1. Introduction
According to the World Health Organization report,
gastric cancer is the 6th most fatal cancer type worldwide, with
783,000 deaths in 2018(1). To date,
the early diagnosis rate of gastric cancer has remained low and the
golden standard for its diagnosis is endoscopic pathological
biopsy. Gastric cancer is mainly treated through surgery,
chemoradiotherapy, immunotherapy and molecular targeted therapies
(2). As a novel treatment method,
immunotherapy includes vaccination, immune checkpoint inhibitors
and adoptive T-cell therapy, but numerous treatments are still in
the research phase (3). With the
development of medical theory and technology, the 5-year survival
rate of patients with early gastric cancer has reached as high as
95% (4). Regrettably, treatment
methods for advanced gastric cancer are still limited and the
5-year survival rate of such patients following neoadjuvant
chemoradiotherapy combined with immunotherapy and targeted therapy
remains low. At present, the overall treatment principle is to
prepare more reasonable and comprehensive therapeutic regimens
through a full assessment of the patients' condition to improve
their quality of life (5).
The nuclear factor-κB (NF-κB) family is a class of
transcription factors that possess multiple biological functions.
There are 5 important genes in the NF-κB signaling pathway,
including NF-κB1 (p50/p105), NF-κB2 (p52/p100), RelA (p65), RelB
and c-Rel (6). The combination of
IκBα (a prototypical member of the IκB family) and NF-κB complexes,
such as NF-κB1 p50-RelA and NF-κB1 p50-c-Rel dimers, can inhibit
the nuclear translocation of NF-κB complexes. The canonical pathway
is activated by immune-related receptors (7). Subsequently, the IKK complex forms and
mediates IκBα phosphorylation and ubiquitin (Ub)-dependent
proteasomal degradation, resulting in nuclear translocation of the
NF-κB dimers. The noncanonical NF-κB pathway is a NF-κB-inducing
kinase, which activates downstream kinase IKKα to process p100 to
p52, thereby forming the RelB/p52 complexes to activate the NF-κB
signaling pathway (7). It has been
indicated that the NF-κB signaling pathway regulates the cell
cycle, apoptosis, migration, adhesion, inflammatory response and
immune response (8). A large number
of studies have demonstrated that the NF-κB signaling pathway is
abnormally activated in different types of malignancies and that it
has important roles in promoting tumor growth, invasion and
migration, as well as in inhibiting apoptosis and enhancing
chemoradiotherapy sensitivity (6).
Furthermore, the NF-κB signaling pathway may serve as a key
effector pathway that maintains the characteristics of cancer stem
cells (6,9). Epithelial-mesenchymal transition (EMT)
has been considered to be closely related to the migration and
invasion of malignant cells. During the transdifferentiation of
epithelial cells into mesenchymal cells, cell-cell adhesions are
weakened, the cell adhesion molecule, E-cadherin, is downregulated
and the mesenchymal molecule vimentin is up-regulated, leading to
increased cell migration and invasion. It has been indicated that
the high expression of cyclooxygenase-2 may lower the expression of
E-cadherin through the NF-κB/Snail pathway, thereby enhancing the
invasion and migration of tumor cells (10). The NF-κB pathway is continuously
activated in the cytoplasm and nucleus of gastric cancer cells, and
its activation is significantly higher than that in para-carcinoma
normal tissues (11). The
activation degree of the NF-κB pathway has significant correlations
with the size, degree of malignancy, depth of infiltration,
lymphatic and peritoneal metastasis and prognosis of gastric cancer
patients. As confirmed in numerous clinicopathological specimens,
the RelA (p65) protein has an increased expression in gastric
adenocarcinoma tissues, where it is involved in tumor growth,
invasion and metastasis (12,13).
Studies have proved that microRNAs (miRNAs/miRs) are
able to act as oncogenes or tumor suppressor genes in tumors
(14-16).
It was also determined that non-coding (nc)RNAs serum profiles are
different between gastric cancer patients and healthy individuals.
miRNAs were indicated to have stable and simple detection indexes,
which highlight their importance as biomarkers for early diagnosis
of gastric cancer (17). miR-331,
miR-21, miR-20b, miR-125a, miR-137, miR-141, miR-146a, miR-196a,
miR-206, miR-218, miR-486-5p and miR-506, have been confirmed as
tumor markers with potential value in gastric cancer diagnosis and
prognostic prediction (18).
Increasing evidence suggests that the imbalance of miRNAs has
important links with tumor proliferation, migration, invasion,
apoptosis, drug resistance and angiogenesis. miR-21, miR-23a,
miR-27a, miR-106b-25, miR-130b, miR-199a, miR-215, miR-222-221 and
miR-370 promote gastric cancer occurrence and development, while
miR-29a, miR-101, miR-125a, miR-129, miR-148b, miR-181c, miR-212,
miR-218, miR-335, miR-375, miR-449, miR-486 and miR-512 suppress
gastric cancer progression (18).
In recent years, the potential value of long ncRNAs
(lncRNAs) as tumor markers has been confirmed (18), and the important regulatory roles of
lncRNAs in tumor occurrence and development have been recognized.
lncRNAs are involved in various malignant biological behaviors;
therefore, lncRNAs are expected to be used in tumor treatments
(19-22).
In addition, chemical modification of lncRNAs may be associated
with cancer development (23).
Non-coding RNA serum profiles are different between
patients with gastric cancer and healthy individuals, which have
significant effects on prognosis. Additionally, the close
association of the NF-κB signaling pathway with gastric cancer
occurrence, development and chemoradiotherapeutic sensitivity may
imply that miRNAs and lncRNAs share certain cascades with the NF-κB
signaling pathway. In the present review, the cascades of miRNAs
and lncRNAs with the NF-κB signaling pathway in gastric cancer were
comprehensively explored.
2. miRNAs and the NF-κB signaling
pathway
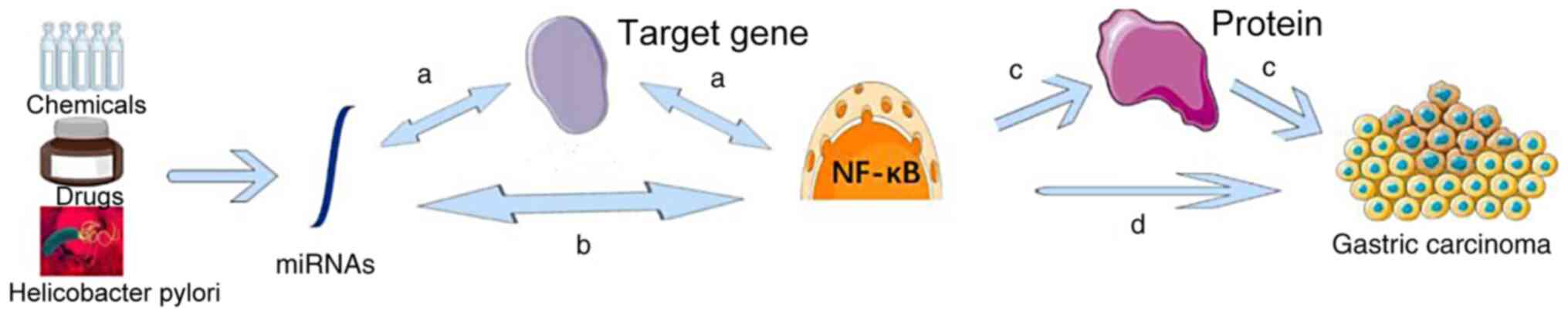
The general regulatory mechanisms of miRNAs and the
NF-κB signaling pathway in gastric cancer are presented in Fig. 1 (24). miRNAs can regulate target genes to
activate or inhibit the NF-κB signaling pathway. The NF-κB
signaling pathway is able to influence miRNA expression by
regulating target genes. miRNAs can also activate or inhibit the
NF-κB signaling pathway directly, with the NF-κB signaling pathway
influencing miRNA expression directly. NF-κB or miRNAs can
therefore influence the development of gastric cancer by regulating
downstream proteins or affecting the development of gastric cancer
directly (24,25).
miR-20a activation of the NF-κB
signaling pathway
The imbalance of miRNAs is frequently considered an
important cause of tumor drug resistance and the continuous
activation of the NF-κB signaling pathway also has an important
role in the decline of tumor chemoradiotherapeutic sensitivity
(26). It was previously indicated
that miR-20a expression is significantly increased in a variety of
gastric cancer cell lines and patient tissues, and that its ectopic
expression facilitates the development of gastric cancer by
promoting the proliferation, migration and invasion of gastric
cancer cells (27). miR-20a has
also been reported to be upregulated in the generated cisplatin
(DDP)-resistant gastric cancer cell line SGC7901/DDP.
