Introduction
Human intervertebral disc degeneration (IDD) refers
to the loss of typical structure and function of intervertebral
disc tissue, gradual disappearance of the nucleus pulposus (NP)
tissue, blurring and disappearing of the boundary between NP and
fibrous rings, accompanied by degenerated NP cells (NPCs) (1). Various types of pathological damage and
aging of physiological functions lead to the degenerative changes
of the intervertebral discs and imbalance in the spine. Once the
progression of IDD begins, it is difficult to reverse (2). Degenerative changes of the disc mainly
occur in the NP of the inner tissue of the intervertebral disc. The
NP is composed of NPCs and the extracellular matrix (ECM). The
former is derived from mesenchymal stem cells and functions to
maintain the healthy metabolism of the NP. Concurrently, NPC
activity is also affected by the physical structure and material
composition of the ECM (3).
Several functional growth factors and their
receptors have been found in normal and degenerative disc tissues,
including insulin-like growth factor-1, basic fibroblast growth
factor and platelet-derived growth factor (4). In recent years, transforming growth
factor (TGF)-β is a cytokine that has been confirmed to be closely
associated with IDD and plays an essential role in the development,
growth and maintenance of disc tissue (5). The TGF-β superfamily includes TGF-βs
(TGF-β1, 2, 3), activins and inhibins, growth differentiation
factors, bone morphogenetic proteins and Nodal. Among the TGF-β
superfamily, TGF-β1 is closely related to the development and
maturation of chondrocytes, as well as the maintenance of
chondrocytes in an undifferentiated state (6). It was reported that TGF-β1 can promote
the proliferation and differentiation of cartilage-like NPCs and
participate in the process of its damage repair (7). However, previous studies have also
demonstrated that TGF-β1 and TGF-β1 receptor expression increased
with the degree of degeneration of intervertebral disc tissue
compared with the normal control disc tissue (8,9). Chen
et al (10) also reported
increased TGF-β1 levels within IDD. An excess of TGF-β activation
exacerbates IDD, and suppression of the excessive TGF-β1
accumulation can prevent IDD development. Therefore, it was
observed that TGF-β1 has a dual effect on the development of IDD.
However, how excess TGF-β1 mediates IDD and NPC function has not
been fully elucidated.
In the TGF-β signaling pathway, its receptor ALK
tyrosine kinase receptor (ALK) 1/5 was reported to mediate Smad,
playing a vital role in the homeostasis of the ECM of the NP
(11-13).
The present study hypothesized that high-dose TGF-β1 regulates NPC
degeneration via the TGF-β receptors ALK1 and ALK5 and the
downstream Smad. Through the inhibition of different TGF receptors,
the present study aimed to explore the role of high-dose TGF-β1 in
the development of NPC degeneration to provide a scientific basis
for clinical prevention and treatment of IDD.
Materials and methods
Ethics statement
All the lumbar intervertebral disc tissues were
obtained with consent from the patients or their families, and the
project was approved by the Ethics Committee of The Second
Affiliated Hospital of Soochow University.
Reagents
Dulbecco's modified Eagle's medium/F12 (DMEM/F12),
fetal bovine serum (FBS), type II collagenase, trypsin,
penicillin-streptomycin, tumor necrosis factor (TNF)-α and bovine
serum albumin (BSA) were purchased from Sigma-Aldrich (Merck KGaA).
Phosphate buffered saline (PBS), goat serum, Elite ABC reagent, ECL
substrate (cat. no. P0018S), and radioimmunoprecipitation assay
(RIPA) lysate buffer (1:500; cat. no. A0277) were purchased from
Beyotime Institute of Biotechnology. Primary antibodies against
TGF-β1 (cat. no. sc-130348; 1:1,000), collagen-II (Col-II; cat. no.
sc-52658; 1:1,000), ALK1 (cat. no. sc-101556; 1:1,000), GAPDH (cat.
no. sc-47724; 1:3,000), and the secondary antibody m-immunoglobulin
G (IgG)κ binding protein conjugated to horseradish peroxidase (cat.
no. sc-516102; 1:500) were purchased from Santa Cruz Biotechnology,
Inc. Primary antibody against ALK5 (cat. no. ab31013; 1:1,000) and
Alexa Fluor 568 (cat. no. ab175473; 1:1,000) were from Abcam.
TRIzol reagent, RETROscript™ reverse transcription kit and SYBR™
Green master mix were purchased from Invitrogen (Thermo Fisher
Scientific, Inc.). The ALK5 inhibitor SB431542 (SB; 100 nM; cat.
no. s1067) was purchased from Selleck Chemicals. The ALK1 inhibitor
San 78-130 (San; 100 nM; cat. no. CS-0020876) was purchased from
AbaChemScene, LLC.
NP cell isolation and culture
NP tissues were collected from 18 patients during
May 2019 (10 male, 8 female; age range, 43 to 65 years) who
underwent spinal surgery for disc herniation in The Second
Affiliated Hospital of Soochow University (Suzhou, China).
Specimens were divided into four groups based on the Pfirrmann
(14) disc magnetic resonance
imaging (MRI) score: i) 2#, 3 samples; ii) 3#, 5 samples; iii) 4#,
4 samples; and iv) 5#, 6 samples. Specimens were washed three times
with sterile PBS. Subsequently, they were cut into small pieces and
sequentially digested with 0.25% trypsin and type II collagenase
for 6 h. The NP cell pellets were obtained after filtration and
seeded (1x105 per ml) in DMEM/F12 medium containing 10%
FBS and 1% penicillin-streptomycin. NP cells were cultured in an
incubator at 37˚C with 95% humidity and 5% CO2. A
TNF-α-induced NPC degeneration model was used as previously
described (15) to investigate
TGF-β1 expression in degenerated NPCs in vitro. NPCs were
treated with TGF-β1 or ALK1/5 inhibitor for 72 h at 37˚C. For the
co-treated group, NPCs were treated with TGF-β1 combined with
ALK1/5 inhibitor for 72 h at 37˚C.
