Introduction
Acute lung injury (ALI) is a life-threatening
disease characterized by increased vascular permeability and
inflammation. Acute respiratory distress syndrome (ARDS) is a
serious form of ALI that is associated with multiple organ failure
and high mortality among patients in the intensive care unit
(1). ARDS may be caused by sepsis,
pneumonia, pancreatitis and trauma (2). Although pharmacological interventions
and ventilatory management are used to treat ARDS, 40% of patients
with ARDS die in hospitals (3).
There are no effective pharmacological therapies for ALI.
Therefore, it is urgent to identify valid therapeutic drugs for the
improvement of ALI treatment.
Lipopolysaccharide (LPS) has often been used in some
animal models of ALI to induce pulmonary inflammation (4). Membrane-bound Toll-like receptors
(TLRs) have been demonstrated to play an important role in the
innate immune system by recognizing specific damage-associated
molecular patterns (DAMPs) and pathogen-associated molecular
patterns (PAMPs) of invading microorganisms (5). TLR4 is widely expressed in various
immune cells. When LPS binds to TLR4, it contributes to the
activation of NF-κB, eventually leading to the production of
proinflammatory cytokines (6-8).
Many studies have shown that oxidative stress plays a major role in
LPS-induced ALI. ROS are mainly generated by NADPH oxidase (Nox),
which expands inflammation by activating downstream signal cascades
(9,10). A body of evidence indicates that LPS
can stimulate the production of ROS, which promotes diverse
intracellular responses via the NF-κB pathway (11).
Itaconate, as a derivate of the tricarboxylic acid
cycle, is derived from the decarboxylation of cis-aconitate
mediated by immunoresponsive gene 1 in the mitochondrial matrix
(12). It has been reported that
itaconate has a direct antimicrobial effect by inhibiting
isocitrate lyase (13). Itaconate
markedly alleviated skin inflammation in a mouse model of psoriasis
and decreased the production of proinflammatory mediators in
LPS-treated macrophages (14,15).
However, whether and how 4-OI plays a protective role in
LPS-induced ALI remains largely unknown. In the present study, the
protective effects of 4-OI against LPS-induced ALI were
investigated, as well as the mechanisms associated with
inflammation and oxidative stress in mice.
Materials and methods
Ethics statement
All animal procedures and experiments were approved
by the Institutional Animal Care and Use Committee of Ningxia
Medical University [registration no. SCXK (Ning) 2018-0025], where
experiments were performed in accordance with the Guide for the
Care and Use of Laboratory Animals published by the National
Institutes of Health, 8th edition (16).
Drugs and antibodies
4-OI was obtained from MedChemExpress. Polyclonal
antibodies against phosphorylated (p)-NF-κB p65 (cat. no. 3033;
1:1,000), NF-κB p65 (cat. no. 8242; 1:1,000), phospho-PI3K (cat.
no. 4228; 1:1,000), PI3K (cat. no. 4249; 1:1,000), phospho-Akt
(cat. no. 4060; 1:1,000), Akt (cat. no. 4691; 1:1,000), lamin B
(cat. no. 9087; 1:1,000) and β-actin (cat. no. 4970; 1:1,000),
anti-rabbit IgG, and horseradish peroxidase (HRP)-linked antibody
(cat. no. 7074; 1:10,000) were purchased from Cell Signaling
Technology, Inc. N-acetyl-L-cysteine (NAC) and the PI3K inhibitor
LY294002 were provided by Beyotime Institute of Biotechnology. LPS
(Escherichia coli 055:B5) was purchased from Sigma-Aldrich
(Merck KGaA).
Animals and treatment
Male C57BL/6 mice (20-25 g; 8 weeks old) were used
in this study. Mice (n=15) were randomly assigned into three groups
of five mice each: Control group, ALI group, and ALI + 4-OI group.
According to a previous study (17), mice were anaesthetized with 2%
sodium pentobarbital (80 mg/kg; Sigma-Aldrich; Merck KGaA) by
intraperitoneal (i.p.) injection. Mice were then treated with an
intratracheal (i.t.) injection of LPS (E. coli O111:B4; 5
mg/kg) in 50 µl saline. Mice in the ALI + 4-OI group received an
i.p. injection of 4-OI (25 mg/kg/dose) in
(2-hydroxypropyl)-β-cyclodextrin in phosphate-buffered saline (PBS)
or vehicle control 2 h before i.t. injection of LPS, as previously
described (18). Control animals
received the same volume of vehicle (vehicle group). Lung tissue
was harvested for subsequent experiments 12 h after LPS
administration. Mice were anesthetized with inhaled isoflurane
(1.5-4%) and euthanized by exsanguination through right ventricle
aspiration, followed by cervical dislocation.
Histological examination
Mice were sacrificed 12 h after LPS administration.
The left lungs of mice were harvested and fixed with 4%
paraformaldehyde at 25˚C for 24 h. After fixation, lung tissue was
embedded in paraffin and sectioned at 5-µm thickness. The sections
were stained with hematoxylin at 25˚C for 10 min and eosin
(H&E) at 25˚C for 3 min and visualized under a light microscope
(magnification, x200). Morphological changes and injury were scored
as follows: 0, no injury; 1, mild; 2, moderate; and 3, severe
injury. This scoring system was based on the presence of exudates,
hyperemia or congestion, infiltration of neutrophils, alveolar
hemorrhage, presence of debris, and cellular hyperplasia (19).
Lung wet/dry (W/D) weight ratios
The lung W/D ratio was calculated as an indicator of
pulmonary edema. The right upper lung of each mouse was excised for
the determination of wet weight and then placed in an 80˚C oven for
4 days and reweighed to determine its dry weight.
Bronchoalveolar lavage fluid (BALF)
acquisition and analysis
After the mice were sacrificed, the BALF was
harvested by three i.t. injections of 0.5 ml cooled PBS. The
harvested fluid was centrifuged for 10 min at 1500 x g at 4˚C.
Total cell, neutrophil and macrophage counts were counted with a
hemocytometer and Wright-Giemsa staining at 25˚C for 30 sec. The
supernatant was collected for total protein content detection and
cytokine examination.