Overexpression of miR-20a leads to inhibition of the expression of
the cylindromatosis (CYLD) gene, activates the NF-κB signaling
pathway, increases the expression of p65, livin and survivin,
reduces apoptosis of gastric cancer cells and enhances their
chemoresistance (28).
miR-300 activation of the NF-κB
signaling pathway
Shikimic acid (SA), also known as Chinese anise, is
a type of hydroaromatic compound that is present in bracken ferns
and that increases the risk of gastric and esophageal cancers.
Through in vitro experiments, Ma and Ning (29) confirmed that SA was able to
upregulate miR-300 expression in the estrogen receptor-positive
breast cancer cell line MCF-7, and inhibit IκBα protein, thereby
upregulating the NF-κB signaling pathway and enhancing breast
cancer cell proliferation. It was previously indicated that miR-300
is able to suppress EMT in malignant tumors, improve the prognosis
of patients with laryngeal squamous cell carcinoma and inhibit the
metastasis of oral squamous cell carcinoma (30-32).
The ability of SA to increase the risk of gastric cancer may be
related to such a mechanism, but relevant research is still
limited.
miR-224 activation of the NF-κB
signaling pathway
Hypoxia is an important factor in the occurrence and
development of a variety of cancer types. Hypoxia-inducible
factor-1 (HIF-1) is able to regulate various miRNAs, while certain
miRNAs are also able to target HIF-1. Currently, miR-224 is
considered as an oncogene that is highly expressed in various tumor
types and that promotes the proliferation and invasion of tumor
cells (33,34). The interactions between HIF-1 and
miRNAs may have important roles in angiogenesis, apoptosis, cell
cycle regulation, proliferation, migration and tumor drug
resistance (35). He et al
(36) confirmed that miR-224 is
highly expressed in gastric cancer tissues and metastatic lymph
nodes. Furthermore, it was confirmed that hypoxia and HIF-1α are
able to upregulate the expression of miR-224 in gastric cancer
cells, while miR-224 downregulated the expression of Ras
association (RalGDS/AF-6) domain family 8 (RASSF8) through target
genes. RASSF8 is closely related to the NF-κB signaling pathway, as
both NF-κB transcriptional activity and p65 translocation were
enhanced following RASSF8 knockout, which promoted the
proliferation, migration and invasion of gastric cancer cells
(36).
miR-21 activation of the NF-κB
signaling pathway
miR-21 expression is upregulated in most tumor
tissues, where it has an important function in tumor cell adhesion,
migration, invasion and angiogenesis. The expression of miR-21 also
correlates with patient prognosis, histological type, lymph node
metastasis and TNM stage. miR-21 is also able to regulate the
radiosensitivity of gastric cancer cells and directly downregulate
the expression of phosphatase and tensin homolog deleted on
chromosome ten (PTEN) (37).
Smoking is a risk factor for a variety of cancers and nicotine is
able to regulate cell proliferation, apoptosis, migration and
invasion through related signaling pathways, which promote tumor
occurrence and development. Shin et al (38) proved that nicotine is able to
activate the NF-κB signaling pathway through the EP2/4 receptor and
promote the expression of miR-21 and miR-16, therefore facilitating
the proliferation of gastric cancer cells. Of note, a study
indicated that miR-21 may be downregulated by celastrol, a natural
component from a plant, resulting in the inhibition of the
activation of the PI3K/Akt/NF-κB signaling pathway and the
promotion of gastric cancer cells apoptosis (39). According to related research,
chromobox protein homolog 7 (CBX7) upregulates the Akt/NF-κB/miR-21
signaling pathway via the inhibition of p16, which positively
regulates the characteristics of gastric cancer stem cells. In
addition, it has been proved that the Akt/NF-κB/miR-21 signaling
pathway is a key effector pathway for CBX7 in inducing the
characteristics of gastric cancer stem cells (40). Furthermore, the NF-κB signaling
pathway and miR-21 appear to activate each other through different
pathways, resulting in a joint promotion of gastric cancer
progression.
miR-17-92 activation of the NF-κB
signaling pathway
It is thought that miR-17-92, also known as
oncomiR-1, is an important carcinogen (41). Overexpression of miR-17-92 promoted
the proliferation and reduced apoptosis of gastric cancer cells in
nude mice, and accelerated tumor growth in vivo (42). Tumor necrosis factor
receptor-associated factor 3 (TRAF3) is thought to be related to
patients' prognosis (43). TRAF3
expression levels in gastric cancer tissues are far lower than
those in para-carcinoma normal tissues and its silencing may
promote proliferation and markedly enhance migration and invasion
of gastric cancer cells. As previously reported, TRAF3 is the
target gene of miR-17-92 and its levels may be decreased by
miR-17-92 overexpression, resulting in the activation of the NF-κB
signaling pathway through non-classical and classical pathways,
which promoted the proliferation, migration and invasion, and
inhibited apoptosis of gastric cancer cells (42).
miR-577 activation of the NF-κB
signaling pathway
miR-577 expression is closely associated with
metastasis and poor prognosis of patients with gastric cancer. It
has been confirmed that gastric cancer cells overexpressing miR-577
are spindle-shaped and display a strong invasive ability. miR-577
does not affect the proliferation of gastric cancer cells, but may
promote the migration, invasion and chemoresistance of gastric
cancer cells. Serum deprivation response protein (SDPR) is a tumor
suppressor gene, and its expression in gastric cancer tissues is
lower than that in normal tissues, which may be related to gastric
cancer prognosis. miR-577 is able to inhibit the expression of its
target gene SDPR and boost the activation of the ERK-NF-κB
signaling pathway through forming an SDPR-ERK protein complex,
which induces EMT. Furthermore, a positive feedback loop is also
formed to upregulate miR-577 expression (44).
miR-216a-3p activation of the NF-κB
signaling pathway
Runt-related transcription factor-1 (RUNX1) has been
identified as a cancer suppressor gene and its expression is
downregulated in gastric cancer tissues (45). miR-216a-3p is highly expressed in
the gastric cancer cell lines AGS, MKN-45 and HGC-27. Analysis of
140 cases of gastric cancer tissue specimens suggested that
miR-216a-3p expression levels are associated with the degree of
malignancy and prognosis of gastric cancer. Furthermore, high
expression of miR-216a-3p was indicated to activate the NF-κB
signaling pathway and increase the expression of its downstream
proteins, MMP2, MMP9, cyclin D1 and Bcl-2, through inhibition of
RUNX1 expression, which enhanced the proliferation, migration and
invasion of gastric cancer cells (46).
miR-362 activation of the NF-κB
signaling pathway
According to the public research database about
cancer-related miRNA microarrays, miR-362 expression is increased
in gastric cancer cells. CYLD is a cancer suppressor gene that
inhibits proliferation and induces apoptosis of tumor cells
(47). A study suggested that
miR-362, through inhibiting the expression of its target gene CYLD,
facilitates the activation of the NF-κB signaling pathway, promotes
the proliferation and inhibits apoptosis of gastric cancer cells,
and induces cisplatin resistance (48).
miR-10b activation of the NF-κB
signaling pathway
miR-10b expression correlates with the size,
invasiveness, lymph node metastasis, distant metastasis and
prognosis of gastric cancer (49).
CUB and Sushi multiple domains-1 (CSMD1) expression is
downregulated in gastric cancer tissues and associated with the
prognosis of patients with gastric cancer. miR-10b is able to
inhibit the expression of part of CSMD1, which further activates
the NF-κB signaling pathway and promotes the EMT of gastric cancer
cells, leading to enhanced migration and invasion of gastric cancer
cells. In nude mouse experiments, the growth and metastasis of
gastric cancer was promoted by overexpression of miR-10b (50).
miR-223-3p activation of miRNAs by the
NF-κB signaling pathway
Through the secretion of cytotoxin-associated gene A
(CagA), CagA-positive Helicobacter pylori is able to
stimulate the activation of the NF-κB signaling pathway in gastric
cancer cells and upregulate miR-223-3p expression, leading to
partial inhibition of the expression of AT-rich interacting domain
containing protein 1A (ARID1A). It has been previously confirmed
that ARID1A is a tumor suppressor gene that restrains the invasion
and migration of tumor cells via upregulating the levels of
E-cadherin and p21. In patients infected with CagA-positive
Helicobacter pylori, the occurrence and development of
gastric cancer may be promoted through the NF-κB/miR-223-3p/ARID1A
axis and miR-223-3p may offer a feasible method for preventing and
treating gastric cancer (51).
miR-135b-5p activation of miRNAs by
the NF-κB signaling pathway
It has been indicated that inflammation and
infection account for ~25% of carcinogenic factors. Long-term
chronic inflammation-induced cell DNA damage and epigenetic changes
may be present in inflammation-related cancers, such as
Helicobacter pylori-infected gastric cancer (52). In tissues of trefoil factor
1-knockout mice and the gastric cancer MKN45 and SNU1 cell lines,
Helicobacter pylori was indicated to promote the expression
of miR-135b-5p through the activation of the NF-κB signaling
pathway and downregulation of Kruppel-like factor 4 (KLF4), which
resulted in the inhibition of apoptosis and enhancement of drug
resistance of gastric cancer cells. It was confirmed that KLF4 acts
as a potential cancer suppressor gene in gastric cancer and that it
has a role in cisplatin resistance (53).
miR-425 activation of miRNAs by the
NF-κB signaling pathway
As a malignant tumor suppressor in vivo, PTEN
is mutated or lost in a variety of cancer types. IL-1β is an
inflammatory factor that activates the NF-κB signaling pathway in
gastric cancer cells, which reduces the expression of target gene
PTEN through upregulating miRNA-425 and promotes the growth of
gastric cancer cells (54).
