Introduction
Fatigue is a common condition that is caused by
strenuous physical activity and harmful diseases, including
grueling exercise (1) and cancer
(2,3). Although symptoms of fatigue can be
eliminated and relieved by appropriate exercise in many cases
(4-6),
people of different nationalities, occupations and ages all
experience fatigue due to current social pressure, such as work and
competitive pressure. If left unchecked, fatigue not only reduces
productivity and that quality of life but can also contribute to
the progression of fatigue itself.
Mitochondria are important intracellular organelles
that produce the adenosine triphosphate (ATP) required for the
contraction of skeletal muscles in mammals. Previous studies have
demonstrated that muscle mitophagy was associated with metabolic
disorders, including obesity and skeletal muscle dyskinesia
concomitant with Parkinson's disease (7,8).
Long-term and appropriate exercise have been found to increase
plasticity and mitochondrial biogenesis in skeletal muscle.
Mitochondria in skeletal muscle can also exhibit physiological
adaptation from oxygen-dependent aerobic glycolysis to oxidative
phosphorylation under anaerobic conditions (9). Skeletal muscles account for 30% of the
body's weight in humans that serve as the storehouse of amino acids
and carbohydrates, in addition to being the converter of chemical
energy into mechanical energy (10).
To maintain a satisfactory level of health and quality of life, a
sufficient number of mitochondria in skeletal muscles is required.
For both professional athletes and the general public, strength
building and the maintenance of various types of movement or
posture require healthy skeletal muscles (11). Increases in the number of
mitochondria can improve the performance of professional athletes
and the health of ordinary people. Indeed, a previous study has
reported that chronic exercise can drive mitochondria biogenesis
and fusion in skeletal muscles of healthy people who trained
regularly for an extended period of time after a previously
sedentary period (12). Notably,
high-intensity exercise can lead to the excess autophagy of
skeletal muscle mitochondria, by process by which damaged
mitochondria are removed by autophagic lysosomal complexes
(11,13). However, a reduction in the number of
mitochondria caused by excessive autophagy can lead to skeletal
muscle weakness (12). Therefore,
understanding the mechanism that lead to exercise-induced
mitochondrial dysfunction can provide insights into novel treatment
options for fatigue.
Rhodiola crenulata oral liquid (RCOL) is a
widely applied agent in traditional Chinese medicine for the
treatment of ailments such as altitude sickness and physical
weakness (14). Rhodiola
crenulata is an endangered species of perennial herbaceous
plant from the Crassulaceae family that is distributed
mainly in Tibet. In traditional Tibetan herbal medicine, it is
believed to increase physical endurance (15-17),
to confer anti-fatigue (18) and
anti-hypoxia properties (19).
Additionally, previous studies have reported that Rhodiola
crenulate, specifically its main active ingredient,
salidroside, have a diverse range of pharmacological effects,
including anti-inflammatory (20,21),
neuroprotective (22-24),
eyesight protective (25),
anti-radiation (26) and anti-cancer
activities (27). However, the
potential effects of RCOL on exhaustive exercise (EE)-induced
fatigue in skeletal muscle remain poorly understood. Therefore, in
the present study, an established in vivo mouse model of
muscle fatigue was used to investigate the potential anti-fatigue
effects of RCOL administration, in addition to the underlying
mechanism, specifically mitophagy.
Materials and methods
Ethical statement
The present study was approved by The Animal
Research Ethics Committee of Chengdu University of Traditional
Chinese Medicine. All experimental protocols were performed in
accordance with The Animal Care Guidelines, conforming to The
Health Guide for the Care and Use of Laboratory Animals (28).
Reagents
Commercial RCOL was purchased from Tibet Tibetan
Medicine Group Co., Ltd. (authorization no. B20070002). Primary
antibodies against PTEN-induced kinase protein 1 (PINK1; cat. no.
BC100-494) were purchased from Novus Biologicals, LLC. Antibodies
specific for Parkin (cat. no. 2132), ubiquitin (cat. no. 3933),
LC3-II/LC3-I (cat. no. 4108), sequestosome 1 (SQSTM 1/p62; cat. no.
5114), citrate synthase (CS, cat. no. 14309), and GAPDH (cat. no.
5174) were obtained from Cell Signaling Technology, Inc.
Lactic acid (LA; cat. no. A019-2), creatine
phosphokinase (CK; cat. no. A032), lactic dehydrogenase (LDH; cat.
no. A020-1), malondialdehyde (MDA; cat. no. A003-1), superoxide
dismutase (SOD; cat. no. A001-2), catalase (CAT; cat. no. A007-1),
total anti-oxidative capability (T-AOC; cat. no. A015), succinate
dehydrogenase (SDH, cat. no. A022), and
Na+-K+-ATPase (cat. no. A070-2) activity
detection kits were purchased from Nanjing Jiancheng Bioengineering
Institute. Glycogen content detection kits (cat. no. BC0345) and
2.5% Gluta transmission electron microscope specimen fixative (cat.
no. P1126) were provided by Beijing Solarbio Science &
Technology Co., Ltd. Xylene, acetone, phosphoric acid, osmium acid,
uranyl acetate, lead citrate and paraformaldehyde were purchased
from Chengdu Chron Chemicals Co., Ltd.
High performance liquid chromatography
(HPLC) analysis of gallic acid, salidroside, tyrosol, catechin,
caffeic acid and p-coumaric acid in RCOL
To quantify the main constituents in RCOL, gallic
acid (cat. no. M-017-181216), salidroside (cat. no. H-040-170403),
tyrosol (cat. no. L-042-180426), catechin (cat. no. E-011-180725),
caffeic acid (cat. no. K-003-181216) and p-coumaric acid (cat. no.
D-032-171216) were used as reference substances (all purchased at
98% purity from Chengdu Ruifen Si Biotechnology Co., Ltd.). The
HPLC protocol was performed as described previously (22,29).
Briefly, a mixture of the reference substances (0.1306 g/ml gallic
acid, 0.1394 g/ml salidroside, 0.0646 g/ml tyrosol, 0.0038 g/ml
catechin, 0.1140 g/ml caffeic acid and 0.0222 g/ml p-coumaric acid)
and RCOL were analyzed using the Agilent 1260 Infinity System
(Agilent Technologies, Inc.). The samples were filtered through
0.45-µm microporous membranes, following which 10 µl of the sample
was loaded onto a C18 chromatographic column (Inner
diameter x length: 4.6x250 mm; particle size: 5 µm; cat. no.
5020-39033, Yili Scientific Instrument Co., Ltd.) and separated
using an initial mobile phase consisting of 0.2% phosphoric acid
solution and acetonitrile. Gradient elution was then performed
using acetonitrile as follows: i) 7% for 0-8 min; ii) 7-13% for
8-27 min; iii) 13-15% for 27-30 min; iv) 15-18% for 30-45 min; and
v) 18% for 45-70 min at a column flow rate of 0.5 ml/min and a
temperature of 25˚C. The curve regression equation was obtained by
diluting 1.306 mg/ml gallic acid, 1.394 mg/ml salidroside, 0.646
mg/ml tyrosol, 0.222 mg/ml p-coumaric acid, 1.114 mg/ml caffeic
acid and 0.038 mg/ml catechin by 2, 4, 8 and 16 times,
respectively. Corresponding peaks were determined at a wavelength
of 275 nm.
Animals
Male, six-week-old Institute of Cancer Research
(ICR) mice (25±2 g) were provided by Chengdu Dashuo Experimental
Animal Co., Ltd. The license number for the use of experimental
animals is SYXK (Chuan) 2013-124. All mice were kept under standard
conditions at 23±2˚C and 50-60% humidity, with a 12-h light/dark
cycle. Mice with similar mean body weights were randomly divided
and housed in groups of five animals per cage. They were provided
with free access to standard diet and sterilized water. All animal
experiments were approved by The Animal Care Committee of Chengdu
University of Traditional Chinese Medicine.