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
Total RNA isolated from lung tissue using
TRIzol® (Applied Biosystems; Thermo Fisher Scientific,
Inc.) and reverse-transcribed with random hexamers and MultiScribe™
reverse transcriptase (Applied Biosystems; Thermo Fisher
Scientific, Inc.) at 50˚C for 15 min and then at 85˚C for 5 sec.
qPCR was performed using HiScript® II One Step qRT PCR
kit (Vazyme Biotech, Co., Ltd.) on a deep-well Real-Time PCR
Detection system (CFX96 Touch™; Bio-Rad Laboratories, Inc.) using
the following thermocycling conditions: Initial denaturation at
95˚C for 5 min, followed by 35 cycles of denaturation at 94˚C for
60 sec, annealing at 58˚C for 60 sec and extension at 72˚C for 50
sec. The relative expression level of the target gene was examined
using the comparative 2-∆∆Cq method (20). β-actin mRNA served as an internal
control. The following mouse primers were used: Interleukin (IL)-1β
forward, 5'-GGGCCTCAAAGGAAAGAATC-3' and reverse,
5'-TACCAGTTGGGGAACTCTGC-3'; tumor necrosis factor (TNF)-α forward,
5'-ACAGCAAGGGACTAGCCAGGAG-3' and reverse,
5'-GGAGTGCCTCTTCTGCCAGT-3'; IL-6 forward,
5'-CTGGGGATGTCTGTAGCTCA-3' and reverse, 5'-CTGTGAAGTCTCCTCTCCGG-3';
inducible nitric oxide synthase (iNOS) forward,
5'-TTTGTGCGAAGTGTCAGTGG-3' and reverse, 5'-AGAAACTTCGGAAGGGAGCA-3';
β-actin forward, 5'-GTGGACATCCGCAAAGAC-3' and reverse,
5'-AAAGGGTGTAACGCAACTA-3'.
Cell culture
RAW264.7 murine macrophage-like cells, purchased
from the CBCAS (The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences), were cultured in DMEM (Invitrogen;
Thermo Fisher Scientific, Inc.) containing 10% (v/v) fetal bovine
serum (FBS, Gibco; Thermo Fisher Scientific, Inc.) in a humidified
atmosphere of 5% CO2 at 37˚C. RAW264.7 cells were
pretreated with 4-OI (125 µM), NAC (10 mM), the PI3K inhibitor
LY294002 (25 µM) or vehicle control for 1 h and then stimulated
with LPS (1 µg/ml) at 37˚C for 30 min to detect the phosphorylated
levels of PI3K, AKT and p65 in LPS-treated macrophages and at 37˚C
for 24 h to quantify the concentration of TNF-α, IL-1β, and IL-6 in
macrophages. The dose and treatment duration of all drugs were
selected according to previous studies (21,22).
Reactive oxygen species
(ROS)-generation assay
After the mice were sacrificed, the BALF was
harvested and lysed by Ammonium-Chloride-Potassium Lysis buffer
(Beyotime Institute of Biotechnology). To detect ROS generation,
the sedimented cells were resuspended in PBS. Briefly, the cells
were incubated with 50 µM of DCFH-DA for 30 min at 37˚C in
darkness. DCF fluorescence intensities were measured by flow
cytometry. At the end of the treatment, the cells were incubated
with 10 µM dichloro-dihydrofluorescein diacetate (DCFH-DA; Beyotime
Institute of Biotechnology) for 1 h at 37˚C in the dark. ROS
generation in the cells was measured immediately using a
fluorescence microscope (magnification, x200; Nikon Corporation).
Myeloperoxidase (MPO) activity, malondialdehyde (MDA) content,
superoxide dismutase (SOD) activity, and glutathione (GSH) content
were also measured. The lung homogenate was dissolved in extraction
buffer to detect the levels of MPO, MDA, SOD and GSH using
commercially available assay kits (cat. nos. A044-1-1, A003-1-2,
A001-3-2 and A005-1-2, respectively; Nanjing Jiancheng
Bioengineering Institute), according to the manufacturer's
instructions.
Cytokine measurements
Cytokines in serum were measured using enzyme-linked
immunosorbent assay (ELISA). Commercially available ELISA kits
(IL-1β; cat. no. MLB00C; IL-6; cat. no. M6000B; TNF-α, cat. no.
MTA00B; R&D Systems, Inc.) were used to determine the levels of
IL-1β, TNF-α and IL-6, as previously described (23,24).
Extraction of nuclear and cytosolic
proteins
To extract nuclear and cytosolic proteins, cells
were washed and lysed with hypotonic buffer [20 mM Hepes (pH 8.0),
10 mM KCl, 1 mM EDTA, 1.5 mM MgCl2, 1 mM DTT, 1 mM
Na3VO4, 1 mM NaF, 1 mM PMSF and 1% (v/v)
protease inhibitor cocktail] on ice for 30 min. NP-40 (0.625%) was
added to cell lysates. The lysates were collected and centrifuged
at 13000 x g and 4˚C for 15 min. The supernatants served as the
cytosolic extract. The nuclear pellets were resuspended in cold
buffer for another 30 min and centrifuged at 15000 x g and 4˚C for
5 min, and the supernatants were collected as the nuclear
extract.
Western blotting
Proteins were extracted with RIPA Lysis Buffer
(Beyotime Institute of Biotechnology) from the cells with protein
concentration determined using bicinchoninic acid protein assay.
Equal amounts of protein (30 µg) were loaded onto 10%
SDS-polyacrylamide gels and transferred onto polyvinylidene
fluoride (PVDF) membranes. The blots were blocked with 5% dry milk
in Tris-buffered saline with 0.1% Tween-20 at 25˚C for 30 min and
incubated with the appropriate primary antibody overnight at 4˚C.
The blots were washed three times and then incubated with an
HRP-conjugated secondary antibody at 25˚C for 1 h. Blots were then
visualized using an enhanced chemiluminescence reagent (Bio-Rad
Laboratories, Inc.). Images were acquired using ImageQuant LAS 4000
mini (Cytvia).
Statistical analysis
All data are normally distributed and are expressed
as the means ± SEM. Differences among multiple groups were analyzed
by one-way ANOVA with Tukey's post hoc test. For the comparison of
lung scoring, Kruskal-Wallis test followed by Dunn's multiple
comparison test was used. P<0.05 was considered to indicate a
statistically significant difference.
