Introduction
Furocoumarins are biologically potent organic
compounds occurring in various traditional herbal medicines and
foods, particularly those belonging to the Umbelliferae and
Rutaceae families (1,2). Xanthotoxin (8-methoxypsoralen) is an
organic substance prevalent in various edible plants. It may be
used in combination with long-wavelength ultraviolet (UV) light as
a treatment for psoriasis, vitiligo and T-cell lymphoma (3). It is a photosensitive substance mostly
used in clinical psoralen UV A (PUVA) therapy for vitiligo,
psoriasis, as well as other skin conditions. PUVA is a UV light
(320-360 nm) therapy for skin diseases, using the sensitizing
effects of the drug psoralen. Previous studies have reported the
modulation of various cytokines and the decontamination of platelet
concentrates by methoxypsoralen and long-wavelength UV radiation
(4,5).
Cytochrome P450 (CYP450) enzymes, which are abundant
in the liver, are responsible for promoting the metabolic
stimulation of a sizable number of xenobiotic compounds. However,
under certain conditions, CYP450 catalysis is able to bring about
reactive species that bind to and inhibit CYP450, and the complex
attaches to other cellular proteins which may lead to cell death
(6). Xanthotoxin is a substance with
a recognized ability to suppress rat and mouse CYP450-mediated
activities (7-11)
and is known as a time-dependent inhibitor of various human
CYP450-mediated activities (12).
Certain studies have suggested that xanthotoxin is able to function
as an efficient mechanism-based inhibitor of CYP2A6 activities,
which discriminatorily incapacitates this enzyme in human liver
microsomes (HLM) when utilized at lower concentrations and for
limited exposure durations (13,14).
In addition, since animal microsomes are routinely
employed for determining metabolic patterns in humans, it is
imperative to identify inter-species differences between humans and
animals with regard to metabolic activities mediated by CYP450
(15-17).
Therefore, the present study sought to identify the similarities
and differences in xanthotoxin metabolism in liver microsomes of 7
mammalian species, including HLM, Rhesus monkey liver microsomes
(RMLM), cynomolgus monkey liver microsomes (CMLM), Sprague Dawley
rat liver microsomes (RLM), mouse liver microsomes (MLM), Dunkin
Hartley guinea pig liver microsomes (PLM) and Beagle dog liver
microsomes (DLM). To date, at least to the best of our knowledge,
no studies are available on the differences in xanthotoxin
metabolites in humans and other mammalian liver microsomes. The
present study therefore aimed to provide insight into metabolic
functions with regard to xanthotoxin and to identify specific
differences among species. The present results may aid future
investigations on xanthotoxin, which may prove to be beneficial to
humans in the future.
Materials and methods
Materials and testing agents
Xanthotoxin was obtained from The Chinese National
Institute for the Control of Pharmaceutical and Biological
Products. NADPH,
KH2PO4-K2HPO4 buffer
and DMSO were purchased from Sigma-Aldrich (Merck KGaA). Human,
Rhesus and Cynomolgus monkey, Sprague Dawley rat, mouse, Dunkin
Hartley guinea pig and Beagle dog microsomes were obtained from the
Research Institute for Liver Diseases Shanghai, Co., Ltd. Liver
microsomes were stored at -80˚C. High-performance liquid
chromatography (HPLC) was used to separate, identify and quantify
all mixture components.
Ultra-high performance liquid
chromatography/quadrupole time-of-flight mass spectrometric
analysis (UPLC/Q-TOF MS)
The UPLC/Q-TOF MS procedure was performed using an
Agilent 6520 Q-TOF LC/MS system and MassHunter Workstation (Agilent
Technologies Inc.). An Extend-C18 column (1.8 µm, 2.1x50 mm) was
used. The elution gradient consisted of mobile phase A (0.1%
aqueous formic acid) and B (100% methanol). The UPLC gradient
curriculum employed was thus motile phase A, which was maintained
at 90% for a duration of 3 min, and thence a linearly set up
gradient lowered moving phase A out of 90% to become 10% at the end
of 14 min and maintained at 10% through a time of 3 min. This was
finally increased to 90% A to equilibrate the column. The column
temperature was maintained at 30˚C together with the flow rate kept
at 0.2 ml/min. Acquisition of mass was performed in the duo of
positive ion mode [electron spray ionization (ESI)+] and
negative ion mode (ESI-) from a mass-to-charge ratio
(m/z) of 80 towards 1,000 Da, making use of a source temperature of
100˚C, a desolvation temperature of 350˚C and a dissolution gas
flow standing at 600 l/h.
Metabolism of xanthotoxin in liver
microsomes
Xanthotoxin (final concentration, 50 µM) was
dissolved in DMSO (final concentration <0.5%) and diluted (PBS;
0.1 m, pH 7.4). For each of the 7 species individually, the liver
microsomes (50 µl; final concentration, 2 mg/ml) were mixed with
standard solution (50 µl) and pre-incubated for 5 min at 37˚C. The
above reaction mixture was added to NADPH solution (100 µl; final
concentration, 1 mM), which was diluted in PBS (0.1 M; pH 7.4). A
total of 400 µl ice-cold methanol was utilized to terminate the
reaction at the end of the 120-min incubation at a temperature of
37˚C. Following centrifugation at 10,000 x g for a total duration
of 10 min at 4˚C, the supernatant was filtered through a 0.22-µm
filter and was collected for analysis by UPLC/Q-TOF MS. A control
reaction was set up without xanthotoxin. An aliquot of 5 µl of the
solution was analyzed by UPLC/Q-TOF MS and HPLC/MS/MS.
