Introduction
The global prevalence of diabetes has increased
markedly. Due to the aging population and continued increase in
obesity rates, the prevalence is expected to rise to 592 million by
2035(1). Concomitantly, the
incidence of type 2 diabetes mellitus (T2DM) has increased rapidly,
and patients with diabetes often experience a wide range of
complications. T2DM is associated with low-grade inflammation, and
this immune inflammatory response can be influenced and regulated
by monocytes and macrophages via the secretion of cytokines and
antigen-presenting cells (2-4).
Previous studies have shown that the tissue macrophage status
regulates the development and progression of T2DM (4,5). In
addition, other studies have shown that peritoneal macrophages
modulate the immune response by regulating cytokines and nitric
oxide production in diabetic rats (6,7). Our
previous study demonstrated that the P2X purinoceptor 7
(P2X7) receptor of the mononuclear phagocyte system is
involved in the pathology of diabetes (8). As expression of the P2X7
receptor may be associated with inflammation (9), it would be interesting to investigate
whether the P2X7 receptor is able to ameliorate the
chronic inflammatory state of T2DM.
Long non-coding RNAs (lncRNAs) are generally RNA
transcripts >200 nucleotides in length that lack the ability to
encode proteins (10). Previous
studies have shown that lncRNAs may contribute to the regulation of
cell apoptosis, proliferation and differentiation at the RNA
epigenetic level via the induction of changes in gene transcription
and post-transcriptional modifications (11). uc.48+ is an lncRNA that has been
observed to be expressed at increased levels, along with the
P2X7 receptor, in the superior cervical ganglia of a rat
model of T2DM, and is associated with cardiac autonomic dysfunction
(12). In our previous study,
uc.48+ small interfering RNA (siRNA) was found to influence immune
and inflammatory responses in a diabetic monophagocyte system
through the P2X7 receptor in vitro (13). A previous study revealed that
macrophages may exist in large numbers in the peritoneum of mice
(14). Therefore, the aim of the
present study was to investigate whether uc.48+ exerts an effect on
mouse abdominal cells through P2X7 receptors in
vivo. The ERK signaling pathway is the key pathway by which
signals are transmitted from surface receptors to the nucleus
(15). Decreased ERK
phosphorylation appears to be associated with decreased
P2X7 receptor expression in RAW264.7 macrophages
(13). In addition, uc.48+ siRNA
has been shown to ameliorate diabetic sympathetic neuropathy in a
rat model of T2DM (12). However,
the exact mechanism by which uc.48+ affects the abdominal cells and
neuropathological changes in mouse models of T2DM requires
elucidation.
The present study aimed to investigate if uc.48+
siRNA has a beneficial effect on the function of abdominal cells,
and whether it ameliorates the neuropathological changes associated
with T2DM through the P2X7 receptor and ERK signaling
pathway. Therefore, the role of uc.48+ and the mechanism underlying
its effects on the pathological changes in T2DM were evaluated.
This was achieved by monitoring and assessing the effects of uc.48+
siRNA on mice with T2DM and their abdominal cells in which
P2X7 receptor expression was upregulated.
Materials and methods
Animals and animal groups
A total of 24 male Kunming mice of clean grade
(32-42 g) were purchased from the Center of Laboratory Animal
Science of Nanchang University at 8 weeks of age. They were
acclimatized for 2 weeks at room temperature with 40-60% relative
humidity and 12-h light/dark cycles and given standard feed with
free access to drinking water, and randomly divided into four
groups (n=6 in each group), comprising the control group, the DM
group, DM treated with uc.48+ siRNA group (DM + uc.48+ si) and DM
treated with scrambled siRNA group (DM + NCsi). All procedures
involving animals were approved by the Animal Care and Use
Committee of the Medical College of Nanchang University (approval
no. 2018 16).
Mice in the DM group were fed with a high-sugar and
-fat diet (consisting of 22% fat, 48% carbohydrate and 20% protein
with a total calorific value of 44.3 kJ/kg) for 4 weeks and
subsequently injected intraperitoneally (i.p.) with streptozotocin
(STZ; 80 mg/kg). Control mice were fed with normal diet (consisting
of 5% fat, 53% carbohydrate and 23% protein, with a total calorific
value of 25 kJ/kg) for 4 weeks and subsequently injected i.p. with
the same concentration of saline. On the third day after STZ
injection (the last day of week 6), the blood glucose levels of the
mice were measured. The successful establishment of the DM model
was confirmed by fasting plasma glucose levels of >11.1 mmol/l
and postprandial plasma glucose levels of >16.7 mmol/l (16,17).
uc.48+ siRNA treatment
The siRNA sequences specific for uc.48+ were
purchased from Invitrogen (Thermo Fisher Scientific, Inc.). The
following target sequence was used: sense,
5'-GGCACUACUACUUGCAGAATT-3' and anti-sense,
5'-UUCUGCAAGUAGUAGUGCCTT-3'. The uc.48+ siRNA was injected i.p.
into DM model mice at the end of week 7 along with
Entranster™-in vivo Transfection Reagent (Engreen
Biosystem Co., Ltd.) to establish the DM + uc.48+ si group.
Similarly, DM model mice were injected with scrambled siRNA: sense,
5'-UUCUCCGAACGUGUCACGUTT-3' and anti-sense,
5'-ACGUGACACGUUCGGAGAATT-3'; Invitrogen; Thermo Fisher Scientific,
Inc.) and transfection reagent to establish the DM + NCsi group.
According to the transfection reagent manufacturer's protocol, a
mixture of 1 µg uc.48+ or scrambled siRNA in diluent (100 µl) and
0.5 µl transfection reagent in diluent (100 µl) was injected i.p.
The control and DM groups were injected with the same volume of
saline.
At the end of week 8, the animals were sacrificed
with CO2 using a chamber displacement rate of
10-30%/min. Subsequently, 0.6-1.0 ml blood was collected by cardiac
puncture and abdominal cells were harvested from the mice by
intra-abdominal lavage.
Nociceptive behavior assays
Since uc.48+ siRNA has been shown to ameliorate
diabetic sympathetic neuropathy in type 2 diabetic rats (8), the ability of uc.48+ siRNA to
alleviate neuropathological changes in diabetic mice was
investigated through behavioral assays in the present study. The
behavioral assays comprised the assessment of mechanical withdrawal
threshold (MWT) and thermal withdrawal latency (TWL) after 6 weeks
(prior to STZ injection), 7 weeks (prior to siRNA injection) and 8
weeks (following siRNA injection), respectively.
Measurement of MWT
Noxious-pressure stimulation was used to evaluate
mechanical hyperalgesia. The experimental protocol of Liu et
al (18) was used.