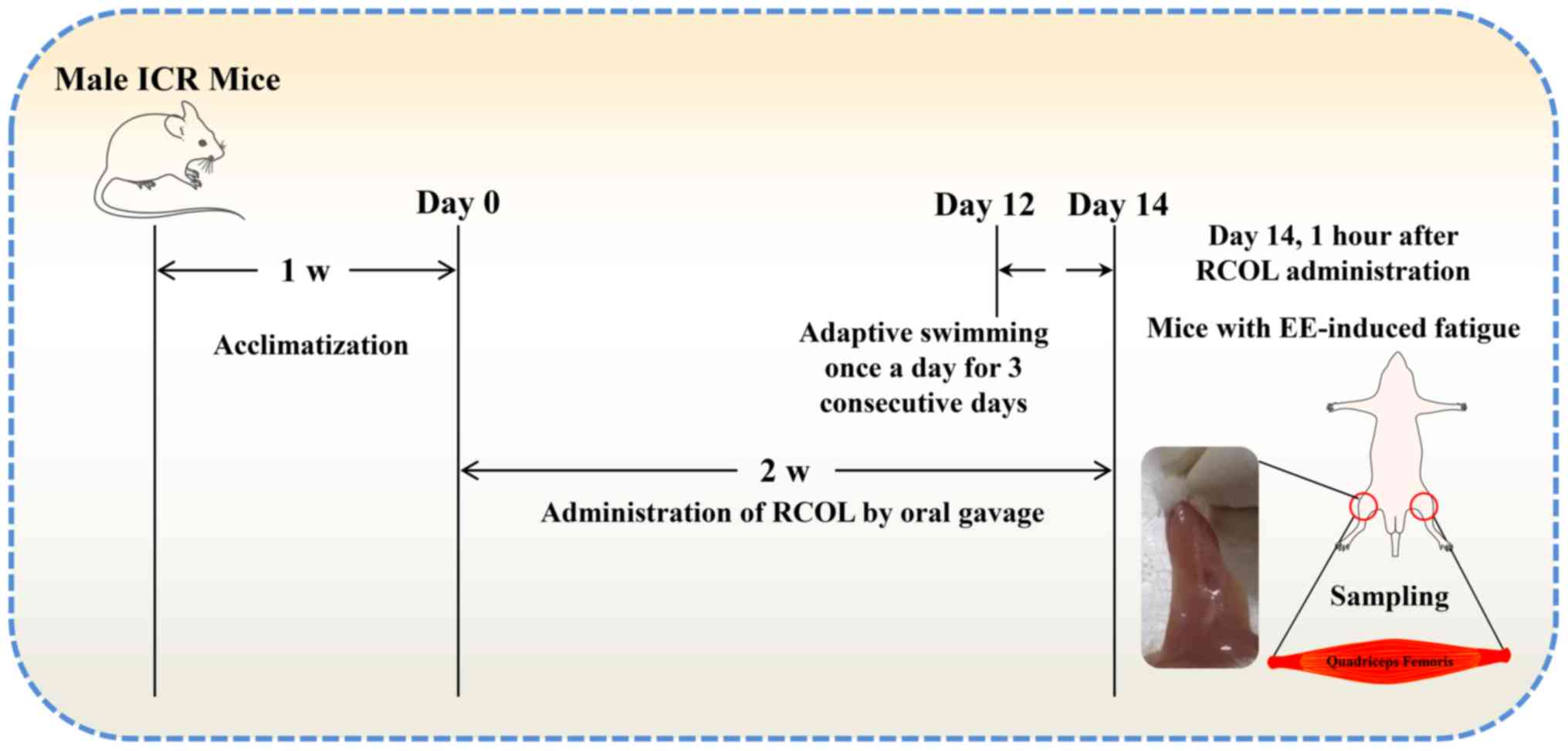
Loaded swimming test
A total of 50 mice were randomly divided into five
experimental groups of ten mice each: i) Control group; ii)
Exhaustive exercise group (EE); iii) Low-Dose group (RCOL-L; 1.02
ml/kg/day); iv) Medium-Dose group (RCOL-M; 3.03 ml/kg/day); and v)
High-Dose group (RCOL-H; 6.06 ml/kg/day). Mice in the three RCOL
groups were given ROCL by oral gavage once daily for two weeks. The
nutritional information for RCOL is summarized in Table I. Mice in the control and EE groups
were given a volume of sterilized water equivalent to the volume of
RCOL administered according to the body weight of the mouse.
 | Table IComposition of Rhodiola
crenulata oral liquid. |
Table I
Composition of Rhodiola
crenulata oral liquid.
| Peak no. | Compound | Retention time
(min) | Concentration
(mg/ml) |
|---|
| 1 | Gallic acid | 9.557 | 1.8690 |
| 2 | Salidroside | 23.077 | 1.2709 |
| 3 | Tyrosol | 27.618 | 0.5359 |
| 4 | Catechin | 31.544 | 0.0109 |
| 5 | Caffeic acid | 36.232 | 0.0989 |
| 6 | p-coumaric
acid | 46.943 | 0.4086 |
All mice except for those in the control group first
underwent free swimming training without any loads in a single
20-min training session each day for 3 days, after which they were
challenged with a loaded swimming test to establish EE-induced
damage in the quadriceps, using a procedure previously described
(30). Briefly, the swimming test
was applied 1 h after the final dosing, with a lead weight ~5% of
the mouse body weight attached at the base of the tails. All mice
were challenged individually under the same conditions (water
temperature, 25±1˚C; depth, 30 cm), where failure to come to the
water surface for breathing within 10 sec was determined as a sign
of exercise fatigue. At the end of the swimming trial, the mice
were sacrificed by cervical dislocation. Subsequently, ~0.6 ml
blood samples from the eyeball were collected, liver and quadriceps
tissues were collected for further experiments. The workflow of the
present experimental protocol is shown in Fig. 1.
Measurement of anti-fatigue biomarkers
and glycogen synthesis ability in the mouse serum and quadriceps
femoris
Serum samples from mice were obtained by
centrifugation at 12,000 x g for 10 min at 4˚C. Levels of
biomarkers associated with fatigue were measured in the serum (CK)
or in the quadriceps femoris tissues of the hind legs (LA, LDH,
MDA, SOD, CAT and T-AOC). Glycogen synthesis ability was assessed
by measuring glycogen levels in the liver and quadriceps femoris
tissues. In addition, CS, SDH and
Na+-K+-ATPase activities were also measured
in quadriceps femoris tissues. All analytes were measured according
to the manufacturer's protocols.
Histopathological estimations of
quadriceps femoris
A portion of the quadriceps tissue (10x5x5 mm) was
carefully dissected from mice in each of the treatment groups and
fixed in 4% formaldehyde overnight at 25˚C. Paraffin blocks were
then prepared after dehydration (75% ethanol for 6 h, 85% ethanol
for 10 h, 95% ethanol for 4 h, anhydrous ethanol I and II for 2 h),
clearing (dimethylbenzene xylene I and II for 20 min and 15 min,
respectively) and wax impregnation for 3 h. The blocks were cut
then into 5-µm sections using a rotary microtome (RM2235; Leica
Microsystems GmbH), deparaffinized in xylene, rehydrated with
deionized water at 25˚C for 20 min and stained with hematoxylin for
30 min and eosin for 5 min at 25˚C. Following the staining
protocol, the quadriceps sections from five mice in each group were
digitally captured using a bright field light microscope
(magnification x200; CX22; Olympus Corporation) equipped with an
image capture system (DM1000; Leica Microsystems GmbH). The number
of muscle fibers was counted in each sample, where the
cross-sectional area of 20 muscle fibers in three random fields was
measured using Image-Pro Plus software version 6.0 (Media
Cybernetics, Inc.).
Ultrastructural analysis of
mitochondria in the quadriceps femoris by transmission electron
microscopy (TEM)
The quadriceps femoris was dissected further into
1-mm3 samples and fixed using a 2.5% Gluta transmission
electron microscope specimen fixative (pH 7.2-7.4) for 4 h at 4˚C,
following which they were rinsed three times for 15 min each with
0.1 M phosphate solution. The samples were then fixed at 25˚C with
1% osmium acid for 3 h and then rinsed again three times for 15 min
each with 0.1 M phosphate solution. The samples were then
dehydrated at 4˚C, using the following dehydration protocol: i) 50%
ethanol for 15-20 min; ii) 70% ethanol for 15-20 min; iii) 90%
ethanol for 15-20 min; iv) 90% ethanol and 90% acetone (1:1) for
15-20 min; v) 90% acetone for 15-20 min; and vi) 100% acetone at
room temperature for 15-20 min three times. Embedding and
solidification were then performed using SPI-Pon 812 embedding kit
(Structure Probe, Inc.) overnight at 37˚C, then for 12 h at 45˚C
and finally for 24 h at 60˚C. Ultrathin 50-60-nm sections were then
cut using an ultramicrotome (EM UC7; Leica Microsystems GmbH),
collected onto copper grids and stained for 5 min with 3% uranyl
acetate and lead citrate at 25˚C. The grids were then washed with
distilled water and viewed under a transmission electron microscope
(magnification x10,000; JEM1230; JEOL) for the observation of the
muscle tissue ultrastructure. A total 10 muscle cells in random
non-overlapping fields per grid were considered for counting
purposes and five grids were assessed in each group using Image-Pro
Plus software version 6.0 (Media Cybernetics, Inc.).
Western blot analysis
The quadriceps femoris of mice were dissected and
immediately separated on ice. Tissue homogenate was obtained from
the quadriceps femoris tissues using the FastPrep-24™ sample
preparation system (MP Biomedicals, LLC) as described previously
(7,31-33).
Total protein was isolated using a whole cell lysis assay (cat. no.
