Introduction
Globally, coronary heart disease is one of the most
common, high-risk diseases and the major reason for death and
disability. Acute myocardial ischemia/reperfusion injury (IRI) is
the leading cause of coronary heart disease worldwide (1). After restoring blood supply to ischemic
myocardium, dysfunction of cell metabolism and structural damage
are further aggravated, which eventually lead to IRI (2). IRI is commonly seen after cardiac
surgery. Oxidative stress is believed to be the major mechanism
underlying the pathogenesis of IRI (3,4). With
the enhanced incidence of coronary heart diseases, myocardial
infarction and cardiac surgery, IRI is a common focus of research.
It is necessary to uncover the pathogenesis of IRI to effectively
prevent its occurrence.
Long non-coding RNA (lncRNA) is a transcript with
200 nt in length and it cannot directly encode proteins (5). Several lncRNAs have been discovered to
be dysregulated during the progression of IRI, which provides a
promising direction for uncovering the pathogenesis of IRI
(6-10).
For example, it has previously been reported that lncRNA nuclear
enriched abundant transcript 1 (NEAT1) can protect cardiomyocytes
from apoptosis via targeting miR-520a by modulating expression
levels of apoptosis-related proteins (11). Gong et al indicated that
upregulation of MALAT1 exerts a critical role in promoting
cardiomyocyte apoptosis by acting as a competing endogenous RNAs
(ceRNA) to sponge miR-144(12). In
addition, knockdown of NEAT1 can protect against IRI by inhibiting
troponin levels and cardiocytes apoptosis through regulation of the
mitogen-activated protein kinase (MAPK) signaling pathway (13). Therefore, clarification of lncRNAs
involved in the occurrence and development of IRI contributes to
improve the prognosis of affected individuals.
lncRNA small nuclear host gene 16 (SNHG16) is
an important member of the SNHG family, serving as an oncogene in
osteosarcoma (14), bladder cancer
(15), lung cancer (16), glioma (17), ovarian cancer (18) and breast cancer (19). Moreover, a previous study showed that
CTCFI-induced upregulation of SNHG16 accelerated
cardiac hypertrophy by targeting the miR-182-5p/IGF1 axis
(20). Furthermore, SNHG16
was found to promote proliferation and inflammatory response of
macrophages through the miR-17-5p/NF-κB signaling pathway in
patients with atherosclerosis (21).
However, it is unclear whether SNHG16 is involved in the
proliferative change in cardiomyocytes following IRI. In the
present study, we constructed both in vivo and in
vitro IRI models by performing ligation of the anterior
descending coronary artery (LAD) in mice and by exposing primary
cardiomyocytes to H2O2, respectively. We
aimed to discover the biological role of SNHG16 in affecting the
proliferative ability of cardiomyocytes.
Materials and methods
Establishment of I/R mouse model
A total of 50 male mice (180-220 g, 6-8 weeks of
age) obtained from the Experimental Animal Center, Qingdao
University were randomly divided into 5 groups (sham group and 4
time-point groups, 10 mice for each group). Mice were housed in a
temperature controlled room (21±2˚C) with a 12:12-h light/dark
cycle (lights on at 06:00). All mice had free access to water and
food. Animal experiments were repeated in triplicate, and a total
of 150 mice were used in this study. The mice were anesthetized
with 10% chloral hydrate at a dose of 200-250 mg/kg, intubated, and
mechanically ventilated. No mice exhibited signs of peritonitis
after the administration of 10% chloral hydrate. After thoracotomy,
the left anterior descending coronary artery was inserted using 4.0
suture at the beginning 2-3 cm. A small soft film was placed
between the suture and the artery. Ligation was performed after
observation of 15-min stabilization of breath and blood pressure.
Thirty minutes later, the ligature was removed and reperfusion was
allowed for 120 min. For the sham group, mice were subjected to
thoracotomy without ligation. During the experiments, mouse
electrocardiogram was monitored. ST segment elevation and upright T
wave were observed after ligation, which were relieved after
reperfusion. All animal protocols were approved by the Animal
Ethics Committee of Qingdao University (Qingdao, Shandong,
China).
Cell culture
According to previously described methods (22,23),
newborn mice (1-2 days of age) were sacrificed by cervical
dislocation (after being anesthetized using 10% chloral hydrate at
a dose of 250 mg/kg). No mice exhibited signs of peritonitis after
the administration of 10% chloral hydrate. Mouse heart was
immediately collected, washed in ice-cold HBSS and cut into
1-mm3 pieces. Heart tissues were digested in trypsin
containing 0.125% EDTA (ethylenediaminetetraacetic acid) overnight
at 4˚C in the dark. On the following day, heart homogenate was
incubated in collagenase II solution in a water bath shaker (37˚C,
100 x g). Dulbecco's modified Eagle's medium (DMEM) (Gibco; Thermo
Fisher Scientific, Inc.) containing 10% fetal bovine serum (FBS)
(Gibco; Thermo Fisher Scientific, Inc.) was utilized to maintain
the activity of cardiomyocytes. In order to construct the IRI model
in vitro, the cells were cultured in the medium containing
200 µmol/l H2O2 for 0, 6, 12, 24 h,
respectively.