Measurement of TWL
Noxious heat stimulation was applied using the
Thermal Paw Stimulation System (BME-410C; Boerni Science and
Technology Co., Ldt.) and hyperalgesia was assessed using thermal
stimulation by Hargreaves' test. The experimental protocol
published by Liu et al (18)
was followed, except the difference in animal strains used.
Measurement of heart rate (HR) and
blood pressure
HR and blood pressure are indicators that reflect
the health of the cardiovascular system. Each mouse was assessed by
measuring its HR and blood pressure, including systolic blood
pressure (SBP) and diastolic blood pressure (DBP), after 6 weeks
(prior to STZ injection), 7 weeks (prior to siRNA injection) and 8
weeks (following siRNA injection). HR and blood pressure were
assessed through an indirect tailcuff method (Softron BP-98A;
Softron Co., Ltd.). A tailcuff 1.5 cm in diameter and 3.2 cm in
length was used. Systolic pulsation was detected using an
electrosphygmograph coupler (ZHHX-Z, MD3000; Anhui Zhenghua
Biologic Apparatus Facilities Co., Ltd.). In brief, the
experimental protocol published by Wu et al (12) was followed, except the difference in
animal strains used. The blood pressure of the mice was measured
five times in the morning at each time point by one person.
Cytokine assays
The inflammatory state of the body can be evaluated
by the determination of cytokines. The cytokine concentrations of
each mouse were detected in the serum at the end of the 8-week
period following animal sacrifice. The cytokines assessed comprised
tumor necrosis factor (TNF)-α, interleukin (IL)-1β and IL-10. The
concentration levels of IL-10 (cat. no. EK0417), IL-1β (cat. no.
EK0394) and TNF-α (cat. no. EK0527) were determined using ELISA
kits (Wuhan Boster Biological Technology, Ltd.) according to the
manufacturer's protocol.
Isolation of abdominal cells
The effect of uc.48+ siRNA on P2X7
receptors in a diabetic mononuclear phagocyte system has previously
been demonstrated in vitro (13). Therefore, the present study aimed to
determine whether uc.48+ affects mouse abdominal cells through
P2X7 receptors in vivo. Abdominal cells were
harvested from the mice at the end of the 8-week period by
intra-abdominal lavage, cultured overnight in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS
(Hyclone; GE Healthcare Life Sciences), 100 U/ml penicillin and 100
mg/ml streptomycin sulphate at 37˚C in a humidified atmosphere
containing 5% CO2, and enriched for abdominal cells by
washing away non-adherent peritoneal cells with lukewarm serum-free
culture medium. Wright-Giemsa dye was used for staining at room
temperature for 5 min and samples were observed using SZ61 Olympus
microscope. The cell seeding density was 5x105/ml for
the Wright-Giemsa and trypan blue staining. The viability of the
purified adherent abdominal cells was determined by trypan blue
exclusion assay. A total of 0.1 ml trypan blue stock solution was
added to 1 ml of cells at room temperature for 5 min. The number of
stained and total cells was counted. Healthy log-phase cultures
exhibit cell viability of ≥95% (19). The following equation was used in
the present study: % Viable cells=[1.00-(number of blue-stained
cells/number of all cells)] x100%.
Total RNA isolation and reverse
transcription-PCR (RT-PCR) analysis
Total RNA was isolated from the abdominal cells
using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The quality assessment of
the RNA and synthesized cDNA was performed according to our
previously published method (13).
The PCR amplification of the P2X7 receptor and β-actin
(internal standard for quantification) genes was performed
according to our previously published method (20). The amplification system included 2
µl cDNA, 12.5 µl PCR mixture (Tiangen Biotech Co., Ltd.), 2 µl
primers (1 µl each of sense and antisense primers) and 8.5 µl
nuclease-free water. The sequences of the primers used for RT-PCR
analysis were as follows: P2X7 receptor (171 bp): sense,
5'-GCACGAATTATGGCACCGTC-3' and antisense,
5'-CCCCACCCTCTGTGACATTC-3'; uc.48+ (231 bp): sense,
5'-GCAAACTGGATGAGGAT-3' and antisense, 5'-GTAGTGCCACAAGGAGA-3';
β-actin (240 bp): sense, 5'-TAAAGACCTCTATGCCAACACAGT-3' and
antisense, 5'-CACGATGGAGGGGCCGGACTCATC-3'. The PCR conditions used
to detect these genes and analyze the PCR products were as
described in the study by Wu et al (13).
Western blot analysis
Total protein was extracted using RIPA buffer
containing a protease/phosphatase inhibitor mixture (diluted 1:100;
Vazyme Biotech Co., Ltd.). The total protein concentration in the
supernatant was measured using a bicinchoninic acid assay (Beyotime
Institute of Biotechnology). The steps of the western blot analysis
for P2X7 receptor and (p-)ERK1/2 were performed as
described previously in the study of Wu et al (13).
Statistical analysis
All experiments were carried out in triplicate to
confirm the accuracy of the results and data are presented as the
mean ± standard deviation. The data of the abdominal cell
experiments were normalized to those of the control group.
Statistical analyses were carried out using SPSS 11.5 software
(SPSS, Inc.). Statistical significance was determined by one-way
analysis of variance. The Tukey's test was used for comparison
between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of uc.48+ siRNA on the
nociceptive behavior of DM mice
The MWTs and TWLs of the mice were measured. Prior
to the STZ injection (at the end of week 6), no significant
differences in MWT were detected among the four groups, while the
TWLs of the mice in the DM groups were lower than those of the
control group. Following STZ injection (at the end of week 7), the
MWTs and TWLs in the DM groups were lower than those of the control
group. Following siRNA injection (at the end of week 8), the MWTs
and TWLs (Fig. 1) in the DM +
uc.48+ si group were higher than those of the DM group.
