Introduction
Sepsis is a systemic inflammatory response syndrome
caused by bacterial infection and a life-threatening organ
dysfunction syndrome. Globally, there are more than 18 million
cases of sepsis every year. Despite considerable progress in
anti-infection therapy and organ function support technology, the
mortality rate of sepsis remains as high as 30-70%. Approximately
14,000 individuals die from complications of sepsis worldwide every
day and it is estimated that 215,000 individuals die in the United
States every year (1). Statistical
data from the United States indicate that the mortality rate of
sepsis has exceeded that of ST-segment elevation myocardial
infarction and has become the main cause of death in noncardiac
patients in hospital intensive care units (2).
The underlying mechanism of sepsis is believed to
involve complex systemic inflammatory network effects, gene
polymorphisms, immune dysfunction, abnormal coagulation function,
tissue damage and abnormal reactions of the host to pathogenic
microorganisms and toxins from various infections. Under
homeostatic conditions, the vascular endothelium can balance the
levels of inflammatory mediators, the innate immune system and the
coagulation system to coordinate the response to inflammation in
the process of systemic inflammation, such as septic shock
(3). However, once endothelial
dysfunction is induced by sepsis, homeostasis imbalance, vascular
reactivity changes and tissue edema may occur (4). Radeva and Waschke (5) demonstrated that inflammatory mediators
could inhibit the signal transduction of the cAMP/Ras-related C3
botulinum toxin substrate 1 pathway and activate the Ras homolog
family member A signaling pathway, leading to endocytosis,
phosphorylation of endothelial adhesion junction proteins and
depolymerization of tight junctions, eventually causing the
destruction of the endothelial barrier. Moreover, Millar et
al (6) established that the
pulmonary vascular endothelium is a key regulator and coordinator
of acute respiratory distress syndrome (ARDS). While describing the
pathophysiological changes of the endothelium in patients with ARDS
in detail, two potential signaling pathways for endothelial
targeted therapy were explored: i) The sphingosine-1-phosphate
pathway; and ii) the angiopoietin (Ang)-Ang1 receptor protein
(Tie2) pathway (3). To the best of
our knowledge, significant progress in the development of targeted
drug therapy against these signaling pathways has notbeen made.
Therefore, exploring new therapeutic methods is the focus of
current research.
Dexmedetomidine (Dex) is an agonist of the
adrenergic α2 receptor. Clinically, Dex can decrease
sympathetic tone; therefore, it is used as a sedative and widely
applied to inhibit the release of norepinephrine, thus terminating
the propagation of pain signals (7). In addition to providing a good
sedative effect, preclinical and clinical studies suggest that Dex
has anti-inflammatory, antioxidative and stress control effects
(8-11).
However, no study has focused on the effects of Dex on endothelial
damage in sepsis. In the present study, the protective effects of
Dex on vascular endothelial injury in a cecal ligation and puncture
(CLP)-induced rat model of sepsis were explored through detection
of the expression levels of endothelial injury markers.
Materials and methods
Animals and group classification
A total of 36 male Sprague-Dawley rats aged at 6
weeks, weighing 200-220 g, were purchased from the Animal
Laboratory of Daping Hospital, Army Medical University. All the
procedures in the present study followed the principles of the
International Guidelines on Animal Welfare (12). The animal experiments were approved
by the Ethics Committee of Daping Hospital, Army Medical University
(Chongqing, China).
The rats were randomly divided into three groups (12
rats/group): the Sham group, the CLP group and the CLP plus Dex
intervention group (CLP + Dex group). Animals were caged with free
access to food and water and were maintained under a 12-h
light/dark cycle at a temperature of 22±1˚C and a humidity of 45%.
All experimental rats were anesthetized by intraperitoneal
injection of 400 mg/kg (0.8 ml of 10%) chloral hydrate (Shanghai
Biochempartner) for CLP, sham operation, exsanguination or sample
collection. Following chloral hydrate administration, no signs of
peritonitis, pain or discomfort were found.
Rat CLP model
The animal model of sepsis in rats was set up
according to the methods described by Rittirsch et al
(13). Briefly, after
intraperitoneal injection of 400 mg/kg (0.8 ml of 10%) chloral
hydrate the rats were pinned on an operating table and the skin of
the mid-lower abdomen of the rats was disinfected and incised. The
cecum was found to the lower-left of the incision, squeezed to the
distal part of the cecum and ligated at the distal end of the
ileocecal base. The length of the ligation was 50% of the total
length of the cecum. The ligated cecum was punctured with a needle
on the side with less vasculature between the ligation line and the
midpoint of the end of the cecum, and then some feces were squeezed
out to kept the puncture hole passable, as previously described
(14). The incision was finally
closed and rinsed with saline to prevent rat dehydration. The CLP
model was deemed to be accomplished with the manifesting signs of
sepsis, including hypothermia, tachycardia, tachypnea, absence of
grooming activities with resulting ruffled fur, reduced intake of
food and water, lethargic conditions, elevated wet/dry (W/D) ratio
of lung tissue and histological alterations. According to the
methods described by Chen et al (15) and Hu et al (9), the rats in the CLP + Dex group were
intraperitoneally injected with Dex (10 µg/kg; Jiangsu Hengrui
Pharmaceutical Co., Ltd.) 30 min prior to the operation. Another
dose of Dex (10 µg/kg) was administered 6 h after the operation.