Cell transfection
Cardiomyocytes were inoculated in a 96-well plate at
6x106 cells/ml. Twenty-four hours later, the cells were
transfected with plasmid using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) following the manufacturer's
instructions. After transfection for 24 h, the cells were then used
for the experiments. The final concentration of the pRL-CMV
plasmids was 0.1 µg/µl. All the mimics, inhibitors and plasmids
were purchased from GenePharma (Shanghai, China). The sequences
were: miR-770-5p mimic, 5'-UCCAGUACCACGUGUCAGGGCCA-3';
miRNA-770-5p-control, 5'-UCGCUUGGUGCAGGUCGGGAA-3 and miR-770-5p
inhibitor, 5'-UGGCCCUGACACGUGGUACUGGA-3'.
RNA extraction and quantitative
real-time polymerase chain reaction (qPCR)
Extraction of total RNA in cells was performed using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). RNA
was reversely transcribed into complementary deoxyribose nucleic
acid (cDNA), and the latter was applied for PCR using SYBR Green
method (Takara) for 5 min at 94˚C, and 40 cycles for 30 sec at
94˚C, 30 sec at 55˚C and 90 sec at 72˚C with Roche LightCycler 480
(Roche, USA). U6 and GAPDH were enrolled as internal controls.
Primers were as below: SNHG16 forward, 5'-CAGAATGCCATGGTTTCCCC-3'
and reverse, 5'-TGGCAAGAGACTTCCTGAGG-3'; GAPDH forward,
5'-CGGAGTCAACGGATTTGGTCGTAT-3' and reverse,
5'-AGCCTTCTCCATGGTGGTGAAGAC-3'; U6 forward, 5'-CTCGCTTCGGCAGCACA-3'
and reverse, 5'-AACGCTTCACGAATTTGCGT-3'.
Dual-luciferase reporter gene
assay
Cardiomyocytes were co-transfected with SNHG16-MT
(mutant)/SNHG16-WT (wild-type) and miRNA-770-5p mimics/NC,
respectively. After transfection of 48 h, cells were lysed for
determining relative luciferase activity with a Promega device
(Promega Corp.). The 3'-UTR of wild-type EGFR and the containing
mutations in the putative binding site were inserted downstream of
the firefly luciferase reporter into the psiCHECK-2 vector (Promega
Corp.). The corresponding mutant construct was created through
mutating the seed regions of the miR-133b binding sites and was
named as 3'-UTR mut EGFR. Cells were seeded into a 12-well plate at
80% confluence and cotransfected with 0.5 µg reporter plasmid, 40
nM miR-133b mimics or negative control with Lipofectamine 2000. For
the reason of transfection efficiency, all samples were
cotransfected with 0.05 µg pRL-CMV plasmid expressing
Renilla Luciferase (Promega Corp.) as an internal control.
Luciferase assay was conducted 48 h after transfection by Dual
Luciferase Reporter Assay System (Promega Corp.). Firefly
luciferase activity was standardized to Renilla luciferase
activity. There experiments were conducted three times for each
assay.
Cell Counting Kit-8 (CCK-8)
Cardiomyocytes were seeded in the 96-well plate at
2x103 cells per well and cultured overnight. Absorbance
(A) at 450 nm was recorded at the appointed time points using the
CCK-8 Kit (Dojindo Laboratories) by the TECAN infinite M200
Multimode microplate reader (Tecan).
5-Ethynyl-2'-deoxyuridine (EdU)
Cardiomyocytes were seeded in a 96-well plate with
300 cells per well. Cardiomyocytes were labeled with 50 µmol/l EdU
at 37˚C for 2 h. After 30-min fixation in 4% paraformaldehyde,
cells were incubated with phosphate-buffered saline (PBS)
containing 0.5% Triton-100 for 20 min. After washing with PBS
containing 3% bovine serum albumin (BSA), 100 µl of dying solution
was applied per well for a 1-h incubation in dark. Subsequently,
the cells were counterstained with 100 µl of Hoechst for 10 min.
EdU-positive cells, Hoechst-labeled cells and merged images were
captured under a fluorescent microscope (IX70; Olympus).
Statistical analyses
Statistical Product and Service Solutions (SPSS)
18.0 (SPSS Inc.) was used for data analyses. Data are expressed as
mean ± standard deviation. Differences between groups were analyzed
by the t-test. Comparisons among multiple groups were performed by
one-way analysis of variance (ANOVA) followed by least significant
difference (LSD) analysis. Comparisons among four groups were
analyzed by Tukey's test. Each experiment was repeated at least
three times. P<0.05 was considered to indicate a statistically
significant difference.
Results
SNHG16 is downregulated in IRI
models
The in vivo IRI model in mice was first
constructed by performing LAD. With the prolongation of
reperfusion, the relative level of SNHG16 was significantly
downregulated (Fig. 1A). Meanwhile,
primary cardiomyocytes were isolated from newborn mice and
subjected to H2O2 treatment. SNHG16
was significantly downregulated as well with the prolongation of
H2O2 treatment in cardiomyocytes (Fig. 1B).