Results
4-OI ameliorates lung tissue injury in
mice with LPS-induced ALI
To confirm the role of 4-OI in the progression of
LPS-induced ALI, cells were pretreated with a dose of 4-OI (25
mg/kg/dose; i.p.) before i.t. injection of LPS in mice. The results
demonstrated that LPS led to severe inflammatory histological
changes in lung tissues, including inflammatory cell infiltration,
damage to the alveolar wall and pulmonary congestion, which were
markedly reversed by 4-OI treatment (Fig. 1A). Moreover, compared with that in
the LPS group, the lung injury score was decreased in the 4-OI
treatment group (Fig. 1B). In
addition, 4-OI treatment significantly decreased the W/D ratios
(Fig. 1C) and the total protein
concentration in BALF (Fig. 1D)
compared with those in the LPS group, which are two indicators of
pulmonary edema.
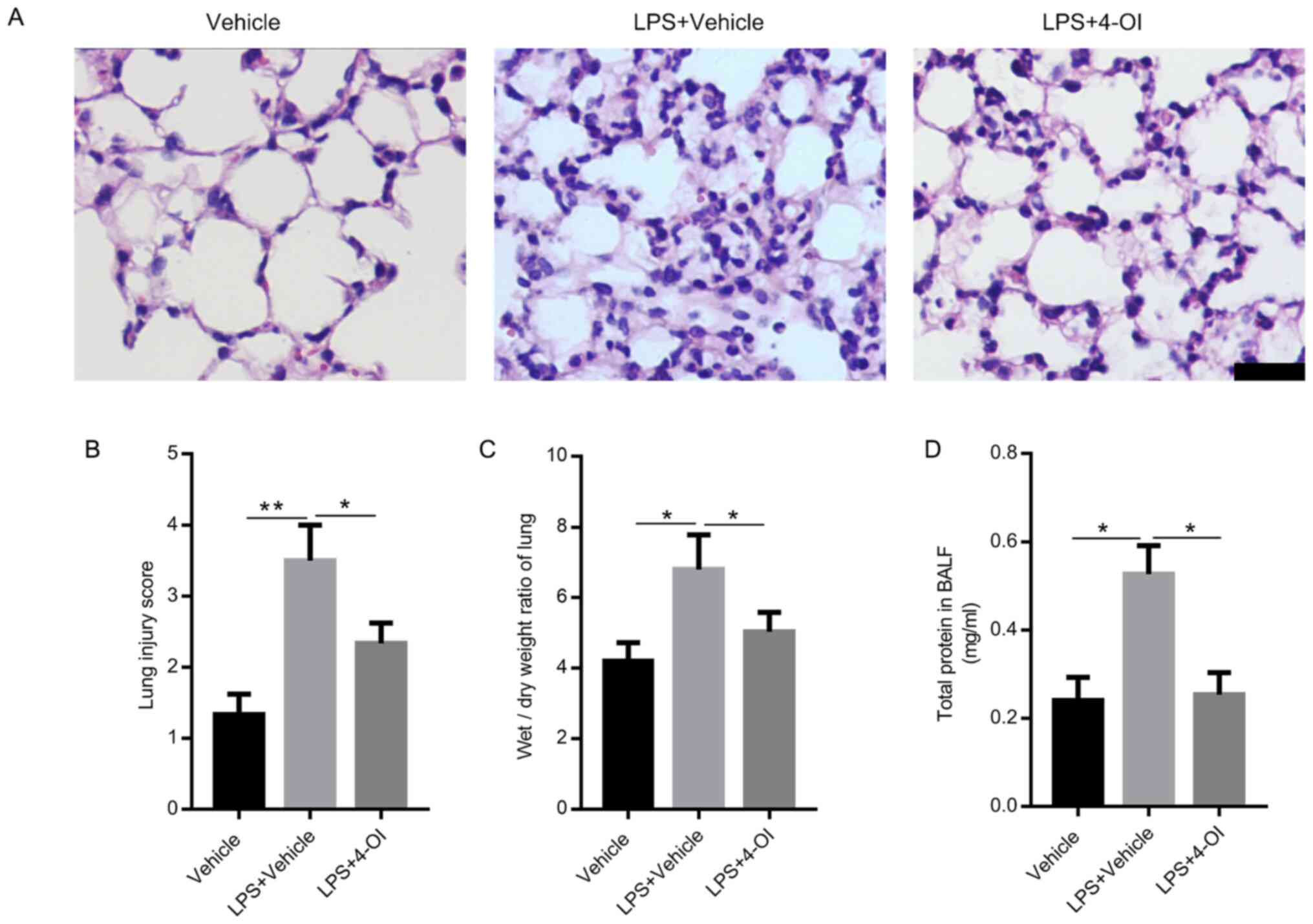 | Figure 14-OI alleviates lung injury induced by
LPS in mice. Treatment of mice with 4-OI at doses of 25 mg/kg was
administered intraperitoneally, and the control group received an
equivalent volume of vehicle [(2-hydroxypropyl)-β-cyclodextrin in
phosphate-buffered saline (PBS)] 2 h before saline or LPS injection
(5 mg/kg, intratracheal). Twelve hours later, the mice were
sacrificed. Hematoxylin and eosin staining (A, bar=100 µm) was used
to detect the lung histopathological changes. (B) Lung injury
scores were determined using four independent parameters, namely
alveolar congestion, hemorrhage, leukocyte infiltration and
alveolar wall thickness. The lung wet/dry ratio (C) and total
protein in BALF (D) were measured to determine lung permeability
(n=5). Data are expressed as the mean ± SEM. *P<0.05
and **P<0.01. 4-OI, 4-octyl itaconate; LPS,
lipopolysaccharides; BALF, bronchoalveolar lavage fluid. |
4-OI suppresses inflammatory responses
in LPS-induced ALI in mice
Subsequently, the effects of 4-OI on LPS-induced
intrapulmonary inflammatory responses were determined in mice. The
results showed that pretreatment with 4-OI decreased the gene
expression and secretion of IL-1β, TNF-α and IL-6 in the lungs
(Fig. 2A-F). Furthermore,
pretreatment with 4-OI notably decreased the number of total cells,
neutrophils and macrophages in BALF induced by LPS (Fig. 2G-I). In addition, pretreatment with
4-OI effectively decreased the LPS-induced increase in MPO
activity, which is a marker of neutrophil infiltration in lung
tissues (Fig. 2J). These results
illustrated that 4-OI prevented LPS-induced ALI by inhibiting
inflammatory responses.
