Introduction
Allergic rhinitis (AR) is an inflammatory reaction
mainly mediated by immunoglobulin (Ig)E after inhaled allergens
enter the body through the nasal mucosa and it is the manifestation
of a systemic allergic disease in the upper respiratory tract
(1). The pathogenesis of allergic
inflammation of the nasal mucosa is the result of the interaction
among environmental pathogenic factors, genetic susceptibility and
the local and systemic immune defense system, in which the
imbalance of immune response is a critical factor (2). However, the mechanism driving the
immunological imbalance in AR remains elusive.
The γδ T cells, which are predominantly located in
the mucosa, have an important role in mucosal immune processes and
are considered to act as the connection between innate and acquired
immunity (3). γδ T cells are
characterized by the expression of the T-cell receptor (TCR) γδ and
may be further divided into different functional subsets, exerting
their regulatory effects on immune balance by secreting IFN-γ or
IL-17A (4,5), as well as by affecting the
differentiation and function of other types of immune cell, such as
regulatory T cells (Tregs) (6,7). It
was previously demonstrated that different subsets of γδ T cells
predominantly expressed in the mucosa and epithelium of the
respiratory tract may have a pro- or anti-inflammatory role under
different conditions, and they are crucial for regulating the
occurrence and persistence of allergic inflammatory conditions,
including AR (8-10).
However, the molecular factors that steer γδ T cells to
differentiate into different functional subsets in AR remain
elusive.
CD27 and CD28 are two major costimulatory molecules
with independent and non-redundant roles in the activation,
proliferation and survival of γδ T cells (11,12).
Studies in mice have demonstrated that CD27+ γδ T cells
produce IFN-γ, whereas CD27- γδ T cells produce
IL-17(13), while the population of
IFN-γ+ and IL-17+ γδ T cells failed to expand
during infection in CD28-deficient mice (14,15),
indicating the essential roles of CD27 and CD28 co-stimulatory
signals in the activation of specific functional subsets of γδ T
cells. However, the roles of CD27 and CD28 on γδ T cells have not
been extensively investigated in humans, and have not yet been
reported in AR. The aim of the present study was to explore the
expression of CD27 and CD28 on γδ T cells in patients with AR.
Materials and methods
Subjects
The present study was approved by the Ethics
Committee of the Shiyan People's Hospital of Baoan District in
Shenzhen City (Shenzhen, China). Written informed consent was
provided by all subjects or their legal guardians prior to
participation. A total of 14 volunteers with AR were enrolled at
the Department of Otolaryngology, Shiyan People's Hospital of Baoan
District (Shenzhen, China) between December 2018 and October 2019.
The diagnosis of AR was made based on the Allergic Rhinitis and Its
Impact on Asthma guidelines (2). To
evaluate the atopic status, the concentrations of serum IgE
specific to local common inhalant allergens were detected using the
automatic system of immunofluorescence quantitative analysis
(UniCAP 100E; Pharmacia Biotech). A concentration >0.7 IU/ml was
considered as the positive diagnostic criterion for AR. In
addition, 12 healthy control (HC) volunteers were recruited, each
with no clinical symptoms of any allergic disease and negative
serum IgE. Subjects who received oral glucocorticoids,
antihistamines or immunotherapy within 1 month were excluded. The
characteristics of AR and HC subjects are listed in Table I. Peripheral venous blood samples
anticoagulated with heparin were collected from each subject.
 | Table IClinical characteristics of HC and AR
subjects. |
Table I
Clinical characteristics of HC and AR
subjects.
| Item | HC subjects
(n=12) | AR subjects
(n=14) | P-value |
|---|
| Age (years) | 21 (9-39) | 16 (8-41) | 0.381 |
| Sex
(male/female) | 6/6 | 8/6 | N/A |
| HDM-specific IgE
(IU/ml) | 0.21 (0.17) | 24.44 (26.96) | <0.001 |
Flow cytometry
Peripheral blood mononuclear cells (PBMCs) were
isolated from whole blood samples using the Ficoll-Hypaque gradient
centrifugation method. First, the cells were suspended in PBS with
0.5% bovine serum albumin (BSA; MP Biomedicals, LLC) to prevent
unspecific binding. Prior to fluorescence antibody staining, cells
were first incubated with Fixable Viability Dye eFluor 780 (1
µg/ml; cat. no. 65-0865-14) for 20 min at room temperature to
confirm their viability. For cell surface staining, cells were
incubated with APC-conjugated CD3 (clone: UCHT1) monoclonal
antibody (2.5 µg/ml; cat. no. 17-0038-41), FITC-conjugated TCR γ/δ
(clone: 5A6.E91) monoclonal antibody (10 µg/ml; cat. no. TCR2061),
PE-Cyanine5-conjugated CD27 (clone: O323) monoclonal antibody (2.5
µg/ml; cat. no. 15-0279-42) and PE-conjugated CD28 (clone: CD28.2)
monoclonal antibody (2.5 µg/ml; cat. no. 12-0289-42) (all from
eBioscience) for 30 min at 4˚C. For transcription factor staining,
cells were first incubated with PE-Cyanine5-conjugated CD3 (clone:
UCHT1) monoclonal antibody (0.6 µg/ml; cat. no. 15-0038-42) for 30
min at 4˚C, then fixed and permeabilized with the Forkhead box p3
(Foxp3)/Transcription Factor Staining Buffer Set following the
manufacturer's protocol at room temperature, and incubated with
APC-conjugated Foxp3 (clone: PCH101) monoclonal antibody (5 µg/ml;
cat. no. 17-4776-42) for 60 min at room temperature (all from
eBioscience). For intracellular cytokine staining, cells were first
cultured in RPMI-1640 medium supplemented with heat-inactivated
fetal bovine serum (10%) and penicillin/streptomycin (1%) (all from
Gibco; Thermo Fisher Scientific, Inc.) for 5 h at 37˚C in
humidified air containing 5% CO2, in the presence of
Cell Stimulation Cocktail (plus protein transport inhibitors;
eBioscience) consisting of phorbol 12-myristate 13-acetate (81 nM),
ionomycin (1.34 µM), brefeldin A (10.6 µM) and monensin (2 µM).
