Introduction
Hyperoxia-induced acute lung injury (HALI) is a type
of iatrogenic pulmonary dysfunction caused by long-term exposure to
high concentrations of oxygen, which is commonly used in the
treatment of refractory hypoxemia (1). The prolonged inhalation of oxygen at
high concentrations can stimulate the production of reactive oxygen
species (ROS), which may lead to an imbalance in the
oxidative-antioxidant system and induce a severe inflammatory
response, causing HALI, which is characterized by damage to the
alveolar epithelium, increased microvascular permeability,
inflammatory cell infiltration, and pulmonary edema. However, it is
believed that HALI cannot be prevented simply by decreasing the
concentration of inhaled oxygen, particularly because the
inhalation of oxygen at high concentrations is necessary to prevent
hypoxemia and to lower the incidence of incision infection,
postoperative nausea, and vomiting, among other conditions
(2,3). Anesthesiologists require elderly
patients with poor oxygen reserves, obese patients, and patients
with airway constriction to inhale oxygen at high concentrations to
increase the arterial partial pressure of oxygen and patient
tolerance to apnea and hypopnea syndrome as well as to ensure
sufficient intubation time for preventing intraoperative hypoxia in
critical situations (such as before induction and during recovery)
(4,5). Therefore, the decrease in oxygen
concentration for preventing hyperoxia-induced lung injury is not
feasible in all patients. However, the contradiction between oxygen
demand and lung oxygen toxicity remains unresolved.
Anti-inflammatory drugs commonly used in clinical
practice remain ineffective for the prevention and treatment of
HALI. Etomidate is an ultrashort-acting, hypnotic, non-barbiturate
intravenous anesthetic agent. It offers advantages such as high
safety, no significant accumulation, and mild effects on
respiratory functions and the circulatory system. Etomidate has
been reported to exert anti-stress and anti-inflammatory effects,
and has also been shown to improve the symptoms of
ischemia-reperfusion injury (6-11).
Nuclear factor-erythroid 2-related factor 2 (Nrf2)
is a transcription factor with antioxidant and anti-inflammatory
properties that plays a key role in maintaining the endogenous
redox balance and controlling the gene networks involved in cell
cycle homeostasis, metabolism, cell protection, immunity and
tumorigenesis (12). At present,
Nrf2 is known as the primary regulator of antioxidative responses.
It induces the expression of multiple genes, most of which encode
antioxidant/detoxification enzymes (13) and plays a key role by inducing both
antioxidants and phase II enzymes [such as NAD(P)H: Quinone
oxidoreductase 1, Glutathione S-transferases (GSTs)], including
heme oxygenase 1 (HO-1) (14). HO-1
is induced at high levels under stress and inflammation. HO-1
performs physiological functions in cell protection, inflammatory
response regulation, antioxidant function, regulation of cell
proliferation, angiogenesis and cardiovascular and pulmonary
homeostasis. HO-1 induction is an adaptive defense mechanism that
protects cells and tissues in various pathological states, such as
in cardiovascular and pulmonary diseases (15). The overall beneficial role of HO-1
is primarily achieved via the HO system. Studies have shown that
H2 supplementation can improve hyperoxic lung injury,
improve blood oxygenation, and alleviate oxidative stress by
regulating the Nrf2 pathway and inducing HO-1(16). Therefore, Nrf2/HO-1 signaling plays
a critical role in the regulation of oxidative stress.
In the present study, it was hypothesized that
etomidate could decrease HALI through its anti-stress and
anti-inflammatory action and its potential effects on the
anti-inflammatory Nrf2/HO-1 pathway were explored.
Materials and methods
Animals
Fifty male specific pathogen-free mice C57BL/6 mice
(6-8 weeks old; weighing 23-25 g) were provided by the Institute of
Experimental Animal Research, Chinese Academy of Medical Sciences.