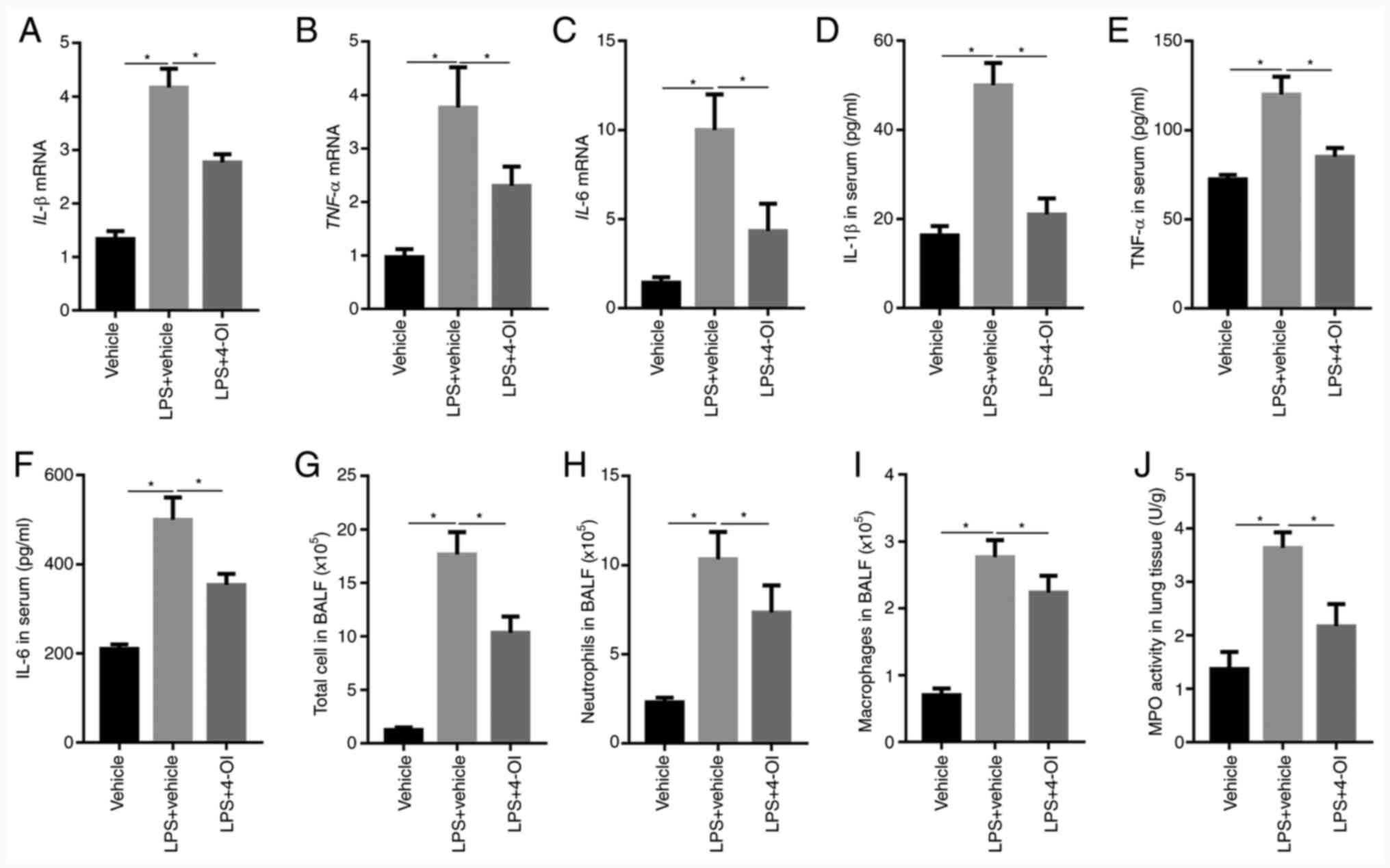 | Figure 24-OI inhibits LPS-induced inflammatory
responses in mice with acute lung injury. Treatment of mice with
4-OI at doses of 25 mg/kg was administered intraperitoneally, and
the control group received an equivalent volume of vehicle
[(2-hydroxypropyl)-β-cyclodextrin in phosphate-buffered saline
(PBS)] 2 h before saline or LPS injection (5 mg/kg, intratracheal).
Twelve hours later, IL-1β (A), TNF-α (B), and IL-6 (C) mRNA levels
in the lungs were determined by reverse transcription-quantitative
PCR (n=5). IL-1β (D), TNF-α (E), and IL-6 (F) protein contents in
serum were determined by enzyme-linked immunosorbent assay (n=5).
Twelve hours later, total cells (G), neutrophils (H) and
macrophages (I) in BALF were assessed (n=5). MPO activity (J) in
lung tissue was determined (n=5). Data are expressed as the mean ±
SEM. *P<0.05. 4-OI, 4-octyl itaconate; LPS,
lipopolysaccharides; IL, interleukin; TNF, tumor necrosis factor;
BALF, bronchoalveolar lavage fluid; MPO, myeloperoxidase. |
4-OI decreases oxidative stress in
mice with LPS-induced ALI
Given that oxidative stress plays an important role
in ALI, whether 4-OI could decrease oxidative stress induced by LPS
was determined. The results showed that LPS increased ROS
generation in the BALF of ALI mice and MDA formation in the lung
tissue of ALI mice, which were inhibited by 4-OI treatment
(Fig. 3A and B). Since increased levels of SOD and GSH
could inhibit oxidative stress, the levels of SOD and GSH were
detected by ELISA. The results showed that LPS decreased the levels
of SOD and GSH in the lung tissue of ALI mice, and this effect was
significantly reversed by pretreatment with 4-OI (Fig. 3C and D). These results indicate that
pretreatment with 4-OI could decrease oxidative stress in the lung
tissue of ALI mice.
