuyang District (Hangzhou, China).
Signed informed consents were obtained from the patients and/or
guardians.
Patients consent for publication
Not applicable.
Competing interests
The author declares that he has no competing
interests.
References
|
1
|
Molinos M, Almeida CR, Caldeira J, Cunha
C, Gonçalves RM and Barbosa MA: Inflammation in intervertebral disc
degeneration and regeneration. J R Soc Interface.
12(20141191)2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hanaei S, Abdollahzade S, Khoshnevisan A,
Kepler CK and Rezaei N: Genetic aspects of intervertebral disc
degeneration. Rev Neurosci. 26:581–606. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Chen J, Xuan J, Gu YT, Shi KS, Xie JJ,
Chen JX, Zheng ZM, Chen Y, Chen XB, Wu YS, et al: Celastrol reduces
IL-1β induced matrix catabolism, oxidative stress and inflammation
in human nucleus pulposus cells and attenuates rat intervertebral
disc degeneration in vivo. Biomed Pharmacother. 91:208–219.
2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Yang RS, Wang YH, Ding C, Su XH and Gong
XB: MiR-146 regulates the repair and regeneration of intervertebral
nucleus pulposus cells via Notch1 pathway. Eur Rev Med Pharmacol
Sci. 23:4591–4598. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Vo NV, Hartman RA, Patil PR, Risbud MV,
Kletsas D, Iatridis JC, Hoyland JA, Le Maitre CL, Sowa GA and Kang
JD: Molecular mechanisms of biological aging in intervertebral
discs. J Orthop Res. 34:1289–1306. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Feng C, Yang M, Lan M, Liu C, Zhang Y,
Huang B, Liu H and Zhou Y: ROS: Crucial intermediators in the
pathogenesis of intervertebral disc degeneration. Oxid Med Cell
Longev. 2017(5601593)2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ishibashi H, Tonomura H, Ikeda T, Nagae M,
Sakata M, Fujiwara H, Tanida T, Mastuda K, Kawata M and Kubo T:
Hepatocyte growth factor/c-met promotes proliferation, suppresses
apoptosis, and improves matrix metabolism in rabbit nucleus
pulposus cells in vitro. J Orthop Res. 34:709–716. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Srivastava RK, Li C, Ahmad A, Abrams O,
Gorbatyuk MS, Harrod KS, Wek RC, Afaq F and Athar M: ATF4 regulates
arsenic trioxide-mediated NADPH oxidase, ER-mitochondrial crosstalk
and apoptosis. Arch Biochem Biophys. 609:39–50. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Luís A, Martins JD, Silva A, Ferreira I,
Cruz MT and Neves BM: Oxidative stress-dependent activation of the
eIF2α-ATF4 unfolded protein response branch by skin sensitizer
1-fluoro-2,4-dinitrobenzene modulates dendritic-like cell
maturation and inflammatory status in a biphasic manner
[corrected]. Free Radic Biol Med. 77:217–229. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Luo J, Xia Y, Luo J, Li J, Zhang C, Zhang
H, Ma T, Yang L and Kong L: GRP78 inhibition enhances ATF4-induced
cell death by the deubiquitination and stabilization of CHOP in
human osteosarcoma. Cancer Lett. 410:112–123. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Oh CH, Kim DY, Ji GY, Kim YJ, Yoon SH,
Hyun D, Kim EY, Park H and Park HC: Cervical arthroplasty for
moderate to severe disc degeneration: Clinical and radiological
assessments after a minimum follow-up of 18 months - Pfirrmann
grade and cervical arthroplasty. Yonsei Med J. 55:1072–1079.
2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yang B and O'Connell GD: Effect of
collagen fibre orientation on intervertebral disc torsion
mechanics. Biomech Model Mechanobiol. 16:2005–2015. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Jin A, Shi XC, Liu Y, Sun J and Ji H:
Docosahexaenoic acid induces PPARγ-dependent preadipocytes
apoptosis in grass carp Ctenopharyngodon idella. Gen Comp
Endocrinol. 266:211–219. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ma Y, Zhu B, Yong L, Song C and Liu X, Yu
H, Wang P, Liu Z and Liu X: Regulation of intrinsic and extrinsic
apoptotic pathways in osteosarcoma cells following oleandrin
treatment. Int J Mol Sci. 23(1950)2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Cai X, Liu Y, Hu Y, Liu X, Jiang H, Yang
S, Shao Z, Xia Y and Xiong L: ROS-mediated lysosomal membrane
permeabilization is involved in bupivacaine-induced death of rabbit
intervertebral disc cells. Redox Biol. 18:65–76. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Tschoeke SK, Hellmuth M, Hostmann A,
Robinson Y, Ertel W, Oberholzer A and Heyde CE: Apoptosis of human
intervertebral discs after trauma compares to degenerated discs
involving both receptor-mediated and mitochondrial-dependent
pathways. J Orthop Res. 26:999–1006. 2008.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Ha KY, Koh IJ, Kirpalani PA, Kim YY, Cho
YK, Khang GS and Han CW: The expression of hypoxia inducible
factor-1alpha and apoptosis in herniated discs. Spine.