After 5 h, cells were collected and incubated with
PE-Cyanine5-conjugated CD3 (clone: UCHT1) monoclonal antibody (0.6
µg/ml; cat. no. 15-0038-42) for 30 min at 4˚C. The cells were then
fixed and permeabilized with the Intracellular Fixation and
Permeabilization Buffer Set following the manufacturer's protocol
at room temperature and incubated with PE-conjugated IFN-γ (clone:
XMG1.2) monoclonal antibody (2.5 µg/ml; cat. no. 12-7311-82) and
APC-conjugated IL-17A (clone: eBio64DEC17) monoclonal antibody (0.6
µg/ml; cat. no. 17-7179-42) for 30 min at room temperature (all
from eBioscience). The same concentrations of species- and
subtype-matched antibodies were used as negative controls:
APC-conjugated Mouse IgG1 kappa Isotype Control (2.5 or 0.6 µg/ml;
cat. no. 17-4714-82), FITC-conjugated Mouse IgG1 Isotype Control
(10 µg/ml; cat. no. MA1-10413), PE-Cyanine 5-conjugated Mouse IgG1
kappa Isotype Control (2.5 or 0.6 µg/ml; cat. no. 15-4714-81),
PE-conjugated Mouse IgG1 kappa Isotype Control (2.5 µg/ml; cat. no.
12-4714-82), APC-conjugated Rat IgG12a kappa Isotype Control (5
µg/ml; cat. no. 17-4321-81) and PE-conjugated Rat IgG1 kappa
Isotype Control (2.5 µg/ml; cat. no. 12-4301-82) (all from
eBioscience). Flow cytometry was performed on a Gallios flow
cytometer (Beckman Coulter, Inc.), 5x104 events were
acquired in each panel and data were analyzed using Kaluza Analysis
software (version 2.1; Kaluza Software).
Statistical analysis
GraphPad Prism 6.0 statistical software (GraphPad
Software, Inc.) was used for statistical analysis. Data were tested
for normality using the Shapiro-Wilk test and are presented as the
mean with standard deviation. Comparisons of the percentages of TCR
γ/δ+ CD3+ cells, Foxp3+
CD3+ cells, IFN-γ+ CD3+ cells,
IL-17A+ CD3+ cells, IFN-γ+ γδ T
cells, and IL-17A+ γδ T cells, and the expression
pattern of CD27 and CD28 on γδ T cells between groups were
performed using Student's t-test. Correlations were analyzed using
Pearson's correlation analysis. P<0.05 was considered to
indicate statistically significant differences.
Results
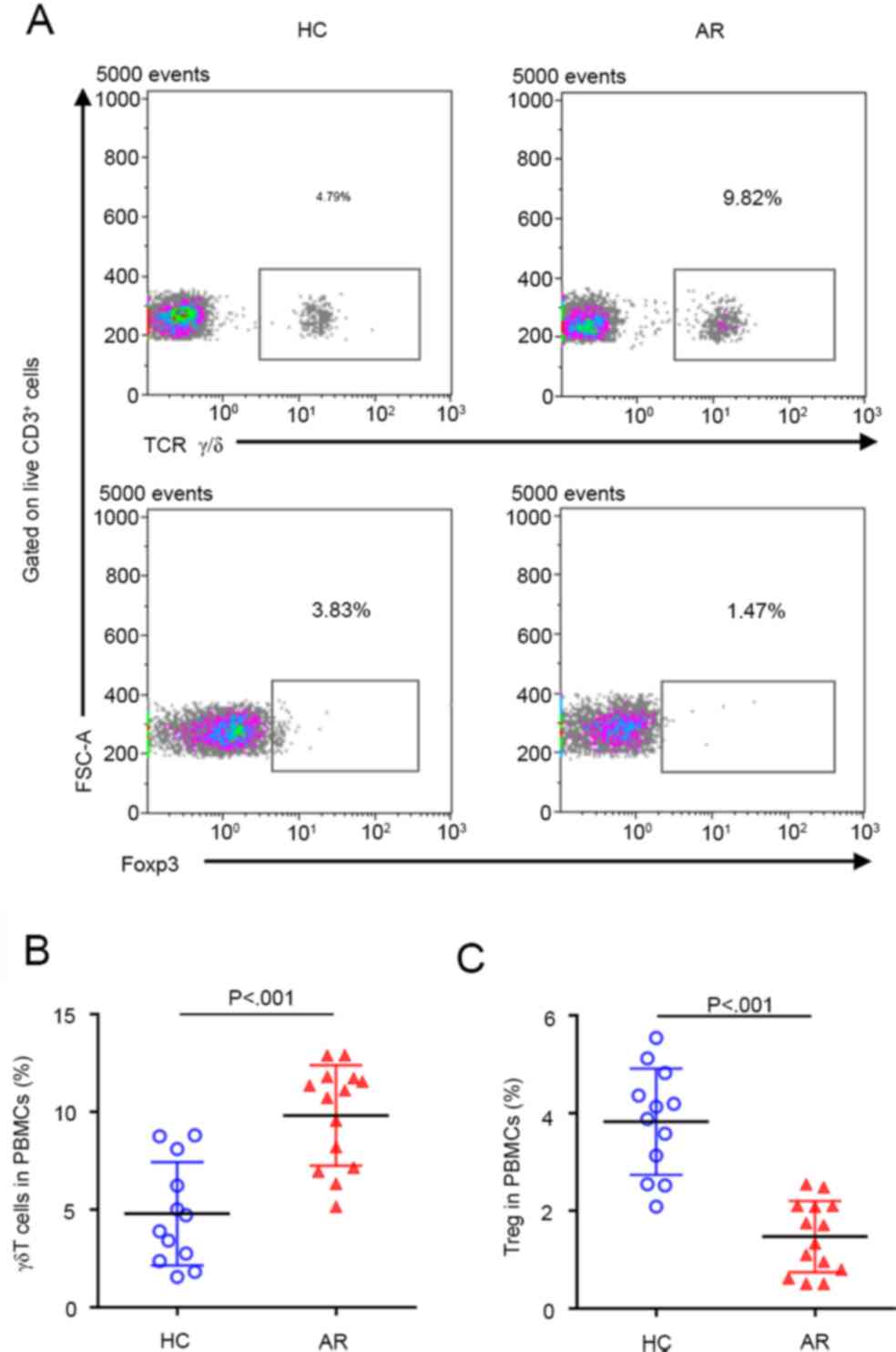
γδ T cells are increased and Tregs are
decreased in AR
The percentages of γδ T cells and Tregs in PBMCs of
HC subjects and patients with AR were first detected. As expected,
compared with HC subjects, the percentage of γδ T cells was
significantly increased, while the percentage of Tregs was
significantly decreased in patients with AR (Fig. 1), suggesting that γδ T cells may
have an important role in the pathogenesis of AR, whereas the
ability of Tregs to regulate immune homeostasis may be
suppressed.