Statistical analysis
The content of each metabolite was compared
according to the peak area of the metabolite. Values are expressed
as the mean ± standard deviation of three samples. One-way ANOVA
with the post hoc Student-Newman-Keuls test was performed to
confirm inter-group differences with SPSS 11.5 software (SPSS
Inc.). P<0.05 was considered to indicate statistical
significance.
Results
Q-TOF MS analysis of xanthotoxin
metabolites
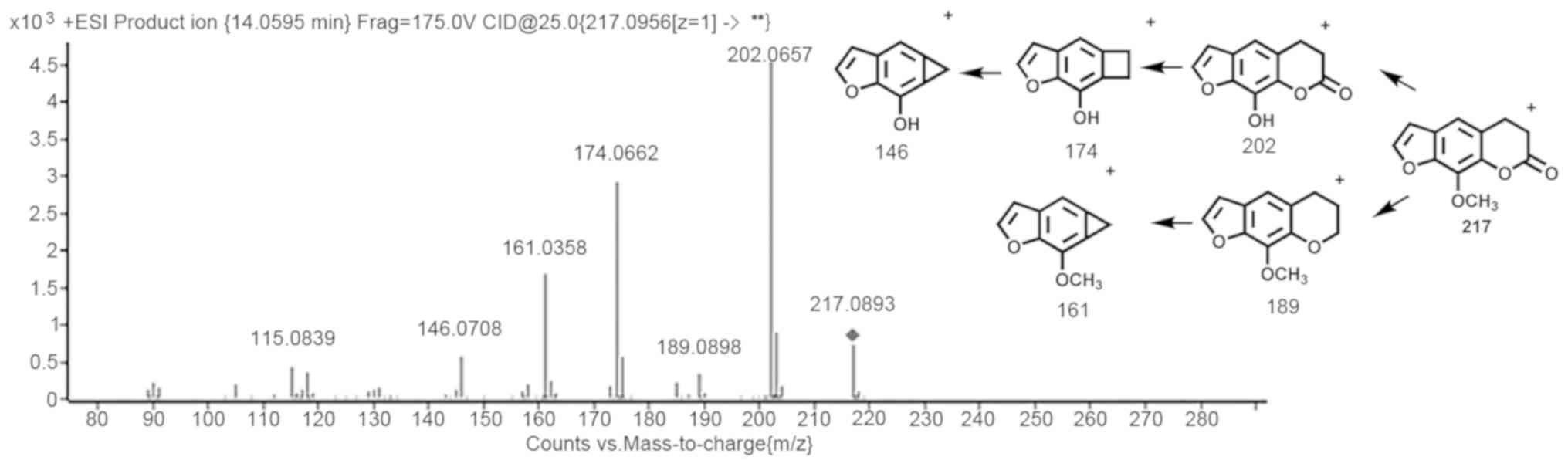
Xanthotoxin had a retention time (Rt) of 14.0595 min
and in ESI+ mode, the protonated molecule
[M+H]+ exhibited a peak at m/z 217.0956. For
xanthotoxin, the noteworthy fragment ions at m/z 202.0657,
189.0898, 174.0662, 161.0958 and 146.0708 in the collision energy
25 eV mass spectrum were generated by means of the loss of
-CH3 and small neutral molecules, e.g. CO. Q-TOF mass
spectra and xanthotoxin metabolite and fragment information are
provided in Fig. 1.