Before the experiment, the mice were provided access to sufficient
food and water, housed at a relative humidity of 40-60% and
temperature of 22-25˚C, with a 12-h day/12-h night cycle. The
experimental protocol was approved by the Laboratory Animal Ethics
Committee of Shanxi Provincial People's Hospital (approval no.
2019-001-05) and was in accordance with the Guide for the Care and
Use of Laboratory Animals published by the National Research
Council (17).
Experimental instruments
The following instruments were used: Optical
microscope (Olympus Corporation); ChampGel 5000 gel imager (Beijing
Sage Venture Technology Co.); ABI Prism®7500
fluorescence quantitative PCR instrument (Applied Biosystems;
Thermo Fisher Scientific, Inc.); Varioskan LUX multifunctional
enzyme labeling instrument (Thermo Fisher Scientific, Inc.); and
SMART Spectro Spectrophotometer (LaMotte).
Drugs and reagents
The following drugs and reagents were procured for
the experiments: Etomidate (Jiangsu Enhua Pharmaceutical Co.); 0.9%
sodium chloride injection (Zhejiang DuBang Pharmaceutical Co.);
TRIzol (Takara Bio Inc.); complete protein extraction kit (Beijing
Solarbio Science & Technology Co., Ltd.); anti-Nrf2 protein
antibody (Santa Cruz, USA); anti-HO-1 protein antibody (Santa Cruz,
USA); anti-β-actin antibody (Santa Cruz Biotechnology, Inc.);
PRIME-SCRIPT RT-PCR kit (Takara Bio Inc.); SYBR PreMix Ex kits
(Takara Bio Inc.); tumor necrosis factor-α (TNF-α), interleukin
(IL)-6, IL-1β and IL-10 ELISA kits (Shanghai Westang Biotechnology
Co., Ltd.); myeloperoxidase (MPO), superoxide dismutase (SOD),
catalase (CAT) and malondialdehyde (MDA) kits (Nanjing Jiancheng
Bioengineering Institute); and anti-mouse IgG antibody (Cell
Signaling Technology, Inc.).
Grouping of experimental animals and
preparation of model
Fifty C57BL/6 mice were randomly divided into five
groups (ten mice in each group): Blank control group (CG), model
group (MG), high oxygen exposure + low etomidate dose group (ELG,
0.3 mg·kg-1), high oxygen exposure + moderate etomidate
dose group (EMG, 3.0 mg·kg-1), and high oxygen exposure
+ high etomidate dose group (EHG, 10 mg·kg-1). The CG
mice were exposed to room air, whereas mice in the other groups
were used as the HALI model. The mice were housed in an airtight
Plexiglas chamber, and 100% oxygen was blown continuously into the
chamber at a flow rate of 4.5-5 l/min. The appropriate dose of
etomidate was calculated according to the body weight of each
mouse. According to a relevant study (18), etomide was mixed with sterile normal
saline to prepare a 10 ml·kg-1 solution. The ELG, EMG,
and EHG mice were intraperitoneally injected with the corresponding
dose of etomidate once a day, whereas the CG and MG mice were
injected with normal saline (10 ml·kg-1) once daily for
3 consecutive days. 24 h after the last injection, the mice were
euthanized by intraperitoneal injection of pentobarbital sodium
(120 mg·kg-1 i.p.), following which the lung tissues
were harvested and the bronchoalveolar lavage fluid (BALF) was
collected.
Pulmonary histopathological
examination
The upper lobe of the right lung was cut, immersed
in 10% formalin, and embedded in paraffin. Routine
hematoxylin-eosin (HE) stained sections (4-5 µm) were prepared at
room temperature for ~120-160 min (the duration was adjusted
according to the staining results), and histopathological changes
in the lung tissues were observed using light microscopy
(magnification, x400) to determine the lung injury score (LIS)
(19) using the following criteria:
0 (no changes in the alveolar area), 1 (diffuse inflammatory cell
infiltration, predominant neutrophil infiltration, no alveolar wall
thickening); 2 (diffuse inflammatory cell, neutrophil and monocyte
infiltration, and mild alveolar wall thickening), 3 (2-3-fold
increase in alveolar wall thickness), 4 (alveolar wall thickening
with 25% lung consolidation), and 5 (alveolar wall thickening with
>50% lung consolidation).