According to related studies, both miR-21 and miR-32 downregulate
the expression of PTEN in gastrointestinal tumors, which
facilitates the migration and invasion of tumor cells (55,56).
miR-107 activation of miRNAs by the
NF-κB signaling pathway
As suggested in previous related studies, miR-107 is
a cancer suppressor gene, which inhibits tumor cell growth in colon
and pancreatic cancers, as well as head-neck tumors (57). However, it was reported that miR-107
expression is upregulated in gastric cancer tissues (58). Overactivation of the NF-κB pathway
may promote the expression of miR-107, while miR-107 is able to
downregulate the downstream target gene forkhead box protein O1
(FOXO1), thereby enhancing the proliferation of gastric cancer
cells. The expression of FOXO1 may be regulated by a variety of
miRNAs as dysregulated miR-223, miR-139 and miR-370 expression may
exert a series of influences on the progression of gastric cancer
via regulating FOXO1(59).
miR-210 activation of miRNAs by the
NF-κB signaling pathway
The uncontrolled expression of phosphatase of
regenerating liver-3 (PRL-3) is closely related to the overall
survival rate and tumor progression. It is believed that miR-210 is
able to regulate the expression of target genes, E2F transcription
factor 3, homeobox A1 (HOXA1) and HOXA3, which are crucial
molecules that regulate cell proliferation and cell cycle
progression. It has been indicated that the transcriptional level
of PRL-3 in gastric cancer tissues positively correlates with the
level of miR-210 expression. Through gene enrichment analysis and
in vitro experiments, Zhang et al (60) confirmed that PRL-3 is able to
increase HIF-1α levels through the activation of the NF-κB
signaling pathway and the upregulation of miR-210 in a
HIF-1α-dependent manner, which enhance the invasion and migration
of gastric cancer.
To sum up, the above-mentioned studies appear to
confirm that miRNAs act as oncogenes that may be involved in the
activation of the NF-κB signaling pathway, which in turn, is able
to upregulate the expression of related miRNAs. The interaction
between the two jointly promotes the occurrence and development of
gastric cancer. However, related mechanisms are yet to be revealed
and whether inhibiting the NF-κB signaling pathway or the
expression of related miRNAs is a novel target in gastric cancer
treatment still requires to be experimentally validated.
miR-338 inhibition of the NF-κB
signaling pathway
Li et al (61) screened differentially expressed
genes from 90 samples in the Gene Expression Omnibus database and
classified gastric cancer into 4 subtypes based on different target
miRNAs. The most important feature of the second subtype was the
miR-338/C-C motif chemokine ligand (CCL)21/NF-κB signaling pathway.
miR-198/protein inhibitor of activated STAT4/NF-κB and
miR-370/CCL21/NF-κB also belonged to this subtype. It was indicated
that miR-338 is able to inhibit the proliferation and migration of
gastric cancer cells (61).
miR-7 inhibition of the NF-κB
signaling pathway
As reported in a previous study, miR-7 is a tumor
suppressor gene that suppresses the proliferation, survival,
migration and invasion of a variety of cancer cell types (62). Using quantitative isobaric Tags for
Relative and Absolute Quantitation, gene expression microarray
analysis and bioinformatics, it was revealed that FOS and RelA are
important functional targets of miR-7 in gastric cancer cells.
Analysis of 106 gastric cancer tissue samples indicated that
gastric cancer tissues with low miR-7 expression had a more
malignant phenotype and that miR-7 expression negatively correlates
with RelA and FOS expression. Gastric cancer cell lines
overexpressing miR-7 have a weakened proliferation ability, a
significantly increased early apoptotic rate and an enhanced
sensitivity to chemotherapeutic drugs (63). In nude mouse experiments,
tumorigenesis is markedly reduced following miR-7 overexpression.
It was also suggested that the expression of RelA, FOS,
proliferating cell nuclear antigen and cyclin D1 markedly declined
in tumor tissues. Furthermore, miR-7 overexpression was determined
to inhibit IKKε expression, which inhibited the activation of RelA,
leading to the suppression of the activation of the NF-κB signaling
pathway (63). The results revealed
that Helicobacter pylori infection is able to induce the
expression of IKKε and increase the expression of RelA in normal
gastric mucosa cells, thereby inhibiting the expression of miR-7.
Through such a mechanism, the NF-κB signaling pathway may be
overactivated in patients with Helicobacter pylori
infection, suggesting that persistent chronic inflammation is
caused and ultimately transforms cells into malignant tumor cells
(63). Furthermore, dysregulated
expression of miR-7 was observed in a variety of tumor tissues
(64). Ye et al (65) used the Cancer Genome Atlas Stomach
Adenocarcinoma and National Center for Bioinformatics Gene
Expression Omnibus databases (accession no.
10.1186/s13046-019-1074-6), and determined that there was a
negative correlation between miR-7 expression and NF-κB RelA (p65)
protein expression in primary gastric cancer tissues. Downregulated
miR-7 corresponded to poor prognosis (66), and mature miR-7 was significantly
reduced in the gastric cancer HGC-27 and MKN-28 cell lines. In the
nude mouse model, hepatic and pulmonary metastasis of gastric
cancer was inhibited in the miR-7 overexpression group of mice. It
was argued that overexpression of miR-7 may not only inhibit the
expression of its downstream protein targets, intercellular
adhesion molecule-1, MMP2, MMP9, VEGF and vascular cell adhesion
molecule-1 through restraining the NF-κB signaling pathway, but
also through reducing inflammatory cell infiltration. As mentioned
above, inflammation has a close link with the occurrence and
development of tumors; therefore, miR-7 may provide a feasible
method for the prevention and treatment of gastric cancer (65).
miR-218 inhibition of the NF-κB
signaling pathway
In patients with metastatic gastric cancer, the
transcription factor POU class 2 homeobox 2 (POU2F2) was indicated
to be abnormally upregulated, while the expression of miR-218 was
downregulated. miR-218 was also demonstrated to inhibit the
migration and invasion of tumor cells. Researchers confirmed that
POU2F2 overexpression may activate the slit guidance ligand
(SLIT)2/Roundabout guidance receptor 1 (ROBO1) signaling pathway
and promote EMT of gastric cancer cells, thereby facilitating
invasion and migration of gastric cancer cells, while its
expression was able to be further enhanced by overactivation of
NF-κB signaling pathway. According to a bioinformatics analysis,
miR-218 is able to directly act on the target gene POU2F2, inhibit
the NF-κB signaling pathway and reduce the expression of POU2F2
through IKK-β suppression. ROBO1 is a single-channel transmembrane
receptor that is able to boost tumor metastasis. Of note, SLIT3 was
able to partially reduce miR-218 expression in metastatic gastric
cancer, thereby relieving the inhibitory effect of miR-218 on
ROBO1. Overall, miR-218 may be an important target that may offer a
novel direction in the diagnosis and treatment of gastric cancer
(67).
miR-185 inhibition of the NF-κB
signaling pathway
The expression of gastrokine 1 (GKN1) and GKN2 in
gastric cancer tissues and the AGS gastric cancer cell line was
lower than that in para-carcinoma tissues and normal gastric
epithelial cell (68,69). GKN2 maintains the homeostasis of
gastric mucosal epithelium and may delay the development of gastric
cancer (70). It was previously
indicated that GKN1 promotes apoptosis of gastric cancer cells and
suppresses their proliferation by causing cell cycle arrest at the
G2/M phase. GKN1 may promote miR-185 expression and decrease the
expression of C-myc, a specific protein involved in maintaining
gastric mucosal homeostasis, GKN2 is able to completely reverse the
function of GKN1 in gastric cancer AGS cell line and that GKN1
inhibits the activity of the NF-κB signaling pathway via
facilitating the expressions of IκB and GKN2(71).
miR-146a inhibition of the NF-κB
signaling pathway
miR-146a, a dependent gene of the NF-κB signaling
pathway, is able to alleviate the inflammatory response. Crone
et al (72) indicated that
there was no expression of miR-146a in normal gastric mucosa.