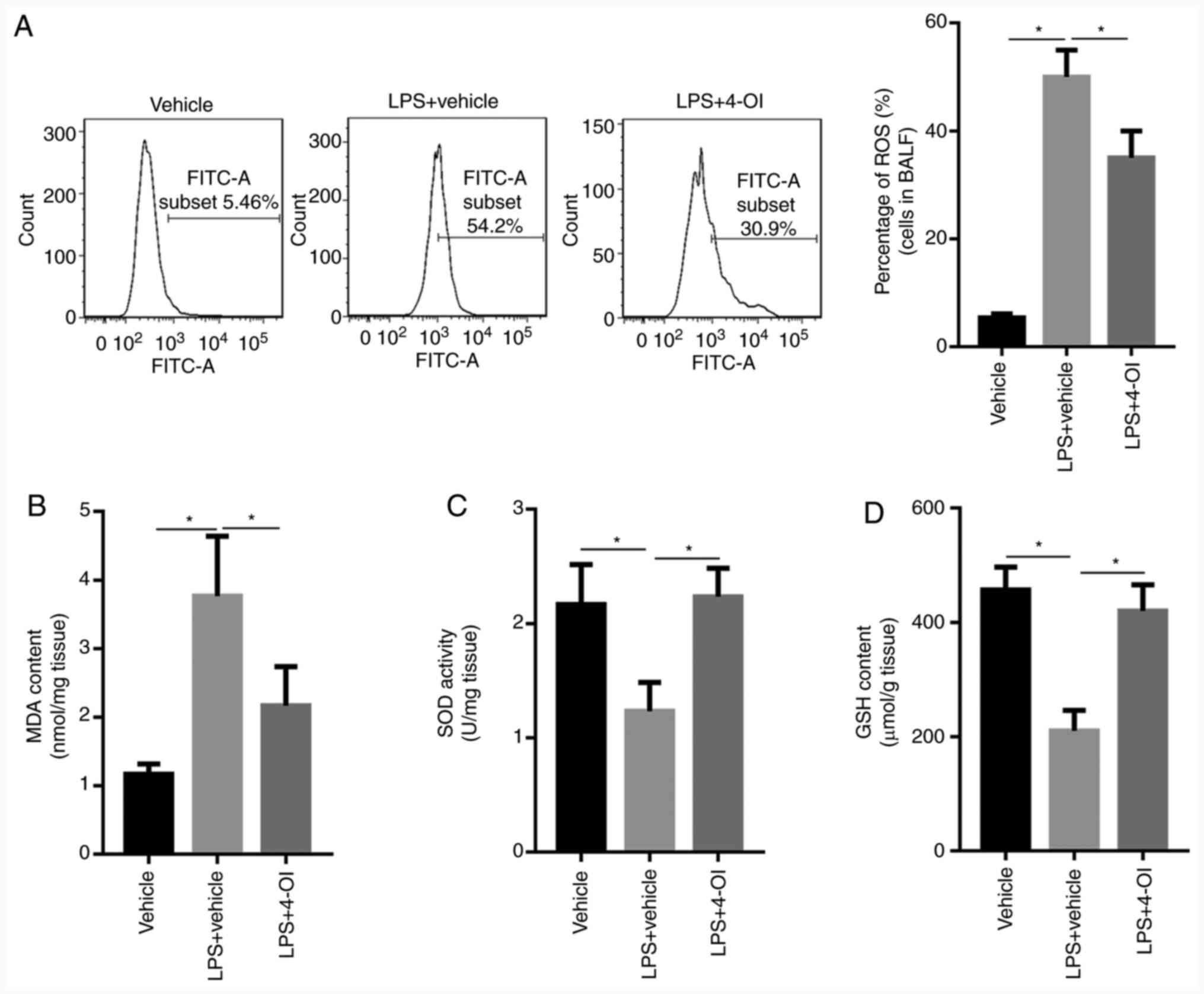 | Figure 34-OI inhibited oxidative stress in
LPS-induced acute lung injury mice. Treatment of mice with 4-OI at
doses of 25 mg/kg was administered intraperitoneally, and the
control group received an equivalent volume of vehicle
[(2-hydroxypropyl)-β-cyclodextrin in phosphate-buffered saline
(PBS)] 2 h before saline or LPS injection (5 mg/kg, intratracheal).
In total, 12 h later, ROS generation (A) in the BALF, MDA content
(B), SOD activity (C) and GSH content (D) in lung tissue were
determined (n=5). Data are expressed as the mean ± SEM.
*P<0.05. 4-OI, 4-octyl itaconate; LPS,
lipopolysaccharides; ROS, reactive oxygen species; MDA,
malondialdehyde; SOD, superoxide dismutase; GSH, glutathione. |
4-OI decreases the production of
inflammatory cytokines by inhibiting ROS-mediated PI3K/Akt/NF-κB
activation
Growing evidence has demonstrated that ROS can
activate the PI3K/Akt pathway, which further induces NF-κB
translocation from the cytosol into the nucleus (21). Therefore, the phosphorylated levels
of PI3K and Akt were detected by western blotting. The results
illustrated that LPS increased the phosphorylated levels of PI3K
and Akt, which were inhibited by the ROS scavenger NAC and 4-OI
(Fig. 4A). To further demonstrate
whether PI3K/Akt is involved in the activation of the NF-κB pathway
in LPS-stimulated macrophages, the expression of NF-κB p65 in the
nuclear and cytosolic fractions was detected by western blotting.
The results illustrated that LY294002, a specific inhibitor of the
PI3K/Akt pathway, significantly inhibited the LPS-induced increased
level of NF-κB p65 in the nuclear fraction (Fig. 4B). Moreover, ROS generation and MDA
formation were increased in LPS-treated RAW264.7 macrophage cells
and were inhibited by 4-OI and NAC treatment (Fig. 4C and D). In addition, the levels of SOD and GSH
were decreased in LPS-treated RAW264.7 macrophage cells, and this
effect was significantly reversed by pretreatment with 4-OI and NAC
(Fig. 4E and F). Furthermore, the production of TNF-α,
IL-1β and IL-6 was also decreased by LY294002 (Fig. 4G-L). Collectively, these data
demonstrate that 4-OI alleviated LPS-induced ALI by suppressing the
ROS-mediated PI3K/Akt/NF-κB pathway.