Determination of the lung wet-to-dry
(W/D) ratio
The degree of pulmonary edema was assessed using the
W/D ratio, with the wet weight (W) obtained by weighing the lung
tissue immediately after isolation, and the dry weight (D) obtained
by weighing the lung tissue after dehydration for 48 h at 80˚C.
Measurement of MDA, MPO, SOD and CAT
levels in lung tissues
Based on the ratio of the weight of the lung tissue
(g) and the volume of 0.9% normal saline (ml) (=1:9), the lung
tissue in the glass homogenizer was manually homogenized to obtain
a 10% homogenate. The homogenate was centrifuged at 4˚C for 10 min
at 16,000 x g, and the supernatant obtained after centrifugation
was used as the sample. The MDA levels were measured using
thiobarbituric acid colorimetry (Malondialdehyde assay kit, cat.
no. A003-1-2), and the absorbance of MDA was measured at 532 nm
wavelength. The activities of MPO, SOD, and CAT were measured using
a colorimetric assay, the xanthine oxidase method, and the UV
absorption method, respectively. The activities of CAT, SOD, and
MPO were measured using Catalase assay kit (cat. no. A007-2-1), SOD
assay kit (cat. no. A001-1-2), and MPO assay kit (cat. no.
A044-1-1), respectively, and the absorbance values were measured at
wavelengths of 405, 550 and 460 nm, respectively.
Expression of Nrf2 and HO-1 mRNA in
lung tissues
Total RNA was extracted from the lung tissues using
TRIzol® (Thermo Fisher Scientific, Inc.) according to
the instructions of the manufacturer. After the RNA concentration
was determined spectrophotometrically, 1 µg RNA was used to
synthesize cDNA using the PRIME-SCRIPT RT-PCR kit [at 37˚C for 15
min (reverse transcription reaction), 1 cycle; at 85 C for 5 sec
(enzyme inactivation reaction), 1 cycle; maintaining at 4˚C, 1
cycle]. Quantitative polymerase chain reaction was performed using
the SYBR PreMix Ex kit and an ABI Prism®7500 fluorescent
quantitative PCR instrument. Thermocycling conditions: 50˚C for 2
min, 95˚C for 10 min, then 95˚C for 15 sec, 60˚C for 1 min, 40
cycles. The primer sequences used were as follows: Nrf2 forward,
5'-GTCAGCGACGGAAAGAGTA-3'; Nrf2 reverse, 5'-ACCTGGGAGTAGTTGGCA-3';
HO-1 forward, 5'-CGCAACAAGCAGAACCCA-3'; HO-1 reverse,
5'-GCGTGCAAGGGATGATTTCC-3'; β-actin forward,
5'-GGGAAATCGTGCGTGACATCAAAG-3'; and β-actin reverse,
5'-AACCGCTCGTTGCCAATAGT-3'. The expression of the relevant genes
was quantified using the 2-ΔΔCq method, using β-actin as
the internal reference (20).
Measurement of Nrf2 and HO-1 protein
expression levels in lung tissues
Total protein was extracted from the lung tissue
(T-PER Tissue Protein Extraction Reagent; cat. no. 78510; Thermo
Fisher Scientific, Inc.), and the Bradford method was used to
quantify total protein. Protein samples were separated by
electrophoresis in 10% SDS-PAGE gels (10 µg protein loaded per
lane) and transferred to polyvinylidene difluoride membranes. The
membranes were blocked by treating with 5% skimmed milk for 1 h at
room temperature and probed with anti-Nrf2 (1:500; cat. no.