Another study confirmed that as a cancer suppressor gene, miR-146a
is able to inhibit the expression of transforming growth
factor-activated kinase-1, resulting in the increase of IκBα
expression and the suppression of the activation of the NF-κB
signaling pathway. This event led to restricted expression of the
downstream protein Bcl-2 and the promotion of apoptosis in the
gastric cancer SGC7901 cell line (73). Another study indicated that the
expression of miR-146a was upregulated in 73% of gastric cancer
tissue samples and that miR-146a also downregulated the expressions
of caspase recruitment domain-containing protein 10 (CARD10), COP9
constitutive photomorphogenic homolog subunit 8 (COPS8) and
interleukin-1 receptor-associated kinase 1 (IRAK1) in vitro
(72). It has previously been
confirmed that IRAK1 is involved in the activation of the NF-κB
signaling pathway (74,75). In this study, it was proved that
miR-146a overexpression is able to decrease CARD10 and COPS8
expression levels, which prevented G protein-coupled receptor from
activating the NF-κB signaling pathway. Furthermore, miR-146a was
able to downregulate the expression of IL-8, IL-23A, CCL5,
colony-stimulatory factor-1 and platelet-derived growth factor
subunit B, which are involved in facilitating the occurrence and
development of tumors (72). The
morbidity rate of Helicobacter pylori infection is high and
half of the world's population may be a carrier of this bacterium
(76), which has been proved to be
able to promote or inhibit the expression of certain miRNAs and
enhance the proliferation and invasion of gastric cancer cells
(77-79).
In the serum of patients with gastric cancer with Helicobacter
pylori infection, miR-375, miR-146 and let-7 were
downregulated, while miR-19 and miR-21 were upregulated and the
Wnt/β-catenin, IRAK and NF-κB signaling pathways were activated
(80). In patients with
Helicobacter pylori infection, IL-17A expression was
increased, which is able to increase the levels of downstream
products, including IL-8 and growth-regulated oncogene α (GRO-α),
and enhance the local inflammatory response through upregulating
the NF-κB signaling pathway. However, IL-17A also promoted the
expression of miR-146a, which was able to reduce the expression
levels of GRO-α and IL-8 in the gastric cancer SGC7901 cell line.
In addition, miR-146a is a dependent gene of the NF-κB signaling
pathway that may inhibit the activation of the NF-κB signaling
pathway via reducing the expression of TRAF6 and IRAK1, thus
reducing the release of inflammatory factors and inhibiting the
inflammatory response (81).
miR-195 inhibition of the NF-κB
signaling pathway
Surgery is currently the preferred treatment method
for gastric cancer (5). Studies
have indicated that certain anesthetics may affect the activation
of the NF-κB signaling pathway through their influence on the
expression of miRNAs (82-84).
Propofol is a commonly used intravenous anesthetic in clinical
anesthesia, characterized by rapid onset and relatively few side
effects. Studies have demonstrated that propofol possesses an
anti-tumor effect against a variety of tumor types through its
inhibitory effect on the proliferation of gastric cancer cells via
regulating MMP2 and inhibitor of growth protein 3 (85,86).
As a tumor suppressor, miR-195 is downregulated in various types of
cancer tissue and cells (87), but
its expression in gastric cancer cells may be upregulated by
propofol, which leads to the activation of the NF-κB and JAK/STAT
signaling pathways, inhibition of cell proliferation, migration and
invasion, and induction cell apoptosis (82). However, the molecular mechanisms
underlying the inhibitory effects of propofol on gastric cancer
growth remain to be fully elucidated.
miR-145 inhibition of the NF-κB
signaling pathway
The amide lidocaine is also commonly used as a local
anesthetic that exerts an important therapeutic effect on rapid
ventricular arrhythmia. A study confirmed that lidocaine was able
to restrain the growth of a variety of tumor types and that the
tumor suppressor miR-145 inhibits the growth of gastric cancer
cells (88). Lidocaine inhibits the
activation of the NF-κB and MEK/ERK signaling pathways by
upregulating the expression of miR-145 in the gastric cancer MKN45
cell line, indicating that miR-145 is a potential therapeutic
target for gastric cancer (83). An
intravenous defined dose of lidocaine exerts a certain treatment
effect on post-operative chronic pain, post-operative cognitive
dysfunction and malignant tumors (89). Perioperative anesthetics have
important effects on gastric cancer, and therefore, understanding
the mechanisms of action of drugs on tumors may serve as an
important guidance for the peri-operative use of anesthetics in
radical gastrectomy.
miR-128b inhibition of the NF-κB
signaling pathway
miR-128b expression declines in gastric cancer
tissues and SGC7901, GC-823 and HGC-27 cell lines, compared with
that in normal tissues and gastric mucosal epithelial GES1 cell
lines. It has been indicated that miR-128b is able to inhibit the
target gene pyruvate dehydrogenase kinase 1, thereby suppressing
the Akt/NF-κB signaling pathway, promoting apoptosis and
restraining proliferation, migration and invasion of gastric cancer
cells (90).
miR-3664-5P inhibition of the NF-κB
signaling pathway
miR-3664-5P is downregulated in gastric cancer
tissues and cell lines and its expression positively correlates
with gastric cancer differentiation, degree of malignancy, tumor
size and patient prognosis. Metadherin (MTDH), a transmembrane
protein, is currently considered to be an oncogene in a variety of
cancer types, which is able to enhance tumor cell proliferation,
invasion, migration, angiogenesis and chemoresistance.
Bioinformatics analysis and a luciferase reporter gene assay
confirmed that MTDH is a target gene of miR-3664-5P and that its
expression is decreased following upregulation of miR-3664-5P,
resulting in the inhibition of the activation of the NF-κB
signaling pathway. Furthermore, miR-3664-5P is able to restrain the
growth and pulmonary metastasis of gastric cancer cells in nude
mice (91).
The above-mentioned miRNAs act as tumor suppressors
in gastric cancer cells and participate in the regulation on the
NF-κB signaling pathway, which is involved in regulating the
expression of a variety of genes. However, the imbalance of miRNAs
and their regulatory mechanisms in gastric cancer remain to be
fully clarified. The mutual regulatory pathways of miRNAs and the
NF-κB signaling pathway in gastric cancer are summarized in
Tables I and II. These schematic summaries may
contribute to subsequent related research and provide a basis for
the development of novel methods for preventing, detecting and
treating gastric cancer.
 | Table ImiRNAs that regulate the NF-κB
signaling pathway in gastric cancer. |
Table I
miRNAs that regulate the NF-κB
signaling pathway in gastric cancer.
| miRNA | Target gene | NF-κB | Protein | Function | (Refs.) |
|---|
| miR-20a | CYLD (-) | (+) | Livin, survivin
(+) | Proliferation,
migration, invasion (+); apoptosis, chemosensitivity (-) | (27,28) |
| miR-21 | - | (+) | Caspase-3 (-) | Apoptosis (-) | (39) |
| miR-17-92 | TRAF3 (-) | (+) | - | Proliferation,
migration, invasion (+) Apoptosis (-) | (42) |
| miR-577 | SDPR (-) | (+) | E-cadherin (-) | Migration, invasion
(+); Chemosensitivity (-) | (44) |
| miR-216a-3p | RUNX1 (-) | (+) | MMP2, MMP9,
CyclinD1, Bcl-2 (+) | Proliferation,
migration, invasion (+) | (46) |
| miR-362 | CYLD (-) | (+) | - | Proliferation (+);
Apoptosis, chemosensitivity (-) | (48) |
| miR-10b | CSMD1 (-) | (+) | c-Myc, cyclin D1
(+) | Migration, invasion
(+) | (50) |
| miR-224 | RASSF8 (-) | (+) | - | Proliferation,
migration, invasion (+) | (36) |
| miR-3664-5p | MTPH (-) | (-) | IL-8, MMP9, VEGF
(-) | Proliferation,
migration, invasion (-); Apoptosis (+) | (91) |
| miR-7 | RELA (-) | (-) | VCAM-1, VEGF, MMP-9
MMP-2, ICAM-1 (-) | Proliferation,
migration, invasion (-) | (63,65) |
| miR-218 | - | (-) | POU2F2/SLIT2/ROBO1
(-) | Migration, invasion
(-) | (67) |
| miR-195 | - | (-) | MMP-2, MMP-9,
Bcl-2, cyclinD1, vimentin (-) Bax, P21, caspase-3, 9 (+) | Proliferation,
migration, invasion (-); Apoptosis (+) | (82) |
| miR-145 | - | (-) | MMP-2, MMP-9,
Bcl-2, cyclinD1, vimentin (-); P21, caspase-3, 7, 9 (+) | Proliferation,
migration, invasion (-); Apoptosis (+) | (83) |
| miR-128b | PDK1 (-) | (-) | Bcl-2 (-) | Proliferation,
invasion (-); Apoptosis (+) | (90) |
| miR-146a | CARD10, COPS8
(-) | (-) | - | Proliferation
(-) | (72) |
| miR-146a | TRAF6/IRAK1
(-) | (-) | GRO-α, IL-8
(-) | Inflammation
(-) | (81) |
| miR-146a | TAK1 (-) | (-) | Bcl-2 (-) | Apoptosis (+) | (73) |
| Table IIRegulation of miRNAs through the
NF-κB signaling pathway in gastric cancer. |
Table II
Regulation of miRNAs through the
NF-κB signaling pathway in gastric cancer.