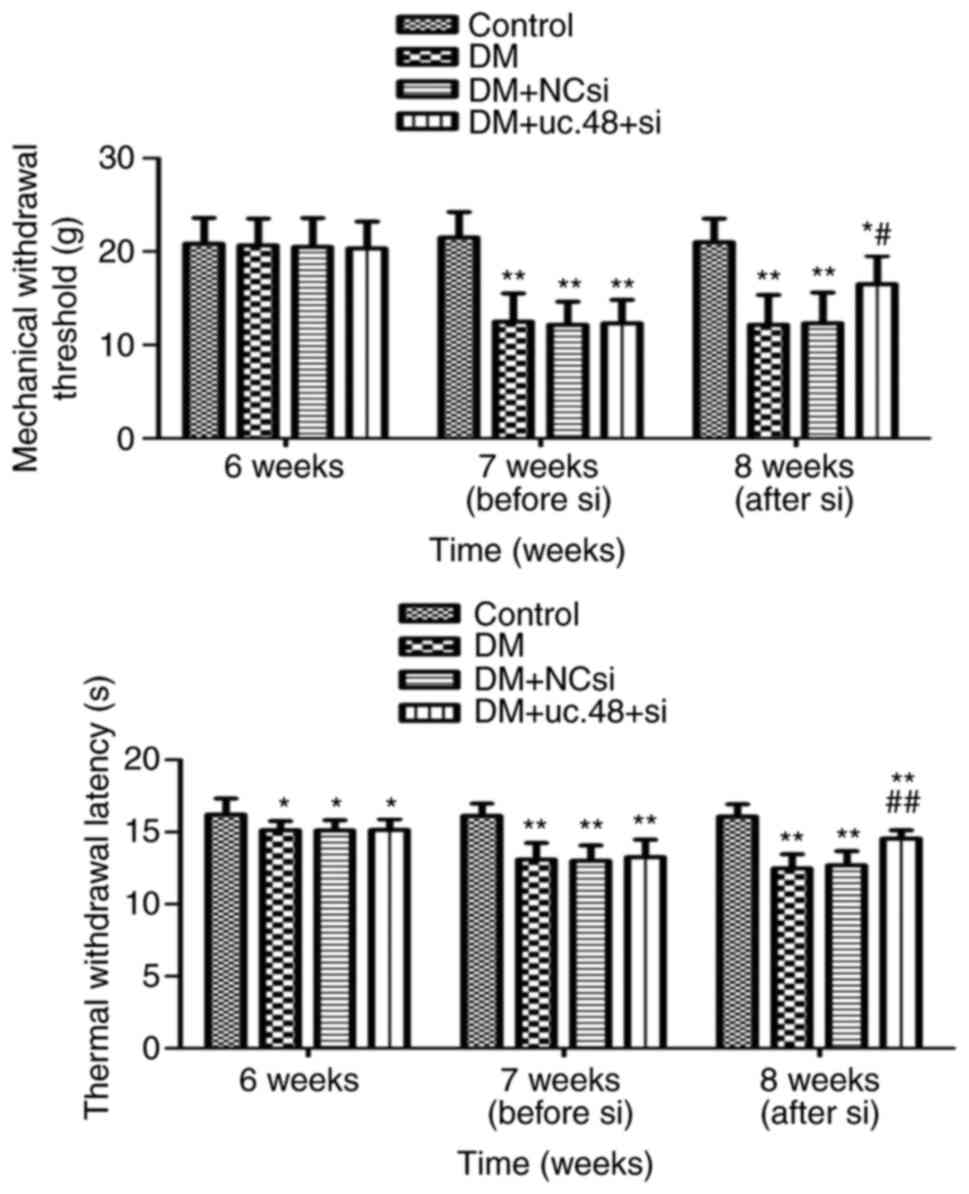 | Figure 1Effects of uc.48+ siRNA on the
nociceptive behavior of type 2 diabetic mice. The behavior of the
mice was assessed using MWT (upper panel) and TWL (lower panel)
assays. At 6W, no significant differences in MWT were detected
among the four groups, while the TWLs of the DM, DM + NCsi and DM +
uc.48+ si groups were lower than that of the control group. At 7W,
the MWTs and TWLs of the DM, DM + NCsi and DM + uc.48+ si groups
were lower compared with those of the control group. At 8W, the MWT
and TWL of the DM + uc.48+ si group were higher than those of the
DM and DM + NCsi groups. n=6/group. *P<0.05,
**P<0.01 vs. the control group;
#P<0.05, ##P<0.01 vs. the DM group.
siRNA, small interfering RNA; uc.48+ si, uc.48+ siRNA; NCsi,
scrambled control siRNA; MWT, mechanical withdrawal threshold; TWL,
thermal withdrawal latency; 6W, 6 weeks (prior to STZ injection);
7W, 7 weeks (following STZ injection); 8W, 8 weeks (after siRNA
injection); DM, diabetes mellitus; STZ, streptozotocin. |
These results reveal that the MWTs and TWLs in the
DM group were significantly decreased compared with those of the
control group. However, such effects were significantly attenuated
following the injection of uc.48+ siRNA. Moreover, the TWL appeared
to be significantly affected by a high-sugar and high-fat diet
while the MTL was not.
Effects of uc.48+ siRNA on the HR and
blood pressure of DM mice
The effects of uc.48+ siRNA treatment on HR and
blood pressure are shown in Table
I. The HR of the DM group was significantly increased compared
with that of the control group. However, no significant differences
were noted in HR between the DM, DM + NCsi and DM + uc.48+ siRNA
groups following uc.48+ siRNA injection. The SBP and DBP in the DM
group were significantly increased compared with those of the
control group. However, these changes were significantly diminished
following uc.48+ siRNA injection (Table
I).
 | Table IEffects of uc.48+ siRNA on the heart
rate, SBP and DBP of DM model mice. |
Table I
Effects of uc.48+ siRNA on the heart
rate, SBP and DBP of DM model mice.
| | Group |
|---|
| Parameter | Control | DM | DM + NCsi | DM + uc.48+ si |
|---|
| Heart rate |
|
6 weeks | 579.80±44.68 | 617.43±37.08 | 617.83±58.58 | 620.15±40.65 |
|
7 weeks | 572.63±46.50 |
636.37±44.46a |
637.62±57.09a |
635.03±52.45a |
|
8 weeks | 568.08±52.03 |
636.93±47.73a |
634.65±58.10a | 601.62±59.16 |
| SBP |
|
6 weeks | 112.15±14.53 |
140.55±16.17b |
140.50±20.08b |
141.50±16.21b |
|
7 weeks | 115.42±11.91 |
146.45±16.83b |
147.75±15.91b |
146.83±14.73b |
|
8 weeks | 115.33±12.89 |
141.20±16.21b |
142.83±11.63b |
122.83±11.63c |
| DBP |
|
6 weeks | 75.57±5.86 | 85.27±11.07 | 86.50±8.04 | 85.67±9.69 |
|
7 weeks | 76.37±8.78 |
90.43±8.47a |
90.33±7.84a |
89.50±8.83a |
|
8 weeks | 75.03±6.80 |
89.73±6.37b |
89.27±8.98b |
80.00±7.44c |
Effects of uc.48+ siRNA on the
cytokine levels of DM mice
The serum TNF-α and IL-1β concentrations of the DM
group were significantly higher compared with those of the control
group, and these increases were significantly attenuated following
the injection of uc.48+ siRNA. The serum IL-10 concentration of the
DM group was significantly decreased compared with the
corresponding concentration in the control group, whereas uc.48+
siRNA injection restored the IL-10 concentration to its initial
levels (Fig. 2).