Sc365949), anti-HO-1 (1:1,000; cat. no. Sc390991), and anti-β-actin
(1:1,000; cat. no. Sc81178) antibodies, using Helicobacter
pylori-coupled anti-mouse IgG (1:1,000; cat. no. 7076S) as the
secondary antibody. Images were recorded using a gel imager and
analyzed using the AlphaImage software v1.4.0 (GraphPad Software,
Inc.).
Detection of inflammatory factors
After the mice were sacrificed, normal saline was
injected into the tracheal tube for bronchoalveolar lavage. After
repeated lavage, BALF was collected in a centrifuge tube,
centrifuged at 110 x g for 10 min at 4˚C, and the supernatant was
collected and stored at -70˚C. The concentrations of TNF-α, IL-1β,
IL-6 and IL-10 in the BALF were determined using ELISA. Absorbance
was measured at 450 nm, and the concentrations of TNF-α (cat. no.
F26260), IL-1β (cat. no. F85180), IL-6 (cat. no. F10830) and IL-10
(cat. no. F10870) were calculated by drawing a standard curve based
on the absorbance value of the standard.
Statistical analysis
Data are expressed as mean ± SD. One-way analysis of
variance (ANOVA) was performed using SPSS software (version 22.0;
IBM Corp.), followed by Bonferroni post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
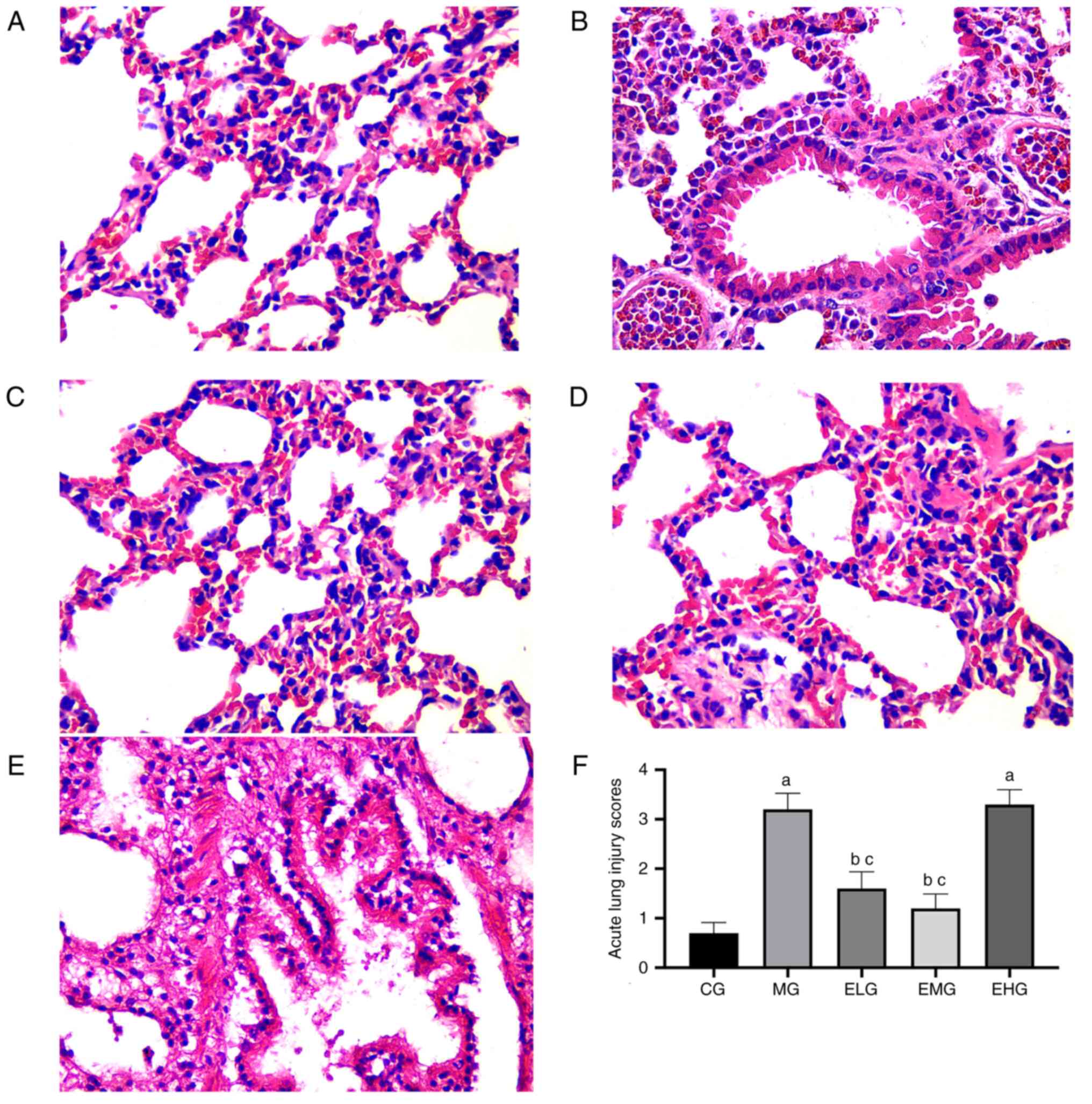
Lung histopathological changes and LIS
values
HE staining showed that the CG mice presented with
normal lung tissue structure, with no pathological changes
(Fig. 1A). Compared with the CG
mice, noticeable alveolar hemorrhage, edema, alveolar wall
thickening, inflammatory cell infiltration, and hyaline membrane
formation were observed in the MG mice after 72 h (Fig. 1B). Following etomidate
administration at low and moderate doses during the continuous
inhalation of oxygen at high concentrations, the ELG and EMG mice
exhibited mild alveolar pathological changes, intact lung tissue,
and fewer inflammatory cells (Fig.
1C and D), whereas the EHG
mice, which were administered with etomidate at high doses, showed
severe pathological changes, similar to that observed in the MG