| Signaling
molecule | NF-κB | miRNAs | Target genes | Function | (Refs.) |
|---|
| CagA (+) | (+) | miR-223-3p | ARID1A
(-)/E-cadherin, P21 (-) | Migration, invasion
(+) | (51) |
| HP (+) | (+) | miR-135-5p | KLF4 (-) | Apoptosis,
chemosensitivity (-) | (53) |
| IL-1β | (+) | miR-425 | PTEN (-) | Proliferation
(+) | (54) |
| - | (+) | miR-107 | FOXO1 (-) | Proliferation
(+) | (59) |
| PRL-3 | (+) | miR-210 | - | Migration, invasion
(+) | (60) |
| EP2/4 | (+) | miR-21, miR-16 | - | Proliferation
(+) | (38) |
| CBX7 | (+) | miR-21 | - | Stem cell-like
properties (-) | (40) |
3. lncRNAs and the NF-κB signaling
pathway
There is also an important link between lncRNAs and
gastric cancer. For instance, in the blood and tissue samples of
patients with cancer, long intergenic non-protein coding RNA CYTOR
(LINC00152) and H19 expression levels positively correlate with the
incidence rate of gastric cancer. NcRNA 1 (GClnc1), LINC02864 and
TMEM132D-AS1 are associated with the survival rate of patients with
gastric cancer and dysregulated lncRNAs may be closely related to
Helicobacter pylori infection (92). Furthermore, lncRNAs are involved in
the regulation of important signaling pathways, such as the NF-κB,
PI3K/AKT, Wnt/β-catenin and ERK/MAPK signaling pathways (93). In the chapter below, the regulation
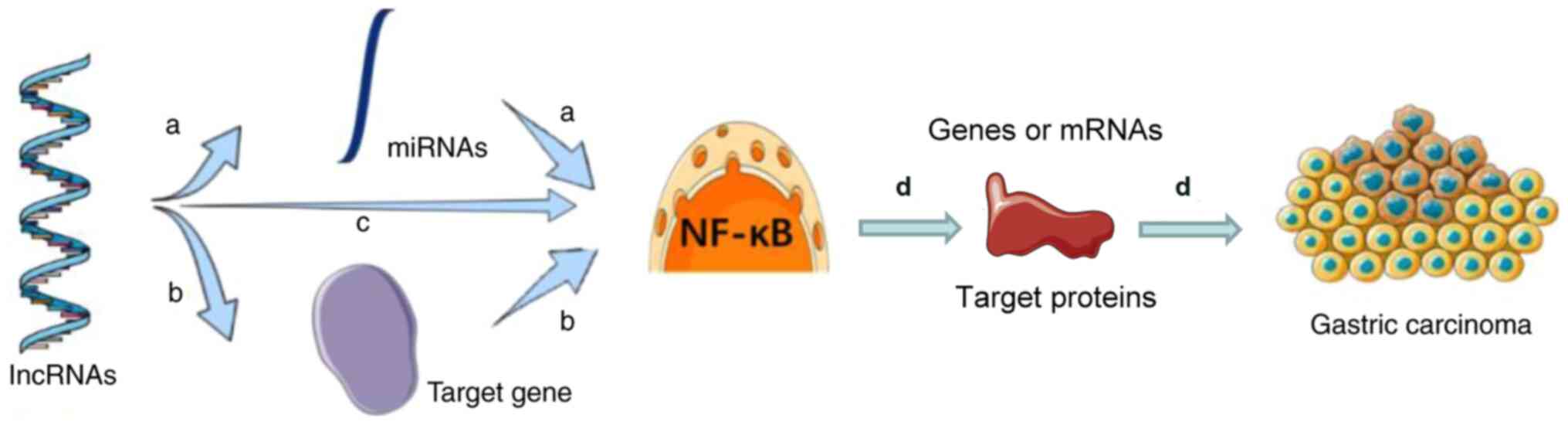
of lncRNAs of the NF-κB signaling pathway is discussed. The general
regulatory mechanisms of lncRNAs and the NF-κB signaling pathway in
gastric cancer are illustrated in Fig.
2. lncRNAs regulate miRNAs to activate or inhibit the NF-κB
signaling pathway. lncRNAs also regulate target genes to activate
or the inhibit NF-κB signaling pathway. Furthermore, lncRNAs can
activate or inhibit the NF-κB signaling pathway directly. The NF-κB
signaling pathway therefore influences the development of gastric
cancer by regulating downstream genes, mRNAs or proteins (94).
CDKN2B-AS1 (lncRNA ANRIL)
It was previously indicated that ten-eleven
translocation-2 (TET2) is an important regulator of certain key
physiological functions of the body. The TET2 protein has a low
expression level in gastric cancer cells, where it is involved in
apoptosis. Its expression also correlates with the prognosis of
patients with gastric cancer. CDKN2B-AS1 (lncRNA ANRIL) is located
at the human CDKN2A/B locus at 9p21.3(95). It is closely related to the
occurrence and development of various tumor types and it is highly
expressed in tumor tissues of certain patients with gastric cancer,
where it promotes migration and inhibits apoptosis of gastric
cancer cells. Using a nude mouse model of metastasis, researchers
confirmed that TET2 protein inhibits pulmonary metastasis of
gastric cancer through suppressing lncRNA ANRIL. The anti-tumor
properties of TET2 depend on lncRNA ANRIL, while this later
promotes the progression of gastric cancer through activating the
NF-κB signaling pathway. Furthermore, the NF-κB signaling pathway
is able to upregulate the expression of p21, baculoviral IAP repeat
containing 2, X-linked inhibitor of apoptosis, cyclin D1, CDK2,
proline rich acidic protein 1, MMP2 and anosmin 1. Therefore,
lncRNA ANRIL may be a potential therapeutic target for gastric
cancer (96).
BANCR
It has been widely accepted that lncRNAs act as
miRNA sponges that affect the expression of genes/mRNAs. BANCR is
an lncRNA containing 693 nucleotides that has four exons located on
chromosome 9(97), whose level in
human gastric cancer tissues is 5 times higher than that in normal
gastric tissues. BGC-803 and MGC-823 cells have a higher level of
BANCR than that of GES-1 cells. It has been confirmed that BANCR
activates the NF-κB signaling pathway, facilitates proliferation
and suppresses apoptosis of gastric cancer cells through inhibiting
the expression of miRNA-9(98).
LINC01410
In gastric cancer cells, LINC01410 was indicated to
inhibit the expression of miR-532-5p, which is involved in the
inhibition of neutrophil cytosolic factor 2 (NCF2) expression, thus
suppressing the activation of the NF-κB pathway. Additionally,
Related genes downstream of the NF-κB pathway such as MMP1, MMP2,
MMP9 and CASP8 and FADD like apoptosis regulator are downregulated.
Through affecting the peripheral angiogenesis and metastasis of
gastric cancer cells, the LINC01410/miR-532/NCF2/NF-κB axis
inhibits the progression of gastric cancer (99).
KRT19P3
Certain lncRNAs may also inhibit the development of
gastric cancer. KRT19P3 is downregulated in gastric cancer tissues.
Through its binding to COP9 signalosome subunit 7A and upregulation
of IκBα, KRT19P3 inhibits the activation of the NF-κB signaling
pathway, which facilitates apoptosis and suppresses the
proliferation of gastric cancer cells. As an important molecular
marker for patients with gastric cancer, KRT19P3 is expected to
become a novel therapeutic tumor target (100).
4. Conclusion
Gastric cancer, a disease with high morbidity and
mortality rates worldwide, has imposed a severe clinical burden
(101). A comprehensive
exploration of the mechanisms of gastric cancer has been performed
and is ongoing, and novel and early screening methods have been
proposed. Surgery is still the major treatment method for gastric
cancer, but numerous targeted drugs and immunotherapy drugs are
being used for treatment of gastric cancer. Physicians and
researchers around the world are making continuous efforts to put
forward novel ideas regarding the etiology, pathogenesis, diagnosis
and treatment of gastric cancer.