Immunohistochemical (IHC)
staining
The NP tissue was fixed with 4% formaldehyde at room
temperature, rehydrated in descending alcohol series, embedded in
paraffin and cut into 5-µm-thick slices. Sections were dewaxed,
hydrated, heated in citric acid at 100˚C and blocked with 10% goat
serum for 1 h at room temperature. Sections were incubated with
TGF-β1 primary antibody overnight at 4˚C. The next day, sections
were incubated with biotinylated IgG Elite ABC reagent at room
temperature for 1 h, developed with 3,3'-diaminobenzidine and
counterstained with hematoxylin at room temperature for 5 min. The
positive area was imaged by a light confocal microscope
(magnification, x400).
Western blotting (WB)
Total protein of NP tissue or NPCs was extracted
with RIPA lysate buffer at a low temperature, and the protein
concentration was determined by bicinchoninic acid
spectrophotometry. Equal amounts of protein (50 µg/lane) were
separated using 10% SDS-PAGE. Proteins were then transferred to a
nitrocellulose membrane and blocked with 5% skimmed milk for 2 h at
room temperature. Subsequently, membranes were incubated with
diluted primary antibodies against collagen II, ALK1, ALK5 and
GAPDH overnight at 4˚C. Following washing, the membranes were
incubated with the secondary antibody for 1 h at room temperature.
The brands were exposed using ECL substrate. Finally, band gray
value analysis was performed with a gel image processing system
(VILBER FUSION FX5; Vilber Lourmat).
Immunofluorescence (IF) staining
NPCs (1x105 per ml) were seeded in
six-well plates. Prior to staining, cells were fixed with 4%
paraformaldehyde for 15 min at room temperature and then treated
with 0.1% Triton-X for 15 min. Subsequently, 5% BSA was used to
block NPCs for 1 h at room temperature. The cells were washed and
incubated with TGF-β1 primary antibody overnight at 4˚C. Following
incubation with Alexa Fluor 568-conjugated secondary antibody for 1
h in the dark, the staining intensity of NPCs was determined using
an inverted fluorescence microscope.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA of NPCs was extracted using TRIzol reagent
according to the manufacturer's instructions. RNA was reverse
transcribed into cDNA with RETROscript at 25˚C for 5 min and then
at 42˚C for 1 h. qPCR analysis of Smad1/2/3/5/8, collagen I/II,
tissue inhibitor of metalloproteinase-3 (TIMP-3), matrix
metalloproteinase-13 (MMP-13) and GAPDH mRNA expression was
performed using SYBR Green master mix with the following reaction
conditions: 95˚C for 5 sec; followed by 45 cycles of 92˚C for 15
sec, 58˚C for 8 sec and 60˚C for 30 sec; and a final extension at
72˚C for 5 min. mRNA levels were normalized to GAPDH expression and
calculated using the 2-∆∆Cq method
(16). The primers used for qPCR are
listed in Table I.
 | Table IPrimer of the genes for RT-qPCR. |
Table I
Primer of the genes for RT-qPCR.
| Gene | Primer sequences
(5'→3') |
|---|
| Collagen II | F:
TGGACGATCAGGCGAAACC |
| | R:
GCTGCGGATGCTCTCAATCT |
| Collagen I | F:
GAGGGCCAAGACGAAGACATC |
| | R:
CAGATCACGTCATCGCACAAC |
| TIMP-3 | F:
CATGTGCAGTACATCCATACGG |
| | R:
CATCATAGACGCGACCTGTCA |
| MMP-13 | F:
ACTGAGAGGCTCCGAGAAATG |
| | R:
GAACCCCGCATCTTGGCTT |
| Smad1 | F:
AGAGACTTCTTGGGTGGAAACA |
| | R:
ATGGTGACACAGTTACTCGGT |
| Smad2 | F:
CGTCCATCTTGCCATTCACG |
| | R:
CTCAAGCTCATCTAATCGTCCTG |
| Smad3 | F:
TGGACGCAGGTTCTCCAAAC |
| | R:
CCGGCTCGCAGTAGGTAAC |
| Smad5 | F:
CCAGCAGTAAAGCGATTGTTGG |
| | R:
GGGGTAAGCCTTTTCTGTGAG |
| Smad8 | F:
CTAGGCTGGAAGCAAGGAGAT |
| | R:
GGGGAATCGTGACGCATTT |
| GAPDH | F:
ACAACTTTGGTATCGTGGAAGG |
| | R:
GCCATCACGCCACAGTTTC |