KGP2100; Ken Gen Biotech. Co., Ltd.). A BCA protein assay kit (cat.
no. AR0146; Boster Biological Technology) was used to evaluate the
protein concentration of each sample. Equivalent amounts (40
µg/lane) of protein samples were separated by SDS-PAGE on
10% gels, following which they were transferred onto 0.45-µm PVDF
membranes (cat no. IPVH00010, EMD Millipore). The membranes were
then blocked with 5% non-fat milk for 2 h at 25˚C and incubated
with primary antibodies against CS, PINK1, Parkin, LC3I/II,
SQSTM-1/p62, ubiquitin and GAPDH overnight at 4˚C (all at 1:1,000
dilution). The membranes were then incubated with horseradish
peroxidase-conjugated goat anti-rabbit IgG secondary antibody
(1:1,000; cat. no. GB23303, Wuhan Servicebio Technology Co., Ltd.)
for 2 h at 25˚C and washed with PBS containing 0.1% Tween-20 (cat.
no. T8220; Beijing Solarbio Science & Technology Co., Ltd.)
three times for 10 min each. Finally, Ultrasignal ECL
electrochemiluminescent substrate (cat. no. 4AW011-100; Beijing 4A
Biotech Co., Ltd.) was used for the visualization of the protein
bands and images were captured using ChampChemi 610 Plus (Beijing
Sage Creation Science Co., Ltd.). Density values of bands were
calculated using Image-Pro Plus software version 6.0 (Media
Cybernetics, Inc.) and normalized to GAPDH.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from quadriceps femoris
tissue using TRIzol® according to manufacturer's
protocol (Invitrogen; Thermo Fisher Scientific, Inc.). cDNA was
subsequently synthesized by reverse transcription using a
commercial Maxima SYBR Green Real-Time PCR kit (cat. no. KGA1339;
Nanjing KeyGen Biotech Co., Ltd.) according to the manufacturer's
instructions. RT-qPCR was performed in an ABI 7300 real-time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). All
reactions were set up in 20 µl volumes using SYBR™ Green PCR Master
Mix according to manufacturer's protocol (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The following thermocycling
conditions were used for the qPCR: Initial denaturation at 95˚C for
30 sec; 40 cycles comprising denaturation at 95˚C for 5 sec,
annealing at 60˚C for 30 sec and elongation at 72˚C for 30 sec; and
final extension at 72˚C for 5 min. PINK1 and Parkin mRNA expression
were normalized to those of GAPDH and presented as
2-ΔΔCq (34). The primer sequences used are listed
in Table II.
| Table IIPrimers used for reverse
transcription-quantitative PCR. |
Table II
Primers used for reverse
transcription-quantitative PCR.
| Gene | GenBank accession
no. | Primer sequence,
5'-3' |
|---|
| GAPDH | NM_017008.4 | F,
TCACTATTGGCAACGAGCGGTTC |
| | | R,
GCACTGTGTTGGCATAGAGGTCTT |
| PINK1 | NM_026880.2 | F,
CAGTGTAGAGCGTGGTGGCAAT |
| | | R,
AGGCACCGACTCAGGCATCT |
| Parkin | NM_016694.4 | F,
GCTTGACACGAGTGGACCTGAG |
| | | R,
AACTGGACCTCTGGCTGCTTCT |
Statistical analysis
Data are presented as the mean ± SD. Statistical
analysis was performed using SPSS version 21.0 (IBM Corp.). Data
were analyzed using one-way ANOVA followed by Tukey's multiple
comparison post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
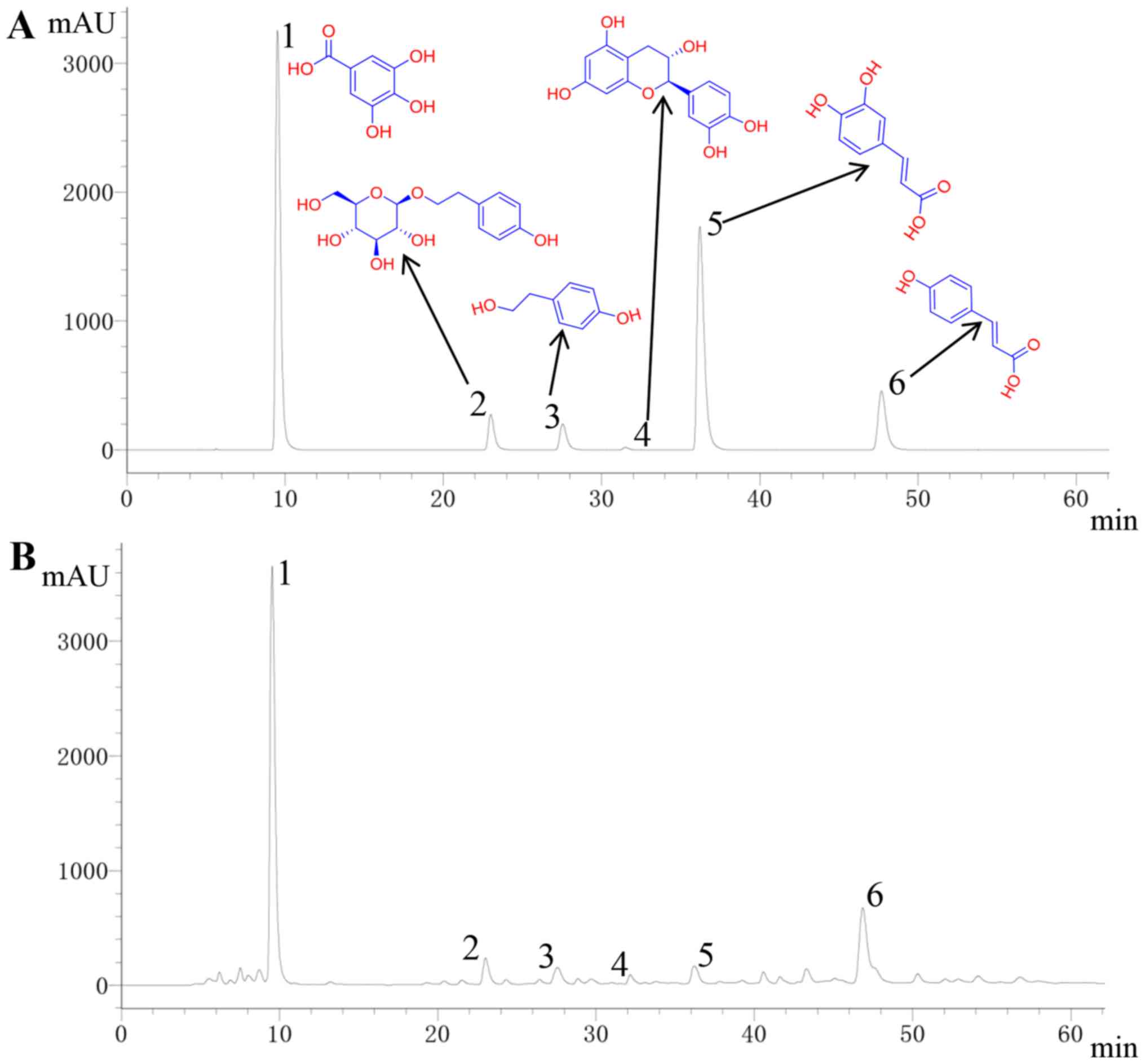
Abundance of the six key compounds
contained within RCOL
Detection and quantification of the six active
compounds of RCOL (gallic acid, salidroside, tyrosol, p-coumaric
acid, caffeic acid, and catechin) were first performed by HPLC.
Concentrations were determined using the following curve regression
equations: i) Gallic acid,
y=5x107x+942161, r=0.9992; ii)
Salidroside, y=5x106x-21828, r=1;
iii) Tyrosol, y=107x-19063, r=1;
iv) p-coumaric acid, y=7x107x-50372,
r=1; v) Caffeic acid,
y=5x107x+115424, r=0.9999, and vi)
Catechin, y=1x107x-7681.9,
r=0.9999. The relative abundance of the six compounds in
RCOL are shown in Fig. 2 and
Table I. Among them, the
concentration of gallic acid and salidroside is the highest, at
1.8690 and 1.2709 mg/ml, respectively.
Effects of RCOL administration on body
weight of mice
The weight of the mice was recorded before and after
RCOL treatment. Beyond the naturally increased body weight observed
in mice, the continuous administration of RCOL for 14 days had no
significant effects on body weight in the mice from each group by
the end of the experiment (Table
III).
| Table IIIEffect of RCOL administration on
mouse BW. |
Table III
Effect of RCOL administration on
mouse BW.
| Title | Control | EE | RCOL-L | RCOL-M | RCOL-H |
|---|
| Initial BW (g) | 27.93±1.19 | 28.27±0.92 | 28.14±1.21 | 27.77±0.86 | 27.55±0.99 |
| Final BW (g) | 35.50±2.11 | 35.85±1.22 | 35.25±1.75 | 32.24±2.11 | 31.28±1.44 |
| ∆BW (g) | 7.50±1.70 | 7.60±0.93 | 7.10±1.72 | 4.50±1.97 | 3.70±1.81 |
Effects of RCOL administration on the
levels of fatigue-related biomarkers in serum and quadriceps
femoris tissues
After 2 weeks of RCOL pre-administration, the mice
underwent 3 days of weight-free adaptive swimming training from day
12 onwards, once per day (Fig. 1).