Meanwhile, the rats in the sham and the CLP groups were
intraperitoneally injected with the equivalent volume of normal
saline. In the sham group, the abdominal cavity of the rats was
opened, the cecum kept outside of the abdominal cavity for 10 min
and then returned into the abdominal cavity, before the incision
was sutured layer by layer. The rats were then returned to their
cages and provided with water and standard chow ad libitum.
All rats were observed every 5 min for 12 h after the operation.
Euthanasia was implemented according to the following symptoms
indicating that the rats were moribund (16): Ruffled fur and piloerection, few or
no activities when provoked, no response to touch stimuli, labored
breathing with gasps and mostly or completely closed eyes when
provoked. The surviving rats were injected intraperitoneally with
400 mg/kg chloral hydrate (0.8 ml of 10%) at 12 h after the
operation. Blood samples (2.5 ml/rat) were collected from the
abdominal aorta and the rats were sacrificed by exsanguination,
before the lung samples were harvested for further analyses.
Plasma Ang1 and 2 detection with ELISA
kits
Blood samples (2.5 ml/rat) were collected from the
rat abdominal aorta. Plasma samples were obtained by collection of
the supernatant from fresh blood after centrifugation at 7,000 x g
and 4˚C for 10 min. The samples were frozen immediately at -80˚C.
The Ang1 and 2 levels in the thawed plasma samples were detected by
a Rat Ang1 ELISA kit (cat. no. EK1295) and a Rat Ang2 ELISA kit
(cat. no. EK1574) (Wuhan Boster Biological Technology, Ltd.).
Hematoxylin and eosin (H&E)
staining
The upper lobe of the right lung was fixed in 4%
paraformaldehyde at 4˚C for 24 h and dehydrated stepwise with
gradient alcohol concentrations, washed with xylene, and embedded
in paraffin. The lung tissue was sectioned at a 5-µm thickness and
stained with H&E as previously described (17).
W/D weight ratio of lung tissue
Rats were sacrificed 12 h after the operation and
the weight of the left lung tissue (wet weight) was measured by an
electronic balance. Subsequently, the lung tissue was dried in an
oven at 80˚C for 48 h and the dry weight was measured. The W/D
weight ratio was calculated as an index to indicate the degree of
pulmonary edema.
Western blot analysis
The middle lobe of the rat right lung was
immediately frozen in liquid nitrogen. Lung samples (~100 mg) were
then homogenized in radioimmunoprecipitation assay buffer (Beyotime
Institute of Biotechnology) containing protease inhibitors. Protein
concentrations were determined using BCA assay kits (Thermo Fisher
Scientific, Inc.; cat. no. 23227). For western blot analysis,
samples containing 25 µg of protein were loaded onto a 10% sodium
dodecyl sulfate-polyacrylamide gel, electrophoresed, transferred
onto a polyvinylidene difluoride membrane, blocked at room
temperature for 60 min in 5% skim milk and treated with primary
antibodies against VE-cadherin (1:1,000 dilution) or GAPDH (1:1,000
dilution) as a protein loading reference (Thermo Fisher Scientific,
Inc.; cat. nos. 36-1900 and MA5-15738-BTIN, respectively) at room
temperature for 2 h. After washing three times with Tris-buffered
saline containing Tween-20 (TBST), the membrane was incubated with
horseradish peroxidase-labeled secondary antibody (Thermo Fisher
Scientific, Inc.; cat. no. A27036; 1:5,000 dilution) at room
temperature for 1 h, and washed again three times with TBST.
Finally, the probed protein was visualized using enhanced
chemiluminescence staining (BeyoECL Plus; Beyotime Institute of
Biotechnology) on an Odyssey® CLx Infrared Imaging
System (LI-COR Biosciences). The results were analyzed by
densitometric measurement (ImageJ version 1.53a; National
Institutes of Health) with a gel imager and the relative expression
of VE-cadherin was normalized to GAPDH.
Statistical analysis
The experimental results were analyzed by using SPSS
software (version 24; IBM Corp.). Data are presented as the mean ±
standard deviation. One-way analysis of variance was used for
multiple-group comparisons followed by the Least Significant
Difference post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Dex relieves CLP-induced symptoms
Among the model rats, six from the CLP group and
three from the CLP + Dex group reached the humane endpoints of the
study. These rats were anesthetized by intraperitoneal injection of
400 mg/kg (0.8 ml of 10%) chloral hydrate and sacrificed by
exsanguination. All rats in the sham group behaved normally when
provoked, while rats in the CLP + Dex group showed an improved
appetite, slightly ruffled fur, more active responses to auditory
or touch stimuli, fewer periods of labored breathing and wider
opened eyes than those of the CLP group (data not shown).