mice (Fig. 1E).
The MG and EHG mice showed elevated LIS values
(P<0.05) compared with that in the CG mice. Compared with the MG
and EHG mice, the ELG and EMG mice showed lower LIS values
(P<0.05; Fig. 1F).
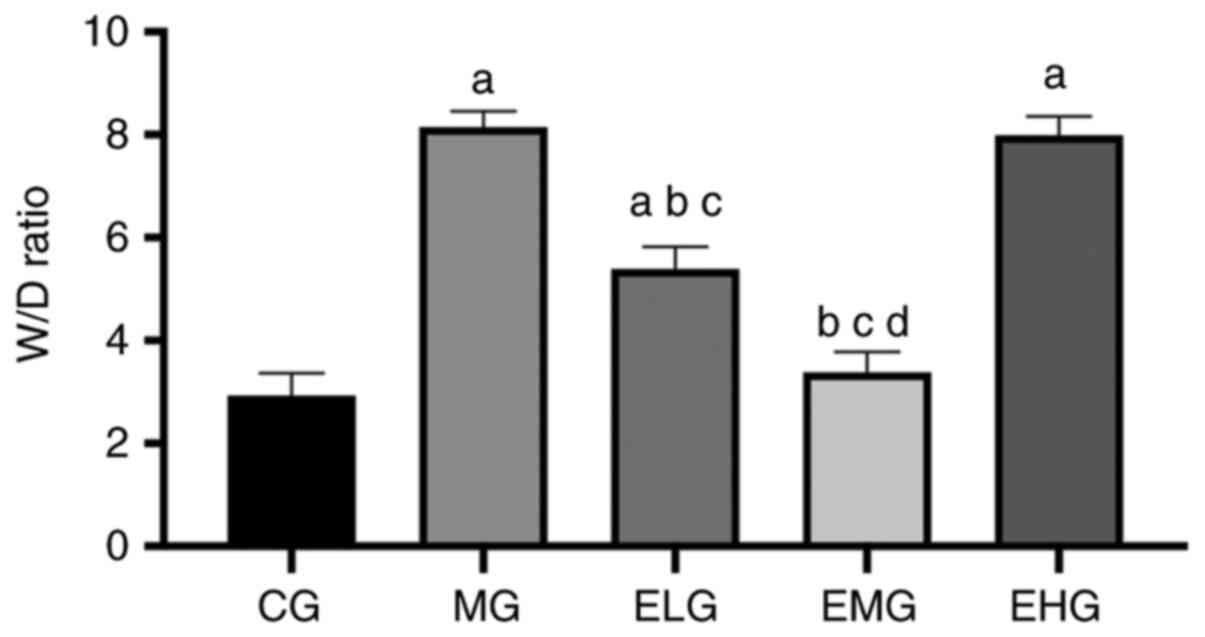
Low and moderate doses of etomidate
decreases the lung W/D ratio in mice with HALI
The MG, ELG and EHG mice exhibited an increased W/D
ratio (P<0.05) compared with that of the CG mice. The ELG and
EMG mice exhibited a decreased W/D ratio (P<0.05) compared with
that of the MG and EHG mice. The EMG mice showed a lower W/D ratio
compared with that of the ELG mice (P<0.05; Fig. 2).
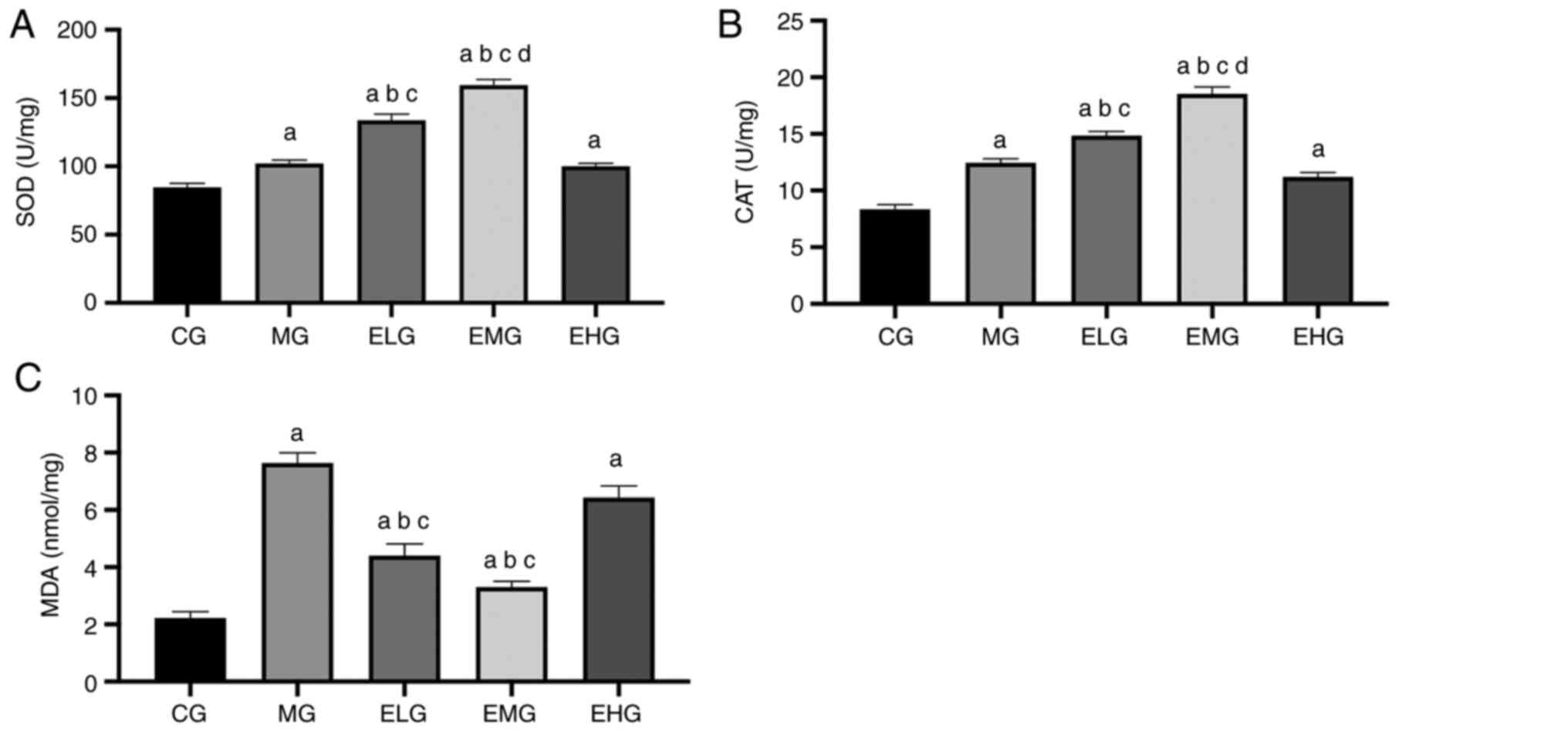
Effects of etomidate on the oxidative
stress response
The MDA content was higher and the SOD and CAT
activities were also enhanced in MG, ELG, EMG and EHG mice compared
with that of the CG mice (P<0.05). The MDA content was lower and
the SOD and CAT activities were higher in the ELG and EMG mice
compared with that of the MG and EHG mice (P<0.05). The EMG mice