Statistical analysis
Data are presented as the mean ± SD. GraphPad
Version 8.4 (GraphPad Software, Inc.) was used to perform
statistical analysis and generate graphs. Differences between two
groups were analyzed using unpaired Student's t-test. Comparison
between multiple groups was performed using one-way ANOVA followed
by Bonferroni as the post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Increased expression of TGF-β1 in
degenerated human NP tissues
To investigate the levels of TGF-β1 in degenerated
human NP tissues, 18 NP samples were collected from patients
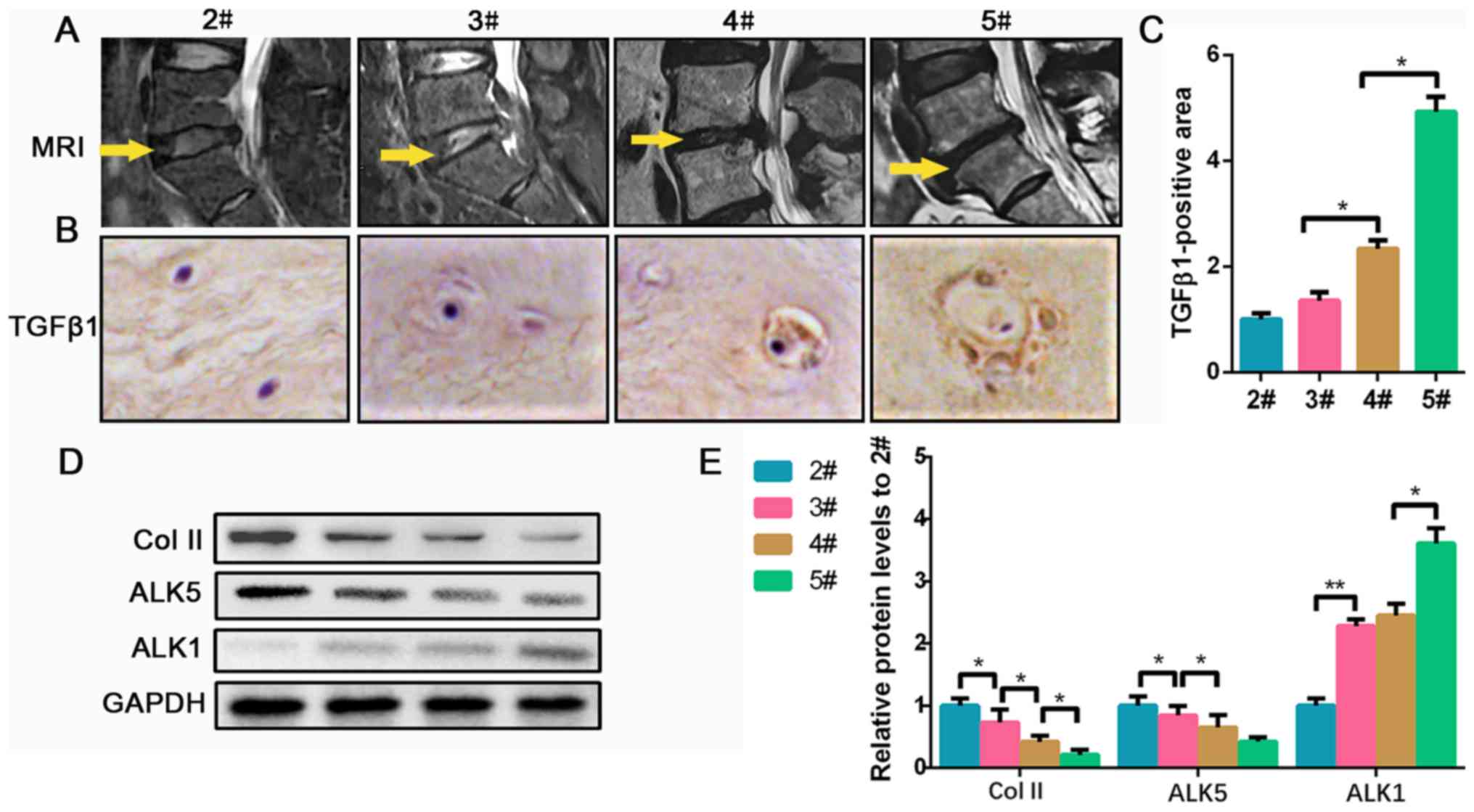
undergoing disc herniation operation in our hospital. As shown in
the MRI in Fig. 1A, the operation
section of 2# is higher compared with the others, and the section
of 3# is brighter and higher compared with 4# and 5#, indicating
that the disc of 2# contained a higher level of water and ECM
compared with the other groups. Furthermore, the IHC results
indicated that the NPCs tend to be hypertrophic and polynuclear in
severely degenerated conditions, and the expression of TGF-β1
increased according to the level of NP degeneration (Fig. 1B and C). Collagen II is an important component
secreted by NPCs in the ECM of the NP, which was found to be
decreased, most likely due to the degeneration of NPCs (17). ALKs (ALK1-7) exert kinase activity
and belong to the type I receptor of the TGF-β superfamily; they
can activate Smad1/2/3/5/8, collectively known as
receptor-regulated Smad protein (15). ALK5-Smad2/3 and ALK1-Smad1/5/8
signaling pathways play opposite roles in regulating the
homeostasis of chondrocytes (18).
Therefore, the expression of ALK1 and ALK5 in degenerated NP
tissues was investigated. Collagen II protein gradually decreased
in groups 2# to 5#. For the ALK5 levels, no significance was found
between 4# and 5#, but 4# was significantly less than 2/3#.
Meanwhile, ALK1 protein gradually increased as the disc
degeneration grade increased (Fig.
1D and E). Collectively, the
data suggested that TGF-β1 accumulates at higher levels as the NP
tissue becomes increasingly degraded, which is accompanied with a
reduction in ALK5 and an increase in ALK1 expression.
TGF-β1 is increased in TNF-α-treated
human NPCs
To investigate TGF-β1 expression in degenerated NPCs
in vitro, a TNF-α-induced NPC degeneration model was used as
previously described (19). NPCs
were isolated from 2# to 5# degenerated NP tissues. Following
treatment with TNF-α, TGF-β1 expression markedly increased compared
with non-treatment cells in each group (Fig. 2A and C). However, no significant differences in
ALK5 expression was found before and after TNF-α treatment in 3#,
4# and 5# groups. ALK1 expression in groups 2#, 3# and 4#
significantly increased following treatment with TNF-α, consistent
with results observed in NP tissues (Fig. 2B and D). Since the ALK5 content of NPCs from
groups 3# to 5# did not significantly differ following TNF-α
treatment, to better reflect the changing trend of ALK5, 2# NPCs
with lower levels of degeneration were used in subsequent
experiments. Additionally, Smad1/2/3/5/8 mRNA expression levels in
NPCs were evaluated. TNF-α induced the decrease of Smad3 and
significantly upregulated Smad1/5/8 levels compared with the
controls (Fig. 2E). ECM mRNA
expression was also analyzed. Collagen II and TIMP-3 decreased
following TNF-α treatment, whereas a marked increase of collagen I
and MMP-13 expression was observed (Fig.
2F). Therefore, these results indicated that TNF-α induced NPC
degeneration and upregulated TGF-β1, ALK1 and Smad1/5/8
expression.