Dex prevents CLP-induced pulmonary
edema
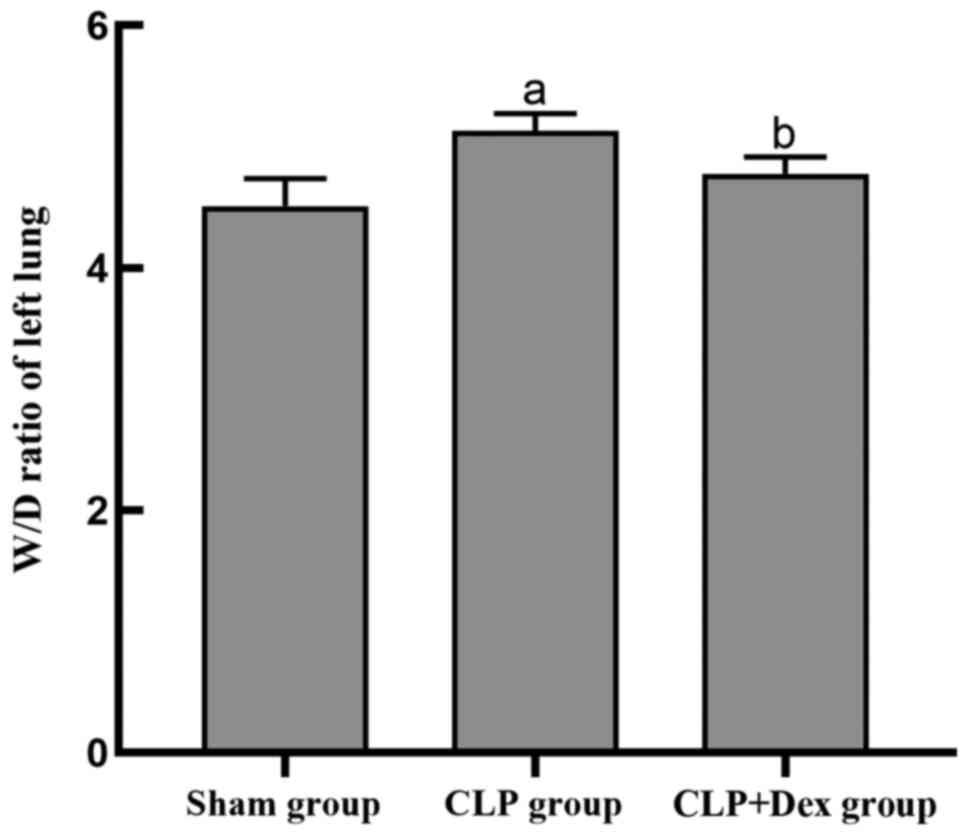
The W/D lung weight ratio of lung tissue reflects
the amount of lung edema (18). To
evaluate the protective effect of Dex on lung tissue edema, the W/D
ratio of the lung tissue in each group was determined. The results
indicated that the W/D ratio in the CLP group was significantly
higher than that in the sham group (P<0.05); however, Dex
treatment significantly reduced the increased ratio due to CLP when
compared with CLP alone (P<0.05; Fig. 1).
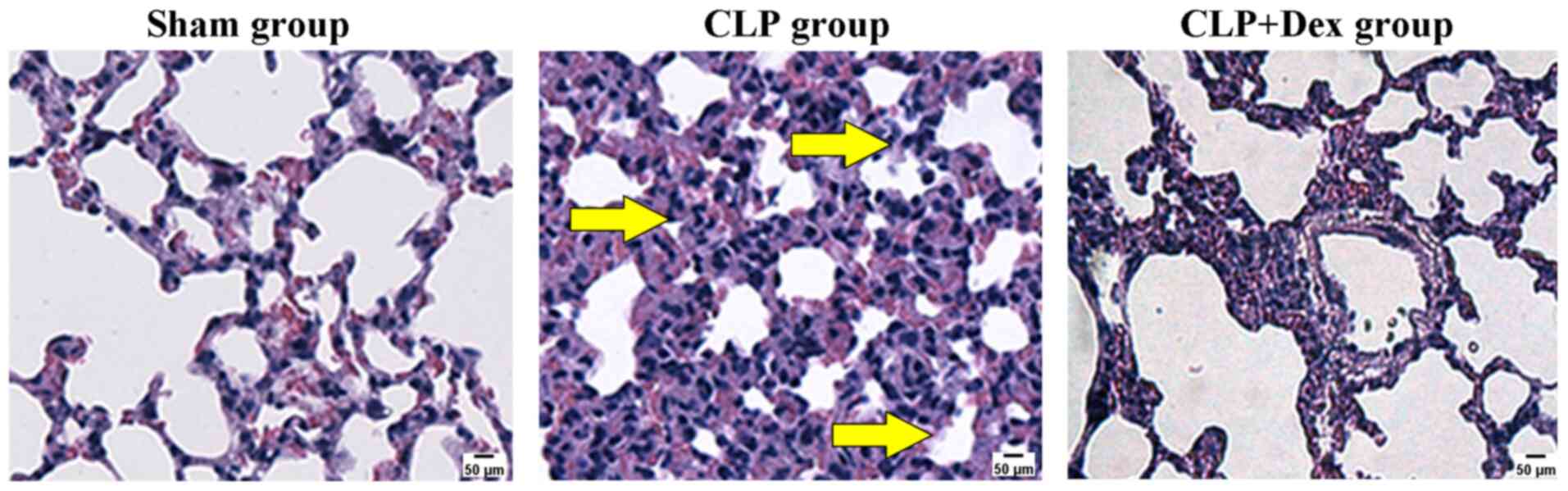
To confirm the edema-protective activity of Dex
histologically, the lung tissues of each group were stained with
H&E. At 12 h after CLP, a widened alveolar septum, a large
number of infiltrating white blood cells and congestion were
clearly observed in the lung of the CLP group when compared with
the sham group. The infiltrating white blood cells, widened
alveolar septum and edema fluid and congestion in the CLP + Dex
group were less obvious than in the CLP group (Fig. 2). These results indicated that CLP