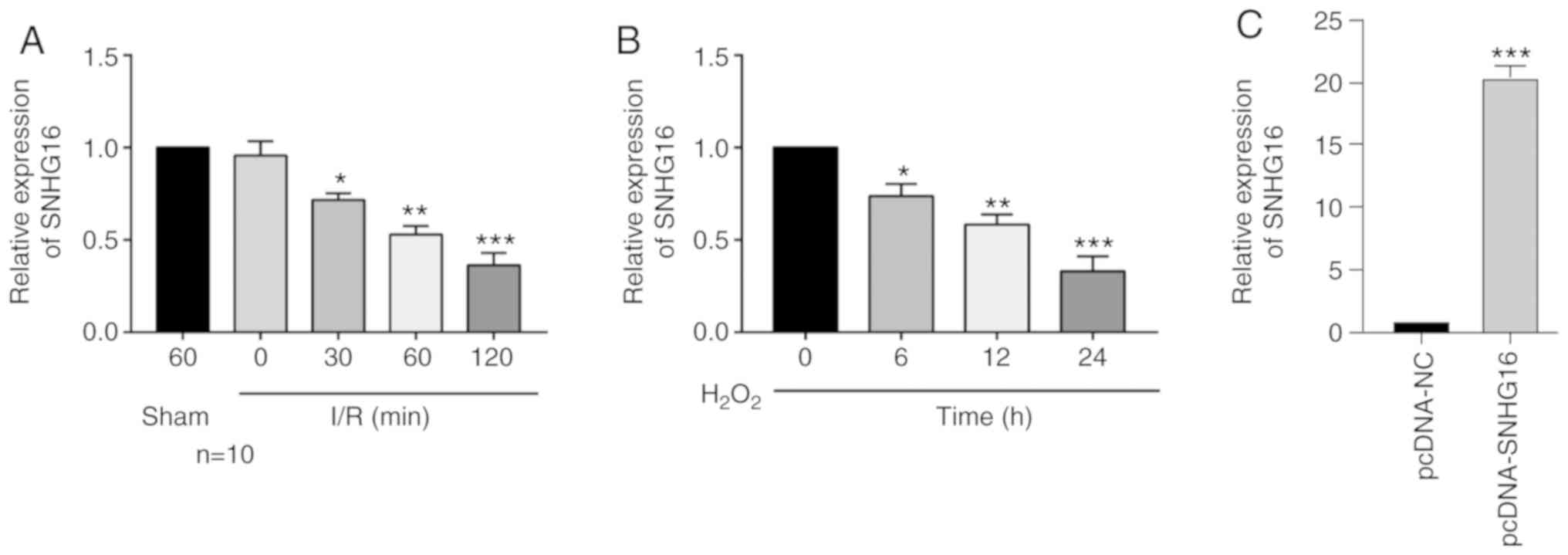 | Figure 1SNHG16 is downregulated in the
IRI models. (A) Relative level of SNHG16 in mice undergoing
sham operations for 60 min, and IRI for 0, 30, 60 and 120 min.
*P<0.05, **P<0.01,
***P<0.001, compared to the sham group. (B) Relative
level of SNHG16 in primary cardiomyocytes exposed to
H2O2 for 0, 6, 12 and 24 h.
*P<0.05, **P<0.01,
***P<0.001, compared to the 0 h group. (C) Relative
level of SNHG16 in primary cardiomyocytes transfected with pcDNA-NC
and pcDNA-SNHG16. ***P<0.001, compared to the
pcDNA-NC group. SNHG16, small nuclear host gene 16; IRI,
ischemia/reperfusion injury. |
SNHG16 accelerates the proliferation
of cardiomyocytes
To uncover the biological function of SNHG16
in IRI, we constructed pcDNA-SNHG16. Transfection of pcDNA-SNHG16
in untreated and H2O2-treated cardiomyocytes
markedly upregulated the SNHG16 level, indicating a high
transfection efficacy (Figs. 1C and
2A). At 48 and 72 h, transfection of
pcDNA-SNHG16 in H2O2-treated cardiomyocytes
enhanced the viability relative to the
H2O2+pcDNA-NC group (Fig. 2B). EdU assay identically showed an
elevation in the EdU-positive rate after overexpression of SNHG16
in cardiomyocytes treated with H2O2 compared
to H2O2+pcDNA-NC group (Fig. 2C).
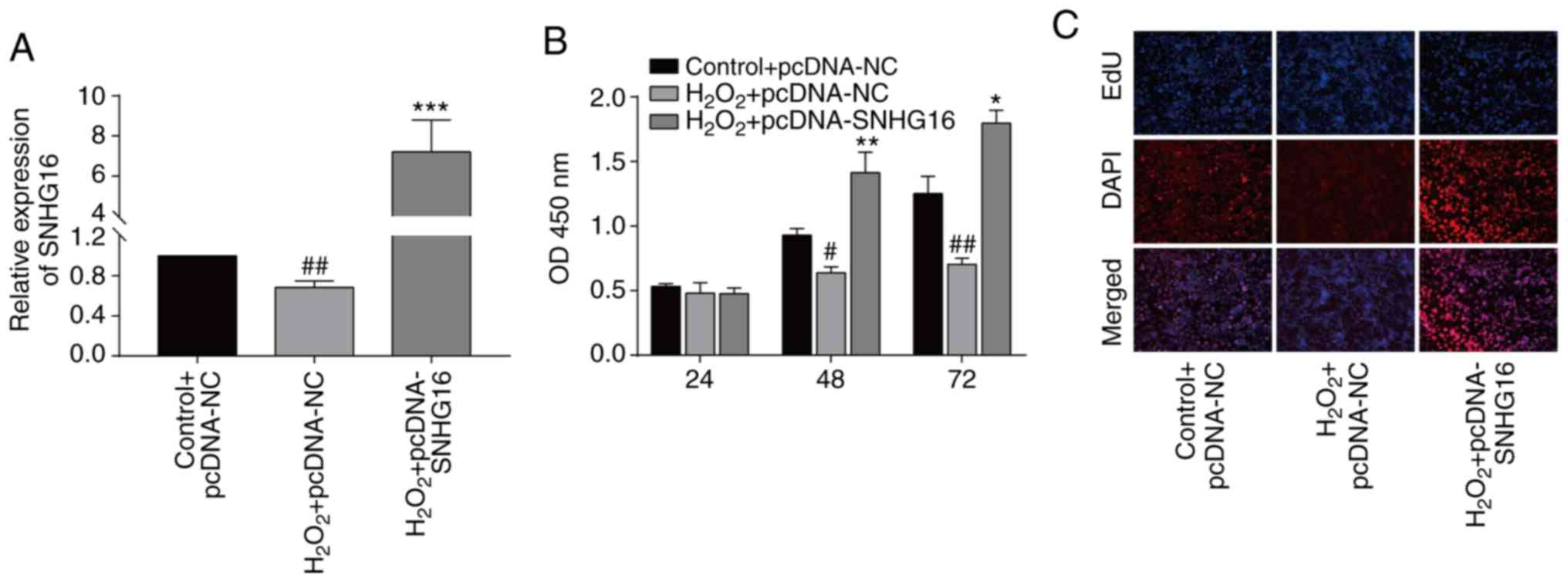 | Figure 2SNHG16 accelerates the
proliferative ability of cardiomyocytes. (A) Relative level of
SNHG16 in the primary cardiomyocytes following transfection and/or
exposure to H2O2 in the following groups,
control+pcDNA-NC, H2O2+pcDNA-NC or
H2O2+pcDNA-SNHG16. ##P<0.01,
H2O2+pcDNA-SNHG16 group vs.
H2O2+pcDNA-NC group,
***P<0.001, H2O2+pcDNA-SNHG16
vs. control+pcDNA-NC group. (B) CCK-8 assay demonstrates the
viability in the primary cardiomyocytes in the groups:
Control+pcDNA-NC, H2O2+pcDNA-NC or
H2O2+pcDNA-SNHG16. #P<0.05,
##P<0.01 H2O2+pcDNA-SNHG16
group vs. H2O2+pcDNA-NC group,
*P<0.05, **P<0.01,
H2O2+pcDNA-SNHG16 group vs. control+pcDNA-NC
group. (C) DAPI-labeled, EdU-labeled and merged images of primary
cardiomyocytes in the group: Control+pcDNA-NC,
H2O2+pcDNA-NC or
H2O2+pcDNA-SNHG16 (magnification, x100).
SNHG16, small nuclear host gene 16. |
SNHG16 was able to bind to
miRNA-770-5p
In order to predict the targets of SNHG16, the
sequence of SNHG16 was obtained from NCBI (https://www.ncbi.nlm.nih.gov), and then the sequence
was uploaded to the online tool miRDB (http://mirdb.org).
The results showed that miRNA-30e-5p, miRNA-205-5p, miRNA-770-5p
and miRNA-564 were predicted to be the targets of SNHG16.
miRNA-770-5p was the only miRNA to be significantly upregulated
after 24-h H2O2 treatment in the
cardiomyocytes (Fig. 3A). In mice
undergoing 120-min I/R injury, only the miRNA-770-5p level was
highly significantly upregulated (Fig.
3B). Potential binding sequences between SNHG16 and
miRNA-770-5p were identified (Fig.
3C). Transfection of the miR-770-5p-mimics and
miR-770-5p-inhibitor significantly upregulated and downregulated
miR-770-5p, respectively, in the cardiomyocytes when compared to
the miR-770-5p-control (Fig. 3D).
Furthermore, luciferase activity was markedly reduced after
co-transfection of SNHG16-WT and miRNA-770-5p mimics, verifying
their binding relationship (Fig.
3E). Transfection of the miRNA-770-5p inhibitor significantly
upregulated the SNHG16 level, and in contrast, transfection
of the miRNA-770-5p mimics significantly elevated the SNHG16
level (Fig. 3F). Moreover,
transfection of pcDNA-SNHG16 significantly downregulated the
miRNA-770-5p level, confirming their negative correlation (Fig. 3G).