On day 14, 1 h after RCOL administration, a weight-loaded
swimming-induced skeletal muscle fatigue protocol was initiated on
the mice, following which serum and quadriceps femoris samples were
collected immediately after the end of the trial to detect changes
in fatigue-related indicators. Compared with those in the EE group,
significantly increased activities of SOD, CAT and T-AOC were
observed in conjunction with significantly reduced LA, CK, LDH and
MDA levels in the three different RCOL dosage groups (Table IV). These results suggested that
RCOL can relieve fatigue induced by EE in mice, possibly by
modulating oxidative stress.
| Table IVEffect of RCOL administration on
anti-fatigue indicators in the serum samples and quadriceps femoris
tissue of mice. |
Table IV
Effect of RCOL administration on
anti-fatigue indicators in the serum samples and quadriceps femoris
tissue of mice.
| Sample | Parameter | Control | EE | RCOL-L | RCOL-M | RCOL-H |
|---|
| Serum | CK, U/ml | 0.77±0.20 |
2.16±0.25a |
1.79±0.08b |
1.55±0.05b |
1.27±0.13b |
| Quadriceps
femoris | LA, mmol/g
prot | 0.51±0.07 |
1.29±0.19a |
1.00±0.083b |
0.75±0.07b |
0.69±0.06b |
| Quadriceps
femoris | LDH, U/g prot | 1.26±0.11 |
1.98±0.18a |
1.76±0.08b |
1.60±0.12b |
1.55±0.12b |
| Quadriceps
femoris | MDA, nmol/mg
prot | 2.95±0.37 |
6.03±0.61a |
4.91±0.06b |
4.48±0.14b |
3.93±0.33b |
| Quadriceps
femoris | SOD, U/mg prot | 158.77±12.95 |
87.86±5.21a |
110.82±5.94b |
121.68±6.77b |
140.39±4.14b |
| Quadriceps
femoris | CAT, U/mg prot | 18.50±3.14 |
4.18±1.64a | 6.68±2.25 |
7.71±1.90c |
14.44±3.12b |
| Quadriceps
femoris | T-AOC, U/mg
prot | 0.67±0.06 |
0.28±0.03a |
0.37±0.03b |
0.46±0.04b |
0.55±0.02b |
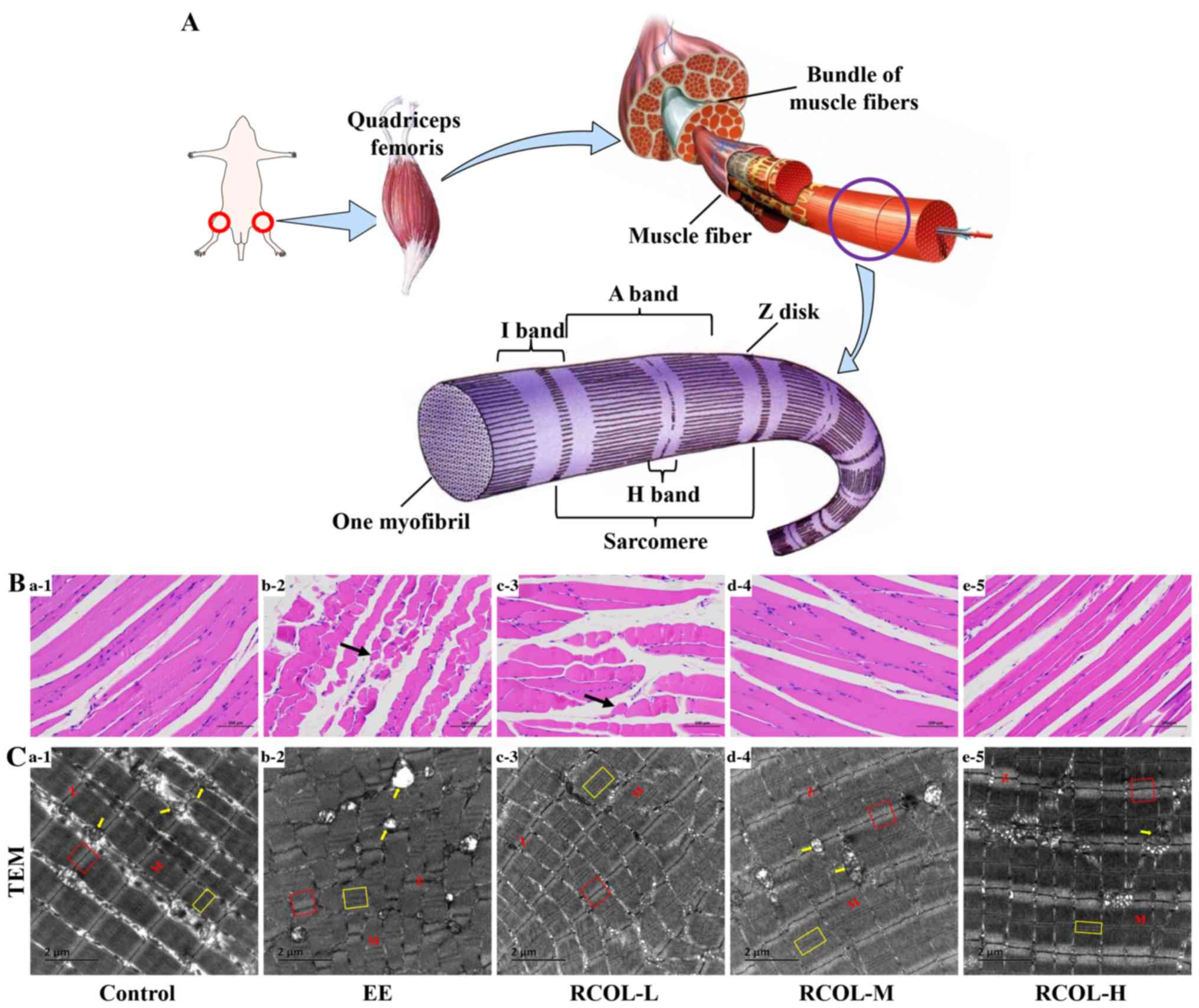
Effect of RCOL administration on the
histopathology and ultrastructure of quadriceps femoris in
mice
A schematic diagram of the quadriceps femoris muscle
structure in mice is presented in Fig.
3A. Morphological analysis of skeletal muscle suggested that
there was slight edema in the local muscle interstroma, looser
connective tissue arrangement, irregular arrangement and shape of
local muscle fibers. Meanwhile, a small amount of muscle fiber area
was visibly reduced, and the spacing was slightly widened in the
control group. By contrast, RCOL pre-administration appeared to
alleviate the above pathological damages (Fig. 3B). Ultrastructural analysis of the
mouse quadriceps femoris using TEM further confirmed the protective
effect of RCOL pre-administration on EE-induced injury (Fig. 3C). Indeed, compared with the control
group, the myofibrils of the quadriceps femoris in the EE group
were found to be significantly damaged, disorganized, partially
disintegrated and even dissolved, where the A- and H-bands appeared
thinner or were missing. The partly or fully disintegrated
mitochondrial cristae, mitochondrial swelling or even vacuoles were
also visible. In addition, the skeletal muscles of the EE group
showed abnormal ultrastructural changes, including disappearance of
the I-band, a blurred, disordered arrangement of the Z-disk and
serrated corrugation (Fig. 3C1-2).
However, other than the changes to the mitochondrial microstructure
aforementioned, results from this experiment did not suggest any
changes in autophagosomes. Collectively, based on the morphological
and ultrastructural characteristics of the skeletal muscle, RCOL
pre-administration can protect the skeletal muscle against injury
induced by EE.
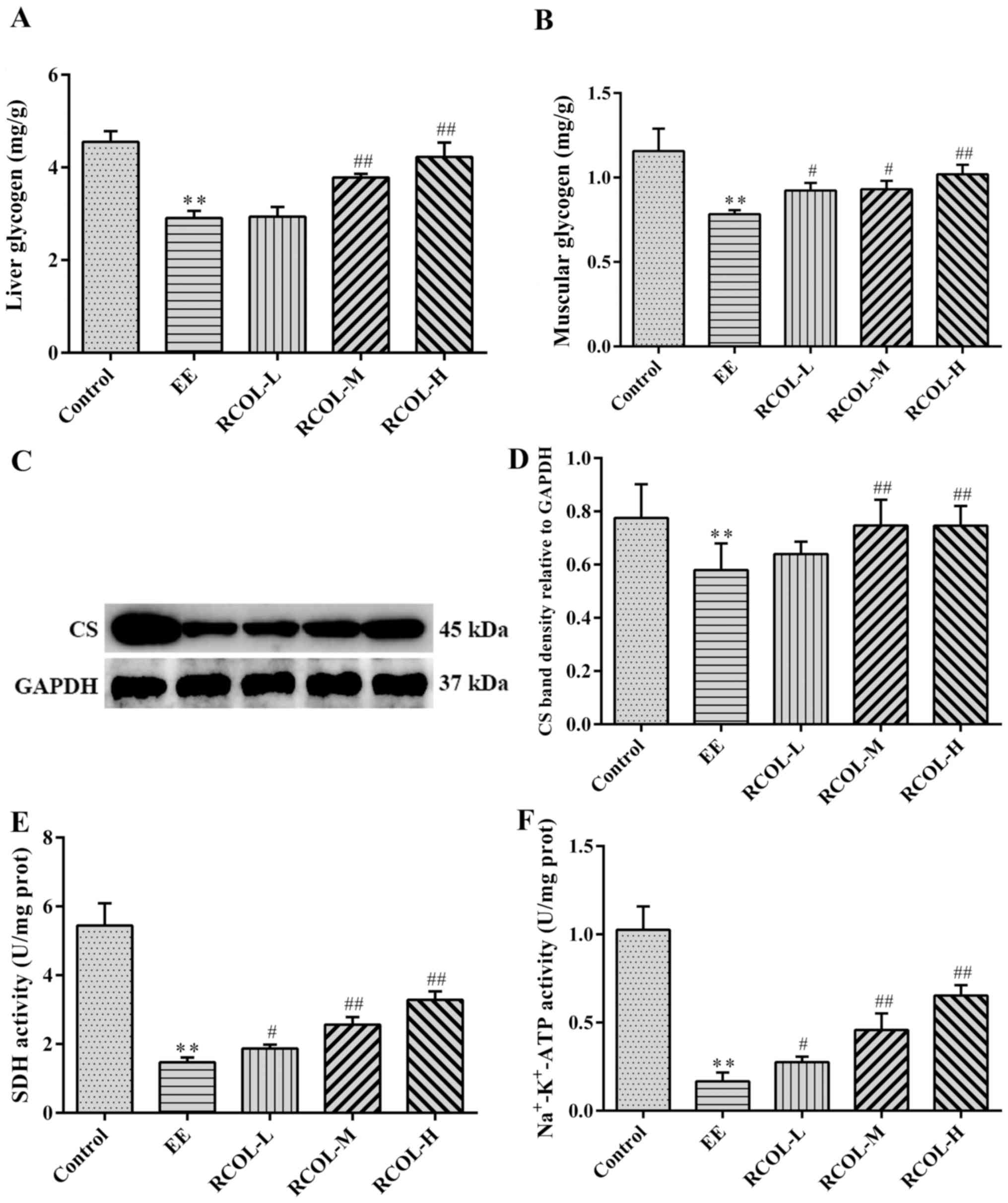
Effect of RCOL administration on
energy production in mice
To evaluate the effect of RCOL on energy production
in mice, the contents of liver and muscle glycogen, in addition to
the activities of SDH and Na+-K+-ATPase in
the quadriceps femoris were measured by biochemical analysis.