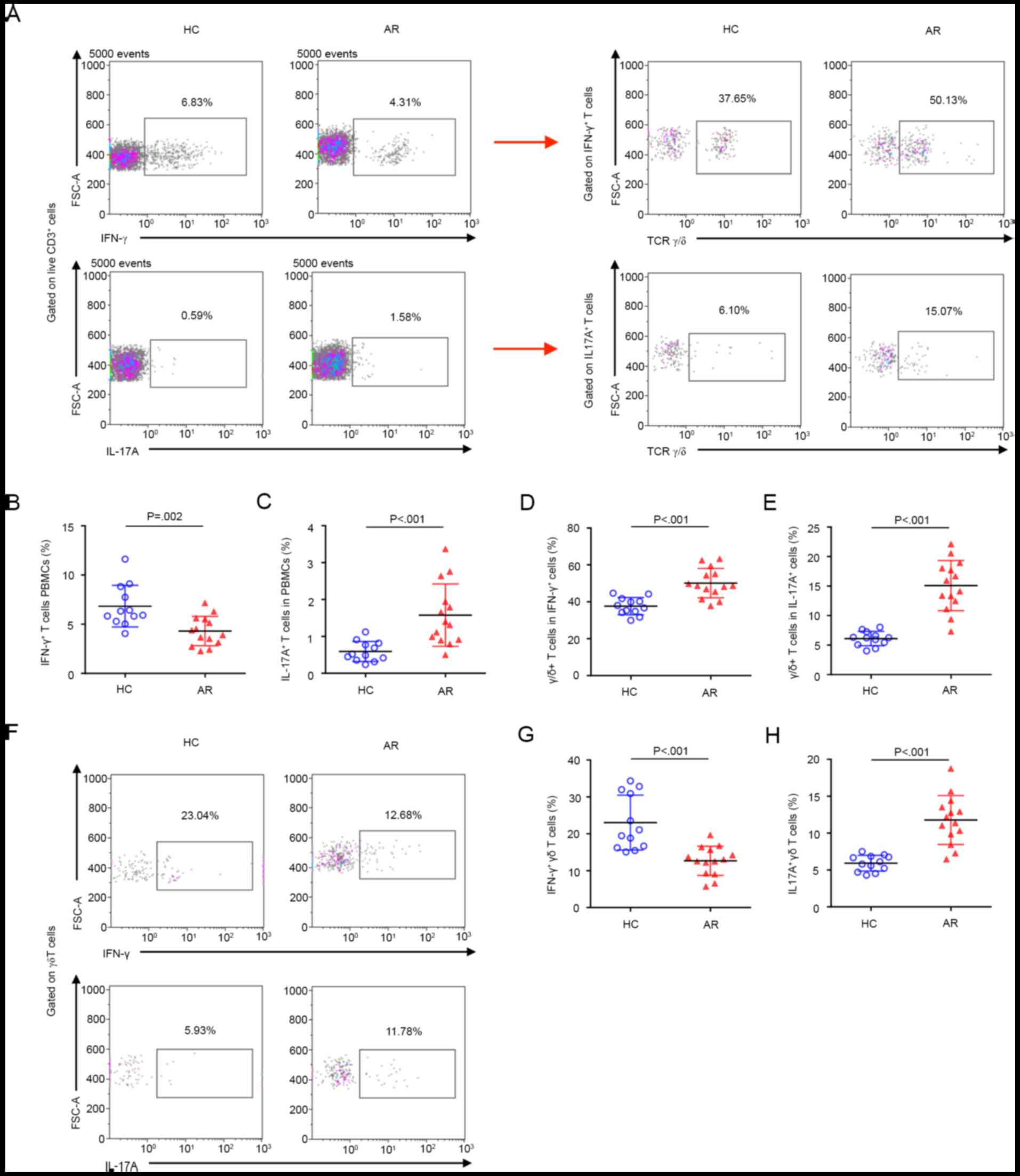
IFN-γ+ γδ T-cell subsets
are decreased and IL-17A+ γδ T-cell subsets are
increased in AR
To further explore the role of γδ T cells in AR, the
percentages of IFN-γ+ and IL-17A+ γδ T cells
in PBMCs of HC and AR subjects were tested. Compared with that in
HC subjects, the percentage of IFN-γ+ T cells was
significantly decreased, while the percentage of IL-17A+
T cells was significantly increased in patients with AR (Fig. 2A-C). Unexpectedly, the expression of
TCR γδ was increased on both IFN-γ+ and
IL-17A+ T cells in AR (Fig.