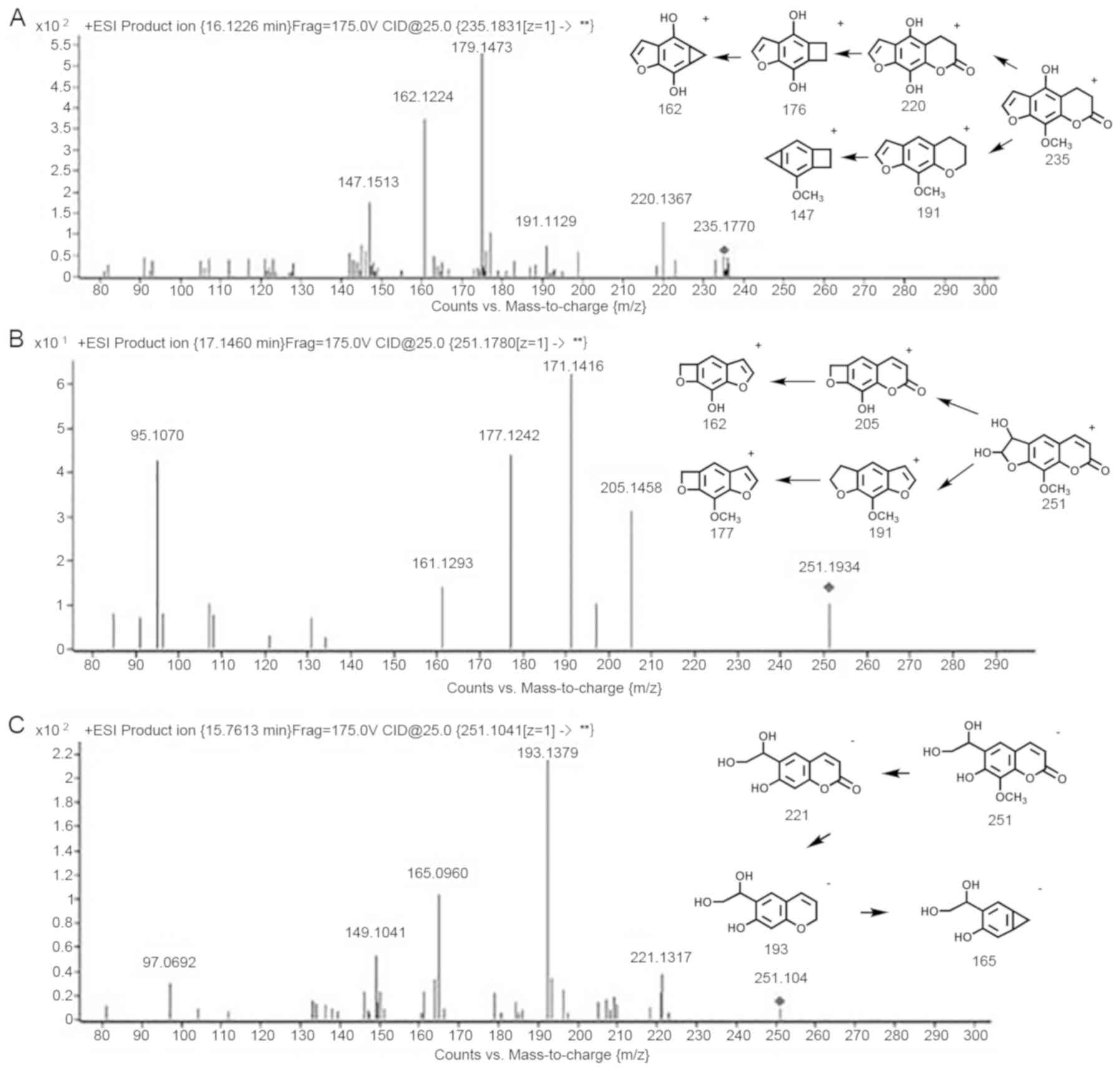
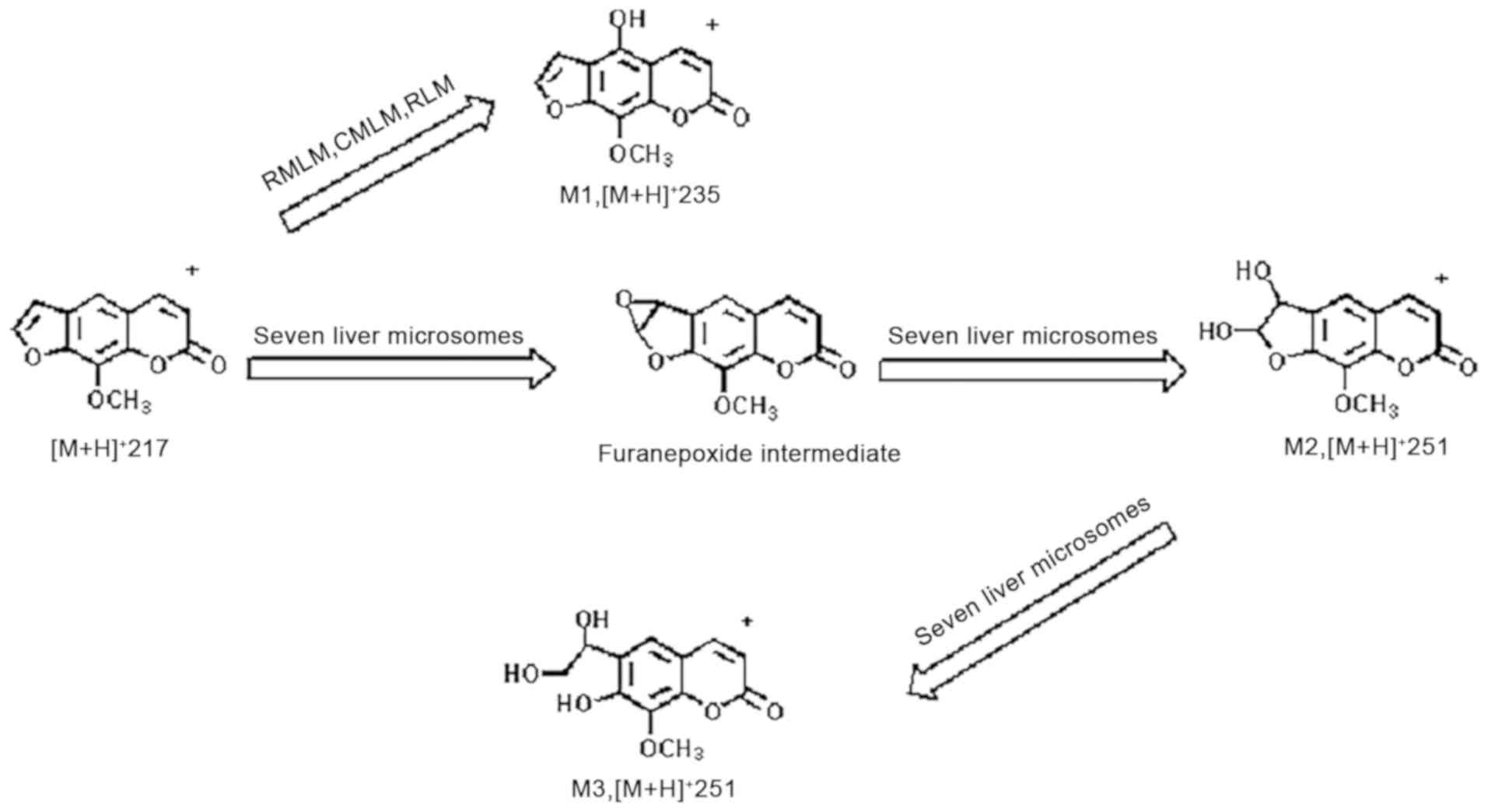
The metabolization of xanthotoxin comprises the
hydroxylation of the phenyl ring, hydrogenation and hydrolysis of
the lactone ring (18,19). Oxidation of the furan ring produces a
furanoepoxide or γ-ketoenal intermediate. Furanoepoxide then forms
a dihydrodiol, while γ-ketoenal is converted to hydroxy coumaryl
acetic acid. The structures and formation of metabolites of
xanthotoxin are presented in detail in Fig. 2. M1-3 are metabolizations of
xanthotoxin detected in RMLM, CMLM and RLM, MLM, HLM, PLM and DLM.
The metabolites M1 (Fig. 2A)
included protonated molecules [M+H]+ in the vicinity of
m/z 235.1831. Meticulous mass analysis revealed the chemical
formula of C12H10O5, propounding
the hydroxylation and hydrogenation of xanthotoxin. The steep
energy mass spectra of M1 revealed a major product ion with m/z
176.1473 and the major fragment ion was observed at m/z 162.1224.
The indicated remnant ions suggested that the alteration took place
at the site C5 of xanthotoxin.
The metabolites M2 (Fig.
2B) included the molecule [M+H]+ at m/z 251.1780
along with [M-H]- at m/z 249.1275. The fragment ions
suggested that xanthotoxin is transformed into an epoxide
intermediate that reacts to become a dihydrodiol (M2). This result
is in agreement with that of an earlier study (20). The deprivation of H2O and
the ensuing deficiency of CO leads to the formation of the compound
with m/z 205 and corroborates the structure of M2.
The metabolite M3 (Fig.
2C), characterized by [M-H]- with m/z 251.1041, is