significantly induced edema of the rat lung and that Dex protected
against CLP-induced pulmonary edema.
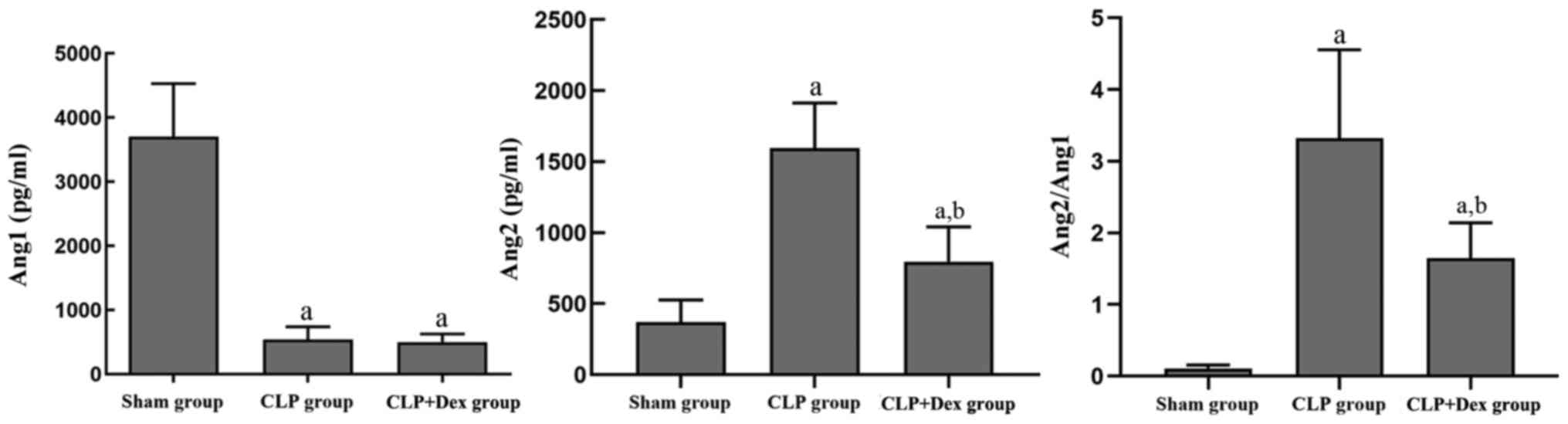
Dex pretreatment diminishes the
CLP-induced increase in the plasma Ang2 level and the Ang2/1
ratio
The plasma Ang1 and 2 levels of the rats in the
different groups were investigated. At 12 h after CLP treatment,
the plasma Ang1 level in the CLP group was significantly lower than
that in the sham group (P<0.01); however, no significant
difference was found between the CLP and the CLP + Dex groups
(P=0.85). However, the CLP group exhibited significantly increased
plasma Ang2 levels compared with those of the sham group
(P<0.01). The increase in plasma Ang2 induced by CLP was reduced
by pretreatment with Dex (P<0.01). Moreover, the ratio of Ang2/1
in the CLP group was significantly higher than that in the sham
group (P<0.01) and that in the CLP + Dex group was lower than
that in the CLP group (P<0.01). These data suggested that Dex
may reverse the CLP-induced elevation of Ang2 expression and the
Ang2/1 ratio (Fig. 3).