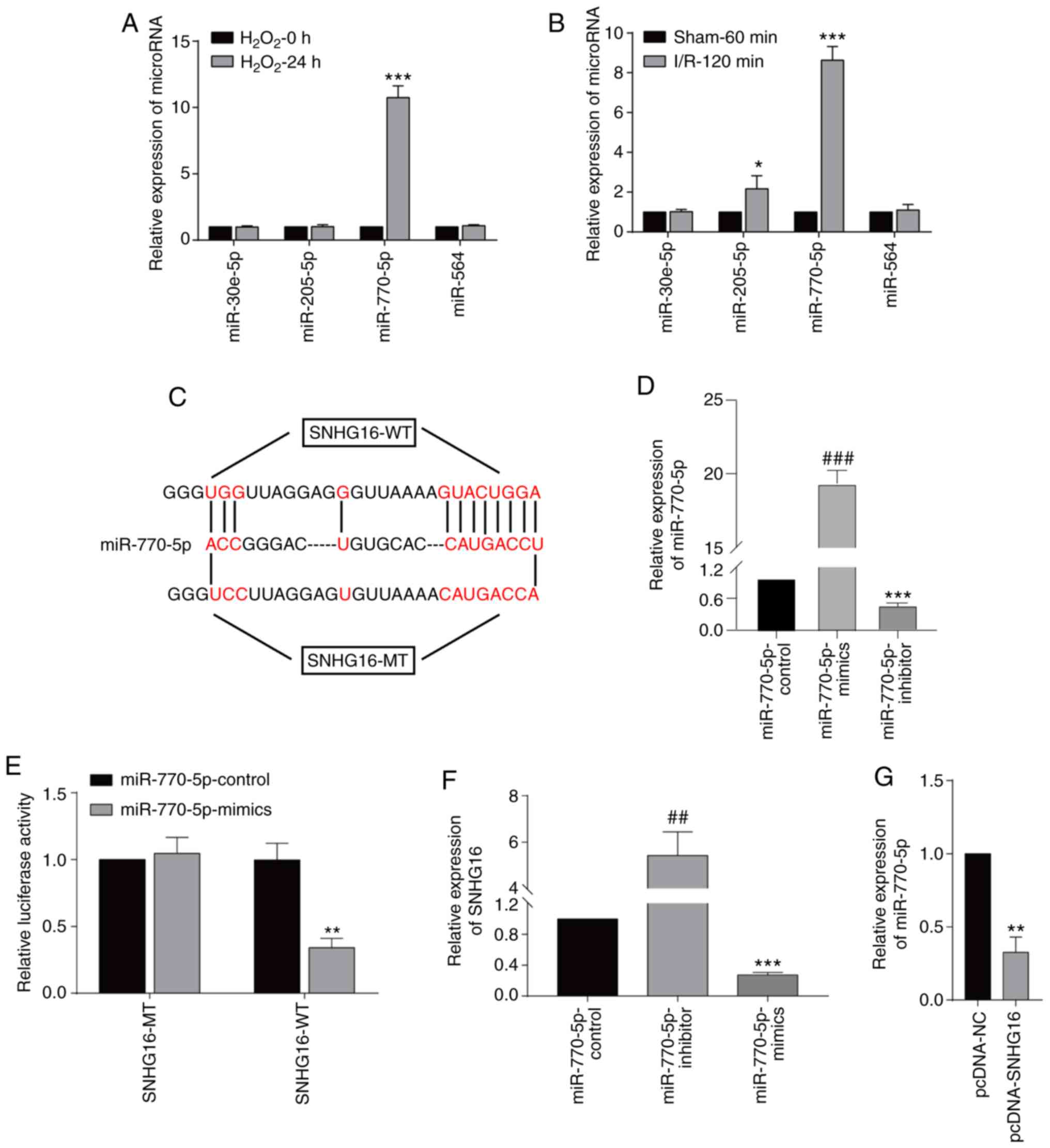 | Figure 3SNHG16 binds to miR-770-5p.
(A) Relative levels of miRNA-30e-5p, miRNA-205-5p, miRNA-770-5p and
miRNA-564 in primary cardiomyocytes undergoing 0 or 24-h
H2O2 exposure. ***P<0.001
H2O2-24 h group vs. H2O2-0 h group (B) Relative levels of
miRNA-30e-5p, miRNA-205-5p, miRNA-770-5p and miRNA-564 in mice
undergoing 60-min sham operation or 120-min IRI.
*P<0.05, ***P<0.001, I/R-120 min vs.
sham 60 group (C) Potential binding sequences between SNHG16
and miR-770-5p. (D) Relative level of miR-770-5p in primary
cardiomyocytes transfected with miR-770-5p-control,
miR-770-5p-mimics and miR-770-5p-inhibitor.
###P<0.001, miR-770-5p-inhibitor group vs.
miR-770-5p-mimics group, ***P<0.001,
miR-770-5p-inhibitor group vs. miR-770-5p-control group. (E)
Dual-luciferase reporter gene assay showed the luciferase activity
in cardiomyocytes co-transfected with SNHG16-MT/SNHG16-WT and
miRNA-770-5p mimics/NC. **P<0.01, miR-770-5p-mimics
group vs. miR-770-5p-control group. (F) SNHG16 expression following
miRNA-770-5p mimics or miRNA-770-5p inhibitor transfection.
##P<0.01, miR-770-5p-inhibitor group vs.
miR-770-5p-mimics group, ***P<0.001,
miR-770-5p-mimics group vs. miR-770-5p-control group. (G) Relative
levels of miR-770-5p in cardiomyocytes transfected with pcDNA-NC or
pcDNA-SNHG16. **P<0.01, pcDNA-NC group vs.
pcDNA-SNHG16 group. SNHG16, small nuclear host gene 16; IRI,
ischemia/reperfusion injury. |
Overexpression of miRNA-770-5p
reversed the role of SNHG16 in cardiomyocytes
Rescue experiments were carried out to explore the
involvement of miRNA-770-5p in SNHG16-mediated cardiomyocyte
cytoactivation. After H2O2 treatment, the
upregulation of SNHG16 in cardiomyocytes transfected with
pcDNA-SNHG16 was partially downregulated by co-transfection of
miRNA-770-5p mimics (Fig. 4A).
Moreover, the enhanced EdU-positive rate in cardiomyocytes
overexpressing SNHG16 was reduced to some extent by miRNA-770-5p
overexpression (Fig. 4B).
Collectively, SNHG16 promotes the cytoactivation of
cardiomyocytes in H2O2-treated cardiomyocytes
by negatively regulating miRNA-770-5p.