in accordance with a hydroxylation on the furan ring, and is also
an oxidation product of the dihydrodiol (M2). The fragments include
ions at m/z 193.1379, 221.1317 and 165.0960.
Identification of the types and amount
of metabolites in liver microsomes from various species
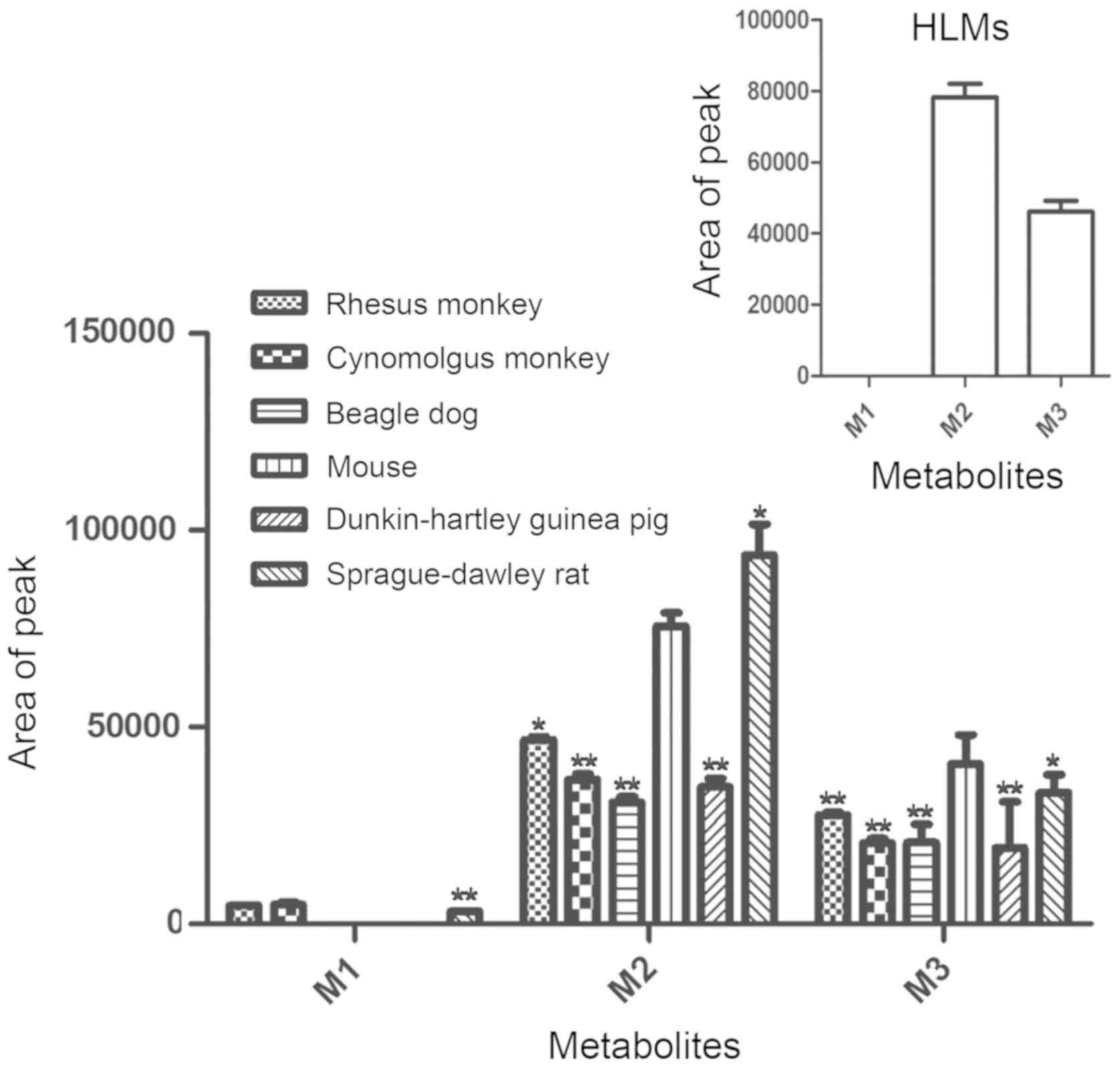
To identify probable metabolic avenues of
xanthotoxin, the metabolites obtained from 7 liver microsome
incubations were initially analyzed by UPLC/Q-TOF MS. A total of 3
metabolites were detected in RMLM, CMLM and RLM, two metabolites
were observed in MLM, HLM, PLM and DLM. The array of the
metabolites may be contradistinguished by MS and Rt.
As presented in Fig.
3, regarding the amount of metabolites, the metabolites M2 and
M3 in MLM were closest to those in HLM.
Table I displays the
chemical formulas, authentic masses and Rts of these metabolites of
xanthotoxin in the liver microsomes of the 7 species. A total of 3
metabolites of xanthotoxin were detected in RMLM, CMLM and RLM. A
duo of metabolites were identified in DLM, PLM, HLM and MLM. The
peaks of ion current chromatograms of the metabolite with m/z 250
in ES+ and ES- modes for the liver microsomes
from all species.
 | Table IIdentification of xanthotoxin
metabolites in liver microsomes. |
Table I
Identification of xanthotoxin
metabolites in liver microsomes.
| | Rt (min) |
|---|
| Metabolites | Molecular
formula | HLM | RMLM | CMLM | RLM | MLM | PLM | DLM |
|---|
| M1 |
C12H10O5 | - | 16.1036 | 16.0826 | 16.1295 | - | - | - |
| M2 |
C12H10O6 | 17.1753 | 17.2637 | 17.1670 | 17.1411 | 17.2245 | 17.2146 | 17.1331 |