Furthermore, CS expression in quadriceps femoris tissue was
measured by western blotting. Compared with those in the EE group,
the mice in the three RCOL groups exhibited significantly higher
liver and muscle glycogen content (Fig.
4A and B), significantly higher
CS expression (Fig. 4C and D) and significantly higher SDH (Fig. 4E), and
Na+-K+-ATPase activity (Fig. 4F). Therefore, enhanced ATP production
may partially underlie the anti-fatigue effects of RCOL observed in
mice following EE.
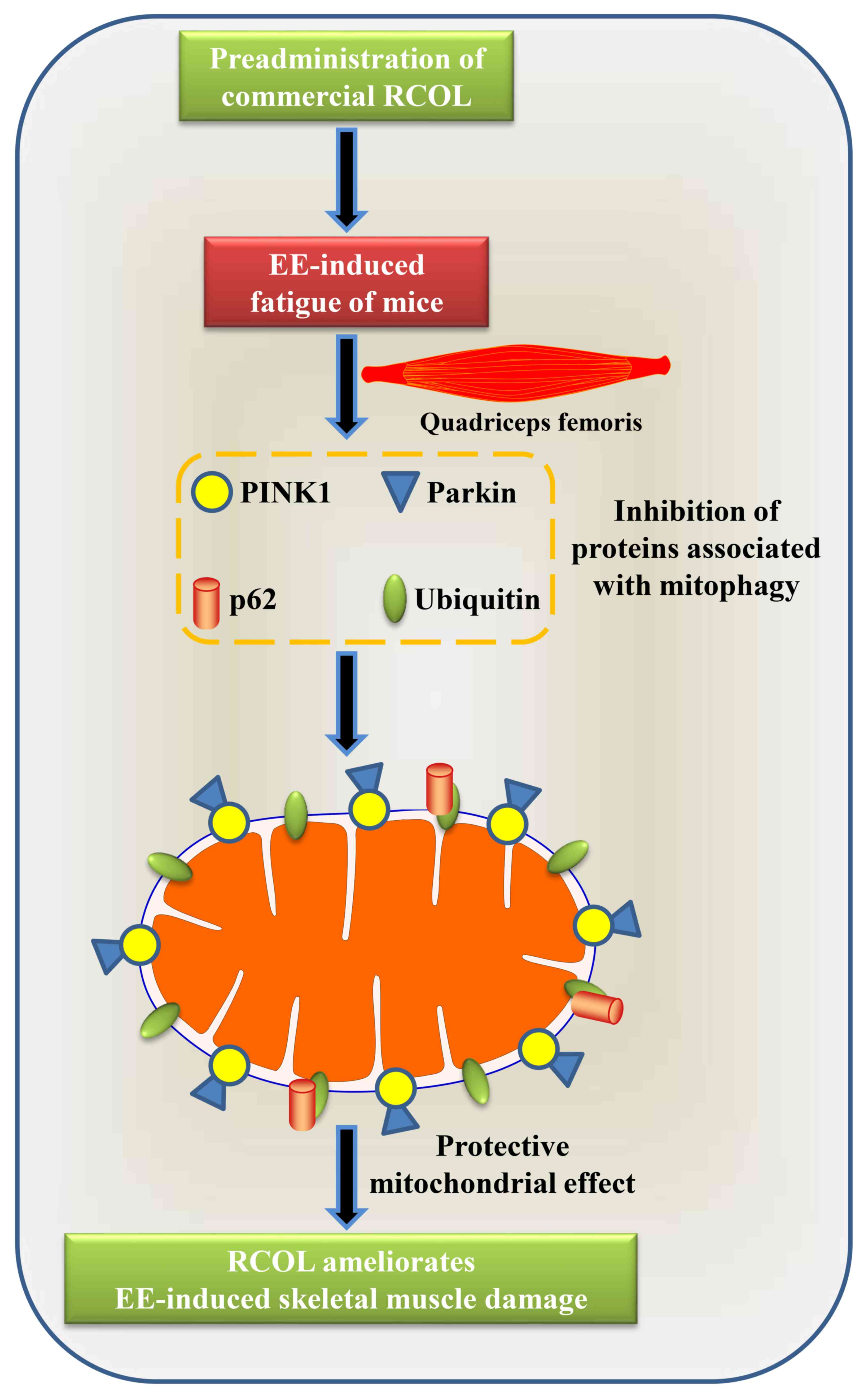
Effects of RCOL administration on the
expression of PINK1/Parkin signaling pathway components
To determine the effects of RCOL on
PINK1/Parkin-mediated mitophagy, western blot analysis was
performed to measure PINK1, Parkin, LC3-Ⅰ/LC3-Ⅱ, p62 and ubiquitin
protein expression of mice quadriceps tissue (Fig. 5A-F). To support this, mRNA expression
levels of Parkin and PINK1 were measured by RT-qPCR (Fig. 5G and H). Compared with those in the control
group, mice in the EE group displayed significantly higher
expression levels of PINK1, Parkin, LC3-Ⅰ/LC3-Ⅱ, p62 and ubiquitin,
proteins associated with mitophagy. RCOL pre-administration was
found to significantly reverse this effect on the expression of
proteins aforementioned. In addition, RT-qPCR results further
confirmed the reduced mRNA expression levels of PINK1 and Parkin in
the medium and high dose RCOL groups compared with those in the EE
group. Altogether, these results suggest that RCOL may protect mice
from EE-induced fatigue by inhibiting the expression of
mitophagy-related proteins.
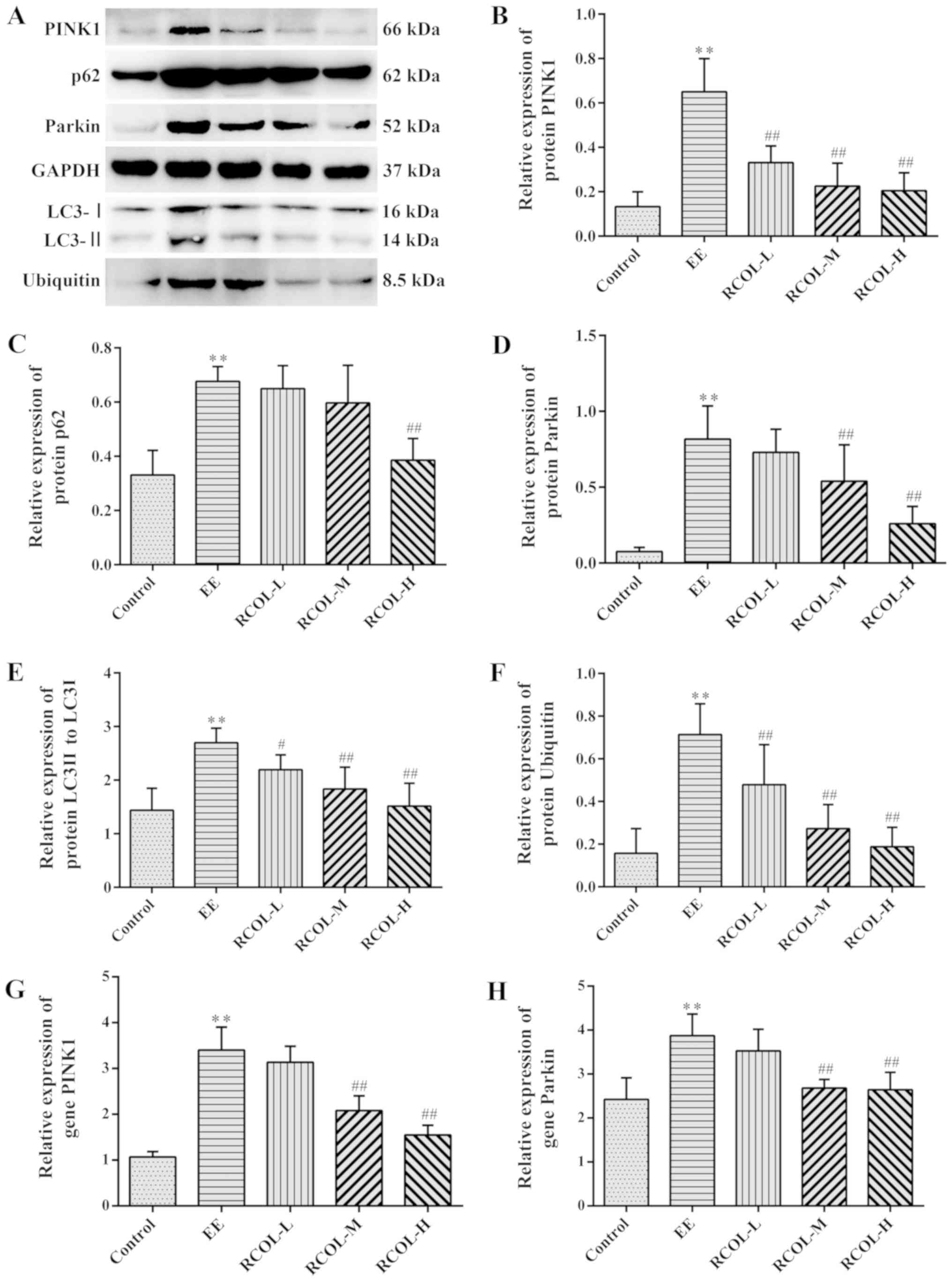 | Figure 5Effects of RCOL administration on the
protein and mRNA expression of key PINK1/Parkin signaling pathway
components. (A) Representative western blotting images showing the
protein expression of PINK1, Parkin, LC3-II/LC3-I, p62 and
ubiquitin in mice quadriceps femoris tissues. Semi-quantitative
densitometric analysis of (B) PINK1, (C) p62, (D) Parkin, (E) LC3I
and (F) ubiquitin. Relative mRNA expression of (G) PINK1 and (H)
Parkin, two key proteins associated with mitophagy. Data are
presented as the mean ± SD. n=6. **P<0.01 vs.