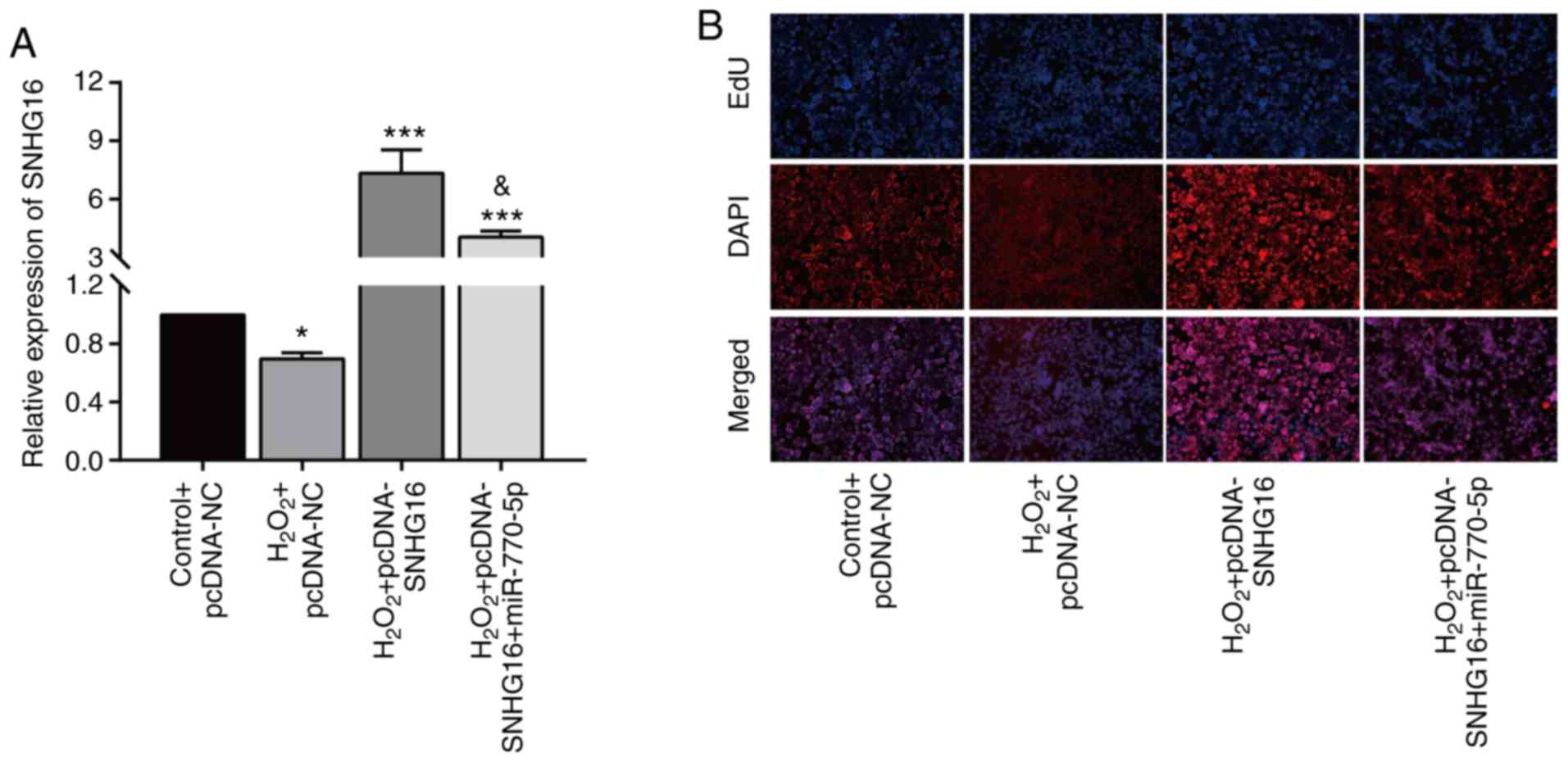 | Figure 4Overexpression of miR-770-5p reverses
the effect of SNHG16 on cardiomyocytes. (A) Relative level
of SNHG16 in primary cardiomyocytes in the control+pcDNA-NC,
H2O2+pcDNA-NC,
H2O2+pcDNA-SNHG16 or
H2O2+pcDNA-SNHG16+miR-770-5p mimic groups.
*P<0.05, ***P<0.001, compared to the
control+pcDNA-NC group; &P<0.05, compared to the
H2O2+pcDNA-SNHG16 group. (B) DAPI-labeled,
EdU-labeled and merged images of primary cardiomyocytes transfected
in the control+pcDNA-NC, H2O2+pcDNA-NC,
H2O2+pcDNA-SNHG16 or
H2O2+pcDNA-SNHG16+miR-770-5p mimic groups
(magnification, x100). SNHG16, small nuclear host gene 16. |
Discussion
Ischemic heart disease following myocardial
infarction and congestive heart failure is the leading cause of
global death (24). With the
improvement of therapeutic strategies, thrombolytic therapy is
widely applied in the treatment of acute myocardial infarction. It
is able to quickly restore the blood flow of the coronary artery in
the ischemic area, causing myocardial reperfusion, and decline of
necrotic myocardial area (25).
However, studies have shown that reperfusion after ischemia can
aggravate cardiomyocyte damage, mainly manifested by accelerated
myocardial necrosis, myocardial infarct size expansion, no-reflow
phenomenon, arrhythmia and other symptoms (25,26).