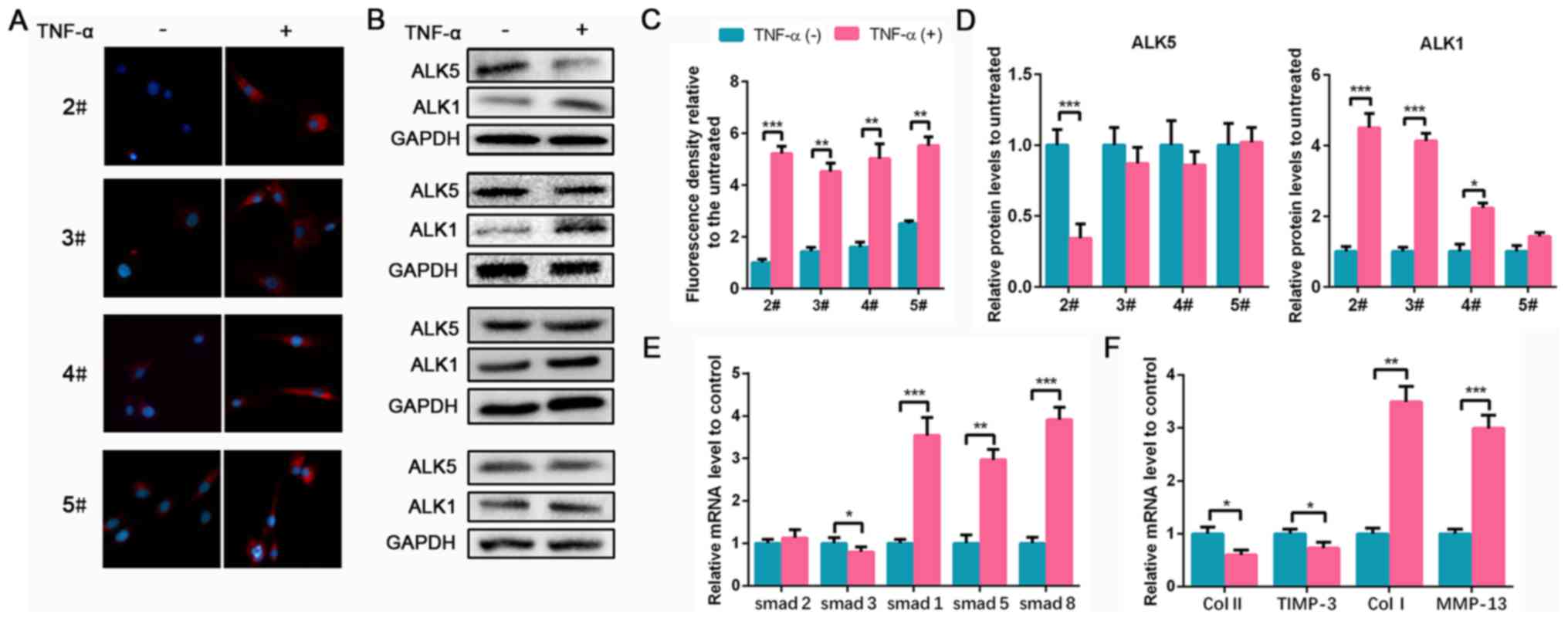 | Figure 2TGF-β1 is upregulated in TNF-α-treated
NPCs. NPCs isolated from specimens in groups 2# to 5# were treated
with 50 ng/ml TNF-α for 24 h. (A) TGF-β1 IF staining of NPCs from
groups 2# to 5# (magnification, x400). (B) Protein expression of
ALK5 and ALK1 in NPCs from groups 2# to 5# was determined by WB.
(C) The quantification of IF. (D) The semi-quantification of WB. (E
and F) The mRNA expression levels of smad1/2/3/5/8, Col II, TIMP-3,
Col I and MMP-13 in NPCs from group 2# were determined using
reverse transcription-quantitative PCR. Data are presented as the
mean ± SD of three independent experiments. *P<0.05,
**P<0.01, ***P<0.001. TGF-β1,
transforming growth factor β1; TNF-α, tumor necrosis factor α; NPC,
nucleus pulposus cell; IF, immunofluorescence; ALK, ALK tyrosine
kinase receptor; WB, western blotting; Col, collagen; TIMP-3,
tissue inhibitor of metalloproteinase-3; MMP-13, matrix
metalloproteinase 13. |
ALK5 suppression aggravates high-dose
TGF-β1-induced NPC degeneration
Due to the upregulation of TGF-β1 in the degenerated
NP tissues, high-dose TGF-β1 was used to stimulate the NPCs, and
the ALK5 inhibitor was also used to explore the effect of ALK5 on
NPC degeneration. As shown in Fig.
3A and B, exogenous TGF-β1
treatment significantly increased TGF-β1 expression in NPCs,
whereas the ALK5 inhibitor SB did not affect TGF-β1 expression. To
evaluate the effect of TGF-β1 treatment on NPCs and the suppressed
efficiency of SB, ALK5 protein expression in NPCs was evaluated.
Results showed that TGF-β1 stimulation upregulated ALK5 expression
and the supplement of SB was sufficient to inhibit ALK5 protein
expression compared with the control group. ALK1 protein expression
was also analyzed. TGF-β1 promoted ALK1 expression as well;
however, SB did not affect ALK1 levels (Fig. 3C and D). Additionally, it was demonstrated that
TGF-β1 increased Smad1/2/3/5/8 mRNA expression. However, SB
significantly downregulated Smad2 and Smad3 expression with or
without the presence of TGF-β1 stimulation. The mRNA levels of
Smad1, Smad5 and Smad8 increased following TGF-β1 stimulation,
however, SB did not affect their expression (Fig. 3E). High-dose TGF-β1 decreased
collagen II expression, and increased TIMP-3, collagen I and MMP-13
expression compared with the control (Fig. 3F). ALK5 plays a positive role in the
mediation of cell viability by regulating the Smad2/3 pathway
(20). Following inhibition of ALK5
expression, collagen II expression was further decreased compared
with the TGF-β1 treated group. TIMP-3 expression also decreased
following suppression of ALK5, whereas no significant differences
were found in collagen I and MMP-13 mRNA expression (Fig. 3F). The results suggested that
high-dose TGF-β1 promoted degeneration of NPCs via suppression of
collagen II and the promotion of collagen I and MMP-13 expression,
whereas ALK5 inhibition aggravated the reduction of collagen II and
decreased TIMP-3 expression.