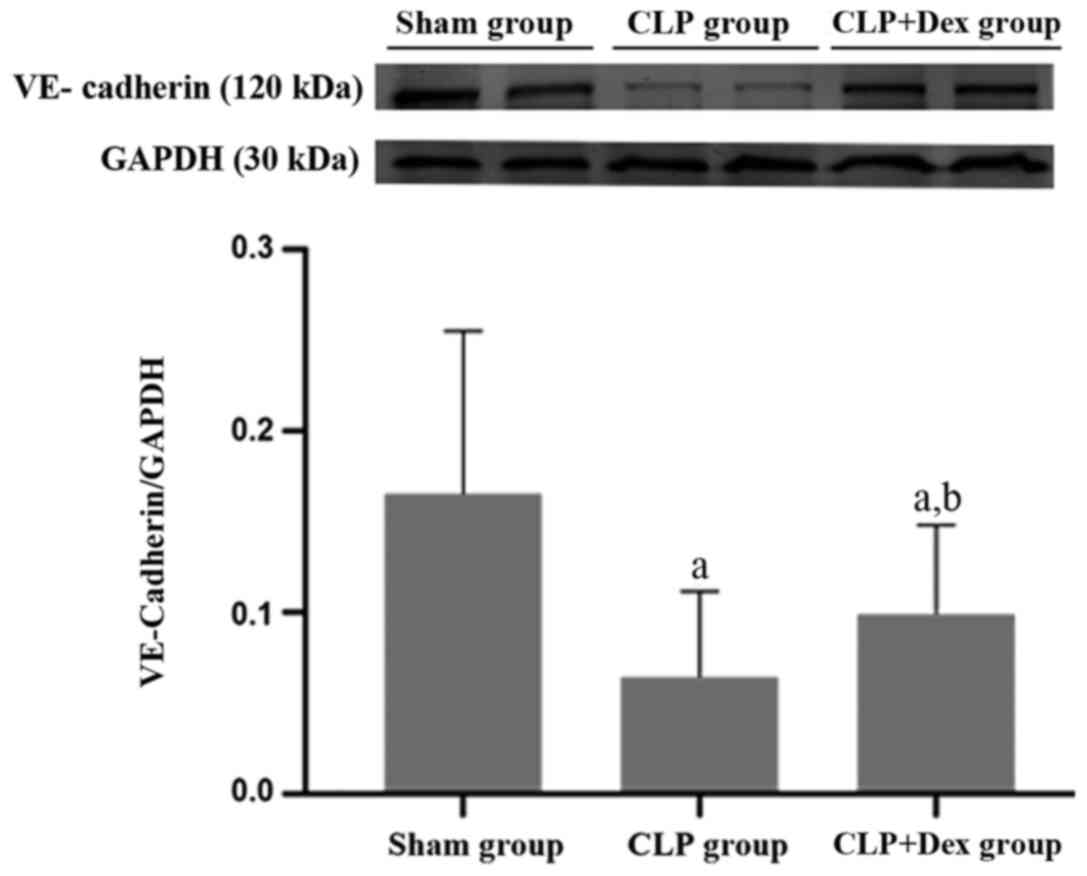
Dex alleviates the CLP-induced
decrease in VE-cadherin protein expression in lung tissue
To investigate whether VE-cadherin is involved in
CLP-induced sepsis, the protein level of VE-cadherin in the three
groups was examined by western blot analysis. In the CLP + Dex
group the protein expression of VE-cadherin in the lung tissue was
significantly lower than that of the sham group (P<0.05). In
addition, the level of VE-cadherin protein in the lung tissue of
the CLP + Dex group was significantly higher than that of the CLP
group (P<0.05; Fig. 4).
Discussion
To explore the complex molecular mechanism of
sepsis, numerous animal models have been developed so far. Among
these models, the most frequently applied one is the CLP model in
rodents, which typically presents with clinical symptoms of sepsis
and septic shock, including hypothermia, tachycardia and shortness
of breath (13). Buras et al
(19) reported that the CLP model
is the ‘gold standard’ for laboratory sepsis research. In the
present study, the advantages of the methods performed by Rittirsch
et al (13) and Liu et
al (14) were combined to
establish a CLP model that successfully mimics typical symptoms of
sepsis, such as hypothermia, tachycardia, tachypnea, absence of
grooming activities with resulting ruffled fur, reduced intake of
food and water and lethargic conditions. The sepsis model used in
the current study was verified by the W/D lung weight ratio
measurements and H&E staining of lung tissue. The pathological
alterations found in the present study were the same as those
described by Li et al (10).
This model provided a quick (12 h), simple, inexpensive and
reproducible method for the study of sepsis.
During the process of sepsis, an excessive immune
response involving the secretion of cytokines, chemokines and other
activated factors has a pronounced impact on the whole body
endothelium. The increased numbers of activated leukocytes adhere
to the endothelium to trigger endothelial coagulation and
destruction of the endothelial barrier. These alterations
eventually lead to extensive global tissue edema (6). Compared with other tissues, there is a
large amount of vasculature in lung tissue, providing abundant
vascular endothelium (6).