| M3 |
C12H12O6 | 15.7499 | 15.8695 | 15.7310 | 15.6837 | 15.6819 | 15.7781 | 15.8149 |
| Number | - | 2 | 3 | 3 | 3 | 2 | 2 | 2 |
Discussion
At present, multitudinous herbal medicines are used
worldwide. Various Chinese traditional medicines and plants have
been approved for nutritional and medical purposes (21,22).
Traditional Chinese Medicine has stood the test of time for
thousands of years and has a long history of usage. Rhizoma ginseng
and Radix Angelicae dahuricae are all medicinal plants
belonging to the plant family of Umbelliferae, which are widely
used in Traditional Chinese Medicine. Xanthotoxin is a medicinal
component of furocoumarins derived from Umbelliferae plants. It is
widely used in Traditional Chinese Medicine, including the Chinese
patent drug Xiaoyan Xuanshi ointment. Xanthotoxin is widely used in
clinical practice for conditions including psoriasis and vitiligo
(4,5). Clinical studies have reported on side
effects of xanthotoxin (23-25).
The present study provided information on the metabolic behavior
and routes of xanthotoxin in liver microsomes of 7 species for the
first time, to the best of our knowledge. This is useful for
investigating the metabolic reactions of xanthotoxin. It provides
an important basis for further research and developments. The
present study elucidated the biotransformation/metabolism of
xanthotoxin, which may have far-reaching and important effects;
this knowledge may contribute to the rational and safe use of
traditional Chinese and western medicine in the future.