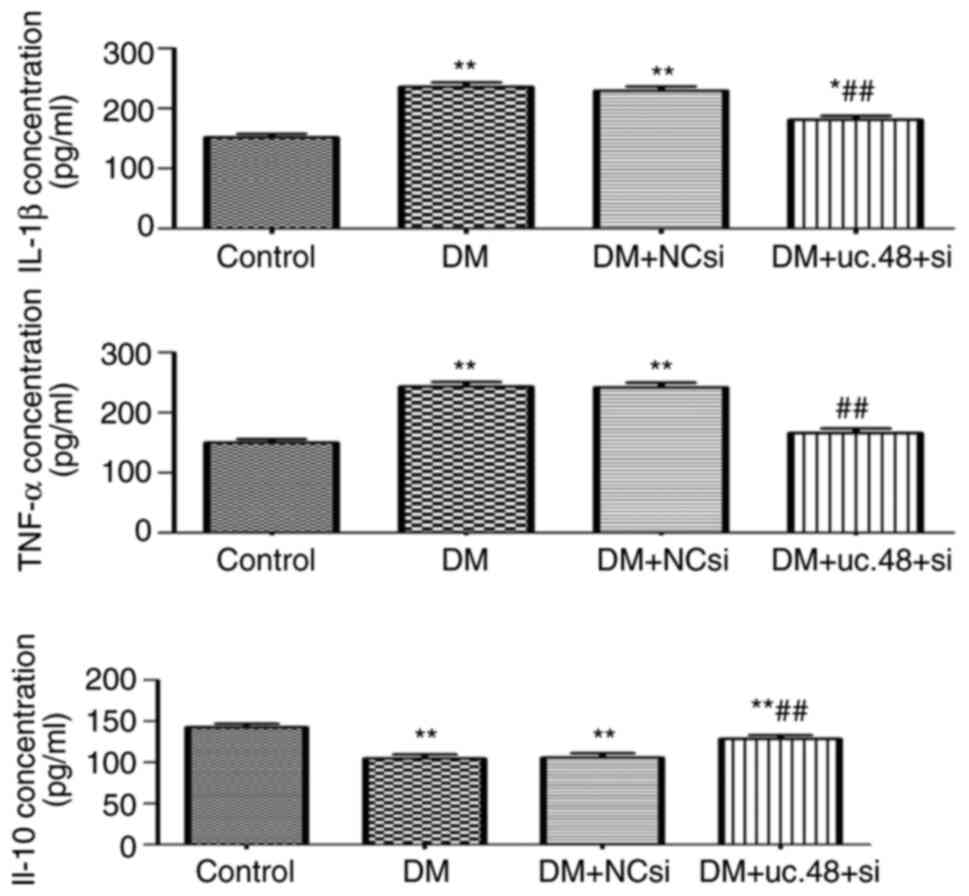 | Figure 2Effects of uc.48+ siRNA on the
cytokine levels of type 2 diabetic mice. The serum expression
levels of the cytokines IL-1β, TNF-α and IL-10 were evaluated by
ELISA. IL-1β and TNF-α levels were increased significantly in the
DM group (235.78±17.84 and 243.07±18.97 pg/ml, respectively)
compared with those in the control group (151.52±14.51 and
149.52±13.84 pg/ml, respectively), but were significantly decreased
by uc.48+ siRNA injection (to 181.30±15.38 and 165.75±19.44 pg/ml,
respectively). The IL-10 expression levels demonstrated opposite
trends compared with those observed for TNF-α and IL-1β. n=6/group.
*P<0.05, **P<0.01 vs. the control
group; ##P<0.01 vs. the DM group. siRNA, small
interfering RNA; uc.48+ si, uc.48+ siRNA; NCsi, scrambled control
siRNA; DM, diabetes mellitus; IL, interleukin; TNF, tumor necrosis
factor. |
Morphological identification and
viability of abdominal cells
The scattered distribution, variable size, irregular
shape and strong refractive index of the adherent abdominal cells
were observed by microscopy (Fig.
3A). Following Wright-Giemsa staining, these cells exhibited
the morphological features of macrophages. They were oval, round or
irregularly shaped. They possessed abundant cytoplasm with
irregular margins and pseudopodia and often exhibited single
eccentric nuclei (Fig. 3B). No
marked changes were noted with regard to the size, morphology and
number of abdominal cells following transfection with uc.48+
siRNA.
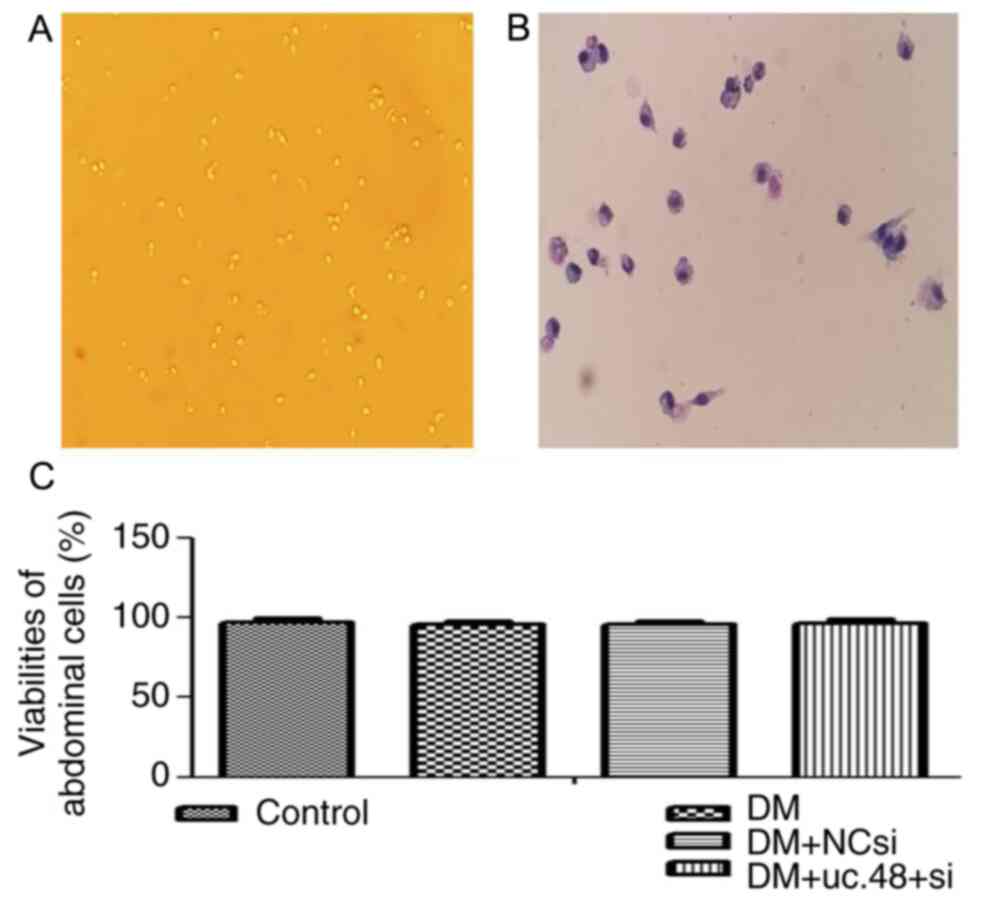 | Figure 3Morphological identification and
viabilities of mouse abdominal cells. (A) Surviving abdominal cells
(magnification, x10) and (B) abdominal cells following
Wright-Giemsa staining (magnification, x20). (C) Viabilities of the
adherent abdominal cells were 97.28±2.46, 95.77±2.29, 95.68±2.38
and 96.63±2.54 in the control, DM, DM + NC si and DM + NC si
groups, respectively. DM, diabetes mellitus; NCsi, scrambled
control siRNA; uc.48+ si, uc.48+ siRNA; siRNA, small interfering
RNA. |
The viabilities of the adherent abdominal cells in
all four groups were >95% as determined by the trypan blue
exclusion assay, and no significant difference was detected among
the groups (Fig. 3C).