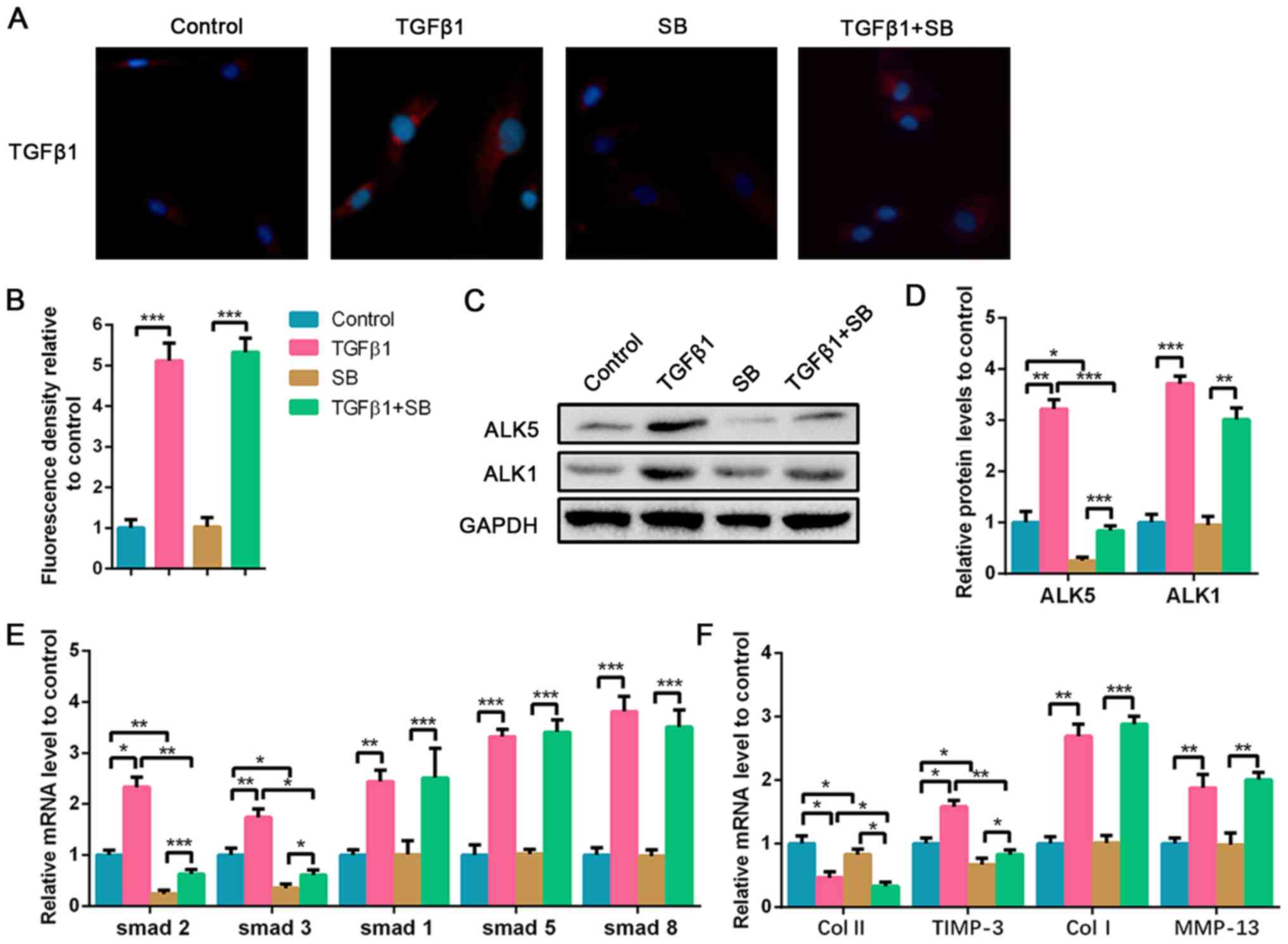 | Figure 3Inhibition of ALK5 aggravates
high-dose TGF-β1-induced degeneration of NPCs. NPCs were treated
with 5 nM TGF-β1 or 100 nM SB for 72 h. For the co-treated group,
NPCs were treated with 5 nM TGF-β1 combined with 100 nM SB for 72
h. (A) Immunofluorescence staining of TGF-β1 (magnification, x400)
and (B) quantification analysis. (C) Protein expression of ALK5 and
ALK1 was determined by western blotting and (D) semi-quantified. (E
and F) The mRNA expression levels of smad1/2/3/5/8, Col II, TIMP-3,
Col I and MMP-13 in NPCs were determined using reverse
transcription-quantitative PCR. Data are presented as the mean ± SD
of three independent experiments. *P<0.05,
**P<0.01, ***P<0.001. TGF-β1,
transforming growth factor β1; NPC, nucleus pulposus cell; ALK, ALK
tyrosine kinase receptor; Col, collagen; TIMP-3, tissue inhibitor
of metalloproteinase-3; MMP-13, matrix metalloproteinase 13; SB,
SB525334. |
ALK1 suppression reverses high-dose
TGF-β1-induced NPC degeneration
To explore the effect of ALK1 in high-dose
TGF-β1-treated NPCs, ALK1 inhibitor was co-cultured with TGF-β1. As
aforementioned, exogenous TGF-β1 upregulated TGF-β1 expression in
NPCs, which was not affected by San supplement (Fig. 4A and B). WB results indicated that San
significantly decreased ALK1 protein expression either with or
without TGF-β1 treatment. ALK5 protein was upregulated following
TGF-β1 treatment and was not affected by San (Fig. 4C and D). As an ALK1 suppressor, San treatment
also led to the suppression of Smad1, Smad5 and Smad8, but had no
effect on Smad2 and Smad3 (Fig. 4E).
Additionally, ALK1 suppression reversed high-dose TGF-β1-induced
NPC degeneration via the upregulation of collagen II and
suppression of collagen I and MMP-13 mRNA expression compared with
the TGF-β1 treated group (Fig. 4F).