In the present review, recent and relevant research
results were summarized. The NF-κB signaling pathway is present in
almost all cells and is involved in numerous physiological
functions. This pathway is overactivated in gastric cancer and in
most cases, it participates in the occurrence and development of
gastric cancer and is associated with unfavorable prognosis of
patients with gastric cancer. The activation of the NF-κB signaling
pathway may induce an immunosuppressive microenvironment that
enhances tumor development, while its inhibition may be a promising
approach for the treatment of gastric cancer (102). miRNAs and lncRNAs were originally
considered meaningless molecules, but they have now been confirmed
to have crucial roles in various functions of the human body. In
gastric cancer, the NF-κB signaling pathway has an important link
with miRNAs and lncRNAs, which exert important effects on the
occurrence and development of gastric cancer. Furthermore, the
interactions of miRNAs/target genes/NF-κB/target proteins,
signaling molecules/NF-κB/miRNAs/target genes,
lncRNAs/miRNAs/NF-κB/genes or mRNAs, lncRNAs/target
genes/NF-κB/target proteins and lncRNAs/NF-κB/target proteins
pathways also appear to offer opportunities for subsequent research
and clinical treatments.
Acknowledgements
Not applicable.
Funding
Funding: This project was supported by the Chinese National
Science Foundation (grant no. 81172210), the China Postdoctoral
Science Foundation (grant no. 2012M521528), the open fund of the
Chinese Hunan Provincial Education Department Innovation Platform
(grant no. 15K109) and the fund of Hunan Provincial Science and
Technology Department (grant no. 2018SK51702).
Availability of data and materials
Not applicable.
Authors' contributions
WH and RC conceived the study, RC and MY performed
the literature search and data analysis and drafted the manuscript
and BW critically revised the manuscript. Data authentication is
not applicable. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Biagioni A, Skalamera I, Peri S, Schiavone
N, Cianchi F, Giommoni E, Magnelli L and Papucci L: Update on
gastric cancer treatments and gene therapies. Cancer Metast Rev.
38:537–548. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Abozeid M, Rosato A and Sommaggio R:
Immunotherapeutic strategies for gastric carcinoma: A review of
preclinical and clinical recent development. Biomed Res Int.
2017(5791262)2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Schernberg A, Rivin Del Campo E, Rousseau
B, Matzinger O, Loi M, Maingon P and Huguet F: Adjuvant
chemoradiation for gastric carcinoma: State of the art and
perspectives. Clin Transl Radiat Oncol. 10:13–22. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Song Z, Wu Y, Yang J, Yang D and Fang X:
Progress in the treatment of advanced gastric cancer. Tumour Biol.
39(1010428317714626)2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhang HD, Jiang LH, Sun DW, Li J and Ji
ZL: The role of miR-130a in cancer. Breast Cancer. 24:521–527.
2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Chen Q, Lu X and Zhang X: Noncanonical
NF-κB signaling pathway in liver diseases. J Clin Transl Hepatol.
9:81–89. 2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Smale ST: Hierarchies of NF-κB target-gene
regulation. Nat Immunol. 12:689–694. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
9
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer.
12(86)2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen Z, Liu M, Liu X, Huang S, Li L, Song
B, Li H, Ren Q, Hu Z, Zhou Y and Qiao L: COX-2 regulates E-cadherin
expression through the NF-kappaB/Snail signaling pathway in gastric
cancer. Int J Mol Med. 32:93–100. 2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Koyama S: Differential expression of
intracellular apoptotic signaling molecules in tumor and
tumor-infiltrating lymphocytes during development of invasion
and/or metastasis of gastric carcinoma. Dig Dis Sci. 48:2290–2300.
2003.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yamanaka N, Sasaki N, Tasaki A, Nakashima
H, Kubo M, Morisaki T, Noshiro H, Yao T, Tsuneyoshi M, Tanaka M and
Katano M: Nuclear factor-kappaB p65 is a prognostic indicator in
gastric carcinoma. Anticancer Res. 24:1071–1075. 2004.PubMed/NCBI
|
|
13
|
Lee BL, Lee HS, Jung J, Cho SJ, Chung HY,
Kim WH, Jin YW, Kim CS and Nam SY: Nuclear factor-kappaB activation
correlates with better prognosis and Akt activation in human
gastric cancer. Clin Cancer Res. 11:2518–2525. 2005.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Dong H, Lei J, Ding L, Wen Y, Ju H and
Zhang X: MicroRNA: Function, detection, and bioanalysis. Chem Rev.
113:6207–6233. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Su W, Mo Y, Wu F, Guo K, Li J, Luo Y, Ye
H, Guo H, Li D and Yang Z: MiR-135b reverses chemoresistance of
non-small cell lung cancer cells by downregulation of FZD1. Biomed
Pharmacother. 84:123–129. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Nezu Y, Hagiwara K, Yamamoto Y, Fujiwara
T, Matsuo K, Yoshida A, Kawai A, Saito T and Ochiya T: MiR-135b, a
key regulator of malignancy, is linked to poor prognosis in human
myxoid liposarcoma. Oncogene. 35:6177–6188. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Majeed W, Iftikhar A, Khaliq T, Aslam B,
Muzaffar H, Atta K, Mahmood A and Waris S: Gastric carcinoma:
Recent trends in diagnostic biomarkers and molecular targeted
therapies. Asian Pac J Cancer Prev. 17:3053–3060. 2016.PubMed/NCBI
|
|
18
|
Matsuoka T and Yashiro M: Biomarkers of
gastric cancer: Current topics and future perspective. World J
Gastroenterol. 24:2818–2832. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Sanchez Calle A, Kawamura Y, Yamamoto Y,
Takeshita F and Ochiya T: Emerging roles of long non-coding RNA in
cancer. Cancer Sci. 109:2093–2100. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Peng L, Yuan X, Jiang B, Tang Z and Li G:
lncRNAs: Key players and novel insights into cervical cancer.
Tumour Biol. 37:2779–2788. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Li J, Meng H, Bai Y and Wang K: Regulation
of lncRNA and its role in cancer metastasis. Oncol Res. 23:205–217.
2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Dinescu S, Ignat S, Lazar AD, Constantin
C, Neagu M and Costache M: Epitranscriptomic Signatures in lncRNAs
and their possible roles in cancer. Genes (Basel).
10(52)2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ghafouri-Fard S, Abak A, Fattahi F, Hussen
BM, Bahroudi Z, Shoorei H and Taheri M: The interaction between
miRNAs/lncRNAs and nuclear factor-κB (NF-κB) in human disorders.
Biomed Pharmacother. 138(111519)2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Li M and Guan H: Noncoding RNAs regulating
NF-κB signaling. Adv Exp Med Biol. 927:317–336. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chen W, Li Z, Bai L and Lin Y: NF-kappaB
in lung cancer, a carcinogenesis mediator and a prevention and
therapy target. Front Biosci (Landmark Ed). 16:1172–1185.
2011.PubMed/NCBI View
Article : Google Scholar
|
|
27
|
Li X, Zhang Z, Yu M, Li L, Du G, Xiao W
and Yang H: Involvement of miR-20a in promoting gastric cancer
progression by targeting early growth response 2 (EGR2). Int J Mol
Sci. 14:16226–16239. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhu M, Zhou X, Du Y, Huang Z, Zhu J, Xu J,
Cheng G, Shu Y, Liu P, Zhu W and Wang T: MiR-20a induces cisplatin
resistance of a human gastric cancer cell line via targeting CYLD.
Mol Med Rep. 14:1742–1750. 2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ma X and Ning S: Shikimic acid promotes
estrogen receptor(ER)-positive breast cancer cells proliferation
via activation of NF-κB signaling. Toxicol Lett. 312:65–71.
2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Yu J, Xie F, Bao X, Chen W and Xu Q:
MiR-300 inhibits epithelial to mesenchymal transition and
metastasis by targeting Twist in human epithelial cancer. Mol
Cancer. 13(121)2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kang Y, Zhang Y, Sun Y, Wen Y and Sun F:
MicroRNA-300 suppresses metastasis of oral squamous cell carcinoma
by inhibiting epithelial-to-mesenchymal transition. Onco Targets
Ther. 11:5657–5666. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
He FY, Liu HJ, Guo Q and Sheng JL: Reduced
miR-300 expression predicts poor prognosis in patients with
laryngeal squamous cell carcinoma. Eur Rev Med Pharmacol Sci.
21:760–764. 2017.PubMed/NCBI
|
|
33
|
Cui R, Kim T, Fassan M, Meng W, Sun HL,
Jeon YJ, Vicentini C, Tili E, Peng Y, Scarpa A, et al: MicroRNA-224
is implicated in lung cancer pathogenesis through targeting
caspase-3 and caspase-7. Oncotarget. 6:21802–21815. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
He X, Zhang Z, Li M, Li S, Ren L, Zhu H,
Xiao B and Shi R: Expression and role of oncogenic miRNA-224 in
esophageal squamous cell carcinoma. BMC Cancer.