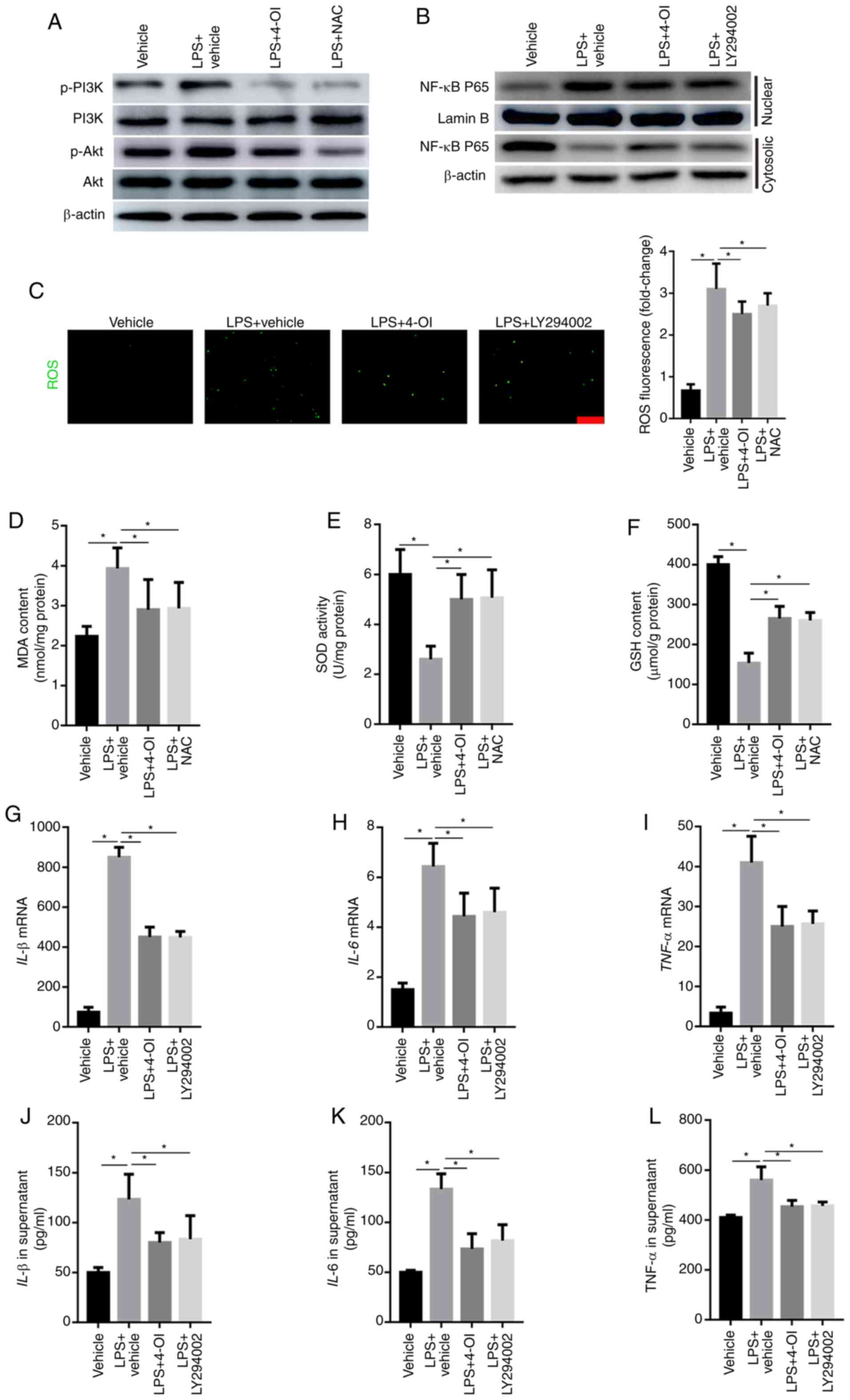 | Figure 44-OI decreased the induction of
inflammatory cytokines by inhibiting ROS-mediated PI3K/Akt/NF-κB
activation in LPS-treated macrophages. RAW264.7 macrophage cells
were pretreated with 4-OI (125 µM), NAC (10 mM), the PI3K inhibitor
LY294002 (25 µM) or vehicle control for 1 h and then stimulated
with LPS (1 µg/ml) for 30 min to detect the phosphorylated levels
of PI3K and Akt and the expression of NF-κB p65 in the nuclear and
cytosolic fractions in LPS-treated macrophages and for 24 h to
quantify ROS production, MDA content, SOD activity and GSH content
and the concentration of TNF-α, IL-1β, and IL-6 in macrophages. The
phosphorylated levels of PI3K and Akt (A) and the expression of
NF-κB p65 in the nuclear and cytosolic fractions (B) of LPS-treated
macrophages were detected by western blotting. ROS generation
(bar=100 µm) (C), MDA content (D), SOD activity (E) and GSH content
(F) in RAW264.7 macrophage cells were determined. The expression of
IL-1β (G), IL-6 (H) and TNF-α (I) in macrophages was measured by
qPCR. IL-1β (J), IL-6 (K) and TNF-α (L) protein contents in the
supernatant of RAW264.7 macrophage cells were determined. Data
represent the means ± SEM of three independent experiments.
*P<0.05. 4-OI, 4-octyl itaconate; LPS,
lipopolysaccharides; NAC, N-acetyl-L-cysteine; ROS, reactive oxygen
species; MDA, malondialdehyde; SOD, superoxide dismutase; GSH,
glutathione; IL, interleukin; TNF, tumor necrosis factor. |
Discussion
Uncontrolled inflammatory responses and/or excessive
oxidative stress are deemed to play an important role in the
pathogenesis of ALI (25). The
present study demonstrated that 4-OI alleviated LPS-induced ALI by
inhibiting the inflammatory response and oxidative stress. 4-OI
suppressed inflammatory cell infiltration and proinflammatory
cytokine generation, decreased lung tissue structural damage,
attenuated ROS generation and inhibited NF-κB activation in
LPS-induced ALI mice in vivo. These results illustrate that
4-OI may be selected as an effective drug for the treatment of ALI
in the future.
It has been reported that 4-OI attenuates hepatic
I/R injury in vivo and protects hepatocytes from injury
resulting from H/R in vitro (18). In addition, octyl itaconate (OI)
markedly prolonged survival, improved body temperature regulation,
decreased the clinical score, and reduced TNF and IL-1β levels in
an LPS model of sepsis by activating the anti-inflammatory
transcription factor Nrf2 (also known as NFE2L2) (15). Itaconate alkylates cysteine residues
of KEAP1 protein, a central player in the antioxidant response,
enabling Nrf2 to accumulate, migrate to the nucleus and further
increase the expression of downstream genes with anti-inflammatory
and antioxidant capacities (26).