Therefore, it was hypothesized that high-dose TGF-β1 degraded NPCs
via the activation of ALK1, which subsequently leads to the
upregulation of Smad1/5/8 expression.
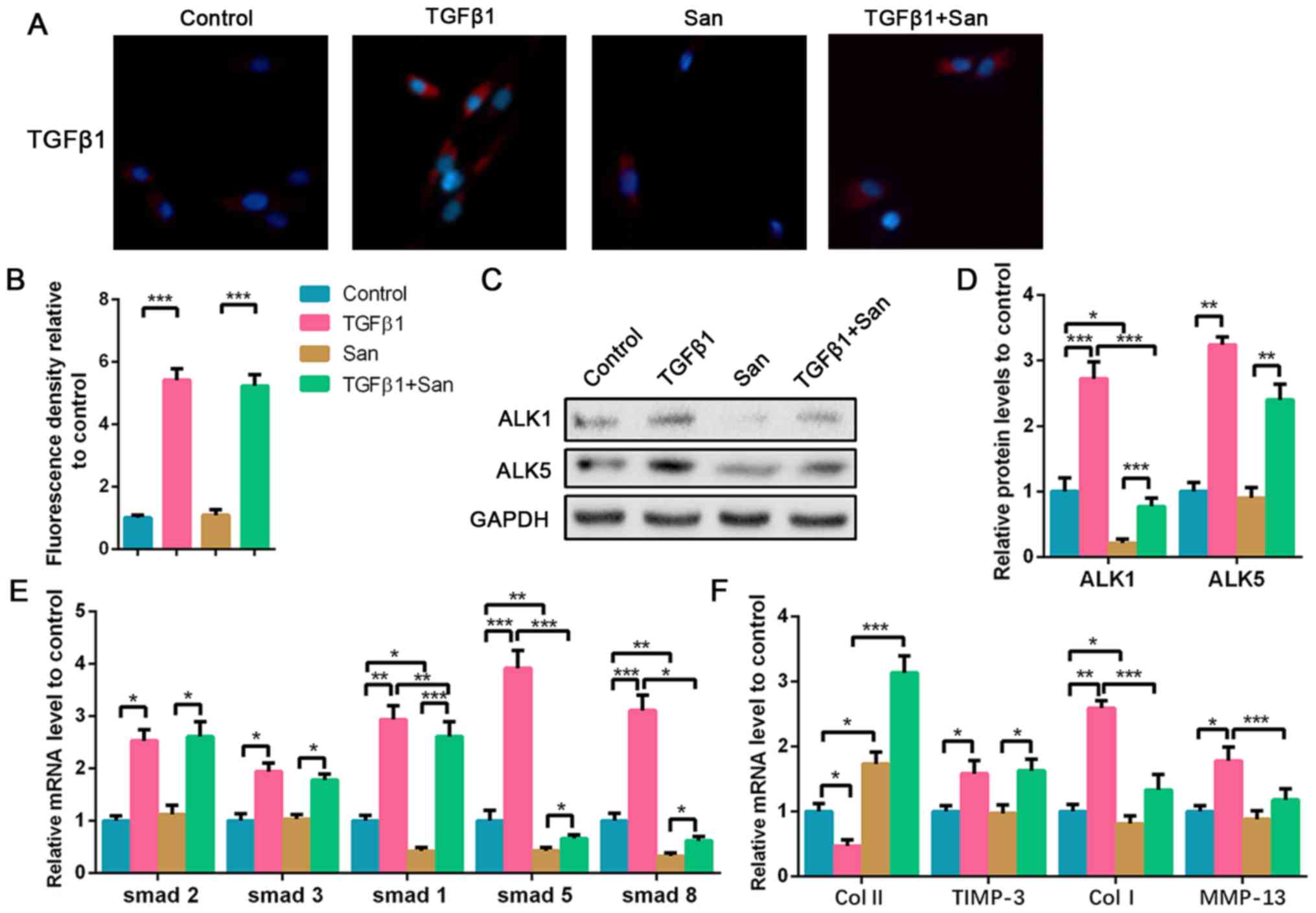 | Figure 4Inhibition of ALK1 alleviates
high-dose TGF-β1-induced NPC degeneration. NPCs were treated with 5
nM TGF-β1 or 100 nM San for 72 h. For the co-treated group, NPCs
were treated with 5 nM TGF-β1 combined with 100 nM San for 72 h.
(A) Immunofluorescence staining of TGF-β1 (magnification, x400) and
(B) quantification analysis. (C) Protein expression of ALK5 and
ALK1 was determined by western blotting and (D) semi-quantified. (E
and F) The mRNA expression levels of smad1/2/3/5/8, Col II, TIMP-3,
Col I and MMP-13 in NPCs were determined using reverse
transcription-quantitative PCR. Data are presented as the mean ± SD
of three independent experiments. *P<0.05,
**P<0.01, ***P<0.001. TGF-β1,
transforming growth factor β1; NPC, nucleus pulposus cell; ALK, ALK
tyrosine kinase receptor; Col, collagen; TIMP-3, tissue inhibitor
of metalloproteinase-3; MMP-13, matrix metalloproteinase 13; San,
San 78-130. |
Discussion
The causes of IDD have not yet been fully
elucidated, but the fundamental pathological changes in the process
of degeneration are now generally evident. The main manifestations
are the gradual reduction of normal NPCs, inflammatory cell
infiltration, ECM and water loss, secondary intervertebral disc
fibrosis and endplate calcification (21). The TGF-β pathway is requisite for the
physiological growth and development of the intervertebral disc
(22,23). However, TGF-β and its receptors
perform different biological functions in various tissues. The
level of secretion, and the distribution and expression of
receptors also play an essential role in their function. TGF-β1 can
promote ECM synthesis in the early stages of disc degeneration, and
repair and protect the disc. Yang et al (24) found that TGF-β1 prevented the
overexpression of MMP-3 caused by TNF-α in NPCs. Additionally,
TGF-β1 exerted anti-inflammatory effects by inhibition of the NF-κB
pathway and promoted collagen II and aggrecan expression in
degraded intervertebral discs (25).