2A, D and E). Approximately half of the
IFN-γ+ T cells were TCR γδ-positive, suggesting that γδ
T cells may be the major source of IFN-γ in AR. Furthermore, the
expression of TCR γδ on IL-17A+ T cells in AR subjects
was more than 2-fold higher compared with that in HC subjects,
suggesting a non-negligible role of γδ T cells in promoting
inflammation in AR by significantly increasing the secretion of
IL-17A. More importantly, the percentage of IFN-γ+ γδ T
cells was significantly decreased and the percentage of
IL-17A+ γδ T cells was increased in AR (Fig. 2F-H), further supporting that the
IFN-γ+ and IL-17A+ γδ T-cell subsets have
important immune regulatory roles in AR.
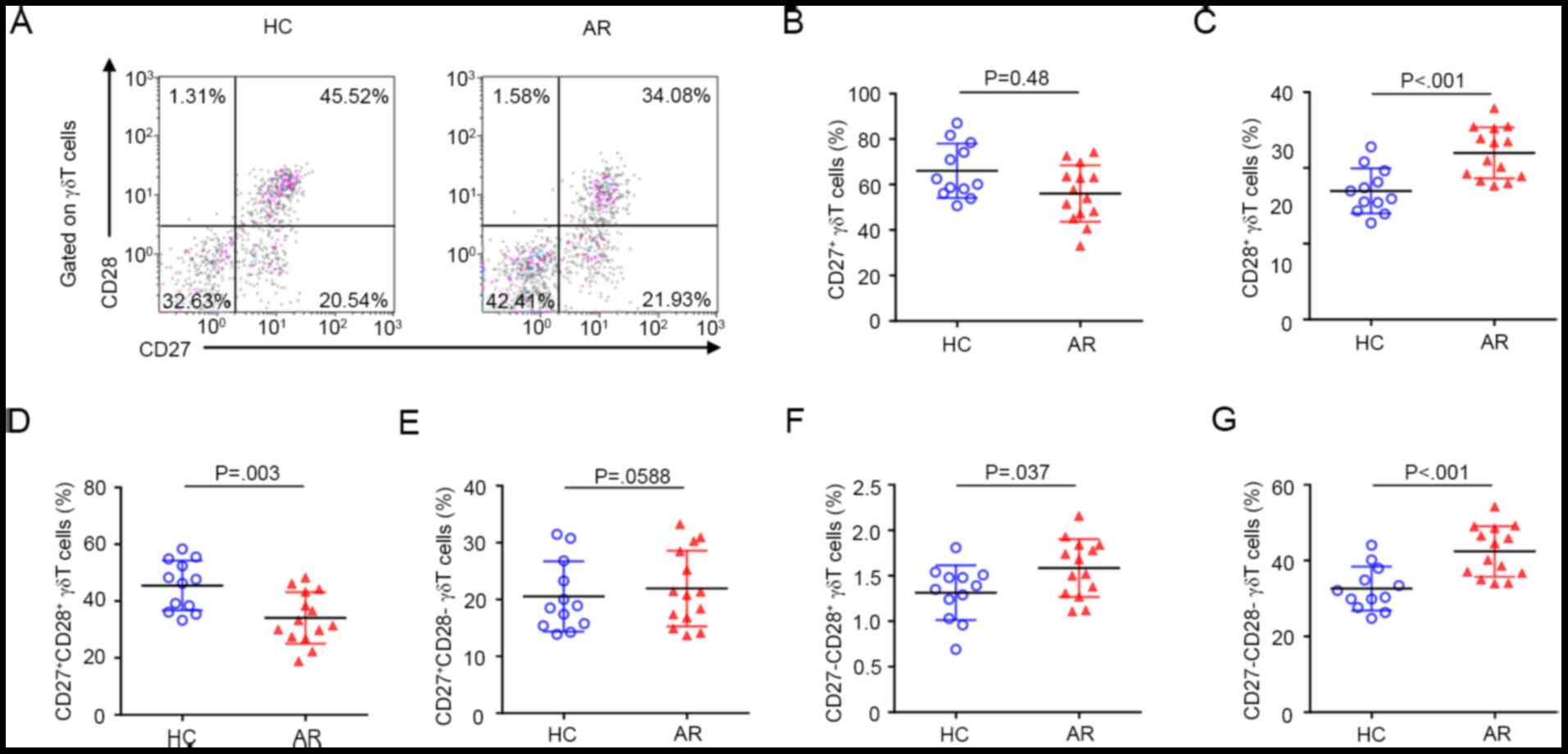
Expression pattern of CD27 and CD28 on
γδ T cells in AR
The expression pattern of CD27 and CD28 on γδ T
cells was
CD27+CD28+>CD27-CD28->CD27+CD28->CD27-CD28+
in PBMCs from HC subjects. By contrast, this pattern was
CD27-CD28->CD27+CD28+>CD27+CD28->CD27-CD28+
in AR subjects (Fig. 3A). Overall,
compared with HC subjects, the percentage of CD27+ γδ T
cells was significantly decreased, while the percentage of
CD28+ γδ T cells was significantly increased in patients
with AR (Fig. 3A-C). Further
analysis demonstrated that the percentage of
CD27+CD28+ γδ T-cell subsets was
significantly decreased in AR (Fig.