It has been indicated that certain dietary
supplements, including grapefruit and hop tea, influence CYP
activity and may affect the pharmacokinetics of drugs (26). The effects of grapefruit juice and
its inhibitory effects against CYP3A4 have been investigated.
Studies have revealed that components of grapefruit juice inhibit
CYP3A4 and alter the absorption and metabolism of midazolam
(27), vinblastine (28) and digoxin (29), in vitro as well as in
vivo. Furanocoumarin derivatives in grapefruit have been
suggested to exert an inhibitory effect on CYPs (30). Previous studies have suggested that
xanthotoxin is an influential mechanism-based inhibitor of CYP2A6
activity and immobilizes this enzyme in HLM when applied at low
concentrations for short exposure times (31). Another study investigated the
reactive metabolites of xanthotoxin as the epoxide intermediate,
demonstrating that CYP450 enzymes are responsible for the metabolic
activity of xanthotoxin (20). These
studies have provided pertinent knowledge for the prudent usage of
xanthotoxin. However, extensive data on the metabolites of
xanthotoxin, particularly the analogy of metabolic traits amidst
liver microsomes from divergent mammalian species, are currently
limited, at least to the best of our knowledge. The scope of the
present study was geared towards obtaining definitive knowledge
regarding the comparison of metabolites and metabolism in liver
microsomes among mammalian species.
UPLC/Q-TOF MS revealed that the metabolic behavior
of xanthotoxin comprises diverse chemical modifications to form a
multitude of metabolites that may have various biological
functions. Fig. 4 illustrates the
suggested metabolic routes of xanthotoxin in the liver microsomes
of the 7 species.
Previous studies focusing on the metabolism of
furanocoumarin compounds have disclosed that the transformations
include hydroxylation of the phenyl ring (32), hydrogenation of unsaturated lactone
ring and hydrolyzation of the lactonic ring (33). However, the production of
furanoepoxide/γ-ketoenal via oxidation of the furan ring is the
fundamental and ultimately interesting metabolic behavior of the
furanocoumarins. The metabolic transformation of furanocoumarins by
CYP450 is presented in Fig. 4.
In the present study on the liver microsomes of 7
species, dihydrodiol or hydroxy coumaryl acetic acid was indicated
to be derived from the furanoepoxide or γ-ketoenal intermediate,
respectively. These results suggest that in diverse species, the
furan ring is one utmost integral site of metabolic transformation
for furanocoumarin compounds. The present investigation on
xanthotoxin may provide a basis for metabolic research with
inclusively furan-containing compounds. It may provide important
information for further study of drug-drug interactions and
food-drug interactions.
In conclusion, in the present study, xanthotoxin and
NADPH were incubated in the liver microsomes of 7 mammalian
species, including human, Rhesus monkey, Cynomolgus monkey, Sprague
Dawley rat, mouse, Dunkin Hartley guinea pig and Beagle dog.
Identification of three metabolites of xanthotoxin was achieved.
The metabolites of xanthotoxin include a dihydrodiol and hydroxy
coumaryl acetic acid from furanoepoxide and γ-ketoenal
intermediates, respectively, originating from the oxidation of the
furan ring, hydroxylation of the phenyl ring, and hydrogenation and
hydrolysis of the unsaturated lactone ester, as well as the
lactonic ring. Umbelliferae medicinal plants, including Rhizoma
ginseng and Angelica dahurica, have been widely used in
Traditional Chinese Medicine. Furocoumarins, including psoralen and
isopsoralen, are widely used in the clinic. Previous studies by our
group have indicated that their metabolites do produce certain
serious drug interactions and toxicity through biotransformation
and other processes (34,35). Relevant documents include studies by
Yang et al (18,19). Xanthotoxin is also derived from a
medicinal plant of the Umbelliferae, and hence, it is speculated
that there may be a similar association between its metabolites.
Further studies by our group will aim to expand on the association
between the formation of these metabolites and their toxicity in
these species.
Due to inter-species differences, different animal
liver microsomes exhibit differences in their metabolites of
xanthotoxin. Only two metabolites were detected in HLM. The present
study mainly focused on the metabolic behavior of xanthotoxin in
the liver microsomes of 7 different mammalian species. The activity
of CYP450 enzymes is reflected indirectly by the metabolism of
substrate. Regarding the types of metabolites, the metabolites of
xanthotoxin in MLM, PLM and DLM were the same as those in HLM;
regarding the amount of metabolites, the metabolites M2 and M3 of
MLM were closest to those in HLM. Analysis of the type and quantity
of metabolites indicated that the metabolism of xanthotoxin in MLM
was most similar to that in HLM. The results of the present study
may provide further knowledge on the metabolic functions of
xanthotoxin in liver microsomes of different species and may prove
to be useful for future study of the metabolic mechanisms of action
of xanthotoxin. Identification of differences among species
regarding drug metabolic processes will aid in the selection of
appropriate animal models for future studies.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81430096), the Key Support
Projects of Tianjin Science and Technology (grant no.