Therefore, the lung is severely affected by endothelial injury.
Damage to the pulmonary vascular endothelial tissue leads to the
accumulation of protein-rich fluid in the pulmonary interstitium,
infiltration of the fluid into the alveolar cavity and eventually
an imbalance of ventilation and blood flow (6). These pathological alterations further
decrease the arterial oxygen partial pressure and pulmonary
compliance, which are the common pathological alterations of ARDS
(3). The present data supported
these alterations and showed the signs of pulmonary edema,
including an increased W/D lung weight ratio, widened pulmonary
interstitium, and increased numbers of white and red blood cells,
in the alveoli of CLP-induced septic rats.
Dex is a highly selective α2 receptor agonist that
is widely used in the clinic. Its anti-inflammatory activity in
preclinical and clinical experiments has attracted the interest of
researchers (20). In addition to
anti-inflammatory and antioxidative stress activities, Yeh et
al (21) found that Dex could
ameliorate the intestinal mucosa, seromuscular layer, aggregated
lymphoid nodules and small vessel perfusion, while it reduced the
endocan level and the destruction of tight junction protein in
endotoxemic rats induced by lipopolysaccharide. Furthermore, the
results of Miranda et al (22) suggested that Dex could reduce the
rolling and adhesion of leukocytes to the endothelium in Syrian
hamsters with endotoxemia as well as improve the condition of
capillary perfusion. The results of the present study are
consistent with the previous findings that pretreatment with Dex
could significantly decrease the CLP-induced elevation of the W/D
lung ratio. In addition, Dex treatment reduced the CLP-induced
widening of the alveolar septum, infiltration of white blood cells,
edema fluid exudation and congestion in sepsis model rats. All of
the above results indicated that Dex may alleviate pulmonary edema
through protection of endothelial barrier integrity.
The dose of Dex used in the present study was chosen
according to previous studies (9,23,24). A
pilot study using Dex at a dose of 50 µg/kg was also conducted, but
some rats showed significantly decreased respiratory frequencies
and one of them manifested signs of hypoxia. Therefore, a dose of
10 µg/kg, at which all experimental rats showed no obvious side
effects, was chosen for the subsequent experiments. However, 10
µg/kg Dex surpasses the conventional dosage in clinical situations
(7). The equivalent dose between
rats and patients as well as the optimal dosage for critical
patients should be further investigated.
Ang1 is synthesized and secreted by cells around the
endothelium, while Ang2 is primarily produced by the endothelium
and stored in Weibel-Palade bodies. Ang1 acts to stabilize
endothelial adhesion through the Ang1-Tie2 signaling pathway, thus
protecting the endothelium. Once inflammatory factors and other
stimulatory molecules appear, Ang2 is rapidly released from
Weibel-Palade bodies (25) to
antagonize the anti-inflammatory effect of Ang1 by competing with
the Tie2 receptor, leading to endothelial barrier destruction,
activation of endothelial cells and promotion of vascular
inflammation (26). The Ang-Tie2
axis is one of the most widely studied pathways in the field of
endothelial barrier function regulation. Elevation of Ang2 levels
in plasma indicates progressive injury of endothelial cells, which
can predict mortality in sepsis-related ARDS patients (26). In sepsis, ARDS, and related cases,
the circulating Ang1 level has been reported to be 2-3 times less
than normal levels; in contrast, the plasma Ang2 levels are
increased by 5-20 times (27).
Therefore, the ratio of Ang2/1 can better reflect the severity of
endothelial damage than either Ang1 or 2 alone. In addition, Han
et al (28) found that the
circulating Ang2 level reached a peak at 12 h after CLP
stimulation. Therefore, stimulation with CLP for 12 h was carried
out in order to investigate the effect of Dex on the expression of
Ang2 and 1 as well as the Ang2/1 ratio in sepsis model rats in the
present study. The results of the present study demonstrated that
CLP stimulation significantly elevated the plasma Ang2 level and
the Ang2/1 ratio in sepsis model rats. Dex successfully reduced the
CLP-upregulated plasma Ang2 level and the Ang2/1 ratio in sepsis
model rats.
VE-cadherin is a key molecule in the adhesion
junctions between vascular endothelial cells (5). Ang1 can bind and activate the Tie2
receptor to trigger downstream guanosine triphosphatase to release
phosphate, which stabilizes the cellular barrier through
upregulation of VE-cadherin in cellular junctions (5). In the present study, the VE-cadherin
levels in the lung tissue of the rats pretreated with Dex were
significantly greater compared with the CLP group, which may be
caused by downregulation of Ang2 expression by Dex through the
Ang-Tie2-VE-cadherin signaling pathway. Although Dex did not
directly affect Ang1 expression compared with CLP alone, the Ang2/1
ratio in the CLP + Dex group was significantly reduced. The reduced
ratio of Ang2/1 may reduce the phosphorylation and internalization
of VE-cadherin through the Ang1-Tie2-VE-cadherin signaling pathway,
ultimately reducing the damage of the endothelial barrier function
and alleviating pulmonary edema.