15(575)2015.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Shen G, Li X, Jia YF, Piazza GA and Xi Y:
Hypoxia-regulated microRNAs in human cancer. Acta Pharmacol Sin.
34:336–341. 2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
He C, Wang L, Zhang J and Xu H:
Hypoxia-inducible microRNA-224 promotes the cell growth, migration
and invasion by directly targeting RASSF8 in gastric cancer. Mol
Cancer. 16(35)2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Huang Y, Yang YB, Zhang XH, Yu XL, Wang ZB
and Cheng XC: MicroRNA-21 gene and cancer. Med Oncol.
30(376)2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Shin VY, Jin H, Ng EK, Cheng AS, Chong WW,
Wong CY, Leung WK, Sung JJ and Chu KM: NF-κB targets miR-16 and
miR-21 in gastric cancer: Involvement of prostaglandin E receptors.
Carcinogenesis. 32:240–245. 2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Sha M, Ye J, Zhang LX, Luan ZY, Chen YB
and Huang JX: Celastrol induces apoptosis of gastric cancer cells
by miR-21 Inhibiting PI3K/Akt-NF-κB signaling pathway.
Pharmacology. 93:39–46. 2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Ni S, Zhao L, Wang X, Wu ZH, Hua RX, Wan
CH, Zhang JY, Zhang XW, Huang MZ, Gan L, et al: CBX7 regulates stem
cell-like properties of gastric cancer cells via p16 and
AKT-NF-κB-miR-21 pathways. J Hematol Oncol. 11(17)2018.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhang X, Li Y, Qi P and Ma Z: Biology of
MiR-17-92 cluster and its progress in lung cancer. Int J Med Sci.
15:1443–1448. 2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Liu F, Cheng L, Xu J, Guo F and Chen W:
MiR-17-92 functions as an oncogene and modulates NF-κB signaling by
targeting TRAF3 in MGC-803 human gastric cancer cells. Int J Oncol.
53:2241–2257. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yang XD and Sun SC: Targeting signaling
factors for degradation, an emerging mechanism for TRAF functions.
Immunol Rev. 266:56–71. 2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Luo Y, Wu J, Wu Q, Li X, Wu J, Zhang J,
Rong X, Rao J, Liao Y, Bin J, et al: MiR-577 regulates TGF-β
induced cancer progression through a SDPR-Modulated
positive-feedback loop with ERK-NF-κB in gastric cancer. Mol Ther.
27:1166–1182. 2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Li N, Zhang QY, Zou JL, Li ZW, Tian TT,
Dong B, Liu XJ, Ge S, Zhu Y, Gao J and Shen L: MiR-215 promotes
malignant progression of gastric cancer by targeting RUNX1.
Oncotarget. 7:4817–4828. 2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Wu Y, Zhang J, Zheng Y, Ma C, Liu XE and
Sun X: MiR-216a-3p inhibits the proliferation, migration, and
invasion of human gastric cancer cells via targeting RUNX1 and
activating the NF-κB signaling pathway. Oncol Res. 26:157–171.
2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Mathis BJ, Lai Y, Qu C, Janicki JS and Cui
T: CYLD-mediated signaling and diseases. Curr Drug Targets.
16:284–294. 2015.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Xia JT, Chen LZ, Jian WH, Wang KB, Yang
YZ, He WL, He YL, Chen D and Li W: MicroRNA-362 induces cell
proliferation and apoptosis resistance in gastric cancer by
activation of NF-κB signaling. J Transl Med. 12(33)2014.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Wang YY, Ye ZY, Zhao ZS, Li L, Wang YX,
Tao HQ, Wang HJ and He XJ: Clinicopathologic significance of
miR-10b expression in gastric carcinoma. Hum Pathol. 44:1278–1285.
2013.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Chen XL, Hong LL, Wang KL, Liu X, Wang JL,
Lei L, Xu ZY, Cheng XD and Ling ZQ: Deregulation of CSMD1 targeted
by microRNA-10b drives gastric cancer progression through the NF-κB
pathway. Int J Biol Sci. 15:2075–2086. 2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Yang F, Xu Y, Liu C, Ma C, Zou S, Xu X,
Jia J and Liu Z: NF-κB/miR-223-3p/ARID1A axis is involved in
Helicobacter pylori CagA-induced gastric carcinogenesis and
progression. Cell Death Dis. 9(12)2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Murata M: Inflammation and cancer. Environ
Health Prev. 23(50)2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Shao L, Chen Z, Soutto M, Zhu S, Lu H,
Romero-Gallo J, Peek R, Zhang S and El-Rifai W: Helicobacter
pylori-induced miR-135b-5p promotes cisplatin resistance in
gastric cancer. FASEB J. 33:264–274. 2019.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Ma J, Liu J, Wang Z, Gu X, Fan Y, Zhang W,
Xu L, Zhang J and Cai D: NF-kappaB-dependent microRNA-425
upregulation promotes gastric cancer cell growth by targeting PTEN
upon IL-1β induction. Mol Cancer. 13(40)2014.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Yang SM, Huang C, Li XF, Yu MZ, He Y and
Li J: MiR-21 confers cisplatin resistance in gastric cancer cells
by regulating PTEN. Toxicology. 306:162–168. 2013.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Wu W, Yang J, Feng X, Wang H, Ye S, Yang
P, Tan W, Wei G and Zhou Y: MicroRNA-32 (miR-32) regulates
phosphatase and tensin homologue (PTEN) expression and promotes
growth, migration, and invasion in colorectal carcinoma cells. Mol
Cancer. 12(30)2013.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Luo Z, Zheng Y and Zhang W: Pleiotropic
functions of miR107 in cancer networks. Onco Targets Ther.
11:4113–4124. 2018.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Zheng Q, Chen C, Guan H, Kang W and Yu C:
Prognostic role of microRNAs in human gastrointestinal cancer: A
systematic review and meta-analysis. Oncotarget. 8:46611–46623.
2017.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Li F, Liu B, Gao Y, Liu Y, Xu Y, Tong W
and Zhang A: Upregulation of MicroRNA-107 induces proliferation in
human gastric cancer cells by targeting the transcription factor
FOXO1. FEBS Lett. 588:538–544. 2014.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Zhang C, Tian W, Meng L, Qu L and Shou C:
PRL-3 promotes gastric cancer migration and invasion through a
NF-κB-HIF-1α-miR-210 axis. J Mol Med (Berl). 94:401–415.
2016.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Li Y, Bai W and Zhang X: Identifying
heterogeneous subtypes of gastric cancer and subtype-specific
subpaths of microRNA-target pathways. Mol Med Rep. 17:3583–3590.
2018.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Kalinowski FC, Brown RA, Ganda C, Giles
KM, Epis MR, Horsham J and Leedman PJ: MicroRNA-7: A tumor
suppressor miRNA with therapeutic potential. Int J Biochem Cell
Biol. 54:312–317. 2014.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Zhao XD, Lu YY, Guo H, Xie HH, He LJ, Shen
GF, Zhou JF, Li T, Hu SJ, Zhou L, et al: MicroRNA-7/NF-κB signaling
regulatory feedback circuit regulates gastric carcinogenesis. J
Cell Biol. 210:613–627. 2015.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Li M, Pan M, You C and Dou J: The
therapeutic potential of miR-7 in cancers. Mini Rev Med Chem.
19:1707–1716. 2019.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Ye T, Yang M, Huang D, Wang X, Xue B, Tian
N, Xu X, Bao L, Hu H, Lv T and Huang Y: MicroRNA-7 as a potential
therapeutic target for aberrant NF-κB-driven distant metastasis of
gastric cancer. J Exp Clin Canc Res. 38(55)2019.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Chen WQ, Hu L, Chen GX and Deng HX: Role
of microRNA-7 in digestive system malignancy. World J Gastrointest
Oncol. 8:121–127. 2016.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Wang SM, Tie J, Wang WL, Hu SJ, Yin JP, Yi
XF, Tian ZH, Zhang XY, Li MB, Li ZS, et al: POU2F2-oriented network
promotes human gastric cancer metastasis. Gut. 65:1427–1438.
2016.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Guo Y, Zhang T, Shi Y, Zhang J, Li M, Lu
F, Zhang J, Chen X and Ding S: Helicobacter pylori inhibits
GKN1 expression via the CagA/p-ERK/AUF1 pathway. Helicobacter.
25(e12665)2020.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Chung Nien Chin S, O'Connor L, Scurr M,
Busada JT, Graham AN, Alipour Talesh G, Tran CP, Sarkar S, Minamoto
T, Giraud AS, et al: Coordinate expression loss of GKN1 and GKN2 in
gastric cancer via impairment of a glucocorticoid-responsive
enhancer. Am J Physiol Gastrointest Liver Physiol. 319:G175–G188.