16YFZCSY00440) and the Program for Changjiang Scholars and
Innovative Research Team in University (grant no. IRT_14R41).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL performed the liver microsome incubation
experiments, analyzed the data and wrote the manuscript. DZ and LZ
were responsible for the UPLC/Q-TOF MS experiments and analysis of
the data. XH, LW, AY and JAM designed the experiments, analyzed the
data and modified of the manuscript. XH received funding for the
project, conceived the study and performed the experiments. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wagstaff DJ: Dietary exposure to
furocoumarins. Regul Toxicol Pharmacol. 14:261–272. 1991.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Guo LQ and Yamazoe Y: Inhibition of
cytochrome P450 by furanocoumarins in grapefruit juice and herbal
medicines. Acta Pharmacol Sin. 25:129–136. 2004.PubMed/NCBI
|
|
3
|
Edelson R, Berger C, Gasparro F, Heald P,
Rook A, Perez M, Wintroub B, Knobler R and Jegasothy B: Treatment
of erythrodermic cutaneous T-cell lymphoma with extracorporeal
photochemotherapy. J Am Acad Dermatol. 27:427–433. 1992.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Corash L, Lin L and Wiesehahn G: Use of
8-methoxypsoralen and long wavelength ultraviolet radiation for
decontamination of platelet concentrates. Blood Cells. 18:57–73.
1992.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Tokura Y: Modulation of cytokine
production by 8-methoxypsoralen and UVA. J Dermatol Sci.
19:114–122. 1999.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhou S, Chan E, Duan W, Huang M and Chen
YZ: Drug bioactivation, covalent binding to target proteins and
toxicity relevance. Drug Metab Rev. 37:41–213. 2005.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Fouin-Fortunet H, Tinel M, Descatoire V,
Letteron P, Larrey D, Geneve J and Pessayre D: Inactivation of
cytochrome P-450 by the drug methoxsalen. J Pharmacol Exp Ther.
236:237–247. 1986.PubMed/NCBI
|
|
8
|
Labbe G, Descatoire V, Beaune P, Letteron
P, Larrey D and Pessayre D: Suicide inactivation of cytochrome
P-450 by methoxsalen. Evidence for the covalent binding of a
reactive intermediate to the protein moiety. J Pharmacol Exp Ther.
250:1034–1042. 1989.PubMed/NCBI
|
|
9
|
Letteron P, Descatoire V, Larrey D, Tinel
M, Geneve J and Pessayre D: Inactivation and induction of
cytochrome P-450 by various psoralen derivatives in rats. J
Pharmacol Exp Ther. 238:685–692. 1986.PubMed/NCBI
|
|
10
|
Mays DC, Hilliard JB, Wong DD and Gerber
N: Activation of 8-methoxypsoralen by cytochrome P-450. Enzyme
kinetics of covalent binding and influence of inhibitors and
inducers of drug metabolism. Biochem Pharmacol. 38:1647–1655.
1989.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Mays DC, Hilliard JB, Wong DD, Chambers
MA, Park SS, Gelboin HV and Gerber N: Bioactivation of
8-methoxypsoralen and irreversible inactivation of cytochrome P-450
in mouse liver microsomes: Modification by monoclonal antibodies,
inhibition of drug metabolism and distribution of covalent adducts.
J Pharmacol Exp Ther. 254:720–731. 1990.PubMed/NCBI
|
|
12
|
Tinel M, Belghiti J, Descatoire V, Amouyal
G, Letteron P, Geneve J, Larrey D and Pessayre D: Inactivation of
human liver cytochrome P-450 by the drug methoxsalen and other
psoralen derivatives. Biochem Pharmacol. 36:951–955.
1987.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Koenigs LL, Peter RM, Thompson SJ, Rettie
AE and Trager WF: Mechanism-based inactivation of human liver
cytochrome P450 2A6 by 8-methoxypsoralen. Drug Metab Dispos.
25:1407–1415. 1997.PubMed/NCBI
|
|
14
|
Goto T, Moriuchi H, Fu X, Ikegawa T,
Matsubara T, Chang G, Uno T, Morigaki K, Isshiki K and Imaishi H:
The effects of single nucleotide polymorphisms in CYP2A13 on
metabolism of 5-methoxypsoralen. Drug Metab Dispos. 38:2110–2116.
2010.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Martignoni M, Groothuis GM and de Kanter
R: Species differences between mouse, rat, dog, monkey and human
CYP-mediated drug metabolism, inhibition and induction. Expert Opin
Drug Metab Toxicol. 2:875–894. 2006.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Li L, Chen X, Zhou J and Zhong D: In vitro
studies on the oxidative metabolism of 20(s)-ginsenoside Rh2 in
human, monkey, dog, rat, and mouse liver microsomes, and human
liver s9. Drug Metab Dispos. 40:2041–2053. 2012.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Li P, Gong Y, Lim HK, Jian W, Edom RW,
Salter R, Silva J and Weng N: Bio-generation of stable isotope
labeled internal standards for absolute and relative quantitation
of drug metabolites in plasma samples by LC-MS/MS. J Chromatogr B
Analyt Technol Biomed Life Sci. 926:92–100. 2013.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yang AH, He X, Chen JX, He LN, Jin CH,
Wang LL, Zhang FL and An LJ: Identification and characterization of
reactive metabolites in myristicin-mediated mechanism-based
inhibition of CYP1A2. Chem Biol Interact. 237:133–140.