Moreover, OI ameliorated renal fibrosis by inhibiting NF-κB
activation, decreasing the generation of ROS and suppressing
autophagy (27). Whether 4-OI can
protect against ALI remains unknown. The present study aimed to
explore the protective effects of 4-OI against ALI in mice, and it
was demonstrated that 4-OI could attenuate the inflammatory cell
infiltration, NF-κB activation and oxidative stress induced by LPS.
However, the exact mechanism needs to be further investigated. LPS
binds to TLR4, which initiates two classic pathways. One pathway is
toll/IL-1 receptor domain-containing adaptor including
interferon-β-dependent, requiring the toll/IL-1 receptor (TIR)
domain-containing adaptor protein. The other pathway is
MyD88-dependent and promotes the translocation of NF-κB from the
cytosol into the nucleus, leading to the release of proinflammatory
cytokines (28). The excessive
release of proinflammatory cytokines by activated macrophages
aggravates tissue injury (29). In
the present study, it was observed that treatment with 4-OI
significantly decreased the LPS-induced expression of IL-1β, IL-6
and TNF-α in lung tissue.
Oxidative stress modulated by ROS plays an essential
role in the progression of ALI (30). Under physiological conditions, ROS
can help prevent pathological injury or noxious stimulation.
However, excessive generation of ROS is thought to lead to cellular
injury and oxidative stress (9). In
the present study, it was found that treatment with 4-OI
significantly decreased LPS-induced ROS generation and MDA
formation and reversed the LPS-induced decrease in SOD and GSH
levels in lung tissue.
The PI3K/Akt pathway plays an important role in
cellular defense against inflammatory stimuli (31). Previous studies have also indicated
that inhibition of the PI3K/Akt pathway attenuates LPS-induced ALI
(32). Moreover, ROS lead to the
activation of PI3K/Akt by inactivating the PTEN protein.
Furthermore, PTEN inhibits the activation of NF-κB via the PI3K/Akt
pathway (33). In addition, ROS, as
secondary messengers, induce the nuclear translocation of NF-κB and
the production of inflammatory cytokines (34). The present study demonstrated that
4-OI and NAC decreased the LPS-induced production of ROS, and the
role of 4-OI in the PI3K/Akt pathway was further explored. In
accordance with a previous study (21), the administration of NAC also
suppressed the phosphorylated levels of PI3K and Akt in LPS-treated
RAW264.7 macrophages. It has been reported that the PI3K and Akt
pathways regulate LPS-induced NF-κB activation (35). Therefore, it was hypothesized that
the PI3K/Akt pathway plays a pivotal role in ROS-mediated NF-κB
activation. The results demonstrated that LY294002, a specific
inhibitor of the PI3K/Akt pathway, markedly inhibited LPS-induced
NF-κB activation. Moreover, LY294002 also decreased the levels of
TNF-α, IL-1β and IL-6. These results demonstrated that 4-OI
decreased the production of inflammatory cytokines by inhibiting
the ROS-mediated PI3K/Akt/NF-κB pathway.
In conclusion, the present study confirmed that 4-OI
exerts potential protective effects against LPS-induced lung injury
in mice, possibly through inhibition of the ROS-mediated
PI3K/Akt/NF-κB pathway.
Acknowledgements
Not applicable.
Funding
This work was supported by the Natural Science
Foundation of Ningxia (grant no. NZ16137).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
SL and YX designed the study. YX and LZ performed
all the experiments and analyzed the data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The experiments were approved by the Institutional
Ethics Committee of Faculty of Ningxia Medical University
(Yinchuan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shen Y, Cai G, Chen S, Hu C and Yan J:
Fluid intake-related association between urine output and mortality
in acute respiratory distress syndrome. Respir Res.
21(24)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Ware LB and Matthay MA: The acute
respiratory distress syndrome. N Engl J Med. 342:1334–1349.
2000.PubMed/NCBI View Article : Google Scholar
|
|
3
|
el-Ebiary M, Torres A, Fàbregas N, de la
Bellacasa JP, González J, Ramirez J, del Baño D, Hernández C and
Jiménez de Anta MT: Significance of the isolation of Candida
species from respiratory samples in critically ill, non-neutropenic
patients. An immediate postmortem histologic study. Am J Respir
Crit Care Med. 156:583–590. 1997.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lv H, Liu Q, Wen Z, Feng H, Deng X and Ci
X: Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute
lung injury via induction of AMPK/GSK3β-Nrf2 signal axis. Redox
Biol. 12:311–324. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Kawai T and Akira S: Toll-like receptors
and their crosstalk with other innate receptors in infection and
immunity. Immunity. 34:637–650. 2011.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kong L and Ge BX: MyD88-independent
activation of a novel actin-Cdc42/Rac pathway is required for
Toll-like receptor-stimulated phagocytosis. Cell Res. 18:745–755.
2008.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Shim JH, Xiao C, Paschal AE, Bailey ST,
Rao P, Hayden MS, Lee KY, Bussey C, Steckel M, Tanaka N, et al:
TAK1, but not TAB1 or TAB2, plays an essential role in multiple
signaling pathways in vivo. Genes Dev. 19:2668–2681.
2005.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Badr G, Al-Sadoon MK, El-Toni AM and
Daghestani M: Walterinnesia aegyptia venom combined with silica
nanoparticles enhances the functioning of normal lymphocytes
through PI3K/AKT, NFκB and ERK signaling. Lipids Health Dis.
11(27)2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Yang CS, Kim JJ, Lee SJ, Hwang JH, Lee CH,
Lee MS and Jo EK: TLR3-triggered reactive oxygen species contribute
to inflammatory responses by activating signal transducer and
activator of transcription-1. J Immunol. 190:6368–6377.
2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Hong HY, Jeon WK and Kim BC: Up-regulation
of heme oxygenase-1 expression through the Rac1/NADPH
oxidase/ROS/p38 signaling cascade mediates the anti-inflammatory
effect of 15-deoxy-delta 12,14-prostaglandin J2 in murine
macrophages. FEBS Lett. 582:861–868. 2008.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Jiang K, Guo S, Yang C, Yang J, Chen Y,
Shaukat A, Zhao G, Wu H and Deng G: Barbaloin protects against
lipopolysaccharide (LPS)-induced acute lung injury by inhibiting
the ROS-mediated PI3K/AKT/NF-κB pathway. Int Immunopharmacol.