Control; #P<0.05 and ##P<0.01 vs. EE.
RCOL, Rhodiola crenulata oral liquid; EE, exhaustive
exercise; L, low dose; M, medium dose; H, high dose; PINK1,
PTEN-induced kinase 1; LC3, microtubule associated protein 1 light
chain 3. |
Discussion
RCOL is a tonic containing an abundance of
pharmacologically active ingredients and nutrients (22). In the present study, RCOL was found
to improve EE-induced skeletal muscle fatigue in mice, increased
antioxidant capacity and glycogen synthesis. The anti-fatigue
effect of RCOL may be associated with the inhibition of mitophagy
through the regulation of the PINK1/Parkin signaling pathway.
Previous studies have suggested that EE can induce
damage through the production of reactive oxygen radicals (35-37).
The imbalanced reduction-oxidation states in turn lead to muscle
weakness and fatigue (35). LA is a
metabolite found in skeletal muscle, where the rapid accumulation
of LA can cause damage. CK serves an important role in energy
transfer, muscle contraction and ATP generation. CK levels have
been previously reported to positively correlate with the
expression of tissue LDH (38,39). EE
has been found to disrupt the redox state, increase MDA expression
and reduce the contents of SOD and CAT, aggravating oxidative
damage to the skeletal muscle (40,41).
Therefore, LA, CK, LDH, MDA, SOD, CAT and T-AOC were chosen as
biomarkers to evaluate the degree of fatigue and oxidation in
response to RCOL in skeletal muscle after EE-induced fatigue. In
the present study, RCOL administration was observed to inhibit the
levels of LA, CK, LDH and MDA, whilst increasing expression of SOD,
CAT and T-AOC in the quadriceps femoris tissue, without affecting
body weight. These findings suggested that the anti-fatigue
properties of RCOL were at least partly associated with its
antioxidant effects.
In the present study, at the cellular and
microstructural levels, morphological and ultrastructural analysis
suggested that EE can disrupt the normal arrangements of skeletal
muscle cells, with incomplete and poorly defined sarcolemma.
Irregular arrangements of muscle fibers can deteriorate further, as
demonstrated by the twisted, broken and dissolved muscle fibers, in
addition to myostromal edema (11).
It is also worth noting that unaccustomed exercise, in a manner
that is dependent on both and intensity, can induce the acute
microdisruption of the myofibrils and cytoskeletal structures
(42). In addition, broken
myofilaments, dilated sarcoplasmic reticula, disruptive
mitochondrial structures, swollen and vacuolated mitochondria were
previously observed in EE-induced skeletal muscle damage (12). The present study suggested that RCOL
administration mitigated EE-induced skeletal muscle damage,
preserved mitochondrial morphology and function, as evidenced by
the restoration of the number and area of healthy muscle fibers and
the amelioration of ultrastructural damage. However, the results of
transmission electron microscopy of skeletal muscle mitochondria in
the present study did not provide direct evidence of mitophagy.
Future studies should aim to couple investigations of
representative markers associated with autophagy, including
lysosomes and autophagy-lysosome complexes, with changes to the
skeletal muscle fiber and mitochondrial ultrastructure.
CS and SDH are two important catalytic enzymes
involved in the tricarboxylic acid cycle. CS and SDH activities are
higher in skeletal muscles with more mitochondria, thereby
providing more ATP (43). Previous
in vitro and in vivo studies suggested that, under
normal conditions, Rhodiola crenulata extract (RCE)
inhibited hepatic glycogen synthesis, reducing blood glucose levels
through activation of the 5'AMP-activated protein kinase (AMPK)
signaling pathway (44). By
contrast, another previous study reported contrasting results,
where RCE treatment increased glycogen synthesis through the AMPK
pathway in HepG2 cells incubated under high-glucose conditions
(45). Therefore, to determine the
effects of RCOL on energy production in mice with EE-induced
fatigue, glycogen content was evaluated in the liver and quadriceps
femoris tissues. RCOL treatment was found to increase glycogen
levels in both tissues, possibly owing to enhanced gluconeogenesis
process. The activities of CS, SDH and
Na+-K+-ATPase in quadriceps femoris were also
assessed. Higher CS, SDH and Na+-K+-ATPase
activities were confirmed in RCOL-administered mice compared with
those in the EE group. Collectively, these findings suggested that
RCOL pre-administration increased glycogen content in the liver and
muscle tissues, thereby augmenting ATP production. The motor
ability and behavioral performance of mice were therefore improved,
evidenced by enhanced energy production and improved
fatigue-related indicators. Therefore, it may be concluded that
RCOL can increase the glycogen content in response to EE. Further
in vivo and in vitro studies are required to verify
the regulatory effects and mechanisms of RCOL on glycogen synthesis
under different conditions, such as diabetes.
Previous studies suggested that autophagy and/or
mitophagy were involved in the physiological and pathological
homeostasis of mitochondrial quality control in skeletal muscle
cell (13,46,47).
Parkin, encoded by the Park2 gene in animals, is a
cytoplasmic E3 ubiquitin ligase with broad physiological roles. It
is highly expressed in skeletal muscle and other tissues, including
the substantia nigra of brain, heart, liver and testis (46). PINK1 belongs to the Ser/Thr protein
kinase family which contains a single protein kinase domain and is
highly expressed in the mitochondrial outer membrane (MOM) of
skeletal muscle. Reduced mitochondrial inner membrane
electrochemical gradients, which are initiated by mitochondrial
uncouplers or toxins, are required for ATP production.
Subsequently, Parkin translocates from the cytosol into the
mitochondria under conditions of impaired and imbalanced
mitochondrial membrane potentials (13). Accumulation of PINK1 on the MOM, then
phosphorylates Parkin on serine 65 to activate the ligase activity
of Parkin, resulting in increased mitophagy (48). Additionally, Parkin conjugates
ubiquitin chains on proteins located on the MOM, causing signal
amplification and recruitment of the autophagosome to initiate
mitophagy (13). Therefore, changes
in the concentration of PINK1 protein can be used to measure the
health status of mitochondria (49,50).
PINK1 and Parkin are considered to be the key mediators of
mitophagy in removing damaged mitochondria from the cell (51).
SQSTM1/p62 is a ubiquitously expressed scaffold
protein that is considered to be a marker of autophagosomes, which
serve an essential role in the regulation of mitophagy (13,52). p62
maintains its own intracellular levels by regulating the mitophagy
process by directly binding to LC3, forming a LC3/p62 complex
(53). Loss of p62 expression
results in the inactivation of autophagy and the accumulation of
cytoplasmic protein inclusions, thereby contributing to the
progression of diseases associated with autophagy deficiency,
including cancer and Parkinson's disease (54). In addition, PINK1/parkin-mediated
mitophagy has also been found to be associated with ubiquitin and
p62(55). Previous studies suggested
that salidroside suppressed bladder cancer cell growth by
inhibiting the mechanistic target of the rapamycin (mTOR) pathway
and translation initiation, whilst inducing autophagy in tumor
cells (56). However, other previous
studies have also suggested that salidroside can increase hypoxia
and hydrogen peroxide-induced autophagy in pulmonary arterial
smooth muscle cells and human umbilical vein endothelial cells
through downregulation of mTOR signaling (57,58).
Salidroside has also been documented to inhibit autophagy in
primary cortical neurons of neonatal Sprague-Dawley rats exposed to
glutamate (59) and suppress the
mitophagy process in the hippocampal CA3 neurons after chronic
hypobaric hypoxia injury (60).