2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yang AH, Zhang L, Zhi DX, Liu WL, Gao X
and He X: Identification and analysis of the reactive metabolites
related to the hepatotoxicity of safrole. Xenobiotica.
48:1164–1172. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Koenigs LL and Trager WF: Mechanism-based
inactivation of P450 2A6 by furanocoumarins. Biochemistry.
37:10047–10061. 1998.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Nakayama S, Takakusa H, Watanabe A, Miyaji
Y, Suzuki W, Sugiyama D, Shiosakai K, Honda K, Okudaira N, Izumi T
and Okazaki O: Combination of GSH trapping and time-dependent
inhibition assays as a predictive method of drugs generating highly
reactive metabolites. Drug Metab Dispos. 39:1247–1254.
2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Peterson LA, Cummings ME, Vu CC and Matter
BA: Glutathione trapping to measure microsomal oxidation of furan
to cis-2-butene-1,4-dial. Drug Metab Dispos. 33:1453–1458.
2005.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Pariser DM and Wyles RJ: Toxic hepatitis
from oral methoxsalen photochemotherapy (PUVA). J Am Acad Dcrmatol.
3:248–250. 1980.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Bjellerup M, Bruze M, Hansson A, Krook G
and Ljunggren B: Liver injury following administration of
8-methoxypsoralen during PUVA therapy. Acta Derm Venereol.
59:371–372. 1979.PubMed/NCBI
|
|
25
|
Zhao G, Xu D, Yuan Z, Jiang Z, Zhou W, Li
Z, Yin M, Zhou Z, Zhang L and Wang T: 8-Methoxypsoralen disrupts
MDR3-mediated phospholipids efflux and bile acid homeostasis and
its relevance to hepatotoxicity. Toxicology. 386:40–48.
2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Rodriguez-Proteau R, Mata JE, Miranda CL,
Fan Y, Brown JJ and Buhler DR: Plant polyphenols and multidrug
resistance: Effects of dietary flavonoids on drug transporters in
Caco-2 and MDCKII-MDR1 cell transport models. Xenobiotica.
36:41–58. 2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Veronese ML, Gillen LP, Burke JP, Dorval
EP, Hauck WW, Pequignot E, Waldman SA and Greenberg HE:
Exposure-dependent inhibition of intestinal and hepatic CYP3A4 in
vivo by grapefruit juice. J Clin Pharmacol. 43:831–839.
2003.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ohnishi A, Matsuo H, Yamada S, Takanaga H,
Morimoto S, Shoyama Y, Ohtani H and Sawadaet Y: Effect of
furanocoumarin derivatives in grapefruit juice on the uptake of
vinblastine by Caco-2 cells and on the activity of cytochrome P450
3A4. Br J Pharmacol. 130:1369–1377. 2000.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Becquemont L, Verstuyft C, Kerb R,
Brinkmann U, Lebot M, Jaillon P and Funck-Brentano C: Effect of
grapefruit juice on digoxin pharmacokinetics in humans. Clin
Pharmacol Ther. 70:311–316. 2001.PubMed/NCBI
|
|
30
|
Guo LQ, Fukuda K, Ohta T and Yamazoe Y:
Role of furanocoumarin derivatives on grapefruit juice-mediated
inhibition of human CYP3A activity. Drug Metab Dispos. 28:766–771.
2000.PubMed/NCBI
|
|
31
|
Draper AJ, Madan A and Parkinson A:
Inhibition of coumarin 7-hydroxylase activity in human liver
microsomes. Arch Biochem Biophys. 341:47–61. 1997.PubMed/NCBI View Article : Google Scholar
|
|
32
|
da Silva VB, Kawano DF, Carvalho I, da
Conceição EC, de Freitas O and da Silva CH: Psoralen and bergapten:
In silico metabolism and toxicophoric analysis of drugs used to
treat vitiligo. J Pharm Pharm Sci. 12:378–387. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
33
|
Marumoto S and Miyazawa M:
Biotransformation of bergapten and xanthotoxin by Glomerella
cingulata. J Agric Food Chem. 58:7777–7781. 2010.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wang LL, Hai Y, Huang NN, Gao X, Liu WL
and He X: Human cytochrome P450 enzyme inhibition profile of three
flavonoids isolated from Psoralea corylifolia: in silico
predictions and experimental validation. New J Chem. 42(Issue
13)2018.
|
|
35
|
Hai Y, Feng S, Wang L, Ma Y, Zhai Y, Wu Z,
Zhang S and He X: Coordination Mechanism and Bio-Evidence: Reactive
γ-Ketoenal Intermediated Hepatotoxicity of Psoralen and Isopsoralen
Based on Computer Approach and Bioassay. Molecules.
22(1451)2017.
|