3D). The percentage of CD27+CD28- γδ
T-cell subsets in patients with AR was similar to that in HC
subjects (Fig. 3E). The percentages
of CD27-CD28+ and
CD27-CD28- γδ T-cell subsets were both
significantly increased in AR (Fig.
3F and G).
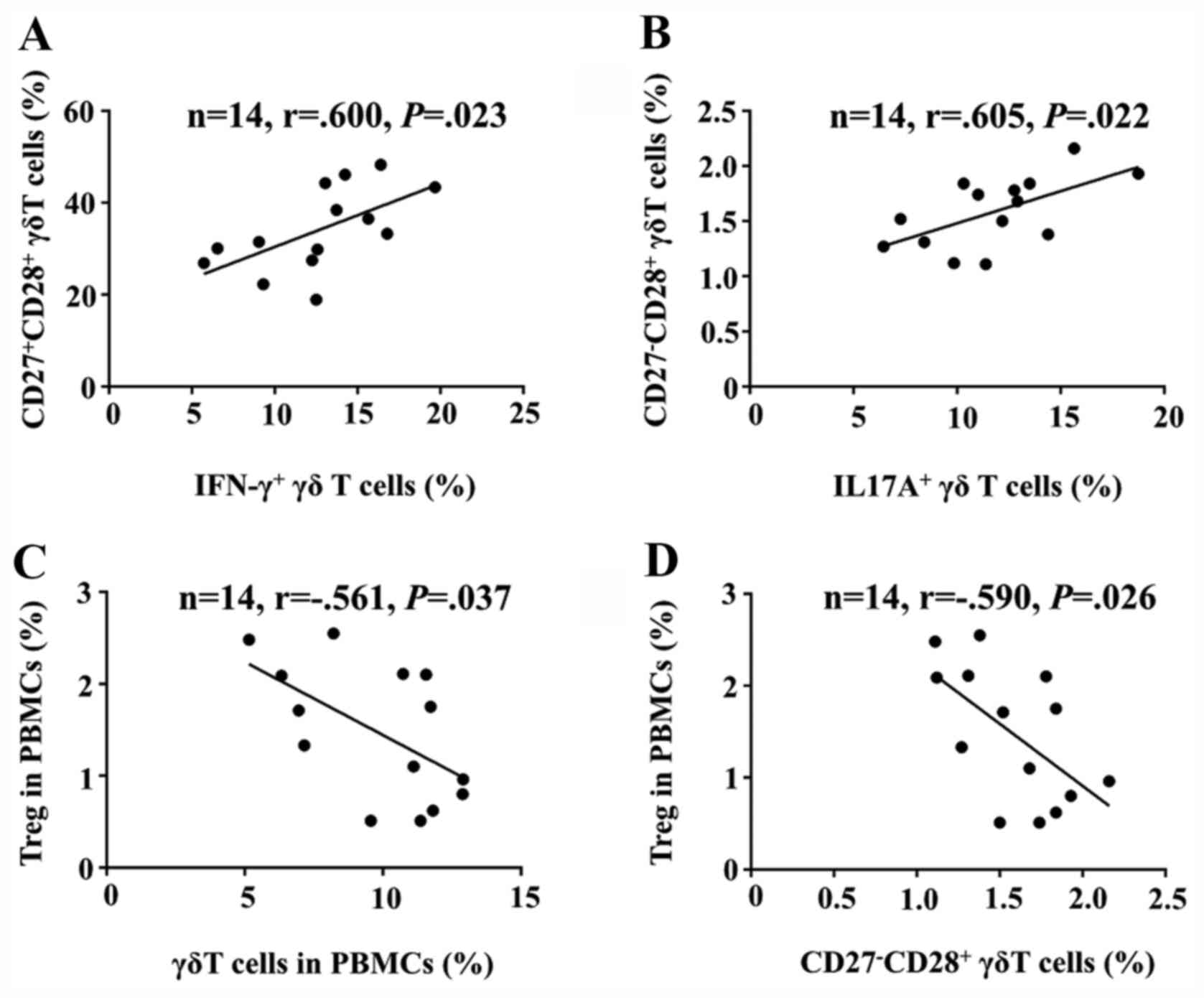
Correlation of CD27 and CD28 with γδ
T-cell subsets in AR
Next, the correlations between CD27 and CD28
expression on γδ T cells and IFN-γ+ or
IL-17A+ γδ T-cell subsets from patients with AR were
analyzed. The results demonstrated that the percentage of the
CD27+CD28+ γδ T-cell subset was positively
correlated with that of the IFN-γ+ γδ T-cell subset
(Fig. 4A) and the percentage of the
CD27-CD28+ γδ T-cell subset was positively
correlated with that of the IL-17A+ γδ T-cell subset
(Fig. 4B). There was no correlation
between the respective other combinations, for instance, between
the CD27+CD28+ γδ T-cell subset with the
IL-17A+ γδ T-cell subset and the
CD27-CD28+ γδ T-cell subset with the
IFN-γ+ γδ T-cell subset (data not shown). These results
suggested that CD28 is necessary for γδ T cells to produce
cytokines, with the CD27+ subset producing IFN-γ and the
CD27- subset producing IL-17A.
Correlation of γδ T-cell subsets with
Tregs in AR
Finally, the correlation between γδ T-cell subsets
and Tregs among the PBMCs of patients with AR was analyzed. The
percentage of γδ T cells was negatively correlated with that of
Tregs (Fig. 4C). In addition, the
percentage of the CD27-CD28+ γδ T-cell subset
was negatively correlated with that of Tregs (Fig. 4D), suggesting that the
CD27-CD28+ γδ T-cell subset may be involved
with the downregulation of the number and function of Tregs in
AR.