Additionally, salidroside alleviated hepatic autophagy in mouse
models of hepatic ischemia-reperfusion injury and hepatic fibrosis
(61,62). In the present study, western blotting
results indicated that RCOL pre-administration inhibited the
expression of PINK1, Parkin, LC3, p62 and ubiquitin. Further gene
expression analysis conducted by RT-qPCR also suggested that RCOL
could downregulate the expression PINK1 and Parkin mRNA. To the
best of the authors' knowledge, these results provided the first
experimental evidence that RCOL could suppress the expression
mitophagy-related proteins in the skeletal muscle following EE
induction, including those in the PINK1/Parkin signaling pathway.
The proposed molecular mechanism underlying the effects of RCOL
pre-administration on alleviating fatigue in mice is presented in
Fig. 6.
In conclusion, the present study demonstrated that
RCOL pre-administration exerted anti-fatigue effects and improved
exercise performance in mice through the regulation of
mitophagy-related markers in skeletal muscle including those of the
PINK1/Parkin signaling pathway. However, the precise molecular
mechanisms underlying the anti-fatigue properties of RCOL require
further investigation.
Acknowledgements
The authors would like to thank Mr. Jiayi Sun,
research assistant at The Innovative Institute of Chinese Medicine
and Pharmacy, Chengdu University of Traditional Chinese Medicine,
Chengdu, China, for providing technical guidance on hematoxylin and
eosin staining, as well as RT-qPCR assays during the present
study.
Funding
The present study was supported by The National Key
R&D Program of China (grant no. 2017YFC1703904), The Major
Science and Technology Project of Sichuan Science and Technology
Department (grant no. 19SYXHZ0095), The National Natural Science
Foundation of China (grant nos. 81773974 and 81203000), The Science
& Technology Department of Sichuan Province (grant no.
2018JY0467), Sichuan Science and Technology Program (grant no.
2019YJ0480), and The First-Class Interdisciplinary Project of
Chengdu University of Traditional Chinese Medicine (grant no.
CZYJC-1903).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XM, JZ and Y Hu provided financial support for the
research, designed the study, supervised the research group,
revised drafts and gave approval for the final version to be
published. Y Hou, XW, XA, HW, YT, and XC performed the experiments.
YZ, Y Hu and XL analyzed and interpreted the data. YH wrote the
manuscript. All authors approved the final version of the
manuscript.
Ethics approval and consent to
participate
The present study was approved by The Animal
Research Ethics Committee of Chengdu University of Traditional
Chinese Medicine. All experimental protocols were performed in
accordance with the Animal Care Guidelines, conforming to the
Health Guide for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
O'Sullivan K, O'Sullivan PB and Gabbett
TJ: Pain and fatigue in sport: Are they so different? Br J Sports
Med. 52:555–556. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kaufman JL, Mina R, Jakubowiak AJ,
Zimmerman TL, Wolf JJ, Lewis C, Gleason C, Sharp C, Martin T,
Heffner LT, et al: Combining carfilzomib and panobinostat to treat
relapsed/refractory multiple myeloma: Results of a multiple myeloma
research consortium Phase I Study. Blood Cancer J.
9(3)2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Witlox L, Hiensch AE, Velthuis MJ, Steins
Bisschop CN, Los M, Erdkamp FL, Bloemendal HJ, Verhaar M, Ten
Bokkel Huinink D, van der Wall E, et al: Four-year effects of
exercise on fatigue and physical activity in patients with cancer.
BMC Med. 16(86)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Fuller JT, Hartland MC, Maloney LT and
Davison K: Therapeutic effects of aerobic and resistance exercises
for cancer survivors: A systematic review of meta-analyses of
clinical trials. Br J Sports Med. 52(1311)2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Cramp F and Byron-Daniel J: Exercise for
the management of cancer-related fatigue in adults. Cochrane
Database Syst Rev. 11(CD006145)2012.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Pilutti LA, Greenlee TA, Motl RW, Nickrent
MS and Petruzzello SJ: Effects of exercise training on fatigue in
multiple sclerosis: A meta-analysis. Psychosom Med. 75:575–580.
2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Fu T, Xu Z, Liu L, Guo Q, Wu H, Liang X,
Zhou D, Xiao L, Liu L, Liu Y, et al: Mitophagy directs
muscle-adipose crosstalk to alleviate dietary obesity. Cell Rep.
23:1357–1372. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Peker N, Donipadi V, Sharma M, McFarlane C
and Kambadur R: Loss of parkin impairs mitochondrial function and
leads to muscle atrophy. Am J Physiol Cell Physiol. 315:C164–C185.
2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Brooks GA: Lactate as a fulcrum of
metabolism. Redox Biol. 35(101454)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Barbalho SM, Flato UAP, Tofano RJ, Goulart
RA, Guiguer EL, Detregiachi CRP, Buchaim DV, Araújo AC, Buchaim RL,
Reina FTR, et al: Physical exercise and myokines: Relationships
with sarcopenia and cardiovascular complications. Int J Mol Sci.
21(3607)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lambert M, Bastide B and
Cieniewski-Bernard C: Involvement of O-GlcNAcylation in the
skeletal muscle physiology and physiopathology: Focus on muscle
metabolism. Front Endocrinol. 9(578)2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Arribat Y, Broskey NT, Greggio C, Boutant
M, Conde Alonso S, Kulkarni SS, Lagarrigue S, Carnero EA, Besson C,
Cantó C and Amati F: Distinct patterns of skeletal muscle
mitochondria fusion, fission and mitophagy upon duration of
exercise training. Acta Physiol (Oxf). 225(e13179)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wang X, Liu Z, Fan F, Hou Y, Yang H, Meng
X, Zhang Y and Ren F: Microfluidic chip and its application in
autophagy detection. TrAC Trends Analyt Chem. 117:300–315.
2019.
|
|
14
|
Chen HI, Ou HC, Chen CY, Yu SH, Cheng SM,
Wu XB and Lee SD: Rhodiola crenulata neuroprotective effect
of in D-galactose-induced aging model. Am J Chin Med. 48:373–390.
2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang X, Hou Y, Li Q, Li X, Wang W, Ai X,
Kuang T, Chen X, Zhang Y, Zhang J, et al: Rhodiola crenulata
attenuates apoptosis and mitochondrial energy metabolism disorder
in rats with hypobaric hypoxia-induced brain injury by regulating
the HIF-1α/microRNA 210/ISCU1/2(COX10) signaling pathway. J
Ethnopharmacol. 241(111801)2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Booker A, Zhai L, Gkouva C, Li S and
Heinrich M: From traditional resource to global commodities:-A
comparison of rhodiola species using NMR spectroscopy-metabolomics
and HPTLC. Front Pharmacol. 7(254)2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Sellami M, Slimeni O, Pokrywka A, Kuvačić
G, D Hayes L, Milic M and Padulo J: Herbal medicine for sports: A
review. J Int Soc Sports Nutr. 15(14)2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhang YZ, Zhu RW, Zhong DL and Zhang JQ:
Nunataks or massif de refuge? A phylogeographic study of
Rhodiola crenulata (Crassulaceae) on the world's
highest sky islands. BMC Evol Biol. 18(154)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Torrens-Spence MP, Pluskal T, Li FS,
Carballo V and Weng JK: Complete pathway elucidation and
heterologous reconstitution of rhodiola salidroside biosynthesis.
Mol Plant. 11:205–217. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Chang PK, Yen IC, Tsai WC, Chang TC and
Lee SY: Protective effects of extract on hypoxia-induced
endothelial damage via regulation of AMPK and ERK pathways. Int J
Mol Sci. 19(2286)2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhang Y and Zhao Q: Salidroside attenuates
interleukin-1β-induced inflammation in human osteoarthritis
chondrocytes. J Cell Biochem. 30(1002)2018.
|
|
22
|
Liu Y, Tang H, Liu X, Chen H, Feng N,
Zhang J, Wang C, Qiu M, Yang J and Zhou X: Frontline science:
Reprogramming COX-2, 5-LOX, and CYP4A-mediated arachidonic acid
metabolism in macrophages by salidroside alleviates gouty
arthritis. J Leukoc Biol. 105:11–24. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wang JM, Qu ZQ, Wu JL, Chung P and Zeng
YS: Mitochondrial protective and anti-apoptotic effects of extract
on hippocampal neurons in a rat model of alzheimer's disease.
Neural Regen Res. 12:2025–2034. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Li FJ, Liu Y, Yuan Y, Yang B, Liu ZM and
Huang LQ: Molecular interaction studies of acetylcholinesterase
with potential acetylcholinesterase inhibitors from the root of
Rhodiola crenulata using molecular docking and isothermal
titration calorimetry methods. Int J Biol Macromol. 104:527–532.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wu H, Chen W, Zhao F, Zhou Q, Reinach PS,
Deng L, Ma L, Luo S, Srinivasalu N, Pan M, et al: Scleral hypoxia
is a target for myopia control. Proc Natl Acad Sci USA.
115:E7091–E7100. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lin KT, Chang TC, Lai FY, Lin CS, Chao HL
and Lee SY: Rhodiola crenulata attenuates γ-ray induced
cellular injury via modulation of oxidative stress in human skin
cells. Am J Chin Med. 46:175–190. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Bassa LM, Jacobs C, Gregory K, Henchey E,
Ser-Dolansky J and Schneider SS: Rhodiola crenulata induces
an early estrogenic response and reduces proliferation and
tumorsphere formation over time in MCF7 breast cancer cells.