In the present study, the plasma Ang2 level in the
CLP group was not as high as reported previously by Ricciuto et
al (27). The reason for this
may be that rodents have a strong resistance to bacteria (19); therefore, the vascular endothelial
damage by CLP is not as obvious as that in humans. Furthermore,
species differences may be involved in the differences as well
(29). Further determination of the
suitable dosage of Dex in rats and reasonable estimation of the
equivalent dosage among different species requires additional
exploration. The results of the present study suggest that Dex
reduced the Ang2/1 ratio by decreasing the Ang2 level, which may
trigger Ang-Tie2 signaling to affect VE-cadherin expression and
improve endothelial barrier function. Sepsis is a highly
heterogeneous disorder with a complex pathogenesis, and endothelial
injury in sepsis involves abnormalities of inflammation (30), immunity (31), complement, coagulation (32), the endothelial system and multiple
signaling pathways (4). There
remain numerous regulatory factors and signaling pathways upstream
of VE-cadherin, which require further exploration to verify whether
Dex can play a role in other stable signaling pathways of
endothelial barrier function.
The present study explored the effects of Dex, a
high-affinity α-2 receptor agonist, on endothelial injury in a
CLP-induced sepsis model. The results demonstrated that Dex
attenuated the elevation of plasma Ang2 to reduce the ratio of
Ang2/1, which may have consequently mitigated the decrease in the
expression level of VE-cadherin. The present study had certain
limitations. The reasons why Dex reduced the plasma Ang2 level but
did not have an impact on the Ang1 level were not determined.
Additionally, considering that other signaling pathways contribute
to the maintenance of endothelial function, further investigations
are warranted to explore the specific mechanisms.
The results of the present study suggested that Dex
may decrease CLP-induced upregulation of Ang2 expression and
increase the CLP-induced decrease of VE-cadherin expression in a
sepsis rat model, and protect against pulmonary edema. These data
provide molecular evidence to support the clinical application of
Dex for the treatment of sepsis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PZ wrote the original draft, conceived and designed
the experiments and performed the animal experiment. JP
participated in the animal experiments and collected the data. YQR
performed the data analysis and contributed to the manuscript
preparation and data presentation. HZ contributed to the reagents,
materials, data analysis and the guidance of experimental
procedures. HY revised and edited the manuscript, as well as
provided the general supervision of the research group. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All the procedures of this study followed the
principles of the International Guidelines on Animal Welfare. The
animal experiments were approved by the Ethics Committee of Daping
Hospital, Army Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cecconi M, Evans L, Levy M and Rhodes A:
Sepsis and septic shock. Lancet. 392:75–87. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Singer M, Deutschman CS, Seymour CW,
Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche
JD, Coopersmith CM, et al: The third international consensus
definitions for sepsis and septic shock (sepsis-3). JAMA.
315:801–810. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Gotts JE and Matthay MA: Sepsis:
Pathophysiology and clinical management. BMJ.
353(i1585)2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Ince C, Mayeux PR, Nguyen T, Gomez H,
Kellum JA, Ospina-Tascón GA, Hernandez G, Murray P and De Backer D:
ADQI XIV Workgroup. The endothelium in sepsis. Shock. 45:259–270.
2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Radeva MY and Waschke J: Mind the gap:
Mechanisms regulating the endothelial barrier. Acta Physiol (Oxf):
222, 2018. doi: 10.1111/apha.12860.
|
|
6
|
Millar FR, Summers C, Griffiths MJ,
Toshner MR and Proudfoot AG: The pulmonary endothelium in acute
respiratory distress syndrome: Insights and therapeutic
opportunities. Thorax. 71:462–473. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Weerink MAS, Struys MMRF, Hannivoort LN,
Barends CRM, Absalom AR and Colin P: Clinical pharmacokinetics and
pharmacodynamics of dexmedetomidine. Clin Pharmacokinet.
56:893–913. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Meng L, Li L, Lu S, Li K, Su Z, Wang Y,
Fan X, Li X and Zhao G: The protective effect of dexmedetomidine on
LPS-induced acute lung injury through the HMGB1-mediated TLR4/NF-κB
and PI3K/Akt/mTOR pathways. Mol Immunol. 94:7–17. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hu H, Shi D, Hu C, Yuan X, Zhang J and Sun
H: Dexmedetomidine mitigates CLP-stimulated acute lung injury via
restraining the RAGE pathway. Am J Transl Res. 9:5245–5258.
2017.PubMed/NCBI
|
|
10
|
Li B, Li Y, Tian S, Wang H, Wu H, Zhang A
and Gao C: Anti-inflammatory effects of perioperative
dexmedetomidine administered as an adjunct to general anesthesia: A
meta-analysis. Sci Rep. 5(12342)2015.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wang XW, Cao JB, Lv BS, Mi WD, Wang ZQ,
Zhang C, Wang HL and Xu Z: Effect of perioperative dexmedetomidine
on the endocrine modulators of stress response: A meta-analysis.