64:140–150. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yu XH, Zhang DW, Zheng XL and Tang CK:
Itaconate: An emerging determinant of inflammation in activated
macrophages. Immunol Cell Biol. 97:134–141. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Rittenhouse JW and McFadden BA: Inhibition
of isocitrate lyase from Pseudomonas indigofera by itaconate. Arch
Biochem Biophys. 163:79–86. 1974.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bambouskova M, Gorvel L, Lampropoulou V,
Sergushichev A, Loginicheva E, Johnson K, Korenfeld D, Mathyer ME,
Kim H, Huang LH, et al: Electrophilic properties of itaconate and
derivatives regulate the IκBζ-ATF3 inflammatory axis. Nature.
556:501–504. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Mills EL, Ryan DG, Prag HA, Dikovskaya D,
Menon D, Zaslona Z, Jedrychowski MP, Costa ASH, Higgins M, Hams E,
et al: Itaconate is an anti-inflammatory metabolite that activates
Nrf2 via alkylation of KEAP1. Nature. 556:113–117. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press (US), Washington, DC, 2011.
|
|
17
|
Zhang Y, Xu T, Pan Z, Ge X, Sun C, Lu C,
Chen H, Xiao Z, Zhang B, Dai Y, et al: Shikonin inhibits myeloid
differentiation protein 2 to prevent LPS-induced acute lung injury.
Br J Pharmacol. 175:840–854. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yi Z, Deng M, Scott MJ, Fu G, Loughran PA,
Lei Z, Li S, Sun P, Yang C, Li W, et al: IRG1/itaconate activates
Nrf2 in hepatocytes to protect against liver ischemia-reperfusion
injury. Hepatology: Jan 30, 2020 (Epub ahead of press).
|
|
19
|
Aziz M, Matsuda A, Yang WL, Jacob A and
Wang P: Milk fat globule-epidermal growth factor-factor 8
attenuates neutrophil infiltration in acute lung injury via
modulation of CXCR2. J Immunol. 189:393–402. 2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Qi S, Xin Y, Guo Y, Diao Y, Kou X, Luo L
and Yin Z: Ampelopsin reduces endotoxic inflammation via repressing
ROS-mediated activation of PI3K/Akt/NF-κB signaling pathways. Int
Immunopharmacol. 12:278–287. 2012.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Li X, Peng F, Xie C, Wu W, Han X and Chen
L:
(E)-3-(3,4-Dimethoxyphenyl)-1-(5-hydroxy-2,2-dimethyl-2H-chromen-6-yl)prop-2-en-1-one
ameliorates the collagen-arthritis via blocking ERK/JNK and NF-κB
signaling pathway. Int Immunopharmacol. 17:1125–1133.
2013.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Sun GY, Yang HH, Guan XX, Zhong WJ, Liu
YP, Du MY, Luo XQ, Zhou Y and Guan CX: Vasoactive intestinal
peptide overexpression mediated by lentivirus attenuates
lipopolysaccharide-induced acute lung injury in mice by inhibiting
inflammation. Mol Immunol. 97:8–15. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Huang XT, Li C, Peng XP, Guo J, Yue SJ,
Liu W, Zhao FY, Han JZ, Huang YH, Yang-Li , et al: An
excessive increase in glutamate contributes to glucose-toxicity in
β-cells via activation of pancreatic NMDA receptors in rodent
diabetes. Sci Rep. 7(44120)2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Huang XT, Liu W, Zhou Y, Sun M, Yang HH,
Zhang CY and Tang SY: Galectin-1 ameliorates
lipopolysaccharide-induced acute lung injury via AMPK-Nrf2 pathway
in mice. Free Radic Biol Med. 146:222–233. 2020.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Hayes JD and Dinkova-Kostova AT: The Nrf2
regulatory network provides an interface between redox and
intermediary metabolism. Trends Biochem Sci. 39:199–218.
2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tian F, Wang Z, He J, Zhang Z and Tan N:
4-Octyl itaconate protects against renal fibrosis via inhibiting
TGF-β/Smad pathway, autophagy and reducing generation of reactive
oxygen species. Eur J Pharmacol. 873(172989)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Biswas SK and Lopez-Collazo E: Endotoxin
tolerance: New mechanisms, molecules and clinical significance.
Trends Immunol. 30:475–487. 2009.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Hussain S, Johnson CG, Sciurba J, Meng X,
Stober VP, Liu C, Cyphert-Daly JM, Bulek K, Qian W, Solis A, et al:
TLR5 participates in the TLR4 receptor complex and promotes
MyD88-dependent signaling in environmental lung injury. Elife: Jan
28, 2020 (Epub ahead of print).
|
|
30
|
Fisher AB, Dodia C, Chatterjee S and
Feinstein SI: A peptide inhibitor of NADPH oxidase (NOX2)
activation markedly decreases mouse lung injury and mortality
following administration of lipopolysaccharide (LPS). Int J Mol
Sci. 20(2395)2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Hyam SR, Lee IA, Gu W, Kim KA, Jeong JJ,
Jang SE, Han MJ and Kim DH: Arctigenin ameliorates inflammation in
vitro and in vivo by inhibiting the PI3K/AKT pathway and polarizing
M1 macrophages to M2-like macrophages. Eur J Pharmacol. 708:21–29.
2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhao M, Li C, Shen F, Wang M, Jia N and
Wang C: Naringenin ameliorates LPS-induced acute lung injury
through its anti-oxidative and anti-inflammatory activity and by
inhibition of the PI3K/AKT pathway. Exp Ther Med. 14:2228–2234.
2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhao M, Zhou A, Xu L and Zhang X: The role
of TLR4-mediated PTEN/PI3K/AKT/NF-κB signaling pathway in
neuroinflammation in hippocampal neurons. Neuroscience. 269:93–101.
2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Niu T, Tian Y, Wang G, Guo G, Tong Y and
Shi Y: Inhibition of ROS-NF-κB-dependent autophagy enhances
Hypocrellin A united LED red light-induced apoptosis in squamous
carcinoma A431 cells. Cell Signal. 69(109550)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhong W, Qian K, Xiong J, Ma K, Wang A and
Zou Y: Curcumin alleviates lipopolysaccharide induced sepsis and
liver failure by suppression of oxidative stress-related
inflammation via PI3K/AKT and NF-κB related signaling. Biomed
Pharmacother. 83:302–313. 2016.PubMed/NCBI View Article : Google Scholar
|