2020.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Zhang Z, Xue H, Dong Y, Zhang J, Pan Y,
Shi L, Xiong P, Zhu J, Li W, Zheng W, et al: GKN2 promotes
oxidative stress-induced gastric cancer cell apoptosis via the
Hsc70 pathway. J Exp Clin Cancer Res. 38(338)2019.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Kim O, Yoon JH, Choi WS, Ashktorab H,
Smoot DT, Nam SW, Lee JY and Park WS: GKN2 contributes to the
homeostasis of gastric mucosa by inhibiting GKN1 activity. J Cell
Physiol. 229:762–771. 2014.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Crone SG, Jacobsen A, Federspiel B,
Bardram L, Krogh A, Lund AH and Friis-Hansen L: MicroRNA-146a
inhibits G protein-coupled receptor-mediated activation of NF-κB by
targeting CARD10 and COPS8 in gastric cancer. Mol Cancer.
11(71)2012.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Chen Y, Zhou B, Xu L, Fan H, Xie J and
Wang D: MicroRNA-146a promotes gastric cancer cell apoptosis by
targeting transforming growth factor β-activated kinase 1. Mol Med
Rep. 16:755–763. 2017.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Meng DF, Sun R, Liu GY, Peng LX, Zheng LS,
Xie P, Lin ST, Mei Y, Qiang YY, Li CZ, et al: S100A14 suppresses
metastasis of nasopharyngeal carcinoma by inhibition of NF-κB
signaling through degradation of IRAK1. Oncogene. 39:5307–5322.
2020.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Wee ZN, Yatim SM, Kohlbauer VK, Feng M,
Goh JY, Bao Y, Lee PL, Zhang S, Wang PP, Lim E, et al: IRAK1 is a
therapeutic target that drives breast cancer metastasis and
resistance to paclitaxel. Nat Commun. 6(8746)2015.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Diaconu S, Predescu A, Moldoveanu A, Pop
CS and Fierbinteanu-Braticevici C: Helicobacter pylori
infection: Old and new. J Med Life. 10:112–117. 2017.PubMed/NCBI
|
|
77
|
Matsushima K, Isomoto H, Inoue N, Nakayama
T, Hayashi T, Nakayama M, Nakao K, Hirayama T and Kohno S: MicroRNA
signatures in Helicobacter pylori-infected gastric mucosa.
Int J Cancer. 128:361–370. 2011.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Darnet S, Moreira FC, Hamoy IG, Burbano R,
Khayat A, Cruz A, Magalhães L, Silva A, Santos S, Demachki S, et
al: High-Throughput sequencing of miRNAs reveals a tissue signature
in gastric cancer and suggests novel potential biomarkers.
Bioinform Biol Insights. 9 (Suppl 1):1–8. 2015.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Assumpcao MB, Moreira FC, Hamoy IG,
Magalhães L, Vidal A, Pereira A, Burbano R, Khayat A, Silva A,
Santos S, et al: High-Throughput miRNA sequencing reveals a field
effect in gastric cancer and suggests an epigenetic network
mechanism. Bioinform Biol Insights. 9:111–117. 2015.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Ranjbar R, Hesari A, Ghasemi F and
Sahebkar A: Expression of microRNAs and IRAK1 pathway genes are
altered in gastric cancer patients with Helicobacter pylori
infection. J Cell Biochem. 119:7570–7576. 2018.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Li N, Wang J, Yu W, Dong K, You F, Si B,
Tang B, Zhang Y, Wang T and Qiao B: MicroRNA-146a inhibits the
inflammatory responses induced by interleukin-17A during the
infection of Helicobacter pylori. Mol Med Rep. 19:1388–1395.
2019.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Zhang W, Wang Y, Zhu Z, Zheng Y and Song
B: Propofol inhibits proliferation, migration and invasion of
gastric cancer cells by up-regulating microRNA-195. INT J Biol
Macromol. 120(Pt A):975–984. 2018.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Sui H, Lou A, Li Z and Yang J: Lidocaine
inhibits growth, migration and invasion of gastric carcinoma cells
by up-regulation of miR-145. BMC CANCER. 19(233)2019.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Xu L, Ge F, Hu Y, Yu Y, Guo K and Miao C:
Sevoflurane postconditioning attenuates hepatic
ischemia-reperfusion injury by limiting HMGB1/TLR4/NF-κB Pathway
via modulating microRNA-142 in vivo and in vitro. Front Pharmacol.
12(646307)2021.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Yang C, Gao J, Yan N, Wu B, Ren Y, Li H
and Liang J: Propofol inhibits the growth and survival of gastric
cancer cells in vitro through the upregulation of ING3.
Oncol Rep. 37:587–593. 2017.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Peng Z and Zhang Y: Propofol inhibits
proliferation and accelerates apoptosis of human gastric cancer
cells by regulation of microRNA-451 and MMP-2 expression. Genet Mol
Res 15, 2016.
|
|
87
|
Yu W, Liang X, Li X, Zhang Y, Sun Z, Liu Y
and Wang J: MicroRNA-195: A review of its role in cancers. Onco
Targets Ther. 11:7109–7123. 2018.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Qiu T, Zhou X, Wang J, Du Y, Xu J, Huang
Z, Zhu W, Shu Y and Liu P: MiR-145, miR-133a and miR-133b inhibit
proliferation, migration, invasion and cell cycle progression via
targeting transcription factor Sp1 in gastric cancer. FEBS Lett.
588:1168–1177. 2014.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Soto G, Naranjo González M and Calero F:
Intravenous lidocaine infusion. Rev Esp Anestesiol Reanim.
65:269–274. 2018.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Zhang L, Lei J, Fang ZL and Xiong JP:
MiR-128b is down-regulated in gastric cancer and negatively
regulates tumour cell viability by targeting PDK1/Akt/NF-κB axis. J
Biosci. 41:77–85. 2016.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Jiao Y, Yang H, Qian J, Gong Y, Liu H, Wu
S, Cao L and Tang L: MiR-3664-5P suppresses the proliferation and
metastasis of gastric cancer by attenuating the NF-κB signaling
pathway through targeting MTDH. Int J Oncol. 54:845–858.
2019.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Dastmalchi N, Khojasteh SMB, Nargesi MM
and Safaralizadeh R: The correlation between lncRNAs and
Helicobacter pylori in gastric cancer. Pathog Dis.
77(ftaa004)2019.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Peng WX, Koirala P and Mo YY:
lncRNA-mediated regulation of cell signaling in cancer. Oncogene.
36:5661–5667. 2017.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Gupta SC, Awasthee N, Rai V, Chava S,
Gunda V and Challagundla KB: Long non-coding RNAs and nuclear
factor-κB crosstalk in cancer and other human diseases. Biochim
Biophys Acta Rev Cancer. 1873(188316)2020.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Kong Y, Hsieh CH and Alonso LC: ANRIL: A
lncRNA at the CDKN2A/B Locus with roles in cancer and metabolic
disease. Front Endocrinol (Lausanne). 9(405)2018.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Deng W, Zhang Y, Cai J, Zhang J, Liu X,
Yin J, Bai Z, Yao H and Zhang Z: lncRNA-ANRIL promotes gastric
cancer progression by enhancing NF-κB signaling. Exp Biol Med
(Maywood). 244:953–959. 2019.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Yu X, Zheng H, Chan MT and Wu WKK: BANCR:
A cancer-related long non-coding RNA. Am J Cancer Res. 7:1779–1787.
2017.PubMed/NCBI
|
|
98
|
Zhang ZX, Liu ZQ, Jiang B, Lu XY, Ning XF,
Yuan CT and Wang AL: BRAF activated non-coding RNA (BANCR)
promoting gastric cancer cells proliferation via regulation of
NF-κB1. Biochem Bioph Res Commun. 465:225–231. 2015.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Zhang JX, Chen ZH, Chen DL, Tian XP, Wang
CY, Zhou ZW, Gao Y, Xu Y, Chen C, Zheng ZS, et al:
LINC01410-miR-532-NCF2-NF-κB feedback loop promotes gastric cancer
angiogenesis and metastasis. Oncogene. 37:2660–2675.
2018.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Zheng J, Zhang H, Ma R, Liu H and Gao P:
Long non-coding RNA KRT19P3 suppresses proliferation and metastasis
through COPS7A-mediated NF-κB pathway in gastric cancer. Oncogene.
38:7073–7088. 2019.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Mattiuzzi C and Lippi G: Current cancer
epidemiology. J Epidemiol Glob Health. 9:217–222. 2019.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Marta ZN, Agnieszka W, Jacek P, Jeleń A,
Adrian K, Dagmara SK, Sałagacka-Kubiak A and Balcerczak E: NFKB2
gene expression in patients with peptic ulcer diseases and gastric
cancer. Mol Biol Rep. 47:2015–2021. 2020.PubMed/NCBI View Article : Google Scholar
|