Clin Exp Pharmacol Physiol. 42:828–836. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
International guidelines on animal
welfare. 156: 723-723, 2005.
|
|
13
|
Rittirsch D, Huber-Lang MS, Flierl MA and
Ward PA: Immunodesign of experimental sepsis by cecal ligation and
puncture. Nat Protoc. 4:31–36. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu X, Wang N, Wei G, Fan S, Lu Y, Zhu Y,
Chen Q, Huang M, Zhou H and Zheng J: Consistency and
pathophysiological characterization of a rat polymicrobial sepsis
model via the improved cecal ligation and puncture surgery. Int
Immunopharmacol. 32:66–75. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chen Q, Yi B, Ma J, Ning J, Wu L, Ma D, Lu
K and Gu J: α22-adrenoreceptor modulated FAK pathway induced by
dexmedetomidine attenuates pulmonary microvascular
hyper-permeability following kidney injury. Oncotarget.
7:55990–56001. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Mai SHC, Sharma N, Kwong AC, Dwivedi DJ,
Khan M, Grin PM, Fox-Robichaud AE and Liaw PC: Body temperature and
mouse scoring systems as surrogate markers of death in cecal
ligation and puncture sepsis. Intensive Care Med Exp.
6(20)2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Cox RA: Endothelin-1 and acute lung injury
in sheeps with smoke inhalation and burn injury[D]. Galveston: The
University of Texas,Graduate School of Biomedical Sciences: 18-19,
2003.
|
|
18
|
Jin Y, Yu G, Peng P, Zhang Y and Xin X:
Down-regulated expression of AQP5 on lung in rat DIC model induced
by LPS and its effect on the development of pulmonary edema. Pulm
Pharmacol Ther. 26:661–665. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Buras JA, Holzmann B and Sitkovsky M:
Animal models of sepsis: Setting the stage. Nat Rev Drug Discov.
4:854–865. 2005.PubMed/NCBI View
Article : Google Scholar
|
|
20
|
Cruz FF, Rocco PR and Pelosi P:
Anti-inflammatory properties of anesthetic agents. Crit Care.
21(67)2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yeh YC, Wu CY, Cheng YJ, Liu CM, Hsiao JK,
Chan WS, Wu ZG, Yu LC and Sun WZ: Effects of Dexmedetomidine on
intestinal microcirculation and intestinal epithelial barrier in
endotoxemic rats. Anesthesiology. 125:355–367. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Miranda ML, Balarini MM and Bouskela E:
Dexmedetomidine attenuates the microcirculatory derangements evoked
by experimental sepsis. Anesthesiology. 122:619–630.
2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wu Y, Liu Y, Huang H, Zhu Y, Zhang Y, Lu F
and Zhou C, Huang L, Li X and Zhou C: Dexmedetomidine inhibits
inflammatory reaction in lung tissues of septic rats by suppressing
TLR4/NF-κB pathway. Mediators Inflamm. 2013(562154)2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ma Y, Yu XY and Wang Y: Dose-related
effects of dexmedetomidine on immunomodulation and mortality to
septic shock in rats. World J Emerg Med. 9:56–63. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Moss A: The angiopoietin: Tie 2
interaction: A potential target for future therapies in human
vascular disease. Cytokine Growth Factor Rev. 24:579–592.
2013.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Parikh SM: Dysregulation of the
angiopoietin-Tie-2 axis in sepsis and ARDS. Virulence. 4:517–524.
2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ricciuto DR, dos Santos CC, Hawkes M,
Toltl LJ, Conroy AL, Rajwans N, Lafferty EI, Cook DJ, Fox-Robichaud
A, Kahnamoui K, et al: Angiopoietin-1 and 2 as clinically
informative prognostic biomarkers of morbidity and mortality in
severe sepsis. Crit Care Med. 39:702–710. 2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Han S, Lee SJ, Kim KE, Lee HS, Oh N, Park
I, Ko E, Oh SJ, Lee YS, Kim D, et al: Amelioration of sepsis by
TIE2 activation-induced vascular protection. Sci Transl Med.
8(335ra355)2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Poli-de-Figueiredo LF, Garrido AG,
Nakagawa N and Sannomiya P: Experimental models of sepsis and their
clinical relevance. Shock. 30 (Suppl 1):S53–S59. 2008.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chousterman BG, Swirski FK and Weber GF:
Cytokine storm and sepsis disease pathogenesis. Semin Immunopathol.
39:517–528. 2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
van der Poll T, van de Veerdonk FL,
Scicluna BP and Netea MG: The immunopathology of sepsis and
potential therapeutic targets. Nat Rev Immunol. 17:407–420.
2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Levi M and van der Poll T: Coagulation and
sepsis. Thromb Res. 149:38–44. 2017.PubMed/NCBI View Article : Google Scholar
|