Discussion
The results of the present study demonstrated that
the elevated γδ T cells in the peripheral blood of patients with AR
had an increased percentage of the CD27-CD28+
γδ T-cell subset, which was positively correlated with the
percentage of IL-17A+ γδ T cells and negatively
correlated with Tregs. Furthermore, the percentage of the
CD27+CD28+ γδ T-cell subset was significantly
decreased and positively correlated with the percentage of
IFN-γ+ γδ T cells. Given that the IFN-γ+ and
IL-17A+ γδ T-cell subsets have been indicated to exert
anti- or pro-inflammatory effects on autoimmune diseases (16), the present results provide evidence
supporting a potential important role for CD27 and CD28 in the
immune regulatory function of γδ T cells in AR.
The major mechanism underlying AR has been indicated
to be the dysfunction of the mucosal barrier, producing an
overactive, unbalanced immune response against certain substances
that are originally ‘harmless’ in vitro, such as allergens
(1). Studies have proven that γδ T
cells predominantly occur in the epithelium and mucosa and have an
indispensable role in orchestrating the immune response of allergic
diseases (17-20).
In the present study, a significant elevation in the percentage of
γδ T cells in PBMCs from patients with AR was observed when
compared with the control subjects, which is largely consistent
with previous studies (21,22), confirming the important role of γδ T
cells in the pathogenesis of AR.
Previous studies have indicated that AR is
characterized by a T helper type (Th) 2-skewed inflammation, with
decreased levels of the Th1-associated cytokine IFN-γ and increased
levels of the Th17-associated cytokine IL-17A (23,24).
Consistently, the data of the present study also demonstrated a
significant decrease in the percentage of IFN-γ+ and an
increase in the percentage of IL-17A+ T cells in PBMCs
of patients with AR compared with control subjects. The average
percentage of IFN-γ+ T cells in HC subjects was 6.83%,
which was slightly lower than the range reported by other studies
using similar experimental conditions (25,26).
This may be due to individual differences between the donors and
future experiments with larger sample sizes may eliminate this
difference. γδ T cells are a major source of IFN-γ and IL-17A in
several autoimmune responses and lower airway allergic
inflammations (16,27); this may explain why the percentage
of TCR γδ+ cells was significantly increased in the
IFN-γ- and IL-17A-producing T cells in patients with AR in the
present study. Of note, the percentage of the IFN-γ+ γδ
T-cell subset was significantly decreased and that of the
IL-17A+ γδ T-cell subset was significantly increased in
AR, further supporting that the IL-17A+ γδ T-cell subset
may have an important pro-inflammatory role in AR.
CD27, a member of the TNF receptor (TNFR)
superfamily, and CD28, a member of the Ig superfamily, are typical
T-cell costimulatory receptors that have specific roles in γδ T
cells. They display different modes of intracellular signaling:
CD27 requires the adaptor protein TNFR-associated factor, whereas
CD28 associates directly with protein kinases to link to downstream
signaling mediators (6). Studies in
mice have demonstrated that CD27 and its ligand, CD70, are
absolutely required for the survival, development, differentiation
and function of γδ T cells (28,29).
Another study in humans suggested that the majority of circulating
Vγ9Vδ2+ T cells express CD27, which acts as a coreceptor
that promotes, in conjunction with TCR-mediated signals, the
production of IFN-γ (30). In αβ T
cells, the interaction of CD28 and its ligands, B7-1 (CD80) and
B7-2 (CD86), provide a ‘second signal’ that is crucial for T-cell
activation, proliferation and survival (31,32).
Although the requirement of CD28/B7 signaling for the activation of
γδ T cells remains controversial (29), certain studies have revealed that
CD28 is expressed on γδ T cells in the lymph nodes and has the
ability to promote the activation, proliferation and survival of γδ
T cells, mainly via IL-2 secretion (12,15).
Data from a study on CD28-deficient mice revealed that the
IFN-γ+ and IL-17+ γδ T-cell populations
failed to expand during infection (15), suggesting that CD28 is also required
for the expansion of γδ T cells. In the present study, the
decreased proportion of CD27+CD28+ and
increased proportion of CD27-CD28+ and
CD27-CD28- γδ T cells in the PBMCs of
patients with AR, as well as the positive correlations of the
CD27+CD28+ γδ T-cell subset with the
IFN-γ+ γδ T-cell subset, and of the
CD27-CD28+ γδ T-cell subset with the
IL-17A+ γδ T-cell subset, suggest an aberrant activation
and proliferation pattern of γδ T cells in AR and a different
cytokine profile compared with that of healthy subjects.
Since first identified in 1995, the immunomodulatory
effect of CD4+CD25+ Tregs on the maintenance
of immune homeostasis has been proven (33). It has been demonstrated that Tregs
may inhibit excessive type I hypersensitivity by producing IL-10
and TGF-β, thereby having a pivotal role in the immune tolerance of
allergens (34). It has been
reported that the number and function of Tregs are significantly
decreased in the peripheral blood during allergic airway
inflammation (35). Furthermore, γδ
T cells and Tregs may exert a synergistic regulatory effect in the
pathogenesis of AR (22,36). Since the specific transcription
factor Foxp3 has a key role in the development and functional
maintenance of Tregs (37), and is
stably and constitutively expressed at a high level in
CD4+CD25+ regulatory T cells,
Foxp3+CD3+ cells in PBMCs were identified as
Tregs in numerous studies (38,39).
Therefore, in the present study, only Foxp3 expression on
CD3+ T cells was analyzed. It was observed that the
percentage of Tregs was significantly decreased in patients with
AR. More importantly, negative correlations were obtained between
the percentage of Tregs with that of the
CD27-CD28+ γδ T-cell subset. These results
suggest that the CD27-CD28+ γδ T-cell subset
may have a role in the decrease of the number and function of Tregs
in AR. However, as the percentage of the
CD27-CD28+ γδ T-cell subset was low, the
function of CD27-CD28+ γδ T cells must still
be proven by further experimental studies.