Therefore, how to minimize the occurrence of reperfusion injury has
become the focus of research.
Recent studies have shown the involvement of lncRNAs
in the regulation of cardiovascular disease (27,28).
Wang et al (29) reported
that lncRNA NRF regulates programmed necrosis and myocardial injury
during ischemia/reperfusion injury (IRI) by targeting miR-873,
providing a potential target for the treatment of myocardial
ischemic diseases. In focal myocardial IRI, lncRNA UCA1 was
found to negatively regulate p27 expression and to exert a
pro-apoptotic role in primary cardiomyocytes (30). In the present paper, long non-coding
RNA small nuclear host gene 16 (lncRNA SNHG16) was time-dependently
downregulated in mice undergoing LAD and in primary cardiomyocytes
treated with H2O2. Both CCK-8 and EdU assays
demonstrated the promotive effect of SNHG16 on the
proliferative ability of cardiomyocytes. The findings indicate that
SNHG16 may be a key regulator involved in IRI.
A growing body of evidence has confirmed that
lncRNAs regulate miRNA expression and functions as competitive
endogenous (ce)RNAs (31,32). The ceRNA theory proposes a complex
regulatory network involving miRNAs, lncRNAs and circular
(circ)RNAs. They usually contain mRNA binding sites and influence
tumor progression by competitively binding microRNA response
elements (MREs) (33-35).
For example, SNHG16 was found to promote the proliferative
ability of osteosarcoma by competitively binding miR-205 and
further upregulating ZEB1 (14). SNHG16 was found to mediate
HMGB1 expression by absorbing miR-218-5p, thus exerting a
carcinogenic effect on pancreatic cancer (36). The present study found many
miRNA-binding sites in the sequences of SNHG16 through
bioinformatics analysis. Furthermore, dual-luciferase reporter gene
assay confirmed the binding between SNHG16 and miRNA-770-5p.
In primary cardiomyocytes, SNHG16 was negatively correlated
with miRNA-770-5p. In addition, overexpression of miRNA-770-5p
partially reversed the role of SNHG16 in accelerating
cardiomyocyte proliferation.
In conclusion, SNHG16 was downregulated in
the IRI mouse model and in the H2O2-exposed
primary cardiomyocytes. Overexpression of SNHG16 accelerated the
proliferative ability of cardiomyocytes following IRI by negatively
regulating miRNA-770-5p.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LY and TW designed the study and performed the
experiments. LY and YL established the animal models. JM and YP
collected the data, RY and YW analyzed the data, and LY and TW
prepared the manuscript. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
This study was approved by the Animal Ethics
Committee of Qingdao University Animal Center.
Patient consent for publication
No patients participated in this study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hausenloy DJ and Yellon DM: Myocardial
ischemia-reperfusion injury: A neglected therapeutic target. J Clin
Invest. 123:92–100. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
2
|
Song XD, Feng JP and Yang RX: Alamandine
protects rat from myocardial ischemia-reperfusion injury by
activating JNK and inhibiting NF-KB. Eur Rev Med Pharmacol Sci.
23:6718–6726. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ibanez B, Heusch G, Ovize M and Van de
Werf F: Evolving therapies for myocardial ischemia/reperfusion
injury. J Am Coll Cardiol. 65:1454–1471. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Martindale JJ and Metzger JM: Uncoupling
of increased cellular oxidative stress and myocardial ischemia
reperfusion injury by directed sarcolemma stabilization. J Mol Cell
Cardiol. 67:26–37. 2014.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Li X, Wu Z, Fu X and Han W: Long noncoding
RNAs: Insights from biological features and functions to diseases.
Med Res Rev. 33:517–553. 2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Schmitz SU, Grote P and Herrmann BG:
Mechanisms of long noncoding RNA function in development and
disease. Cell Mol Life Sci. 73:2491–2509. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Dabek J, Owczarek A, Gasior Z, Ulczok R,
Skowerski M, Kulach A, Mazurek U and Bochenek A: Oligonucleotide
microarray analysis of genes regulating apoptosis in chronically
ischemic and postinfarction myocardium. Biochem Genet. 46:241–247.
2008.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Pitts KR, Derry JM, Kerkof K, Lawrence WA
and Toombs CF: Differentially regulated functional gene clusters
identified during ischemia and reperfusion in isolated cardiac
myocytes using coverslip hypoxia. J Pharmacol Toxicol Methods.
57:42–51. 2008.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Small EM and Olson EN: Pervasive roles of
microRNAs in cardiovascular biology. Nature. 469:336–342.
2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wu HJ, Tang GM, Shao PY, Zou HX, Shen WF,
Huang MD, Pan HH, Zhai CL and Qian G: Long non-coding RNA NEAT1
modulates hypoxia/reoxygenation-induced cardiomyocyte injury via
targeting microRNA-520a. Exp Ther Med. 18:2199–2206.
2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Gong X, Zhu Y, Chang H, Li Y and Ma F:
Long noncoding RNA MALAT1 promotes cardiomyocyte apoptosis after
myocardial infarction via targeting miR-144-3p. Biosci Rep.
39(BSR20191103)2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Du XJ, Wei J, Tian D, Yan C, Hu P, Wu X,
Yang W and Hu X: NEAT1 promotes myocardial ischemia-reperfusion
injury via activating the MAPK signaling pathway. J Cell Physiol.