31:1309–1313. 2006.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhao CQ, Jiang LS and Dai LY: Programmed
cell death in intervertebral disc degeneration. Apoptosis.
11:2079–2088. 2006.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang Z, Wang D, Li Y and Zhang X:
Protective effects of verapamil against
H2O2-induced apoptosis in human lens
epithelial cells. Biomol Ther (Seoul). 22:553–557. 2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Xian M, Cao H, Cao J, Shao X, Zhu D, Zhang
N, Huang P, Li W, Yang B, Ying M, et al: Bortezomib sensitizes
human osteosarcoma cells to adriamycin-induced apoptosis through
ROS-dependent activation of p-eIF2α/ATF4/CHOP axis. Int J Cancer.
141:1029–1041. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zong Y, Feng S, Cheng J, Yu C and Lu G:
Upregulated ATF4 Expression increases cell sensitivity to apoptosis
in response to radiation. Cell Physiol Biochem. 41:784–794.
2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ameri K and Harris AL: Activating
transcription factor 4. Int J Biochem Cell Biol. 40:14–21.
2008.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Averous J, Bruhat A, Jousse C, Carraro V,
Thiel G and Fafournoux P: Induction of CHOP expression by amino
acid limitation requires both ATF4 expression and ATF2
phosphorylation. J Biol Chem. 279:5288–5297. 2004.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Sok J, Wang XZ, Batchvarova N, Kuroda M,
Harding H and Ron D: CHOP-Dependent stress-inducible expression of
a novel form of carbonic anhydrase VI. Mol Cell Biol. 19:495–504.
1999.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Fu HY, Okada K, Liao Y, Tsukamoto O,
Isomura T, Asai M, Sawada T, Okuda K, Asano Y, Sanada S, et al:
Ablation of C/EBP homologous protein attenuates endoplasmic
reticulum-mediated apoptosis and cardiac dysfunction induced by
pressure overload. Circulation. 122:361–369. 2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yang Y, Liu L, Naik I, Braunstein Z, Zhong
J and Ren B: Transcription factor C/EBP homologous protein in
health and diseases. Front Immunol. 8(1612)2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Li P, Zhou L, Zhao T, Liu X, Zhang P, Liu
Y, Zheng X and Li Q: Caspase-9: Structure, mechanisms and clinical
application. Oncotarget. 8:23996–24008. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Park GB, Choi Y, Kim YS, Lee HK, Kim D and
Hur DY: ROS and ERK1/2-mediated caspase-9 activation increases XAF1
expression in dexamethasone-induced apoptosis of EBV-transformed B
cells. Int J Oncol. 43:29–38. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chen Y, Sun P, Bai W and Gao A: MiR-133a
regarded as a potential biomarker for benzene toxicity through
targeting Caspase-9 to inhibit apoptosis induced by benzene
metabolite (1,4-Benzoquinone). Sci Total Environ. 571:883–891.
2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Tamaki H, Harashima N, Hiraki M, Arichi N,
Nishimura N, Shiina H, Naora K and Harada M: Bcl-2 family
inhibition sensitizes human prostate cancer cells to docetaxel and
promotes unexpected apoptosis under caspase-9 inhibition.
Oncotarget. 5:11399–11412. 2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Roberts JL, Booth L, Conley A,
Cruickshanks N, Malkin M, Kukreja RC, Grant S, Poklepovic A and
Dent P: PDE5 inhibitors enhance the lethality of standard of care
chemotherapy in pediatric CNS tumor cells. Cancer Biol Ther.
15:758–767. 2014.PubMed/NCBI View Article : Google Scholar
|