In conclusion, the increased numbers of circulating
γδ T cells were characterized by an increased percentage of the
CD27-CD28+ γδ T-cell subset, which was
indicated to be positively correlated with the percentage of
IL-17A+ γδ T cells and negatively correlated with Tregs
in patients with AR. Furthermore, the decreased
CD27+CD28+ γδ T-cell subset was positively
correlated with the percentage of IFN-γ+ γδ T cells.
These data indicate the potentially important roles of CD27 and
CD28 in the immune regulatory role of γδ T cells in AR. Given the
limitations of the correlation analysis, it is necessary to fully
verify the specific functions of CD27 and CD28 on γδ T cells in AR
via further experiments, such as cell experiments.
Acknowledgements
The authors gratefully acknowledge the guidance of
the experiments by Dr Rui Zheng (Department of
Otorhinolaryngology-Head and Neck Surgery, The Third Affiliated
Hospital, Sun Yat-sen University, Guangzhou, China).
Funding
The present study was supported by the Science and
Technology Program of Shenzhen Science and Technology Innovation
Committee (grant no. JCYJ20180305163629056). The funding body has
not influenced the study design, data collection, analysis or its
interpretations.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QW contributed to the conceptualization and design
of the study and drafted the manuscript. QS and QC performed the
flow cytometry assays and helped draft the manuscript. HL and DL
helped to analyze data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Shiyan People's Hospital of Baoan District in
Shenzhen City (Shenzhen, China). Written informed consent was
provided by all subjects or their legal guardians prior to
participation.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Greiner AN, Hellings PW, Rotiroti G and
Scadding GK: Allergic rhinitis. Lancet. 378:2112–2122.
2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Brozek JL, Bousquet J, Agache I, Agarwal
A, Bachert C, Bosnic-Anticevich S, Brignardello-Petersen R,
Canonica GW, Casale T, Chavannes NH, et al: Allergic rhinitis and
its impact on asthma (ARIA) guidelines-2016 revision. J Allergy
Clin Immunol. 140:950–958. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Pang DJ, Neves JF, Sumaria N and
Pennington DJ: Understanding the complexity of γδ T-cell subsets in
mouse and human. Immunology. 136:283–290. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Costa MF, Bornstein VU, Candea AL,
Henriques-Pons A, Henriques MG and Penido C: CCL25 induces
α4β7 integrin-dependent migration of
IL-17+ γδ T lymphocytes during an allergic reaction. Eur
J Immunol. 42:1250–1260. 2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Dyring-Andersen B, Skov L, Løvendorf MB,
Bzorek M, Søndergaard K, Lauritsen JP, Dabelsteen S, Geisler C and
Bonefeld CM: CD4+ T cells producing interleukin (IL)-17,
IL-22 and interferon-γ are major effector T cells in nickel
allergy. Contact Dermatitis. 68:339–347. 2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kohlgruber AC, Gal-Oz ST, LaMarche NM,
Shimazaki M, Duquette D, Koay HF, Nguyen HN, Mina AI, Paras T,
Tavakkoli A, et al: γδ T cells producing interleukin-17A regulate
adipose regulatory T cell homeostasis and thermogenesis. Nat
Immunol. 19:464–474. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Petermann F, Rothhammer V, Claussen MC,
Blanco LR, Heink S, Prinz I, Hemmer B, Kuchroo VK, Oukka M and Korn
T: γδ T cells enhance autoimmunity by restraining regulatory T cell
responses via an interleukin-23-dependent mechanism. Immunity.
33:351–363. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Huang Y, Yang Z, McGowan J, Huang H,
O'Brien RL and Born WK: Regulation of IgE responses by γδ T cells.
Curr Allergy Asthma Rep. 15(13)2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ullah MA, Revez JA, Loh Z, Simpson J,
Zhang V, Bain L, Varelias A, Rose-John S, Blumenthal A, Smyth MJ,
et al: Allergen-induced IL-6 trans-signaling activates γδ T cells
to promote type 2 and type 17 airway inflammation. J Allergy Clin
Immunol. 136:1065–1073. 2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zheng R and Yang Q: The role of the γ δ T
cell in allergic diseases. J Immunol Res.
2014(963484)2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ribeiro ST, Ribot JC and Silva-Santos B:
Five layers of receptor signaling in γδ T-cell differentiation and
activation. Front Immunol. 6(15)2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ribot JC and Silva-Santos B:
Differentiation and activation of γδ T lymphocytes: Focus on CD27
and CD28 costimulatory receptors. Adv Exp Med Biol. 785:95–105.
2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ribot JC, deBarros A, Pang DJ, Neves JF,
Peperzak V, Roberts SJ, Girardi M, Borst J, Hayday AC, Pennington
DJ and Silva-Santos B: CD27 is a thymic determinant of the balance
between interferon-gamma- and interleukin 17-producing gammadelta T
cell subsets. Nat Immunol. 10:427–436. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Beyersdorf N, Kerkau T and Hünig T: CD28
co-stimulation in T-cell homeostasis: A recent perspective.
Immunotargets Ther. 4:111–122. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ribot JC, Debarros A, Mancio-Silva L,
Pamplona A and Silva-Santos B: B7-CD28 costimulatory signals
control the survival and proliferation of murine and human γδ T
cells via IL-2 production. J Immunol. 189:1202–1208.
2012.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Peters C, Kabelitz D and Wesch D:
Regulatory functions of γδ T cells. Cell Mol Life Sci.
75:2125–2135. 2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Akitsu A and Iwakura Y:
Interleukin-17-producing γδ T (γδ17) cells in inflammatory
diseases. Immunology. 155:418–426. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Reyes NJ, Mayhew E, Chen PW and Niederkorn
JY: γδ T cells are required for maximal expression of allergic
conjunctivitis. Invest Ophthalmol Vis Sci. 52:2211–2216.