234:18773–18780. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhu C, Cheng D, Qiu X, Zhuang M and Liu Z:
Long noncoding RNA SNHG16 promotes cell proliferation by sponging
MicroRNA-205 and upregulating ZEB1 expression in osteosarcoma. Cell
Physiol Biochem. 51:429–440. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Feng F, Chen A, Huang J, Xia Q, Chen Y and
Jin X: Long noncoding RNA SNHG16 contributes to the development of
bladder cancer via regulating miR-98/STAT3/Wnt/β-catenin pathway
axis. J Cell Biochem. 119:9408–9418. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Han W, Du X, Liu M, Wang J, Sun L and Li
Y: Increased expression of long non-coding RNA SNHG16 correlates
with tumor progression and poor prognosis in non-small cell lung
cancer. Int J Biol Macromol. 121:270–278. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yang BY, Meng Q, Sun Y, Gao L and Yang JX:
Long non-coding RNA SNHG16 contributes to glioma malignancy by
competitively binding miR-20a-5p with E2F1. J Biol Regul Homeost
Agents. 32:251–261. 2018.PubMed/NCBI
|
|
18
|
Yang XS, Wang GX and Luo L: Long
non-coding RNA SNHG16 promotes cell growth and metastasis in
ovarian cancer. Eur Rev Med Pharmacol Sci. 22:616–622.
2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Cai C, Huo Q, Wang X, Chen B and Yang Q:
SNHG16 contributes to breast cancer cell migration by competitively
binding miR-98 with E2F5. Biochem Biophys Res Commun. 485:272–278.
2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang D, Lin B, Zhang W and Wang X:
Up-regulation of SNHG16 induced by CTCF accelerates cardiac
hypertrophy by targeting miR-182-5p/IGF1 axis. Call Biol Int, Mar
3, 2020 (Epub ahead of print).
|
|
21
|
An JH, Chen ZY, Ma QL, Wang HJ, Zhang JQ
and Shi FW: LncRNA SNHG16 promoted proliferation and inflammatory
response of macrophages through miR-17-5p/NF-κB signaling pathway
in patients with atherosclerosis. Eur Rev Med Pharmacol Sci.
23:8665–8677. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Goldenberg I, Shainberg A, Jacobson KA,
Shneyvays V and Grossman E: Adenosine protects against angiotensin
II-induced apoptosis in rat cardiocyte cultures. Mol Cell Biochem.
252:133–139. 2003.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Chen J, Larsson L, Haugen E, Fedorkova O,
Angwald E, Waagstein F and Fu M: Effects of autoantibodies removed
by immunoadsorption from patients with dilated cardiomyopathy on
neonatal rat cardiomyocytes. Eur J Heart Fail. 8:460–467.
2006.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Pagidipati NJ and Gaziano TA: Estimating
deaths from cardiovascular disease: A review of global
methodologies of mortality measurement. Circulation. 127:749–756.
2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Dong J, Zhao Y and He XK: Down-regulation
of miR-192 protects against rat ischemia-reperfusion injury after
myocardial infarction. Eur Rev Med Pharmacol Sci. 22:6109–6118.
2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wu H, Ye M, Yang J and Ding J: Modulating
endoplasmic reticulum stress to alleviate myocardial ischemia and
reperfusion injury from basic research to clinical practice: A long
way to go. Int J Cardiol. 223:630–631. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wang K, Long B, Zhou LY, Liu F, Zhou QY,
Liu CY, Fan YY and Li PF: CARL lncRNA inhibits anoxia-induced
mitochondrial fission and apoptosis in cardiomyocytes by impairing
miR-539-dependent PHB2 downregulation. Nat Commun.
5(3596)2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Viereck J, Kumarswamy R, Foinquinos A,
Xiao K, Avramopoulos P, Kunz M, Dittrich M, Maetzig T, Zimmer K,
Remke J, et al: Long noncoding RNA Chast promotes cardiac
remodeling. Sci Transl Med. 8(326ra22)2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang K, Liu F, Liu CY, An T, Zhang J, Zhou
LY, Wang M, Dong YH, Li N, Gao JN, et al: The long noncoding RNA
NRF regulates programmed necrosis and myocardial injury during
ischemia and reperfusion by targeting miR-873. Cell Death Differ.
23:1394–1405. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liu Y, Zhou D, Li G, Ming X, Tu YF, Tian
J, Lu H and Yu B: Long non coding RNA-UCA1 contributes to
cardiomyocyte apoptosis by suppression of p27 expression. Cell
Physiol Biochem. 35:1986–1998. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Ulitsky I: Interactions between short and
long noncoding RNAs. FEBS Lett. 592:2874–2883. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Chan JJ and Tay Y: Noncoding RNA: RNA
regulatory networks in cancer. Int J Mol Sci.
19(1310)2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Kartha RV and Subramanian S: Competing
endogenous RNAs (ceRNAs): New entrants to the intricacies of gene
regulation. Front Genet. 5(8)2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Karreth FA and Pandolfi PP: ceRNA
cross-talk in cancer: When ce-bling rivalries go awry. Cancer
Discov. 3:1113–1121. 2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Liu S, Zhang W, Liu K and Liu Y: LncRNA
SNHG16 promotes tumor growth of pancreatic cancer by targeting
miR-218-5p. Biomed Pharmacother. 114(108862)2019.PubMed/NCBI View Article : Google Scholar
|