Phytomedicine. 23:87–94. 2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Couto M: Laboratory guidelines for animal
care. Methods Mol Biol. 770:579–599. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhao CY, Jia GF, He ZJ, Luo QF, Fan G,
Zhang Y and Zhao WJ: Effect of fertilization combinations of
nitrogen, phosphorus, and potassium on four phenolic compounds of
cultivated Rhodiola crenulata. Zhongguo Zhong Yao Za Zhi.
43:1812–1817. 2018.(In Chinese). PubMed/NCBI View Article : Google Scholar
|
|
30
|
Hu Y, Zhang J, Zhang Y, Wang P and Meng X:
Metabonomic study on fatigue elimination of exhaustive exercise
mouse by rhodiola based on UFLC-Q-TOF. World Science and
Technology/Modernization of Traditional Chinese Medicine and
Materia Medica. 17:2209–2214. 2015.(In Chinese).
|
|
31
|
Hou Y, Wang X, Chen X, Zhang J, Ai X,
Liang Y, Yu Y, Zhang Y, Meng X, Kuang T and Hu Y: Establishment and
evaluation of a simulated high-altitude hypoxic brain injury model
in SD rats. Mol Med Rep. 19:2758–2766. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Hou Y, Qieni X, Li N, Bai J, Li R, Gongbao
D, Liang Y, Fan F, Wencheng D, Wang Z, et al: Longzhibu disease and
its therapeutic effects by traditional Tibetan medicine: Ershi-wei
chenxiang pills. J Ethnopharmacol. 249(112426)2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Moore TM, Zhou Z, Cohn W, Norheim F, Lin
AJ, Kalajian N, Strumwasser AR, Cory K, Whitney K, Ho T and et al:
The impact of exercise on mitochondrial dynamics and the role of
Drp1 in exercise performance and training adaptations in skeletal
muscle. Mol Metab. 21:51–67. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Liu R, Wu L, Du Q, Ren JW, Chen QH, Li D,
Mao RX, Liu XR and Li Y: Small molecule oligopeptides isolated from
walnut (Juglans regia L.) and their anti-fatigue effects in
mice. Molecules. 24(45)2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Lekhi C, Gupta PH and Singh B: Influence
of exercise on oxidant stress products in elite Indian cyclists. Br
J Sports Med. 41:691–693. 2007.PubMed/NCBI View Article : Google Scholar
|
|
37
|
McAnulty SR, McAnulty LS, Nieman DC,
Morrow JD, Shooter LA, Holmes S, Heward C and Henson DA: Effect of
alpha-tocopherol supplementation on plasma homocysteine and
oxidative stress in highly trained athletes before and after
exhaustive exercise. J Nutr Biochem. 16:530–537. 2005.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Nonaka K, Ozaki Y, Ito K, Sakita M, Une S
and Akiyama J: Endurance exercise increases the protein levels of
PGC-1α and respiratory chain complexes in mouse skeletal muscle
during atorvastatin administration. J Physiol Sci. 69:327–333.
2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Assunção Carvalho LC, de Freitas MC, Silva
AS, Biasoto AC, Martins MD, de Moura RC, Brito AK, Silva AS,
Ribeiro SL, Rossi FE and Santos MA: Nectar supplementation reduced
biomarkers of oxidative stress, muscle damage, and improved
psychological response in highly trained young handball players.
Front Physiol. 9(1508)2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Vollaard NB, Shearman JP and Cooper CE:
Exercise-induced oxidative stress: Myths, realities and
physiological relevance. Sports Med. 35:1045–1062. 2005.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Powers SK and Jackson MJ: Exercise-induced
oxidative stress: Cellular mechanisms and impact on muscle force
production. Physiol Rev. 88:1243–1276. 2008.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Minetto MA, Giannini A, McConnell R, Busso
C and Massazza G: Effects of exercise on skeletal muscles and
tendons. Curr Opin Endocrine Metabolic Res. 9:90–95. 2019.
|
|
43
|
Ratkevicius A, Carroll AM, Kilikevicius A,
Venckunas T, McDermott KT, Gray SR, Wackerhage H and Lionikas A:
H55N polymorphism as a likely cause of variation in citrate
synthase activity of mouse skeletal muscle. Physiol Genomics.
42:96–102. 2010.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Lee SY, Lai FY, Shi LS, Chou YC, Yen IC
and Chang TC: Rhodiola crenulata extract suppresses hepatic
gluconeogenesis via activation of the AMPK pathway. Phytomedicine.
22:477–486. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Lin KT, Hsu SW, Lai FY, Chang TC, Shi LS
and Lee SY: Rhodiola crenulata extract regulates hepatic
glycogen and lipid metabolism via activation of the AMPK pathway.
BMC Comp Altern Med. 16(127)2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Gouspillou G, Godin R, Piquereau J, Picard
M, Mofarrahi M, Mathew J, Purves-Smith FM, Sgarioto N, Hepple RT,
Burelle Y and Hussain SN: Protective role of Parkin in skeletal
muscle contractile and mitochondrial function. J Physiol.
596:2565–2579. 2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Chen CC, Erlich AT and Hood DA: Role of
parkin and endurance training on mitochondrial turnover in skeletal
muscle. Skelet Muscle. 8(10)2018.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Youle RJ and Narendra DP: Mechanisms of
mitophagy. Nat Rev Mol Cell Biol. 12:9–14. 2011.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Narendra DP, Jin SM, Tanaka A, Suen DF,
Gautier CA, Shen J, Cookson MR and Youle RJ: PINK1 is selectively
stabilized on impaired mitochondria to activate parkin. PLoS Biol.
8(e1000298)2010.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Matsuda N, Sato S, Shiba K, Okatsu K,
Saisho K, Gautier CA, Sou YS, Saiki S, Kawajiri S, Sato F, et al:
PINK1 stabilized by mitochondrial depolarization recruits parkin to
damaged mitochondria and activates latent Parkin for mitophagy. J
Cell Biol. 189:211–221. 2010.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Botella J, Saner N and Granata C: Guardian
of mitochondrial function: An expanded role of parkin in skeletal
muscle. J Physiol. 596:6139–6140. 2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Moscat J and Diaz-Meco MT: p62 at the
crossroads of autophagy, apoptosis, and cancer. Cell.
137:1001–1004. 2009.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Ichimura Y, Kumanomidou T, Sou YS,
Mizushima T, Ezaki J, Ueno T, Kominami E, Yamane T, Tanaka K and
Komatsu M: Structural basis for sorting mechanism of p62 in
selective autophagy. J Biol Chem. 283:22847–22857. 2008.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Komatsu M, Waguri S, Koike M, Sou YS, Ueno
T, Hara T, Mizushima N, Iwata J, Ezaki J, Murata S, et al:
Homeostatic levels of p62 control cytoplasmic inclusion body
formation in autophagy-deficient mice. Cell. 131:1149–1163.
2007.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Geisler S, Holmström KM, Skujat D, Fiesel
FC, Rothfuss OC, Kahle PJ and Springer W: PINK1/parkin-mediated
mitophagy is dependent on VDAC1 and p62/SQSTM1. Nat Cell Biol.
12:119–131. 2010.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Liu Z, Li X, Simoneau AR, Jafari M and Zi
X: Rhodiola rosea extracts and salidroside decrease the growth of
bladder cancer cell lines via inhibition of the mTOR pathway and
induction of autophagy. Mol Carcinog. 51:257–267. 2012.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Gui D, Cui Z, Zhang L, Yu C, Yao D, Xu M,
Chen M, Wu P, Li G, Wang L and Huang X: Salidroside attenuates
hypoxia-induced pulmonary arterial smooth muscle cell proliferation
and apoptosis resistance by upregulating autophagy through the
AMPK-mTOR-ULK1 pathway. BMC Pulm Med. 17(191)2017.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Zheng XT, Wu ZH, Wei Y, Dai JJ, Yu GF,
Yuan F and Ye LC: Induction of autophagy by salidroside through the
AMPK-mTOR pathway protects vascular endothelial cells from
oxidative stress-induced apoptosis. Mol Cell Biochem. 425:125–138.
2017.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Yin WY, Ye Q, Huang HJ, Xia NG, Chen YY,
Zhang Y and Qu QM: Salidroside protects cortical neurons against
glutamate-induced cytotoxicity by inhibiting autophagy. Mol Cell
Biochem. 419:53–64. 2016.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Biswal S, Barhwal KK, Das D, Dhingra R,
Dhingra N, Nag TC and Hota SK: Salidroside mediated stabilization
of Bcl-xL prevents mitophagy in CA3 hippocampal neurons
during hypoxia. Neurobiol Dis. 116:39–52. 2018.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Feng J, Chen K, Xia Y, Wu L, Li J, Li S,
Wang W, Lu X, Liu T and Guo C: Salidroside ameliorates autophagy
and activation of hepatic stellate cells in mice via NF-κB and
TGF-β1/Smad3 pathways. Drug Des Devel Ther. 12:1837–1853.
2018.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Feng J, Zhang Q, Mo W, Wu L, Li S, Li J,
Liu T, Xu S, Fan X and Guo C: Salidroside pretreatment attenuates
apoptosis and autophagy during hepatic ischemia-reperfusion injury
by inhibiting the mitogen-activated protein kinase pathway in mice.
Drug Des Devel Ther. 11:1989–2006. 2017.PubMed/NCBI View Article : Google Scholar
|