However, the accumulation of TGF-β1 increases the risk of disc
degeneration as the disease develops, accelerating the process of
degeneration in the middle and late stages of the degeneration
(26). In the present study, TGF-β1
was widely expressed both in the highest degenerated NP tissue and
in TNF-α-induced degenerated NPCs in vitro, indicating that
TGF-β1 is associated with the development of NP degeneration.
The present study found that TGF-β1 accumulates in
degenerated NP and confirmed that high-dose TGF-β1 contributed to
the degenerated phenotype of NPCs, with an upregulation in ALK1/5
and Smad1/2/3/5/8. It was reported that TGF-β upregulation can
increase the activation of type I receptors (ALK5 or ALK1), and
further activate Smad2/3 or Smad1/5/8(27). Kwon et al (28) also illustrated that both Smad2/3 and
Smad1/5/8 were upregulated in degenerative bovine NPCs, and
activated Smad1/5/8 could inhibit Smad2/3 expression, leading to
further IDD. Accumulating evidence has indicated that TGF-β can
stimulate ECM production via Smad2/3 overexpression (29,30).
TIMP-3 has a specific protective effect on cartilage, and the
TGF-β/Smad2/3 pathway can enhance the expression of TIMP-3(31), thereby explaining the protective
effect on the ECM. However, activated ALK1 can mediate Smad1/5/8
phosphorylation and has the opposite effect in numerous tissues
(32,33). The TGF-β/Smad1/5/8 signaling pathway
produces the mineralization marker MMP-13 and causes chondrocyte
hypertrophy (34). Hence,
suppression of ALK1 and the downstream downregulation of Smad1/5/8
could exert a positive effect on NPCs. Most tissue and organ
fibrotic diseases where the pathological basis includes the
excessive deposition of collagen I are closely related to the
overexpression of TGF-β (35).
Therefore, the present study observed high-dose TGF-β1-induced
overexpression of collagen I, which was also inhibited by the
suppression of ALK1.
In summary, the present study elucidated the
degenerative effect of high-dose TGF-β1 on NPCs, which is mainly
associated with the activation of ALK1 and Smad1/5/8 expression.
High-dose TGF-β1 caused an increase in ALK1 and ALK5 expression,
but the adverse effect of ALK1 was more pronounced than the
protective effect of ALK5. The combination of high-dose TGF-β1 and
ALK1 inhibitor had a robust protective impact on ECM stability. The
present study provided a novel basis for future research on the
association between these two. However, as it was difficult for us
to obtain specimens without any degermation, no healthy disc
samples were obtained from individuals without IDD, which is a
limitation of the present study. Therefore, the relationship
between TGF-β and IDD remains unclear, so we plan to confirm the
experimental results in animals in the future so that we can have a
control group without degeneration.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZQ and YS designed the study and performed the
experiments, ZQ and FZ collected the data, WC and TL analyzed the
data, ZQ and YS prepared the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
The Second Affiliated Hospital of Soochow University (approval no.
AK2017-7K8J). Written informed consent was obtained from the
patients and/or guardians.
Patient consent for publication
Patients or their guardians have provided written
informed consents for publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhao CQ, Wang LM, Jiang LS and Dai LY: The
cell biology of intervertebral disc aging and degeneration. Ageing
Res Rev. 6:247–261. 2007.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhou TY, Wu YG, Zhang YZ, Bao YW and Zhao
Y: SIRT3 retards intervertebral disc degeneration by anti-oxidative
stress by activating the SIRT3/FOXO3/SOD2 signaling pathway. Eur
Rev Med Pharmacol Sci. 23:9180–9188. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Sampara P, Banala RR, Vemuri SK, Av GR and
Gpv S: Understanding the molecular biology of intervertebral disc
degeneration and potential gene therapy strategies for
regeneration: A review. Gene Ther. 25:67–82. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Vo NV, Hartman RA, Patil PR, Risbud MV,
Kletsas D, Iatridis JC, Hoyland JA, Le Maitre CL, Sowa GA and Kang
JD: Molecular mechanisms of biological aging in intervertebral
discs. J Orthop Res. 34:1289–1306. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ashley JW, Enomoto-Iwamoto M, Smith LJ,
Mauck RL, Chan D, Lee J, Heyworth MF, An H and Zhang Y:
Intervertebral disc development and disease-related genetic
polymorphisms. Genes Dis. 3:171–177. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Wu M, Chen G and Li YP: TGF-β and BMP
signaling in osteoblast, skeletal development, and bone formation,
homeostasis and disease. Bone Res. 4(16009)2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yang Y, He X, Li Y, Feng J, Pang H, Wang J
and Liu Q: Association of transforming growth factor-β1 with
pathological grading of intervertebral disc degeneration. Nan Fang
Yi Ke Da Xue Xue Bao. 32:897–900. 2012.(In Chinese).
|
|
8
|
Tolonen J, Gronblad M, Vanharanta H, Virri
J, Guyer RD, Rytomaa T and Karaharju EO: Growth factor expression
in degenerated intervertebral disc tissue. An immunohistochemical
analysis of transforming growth factor beta, fibroblast growth
factor and platelet-derived growth factor. Eur Spine J. 15:588–596.
2006.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Tolonen J, Gronblad M, Virri J, Seitsalo
S, Rytomaa T and Karaharju E: Transforming growth factor beta
receptor induction in herniated intervertebral disc tissue: An
immunohistochemical study. Eur Spine J. 10:172–176. 2001.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen S, Liu S, Ma K, Zhao L, Lin H and
Shao Z: TGF-β signaling in intervertebral disc health and disease.