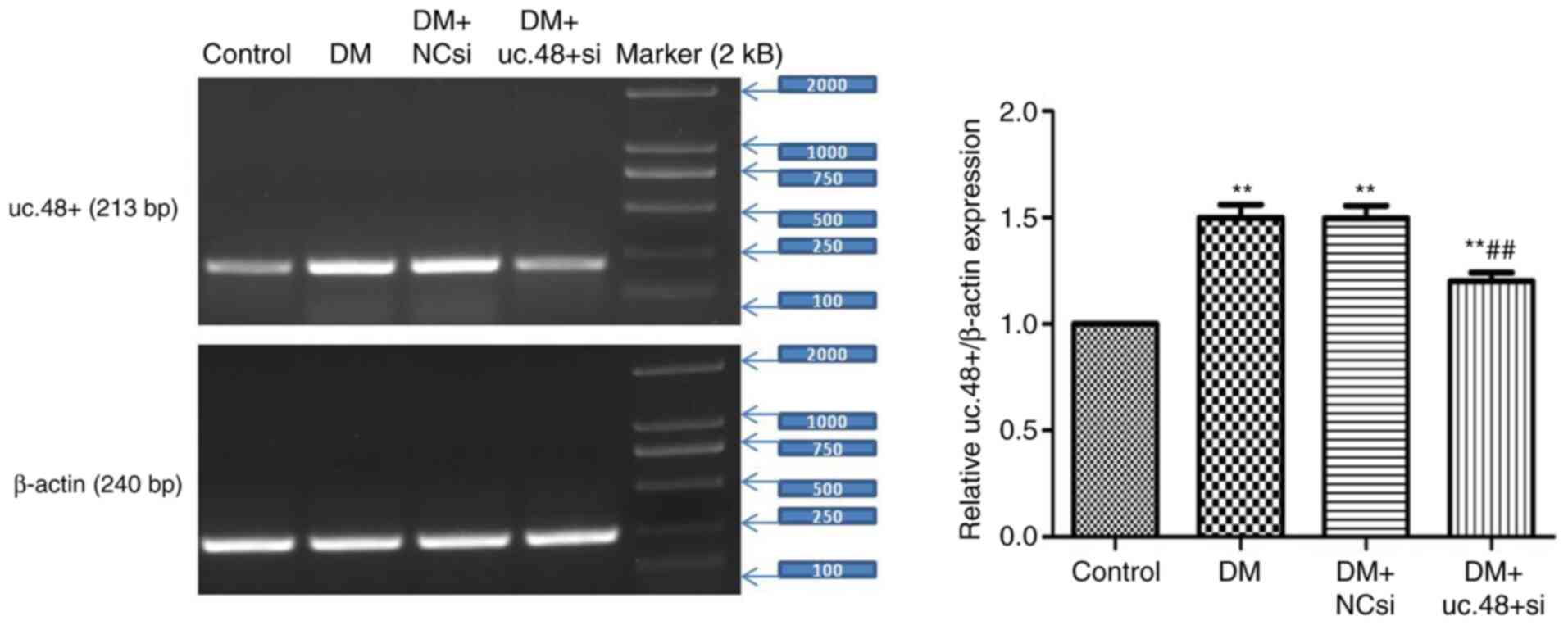
Changes in the uc.48+ expression
levels in the abdominal cells of DM mice following uc.48+ siRNA
treatment
The expression levels of uc.48+ in the abdominal
cells were significantly reduced following treatment with uc.48+
siRNA (1.20±0.10) compared with the DM group, whereas no
significant differences were noted between the DM (1.50±0.15) and
the DM + NCsi (1.49±0.14) groups (Fig.
4). These results indicate that the targeting of uc.48+ with
siRNA effectively suppressed the expression of uc.48+ in the
abdominal cells of DM model mice.
Changes in the expression levels of
P2X7 receptor mRNA and protein in the abdominal cells of
DM mice following uc.48+ siRNA treatment
As shown in Fig. 5,
the upregulated mRNA and protein levels of the P2X7
receptor were significantly decreased following uc.48+ siRNA
transfection in vivo (1.46±0.19 and 2.46±0.38, respectively)
compared with those in the DM (2.29±0.29 and 4.41±0.63,
respectively) group. No significant differences were detected
between the DM and the DM + NCsi groups.