2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Takai T and Ikeda S: Barrier dysfunction
caused by environmental proteases in the pathogenesis of allergic
diseases. Allergol Int. 60:25–35. 2011.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wu J, Xu L, Han X, Hu H, Qi F, Bai S, Chai
R, Teng Y and Liu B: Role of γδ T cells in exacerbated airway
inflammation during reinfection of neonatally primed mice in
adulthood. J Med Virol. 89:2108–2115. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Xuekun H, Qintai Y, Yulian C and Gehua Z:
Correlation of gammadelta-T-cells, Th17 cells and IL-17 in
peripheral blood of patients with allergic rhinitis. Asian Pac J
Allergy Immunol. 32:235–239. 2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yang Q, Li C, Wang W, Zheng R, Huang X,
Deng H, Jin P, Tan K, Yan Y and Wang D: Infiltration pattern of
gammadelta T cells and its association with local inflammatory
response in the nasal mucosa of patients with allergic rhinitis.
Int Forum Allergy Rhinol. 9:1318–1326. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Bajoriuniene I, Malakauskas K, Lavinskiene
S, Jeroch J, Gasiuniene E, Vitkauskiene A and Sakalauskas R:
Response of peripheral blood Th17 cells to inhaled
Dermatophagoides pteronyssinus in patients with allergic
rhinitis and asthma. Lung. 190:487–495. 2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
König K, Klemens C, Eder K, San Nicoló M,
Becker S, Kramer MF and Gröger M: Cytokine profiles in nasal fluid
of patients with seasonal or persistent allergic rhinitis. Allergy
Asthma Clin Immunol. 11(26)2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Baeten D, Van Damme N, Van den Bosch F,
Kruithof E, De Vos M, Mielants H, Veys EM and De Keyser F: Impaired
Th1 cytokine production in spondyloarthropathy is restored by
anti-TNFalpha. Ann Rheum Dis. 60:750–755. 2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Migalovich Sheikhet H, Villacorta Hidalgo
J, Fisch P, Balbir-Gurman A, Braun-Moscovici Y and Bank I:
Dysregulated CD25 and cytokine expression by γδ T cells of systemic
sclerosis patients stimulated with cardiolipin and zoledronate.
Front Immunol. 9(753)2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhao Y, Yang J and Gao YD: Altered
expressions of helper T cell (Th)1, Th2, and Th17 cytokines in
CD8+ and γδ T cells in patients with allergic asthma. J
Asthma. 48:429–436. 2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Born WK and O'Brien RL: γδ T cells
develop, respond and survive-with a little help from CD27. Eur J
Immunol. 41:26–28. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ribot JC, debarros A and Silva-Santos B:
Searching for 'signal 2': Costimulation requirements of γδ T cells.
Cell Mol Life Sci. 68:2345–2355. 2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
DeBarros A, Chaves-Ferreira M, d'Orey F,
Ribot JC and Silva-Santos B: CD70-CD27 interactions provide
survival and proliferative signals that regulate T cell
receptor-driven activation of human γδ peripheral blood
lymphocytes. Eur J Immunol. 41:195–201. 2011.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Lenschow DJ, Walunas TL and Bluestone JA:
CD28/B7 system of T cell costimulation. Annu Rev Immunol.
14:233–258. 1996.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Smith-Garvin JE, Koretzky GA and Jordan
MS: T cell activation. Annu Rev Immunol. 27:591–619.
2009.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Sakaguchi S, Sakaguchi N, Asano M, Itoh M
and Toda M: Pillars article: Immunologic self-tolerance maintained
by activated T cells expressing IL-2 receptor α-chains (CD25).
Breakdown of a single mechanism of self-tolerance causes various
autoimmune diseases. J. Immunol. 1995. J Immunol. 186:3808–3821.
2011.PubMed/NCBI
|
|
34
|
Palomares O, Martin-Fontecha M, Lauener R,
Traidl-Hoffmann C, Cavkaytar O, Akdis M and Akdis CA: Regulatory T
cells and immune regulation of allergic diseases: Roles of IL-10
and TGF-β. Genes Immun. 15:511–520. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
van de Veen W, Wirz OF, Globinska A and
Akdis M: Novel mechanisms in immune tolerance to allergens during
natural allergen exposure and allergen-specific immunotherapy. Curr
Opin Immunol. 48:74–81. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zheng R, Wu X, Huang X, Chen Y, Yang Q, Li
Y and Zhang G: Gene expression pattern of Treg and TCR Vγ subfamily
T cells before and after specific immunotherapy in allergic
rhinitis. J Transl Med. 12(24)2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Hori S, Nomura T and Sakaguchi S: Pillars
article: Control of regulatory T cell development by the
transcription factor Foxp3. Science 2003 299: 1057-1061. J Immunol.
198:981–985. 2017.PubMed/NCBI
|
|
38
|
Xue M, Liang H, Tang Q, Xue C, He X, Zhang
L, Zhang Z, Liang Z, Bian K, Zhang L and Li Z: The protective and
immunomodulatory effects of fucoidan against 7,12-Dimethyl
benz[a]anthracene-induced experimental mammary carcinogenesis
through the PD1/PDL1 signaling pathway in rats. Nutr Cancer.
69:1234–1244. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhang X, Huang T and Wu Y, Peng W, Xie H,
Pan M, Zhou H, Cai B and Wu Y: Inhibition of the PI3K-Akt-mTOR
signaling pathway in T lymphocytes in patients with active
tuberculosis. Int J Infect Dis. 59:110–117. 2017.PubMed/NCBI View Article : Google Scholar
|