Osteoarthritis Cartilage. 27:1109–1117. 2019.
|
|
11
|
Uchiyama Y, Guttapalli A, Gajghate S,
Mochida J, Shapiro IM and Risbud MV: SMAD3 functions as a
transcriptional repressor of acid-sensing ion channel 3 (ASIC3) in
nucleus pulposus cells of the intervertebral disc. J Bone Miner
Res. 23:1619–1628. 2008.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tian Y, Yuan W, Li J, Wang H, Hunt MG, Liu
C, Shapiro IM and Risbud MV: TGFβ regulates Galectin-3 expression
through canonical Smad3 signaling pathway in nucleus pulposus
cells: Implications in intervertebral disc degeneration. Matrix
Biol. 50:39–52. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Colombier P, Clouet J, Boyer C, Ruel M,
Bonin G, Lesoeur J, Moreau A, Fellah BH, Weiss P, Lescaudron L, et
al: TGF-β1 and GDF5 act synergistically to drive the
differentiation of human adipose stromal cells toward nucleus
pulposus-like cells. Stem Cells. 34:653–667. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Griffith JF, Wang YX, Antonio GE, Choi KC,
Yu A, Ahuja AT and Leung PC: Modified Pfirrmann grading system for
lumbar intervertebral disc degeneration. Spine (Phila Pa 1976).
32:E708–E712. 2007.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ten DP, Yamashita H, Ichijo H, Franzen P,
Laiho M, Miyazono K and Heldin CH: Characterization of type I
receptors for transforming growth factor-beta and activin. Science.
264:101–104. 1994.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Patil P, Niedernhofer LJ, Robbins PD, Lee
J, Sowa G and Vo N: Cellular senescence in intervertebral disc
aging and degeneration. Curr Mol Biol Rep. 4:180–190.
2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
van der Kraan PM, Blaney DE, Blom A and
van den Berg WB: TGF-beta signaling in chondrocyte terminal
differentiation and osteoarthritis: Modulation and integration of
signaling pathways through receptor-Smads. Osteoarthritis
Cartilage. 17:1539–1545. 2009.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang XH, Hong X, Zhu L, Wang YT, Bao JP,
Liu L, Wang F and Wu XT: Tumor necrosis factor alpha promotes the
proliferation of human nucleus pulposus cells via nuclear
factor-κB, c-Jun N-terminal kinase, and p38 mitogen-activated
protein kinase. Exp Biol Med (Maywood). 240:411–417.
2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhu Y, Tao H, Jin C, Liu Y, Lu X, Hu X and
Wang X: Transforming growth factor-beta1 induces type II collagen
and aggrecan expression via activation of extracellular
signal-regulated kinase 1/2 and Smad2/3 signaling pathways. Mol Med
Rep. 12:5573–5579. 2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Roberts S, Evans H, Trivedi J and Menage
J: Histology and pathology of the human intervertebral disc. J Bone
Joint Surg Am. 88 (Suppl 2):S10–S14. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Peck SH, McKee KK, Tobias JW, Malhotra NR,
Harfe BD and Smith LJ: Whole Transcriptome analysis of
notochord-derived cells during embryonic formation of the nucleus
pulposus. Sci Rep. 7(10504)2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jin H, Shen J, Wang B, Wang M, Shu B and
Chen D: TGF-β signaling plays an essential role in the growth and
maintenance of intervertebral disc tissue. FEBS Lett.
585:1209–1215. 2011.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yang H, Gao F, Li X, Wang J, Liu H and
Zheng Z: TGF-β1 antagonizes TNF-α induced up-regulation of matrix
metalloproteinase 3 in nucleus pulposus cells: Role of the ERK1/2
pathway. Connect Tissue Res. 56:461–468. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Yang H, Cao C, Wu C, Yuan C, Gu Q, Shi Q
and Zou J: TGF-β1 suppresses inflammation in cell therapy for
intervertebral disc degeneration. Sci Rep. 5(13254)2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Singh K, Masuda K, Thonar EJ, An HS and
Cs-Szabo G: Age-related changes in the extracellular matrix of
nucleus pulposus and anulus fibrosus of human intervertebral disc.
Spine (Phila Pa 1976). 34:10–16. 2009.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Mehra A and Wrana JL: TGF-beta and the
Smad signal transduction pathway. Biochem Cell Biol. 80:605–622.
2002.PubMed/NCBI View
Article : Google Scholar
|
|
28
|
Kwon YJ, Lee JW, Moon EJ, Chung YG, Kim OS
and Kim HJ: Anabolic effects of Peniel. 2000, a peptide that
regulates TGF-beta1 signaling on intervertebral disc degeneration.
Spine (Phila Pa 1976). 38:E49–E58. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Hu B, Xu C, Cao P, Tian Y, Zhang Y, Shi C,
Xu J, Yuan W and Chen H: TGF-β stimulates expression of chondroitin
polymerizing factor in nucleus pulposus cells through the Smad3,
RhoA/ROCK1, and MAPK signaling pathways. J Cell Biochem.
119:566–579. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chen MH, Sun JS, Liao SY, Tai PA, Li TC
and Chen MH: Low-intensity pulsed ultrasound stimulates matrix
metabolism of human annulus fibrosus cells mediated by transforming
growth factor β1 and extracellular signal-regulated kinase pathway.
Connect Tissue Res. 56:219–227. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wang X, Zhu Y, Tao H, Jin C, Liu Y, Lu X,
Hu X and Fan C: Interaction of ERK1/2 and Smad2/3 signaling
pathways in TGF-β1-induced TIMP-3 expression in rat chondrocytes.
Arch Biochem Biophys. 564:229–236. 2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Van Caam A, Madej W, Garcia DVA, Goumans
MJ, Ten DP, Blaney DE and van der Kraan P: TGFβ1-induced SMAD2/3
and SMAD1/5 phosphorylation are both ALK5-kinase-dependent in
primary chondrocytes and mediated by TAK1 kinase activity.
Arthritis Res Ther. 19(112)2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Derynck R and Budi EH: Specificity,
versatility, and control of TGF-β family signaling. Sci Signal.
12(eaav5183)2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Mariani E, Pulsatelli L and Facchini A:
Signaling pathways in cartilage repair. Int J Mol Sci.
15:8667–8698. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Brenner DA, Rippe RA, Rhodes K, Trotter JF
and Breindl M: Fibrogenesis and type I collagen gene regulation. J
Lab Clin Med. 124:755–760. 1994.PubMed/NCBI
|