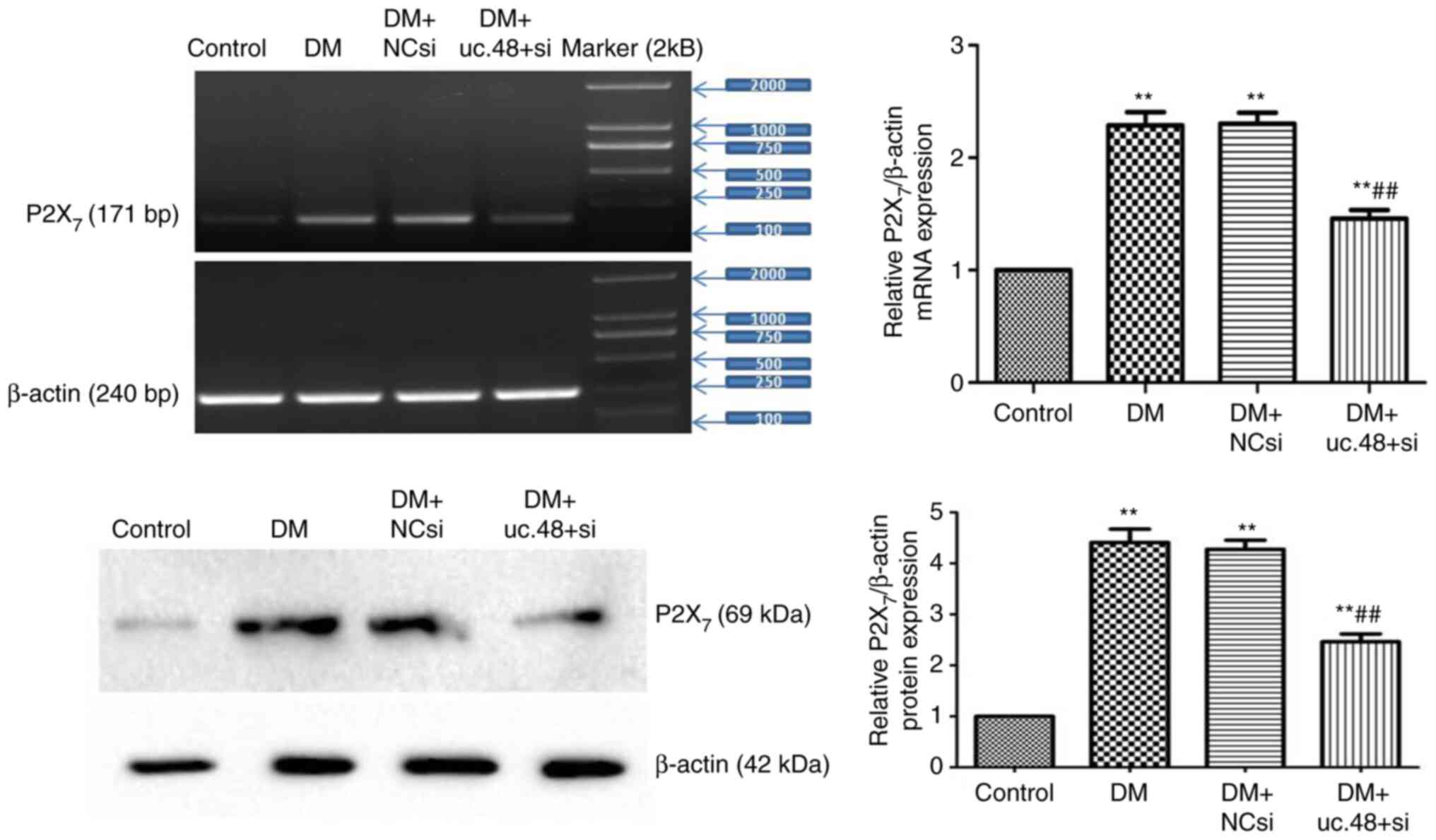 | Figure 5Changes in the expression levels of
P2X7 receptor mRNA and protein in the abdominal cells of
type 2 diabetic mice following uc.48+ siRNA treatment. The mRNA and
protein levels (top panel and bottom panel, respectively) of the
P2X7 receptor were downregulated in the uc.48+ siRNA
group (1.46±0.19 and 2.46±0.38, respectively) as determined by
RT-PCR and western blotting, respectively, following transfection
of uc.48+ siRNA in vivo, while no significant differences
were detected between the DM (2.29±0.29 and 4.41±0.63,
respectively) and the DM + NCsi groups (2.30±0.24 and 4.27±0.44,
respectively). n=6/group. **P<0.01 vs. the control
group; ##P<0.01 vs. the DM group. P2X7,
P2X purinoceptor 7; DM, diabetes mellitus; NCsi, scrambled control
siRNA; uc.48+ si, uc.48+ siRNA; siRNA, small interfering RNA;
RT-PCR, reverse transcription PCR. |
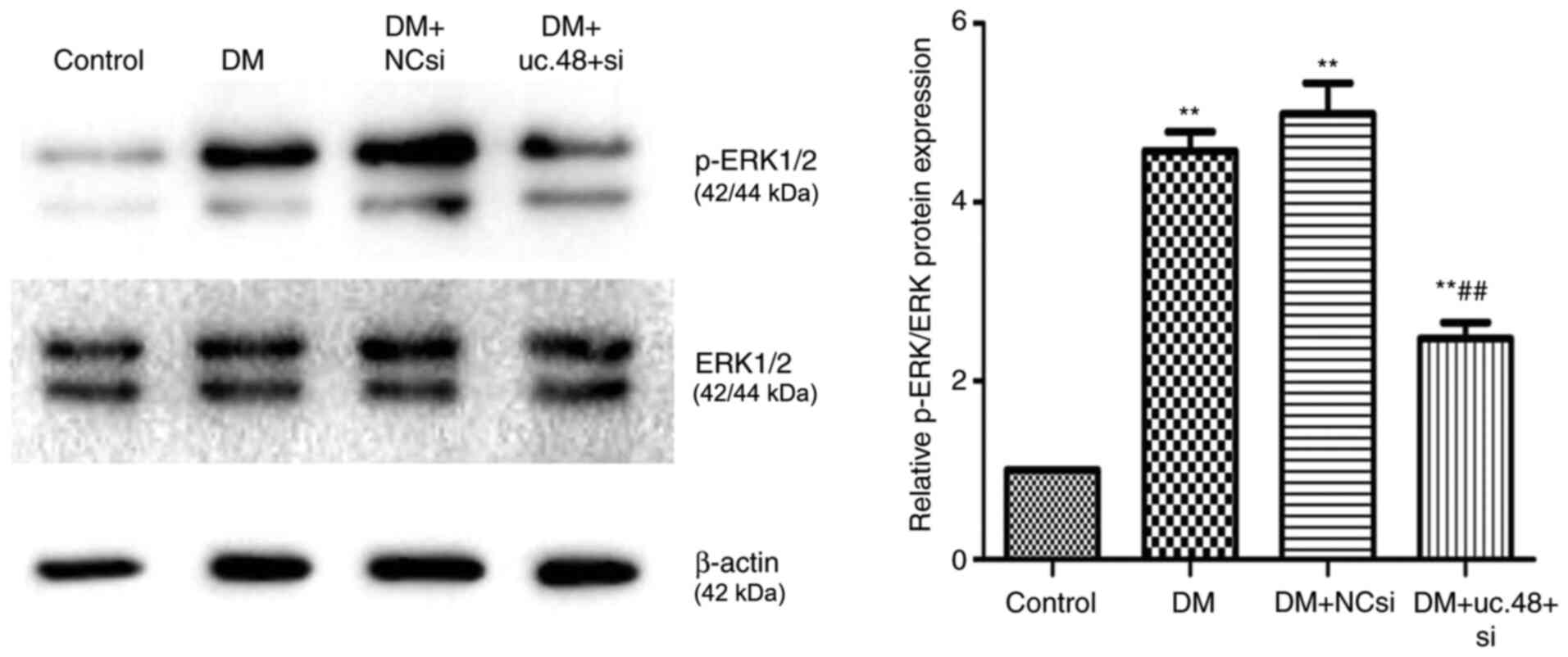
Changes in phosphorylated (p-)ERK1/2
levels in the abdominal cells of DM mice following uc.48+ siRNA
treatment
The levels of p-ERK1/2 were normalized to the total
ERK1/2 protein levels. The normalized p-ERK1/2 levels were
significantly increased in the DM group (4.57±0.52) compared with
the control group (1.00±0.00; Fig.
6). The knockdown of uc.48+ with uc.48+ siRNA (2.47±0.44)
significantly decreased the ratio of p-ERK1/2 to total ERK1/2
compared with the DM group, while scrambled siRNA (4.99±0.85)
exhibited no significant effects on p-ERK levels (Fig. 6).
Discussion
Diabetes is a considerable global health problem and
has been classified as a major disease that requires prevention and
control by the World Health Organization (21). The etiology of T2DM has not been
fully clarified. Previous studies have shown that diabetic
autonomic neuropathy, a complication of T2DM, can lead to cardiac
dysfunction (22), while a chronic
low-grade inflammatory response serves an important role in the
occurrence and development of T2DM (2). In the present study, it was observed
that the MWT and TWL of mice in the DM group were significantly
lower than those in the control group, indicating that DM damages
autonomic nerves and causes autonomic dysfunction, resulting in
significant increases in blood pressure and HR. Concomitantly, the
DM group displayed significantly higher serum levels of the
anti-inflammatory cytokines TNF-α and IL-1β in comparison with
those in the control group. The results presented in the current
study indicate that inflammatory reactions occurred in the T2DM
model mice, which exhibited effects on multiple systems, such as
the central nervous and cardiovascular systems, and resulted in
physiological dysfunction.
In vitro studies have shown that lncRNAs
exhibit important cellular functions (23-25).
Experiments using knockout animal models have confirmed that
multiple lncRNAs serve roles in disease pathogenesis (26,27). A
number of specific lncRNAs have been shown to participate in
pathological processes of the endocrine system, including DM
(28,29). The RT-PCR results of the present
study indicate that the expression levels of uc.48+ in the
abdominal cells of mice were significantly higher in the DM group
than in the control group. The results further revealed that the
MWT and TWL of mice with DM were significantly reduced following
uc.48+ siRNA injection, suggesting that uc.48+ siRNA treatment may
relieve diabetic neuropathic pain. Concomitantly, the blood
pressure and expression levels of the anti-inflammatory cytokines
IL-1β and TNF-α were also significantly decreased in DM model mice
following uc.48+ siRNA injection, demonstrating that the
downregulation of uc.48+ is associated with changes that relieve
the diabetic inflammatory state and cardiovascular disease.
Monocytes/macrophages play an important role in the
occurrence and development of T2DM, which is regarded as a type of
low-grade inflammation (4,5,30,31).
The in vitro results of our previous study using RAW264.7
macrophages revealed that uc.48+ siRNA is able to regulate immune
and inflammatory responses, thus influencing the course and outcome
of these effects, which are mediated by the P2X7
receptor (13). In the present
study, the role of uc.48+ in the pathological changes of T2DM were
investigated by monitoring the effects of uc.48+ siRNA on the
abdominal cells of a mouse model of T2DM. The results indicated
that the mRNA and protein expression levels of the P2X7
receptor and p-ERK1/2 level in the abdominal cells were
significantly increased in DM model mice compared with the control
group. However, these changes were significantly attenuated
following transfection with uc.48+ siRNA in vivo. The
experimental results indicate that uc.48+ serves an important role
in the pathological changes of T2DM via regulation of the function
of abdominal cells, while uc.48+ siRNA treatment may influence the
ERK signaling pathway via the P2X7 receptor. However, it
is unclear whether uc.48+ regulates the expression of cytokines
through the ERK signaling pathway, or the expression of cytokines
activates the ERK signaling pathway. Therefore, the specific
mechanism requires further study. In particular, more experiments
to verify that the effects of uc.48+ siRNA therapy are mediated by
the P2X7 receptor and ERK signaling are necessary. In
future studies, rescue experiments in which P2X7
receptor functions or ERK activity are blocked will be conducted to
elucidate the mechanism of uc.48+ siRNA treatment.
In conclusion, the present study demonstrated that
the uc.48+ expression levels of abdominal cells were significantly
increased in a mouse model of DM compared with those in
non-diabetic controls. Treatment of the mice with uc.48+ siRNA
ameliorated the blood pressure and neuropathological changes
associated with T2DM, and also downregulated the expression levels
of the P2X7 receptor. In addition, uc.48+ siRNA
regulated the inflammatory response and ERK signaling pathway in
the abdominal cells of the diabetic mice. These effects may be
mediated by the P2X7 receptor in T2DM. It is suggested
that uc.48+ may serve an important role in T2DM via regulation of
the P2X7 receptor, and thereby exert an effect on
pathological changes in blood pressure, neuropathological changes
and abdominal cells function.
Acknowledgements
The authors would like to thank Professor Shangdong
Liang [Department of Physiology, Medical College of Nanchang
University (Nanchang, China)] for guidance us during the present
study.
Funding
The present study was supported by a grant from the
National Natural Science Foundation of China (grant no.
81660144).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YN and HW were responsible for the conception and
design of the study. YN, HW, FW, MJ and QL acquired the data. HW
drafted the manuscript and YN revised the manuscript for important
intellectual content. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All procedures involving animals were approved by
the Animal Care and Use Committee of the Medical College of
Nanchang University (approval no. 2018 16).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Harris-Hayes M, Schootman M, Schootman JC
and Hastings MK: The role of physical therapists in fighting the
type 2 diabetes epidemic. J Orthop Sports Phys Ther. 50:5–16.
2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Prattichizzo F, De Nigris V, Spiga R,
Mancuso E, La Sala L, Antonicelli R, Testa R, Procopio AD, Olivieri
F and Ceriello A: Inflammageing and metaflammation: The yin and
yang of type 2 diabetes. Ageing Res Rev. 41:1–17. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bonnet F and Scheen AJ: Effects of SGLT2
inhibitors on systemic and tissue low-grade inflammation: The
potential contribution to diabetes complications and cardiovascular
disease. Diabetes Metab. 44:457–464. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Ward MG, Li G and Hao M: Apoptotic β-cells
induce macrophage reprogramming under diabetic conditions. J Biol
Chem. 293:16160–16173. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Saika F, Kiguchi N, Matsuzaki S, Kobayashi
D and Kishioka S: Inflammatory macrophages in the sciatic nerves
facilitate neuropathic pain associated with type 2 diabetes
mellitus. J Pharmacol Exp Ther. 368:535–544. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Maciel FR, Punaro GR, Rodrigues AM, Bogsan
CS, Rogero MM, Oliveira MN, Mouro MG and Higa EM: Immunomodulation
and nitric oxide restoration by a probiotic and its activity in gut
and peritoneal macrophages in diabetic rats. Clin Nutr.
35:1066–1072. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Breuillard C, Bonhomme S, Couderc R,
Cynober L and De Bandt JP: In vitro anti-inflammatory effects of
citrulline on peritoneal macrophages in Zucker diabetic fatty rats.
Br J Nutr. 113:120–124. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wu H, Nie Y, Xiong H, Liu S, Li G, Huang
A, Guo L, Wang S, Xue Y, Wu B, et al: P2X7 receptor expression in
peripheral blood monocytes is correlated with plasma C-reactive
protein and cytokine levels in patients with type 2 diabetes
mellitus: A preliminary report. Inflammation. 38:2076–2081.
2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Adinolfi E, Giuliani AL, De Marchi E,
Pegoraro A, Orioli E and Di Virgilio F: The P2X7 receptor: A main
player in inflammation. Biochem Pharmacol. 151:234–244.
2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bhat SA, Ahmad SM, Mumtaz PT, Malik AA,
Dar MA, Urwat U, Shah RA and Ganai NA: Long non-coding RNAs:
Mechanism of action and functional utility. Noncoding RNA Res.
1:43–50. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Dinescu S, Ignat S, Lazar AD, Constantin
C, Neagu M and Costache M: Epitranscriptomic signatures in lncRNAs
and their possible roles in cancer. Genes (Basel).
10(52)2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wu B, Zhang C, Zou L, Ma Y, Huang K, Lv Q,
Zhang X, Wang S, Xue Y, Yi Z, et al: LncRNA uc.48+ siRNA improved
diabetic sympathetic neuropathy in type 2 diabetic rats mediated by
P2X7 receptor in SCG. Auton Neurosci. 197:14–18. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wu H, Wen F, Jiang M, Liu Q and Nie Y:
LncRNA uc.48+ is involved in the diabetic immune and inflammatory
responses mediated by P2X7 receptor in RAW264.7 macrophages. Int J
Mol Med. 42:1152–1160. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Dos Anjos Cassado A: F4/80 as a major
macrophage marker: The case of the peritoneum and spleen. Results
Probl Cell Differ. 62:161–179. 2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Jain R, Watson U, Vasudevan L and Saini
DK: ERK activation pathways downstream of GPCRs. Int Rev Cell Mol
Biol. 338:79–109. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ma L, Zhang S and Du M: Cordycepin from
Cordyceps militaris prevents hyperglycemia in
alloxan-induced diabetic mice. Nutr Res. 35:431–439.
2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang J, Qiu H, Huang J, Ding S, Huang B,
Wu Q and Jiang Q: Establishment of a diabetic myocardial
hypertrophy model in Mus musculus castaneus mouse. Int J Exp
Pathol. 99:295–303. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liu S, Zou L, Xie J, Xie W, Wen S, Xie Q,
Gao Y, Li G, Zhang C, Xu C, et al: LncRNA NONRATT021972 siRNA
regulates neuropathic pain behaviors in type 2 diabetic rats
through the P2X7 receptor in dorsal root ganglia. Mol Brain.
9(44)2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Adan A, Kiraz Y and Baran Y: Cell
proliferation and cytotoxicity assays. Curr Pharm Biotechnol.
17:1213–1221. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang S, Xu H, Zou L, Xie J, Wu H, Wu B, Yi
Z, Lv Q, Zhang X, Ying M, et al: LncRNA uc.48+ is involved in
diabetic neuropathic pain mediated by the P2X3 receptor in the
dorsal root ganglia. Purinergic Signal. 12:139–148. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Begum M, Lewison G, Sommariva S, Ciani O,
Tarricone R and Sullivan R: European diabetes research and its
funding, 2002-2013. Diabet Med. 34:1354–1360. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Tarquini R, Lazzeri C, Pala L, Rotella CM
and Gensini GF: The diabetic cardiomyopathy. Acta Diabetol.
48:173–181. 2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Andersen RE and Lim DA: Forging our
understanding of lncRNAs in the brain. Cell Tissue Res. 371:55–71.
2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Dong Y, Yoshitomi T, Hu JF and Cui J: Long
noncoding RNAs coordinate functions between mitochondria and the
nucleus. Epigenetics Chromatin. 10(41)2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Lennox KA and Behlke MA: Cellular
localization of long non-coding RNAs affects silencing by RNAi more
than by antisense oligonucleotides. Nucleic Acids Res. 44:863–877.
2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chen X, Yan CC, Zhang X and You ZH: Long
non-coding RNAs and complex diseases: From experimental results to
computational models. Brief Bioinform. 18:558–576. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Magagula L, Gagliardi M, Naidoo J and
Mhlanga M: Lnc-ing inflammation to disease. Biochem Soc Trans.
45:953–962. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
He X, Ou C, Xiao Y, Han Q, Li H and Zhou
S: LncRNAs: Key players and novel insights into diabetes mellitus.
Oncotarget. 8:71325–71341. 2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Li X, Zhao Z, Gao C, Rao L, Hao P, Jian D,
Li W, Tang H and Li M: The diagnostic value of whole blood lncRNA
ENST00000550337.1 for pre-diabetes and type 2 diabetes mellitus.
Exp Clin Endocrinol Diabetes. 125:377–383. 2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Alvarado-Vázquez PA, Grosick RL,
Moracho-Vilrriales C, Ward E, Threatt T and Romero-Sandoval EA:
Cytokine production capabilities of human primary monocyte-derived
macrophages from patients with diabetes mellitus type 2 with and
without diabetic peripheral neuropathy. J Pain Res. 12:69–81.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Dhananjayan K, Gunawardena D, Hearn N,
Sonntag T, Moran C, Gyengesi E, Srikanth V and Münch G: Activation
of macrophages and microglia by interferon-γ and lipopolysaccharide
increases methylglyoxal production: A new mechanism in the
development of vascular complications and cognitive decline in type
2 diabetes mellitus? J Alzheimers Dis. 59:467–479. 2017.PubMed/NCBI View Article : Google Scholar
|