showed higher CAT and SOD activities compared with that of the ELG
mice (P<0.05; Fig. 3, Table I).
 | Table IQuantitative analysis of CAT, MDA,
SOD and MPO in different groups. |
Table I
Quantitative analysis of CAT, MDA,
SOD and MPO in different groups.
| Group | CAT (U/mg) | MDA (nmol/mg) | SOD (U/mg) | MPO (U/g) |
|---|
| CG | 8.04±0.41 | 2.38±0.29 | 82.68±7.67 | 2.52±0.37 |
| MG |
12.91±0.50a |
7.69±0.40a |
103.44±8.01a |
12.13±.045a |
| ELG |
14.87±0.54a-c |
4.32±0.47a-c |
132.56±10.91a-c |
6.48±0.33a-c |
| EMG |
17.72±0.59a-d |
3.11±0.30a-c |
150.83±11.04a-d |
5.79±.039a-c |
| EHG |
10.05±0.46a |
6.73±0.42a |
104.25±9.82a |
11.08±0.50a |
Low and moderate etomidate doses
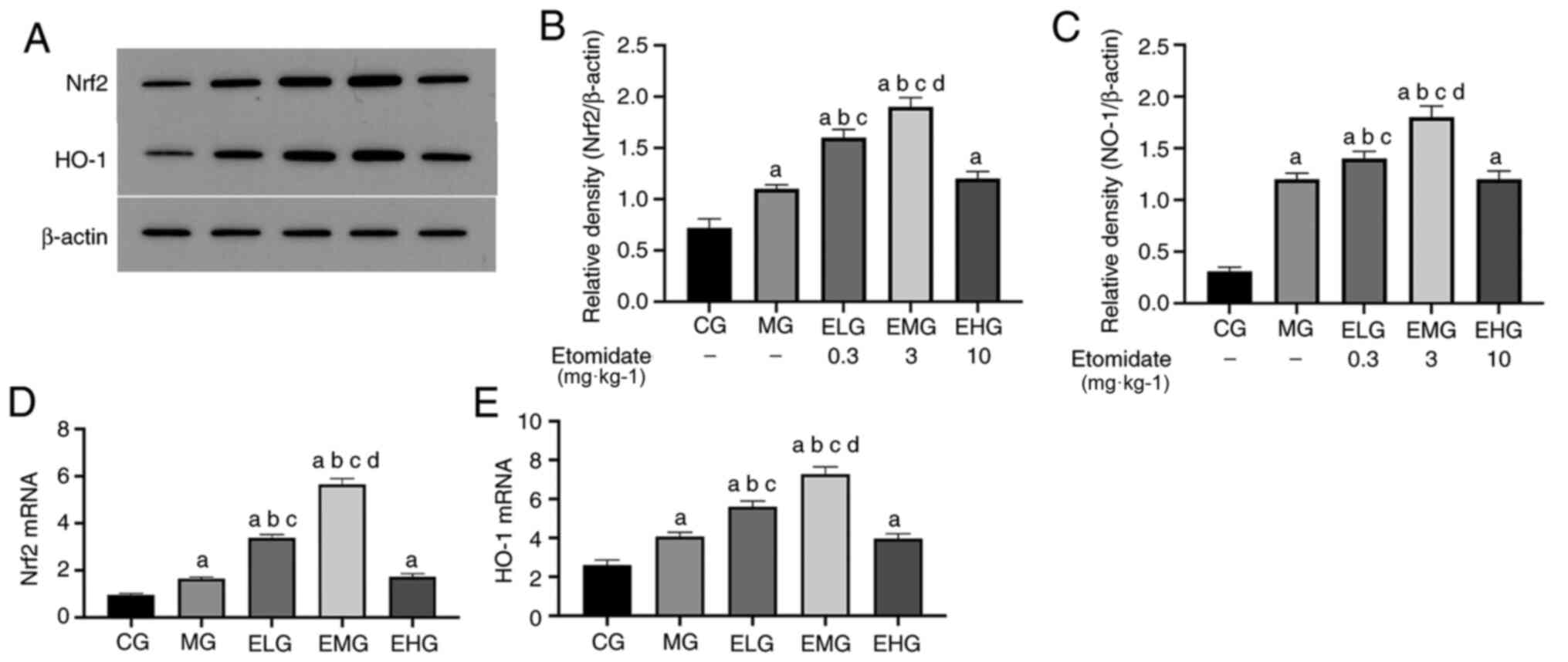
upregulates Nrf2 and HO-1 mRNA levels
The mRNA expression levels of Nrf2 and HO-1 were
elevated (P<0.05) in MG, ELG, EMG and EHG mice compared with
that of the CG mice. The mRNA expression levels of Nrf2 and HO-1
were elevated (P<0.05) in the ELG and EMG mice compared with
that of the MG and EHG mice. The mRNA expression of Nrf2 and HO-1
was elevated in the EMG mice compared with that of the ELG mice
(P<0.05; Fig. 4D and E; Table
II).
| Table IIRelative expression levels of Nrf2
and HO-1 mRNA in different groups. |
Table II
Relative expression levels of Nrf2
and HO-1 mRNA in different groups.
| Group | Nrf2 | HO-1 |
|---|
| CG | 0.823±0.016 | 2.621±0.040 |
| MG |
1.707±0.029a |
4.086±0.031a |
| ELG |
3.375±0.039a-c |
5.472±0.042a-c |
| EMG |
5.436±0.052a-d |
7.391±0.056a-d |
| EHG |
1.612±0.044a |
4.003±0.039a |
Etomidate at low to moderate doses
increases Nrf2 and HO-1 protein levels
Western blotting results revealed that the protein
expression levels of Nrf2 and HO-1 were elevated (P<0.05) in MG,
ELG, EMG and EHG mice compared with that of CG mice. Furthermore,
the protein expression levels of Nrf2 and HO-l were significantly
elevated in ELG and EMG mice, with the levels in EMG mice
comparable to those of the MG and ELG mice, respectively
(P<0.05; Fig. 4A-C).
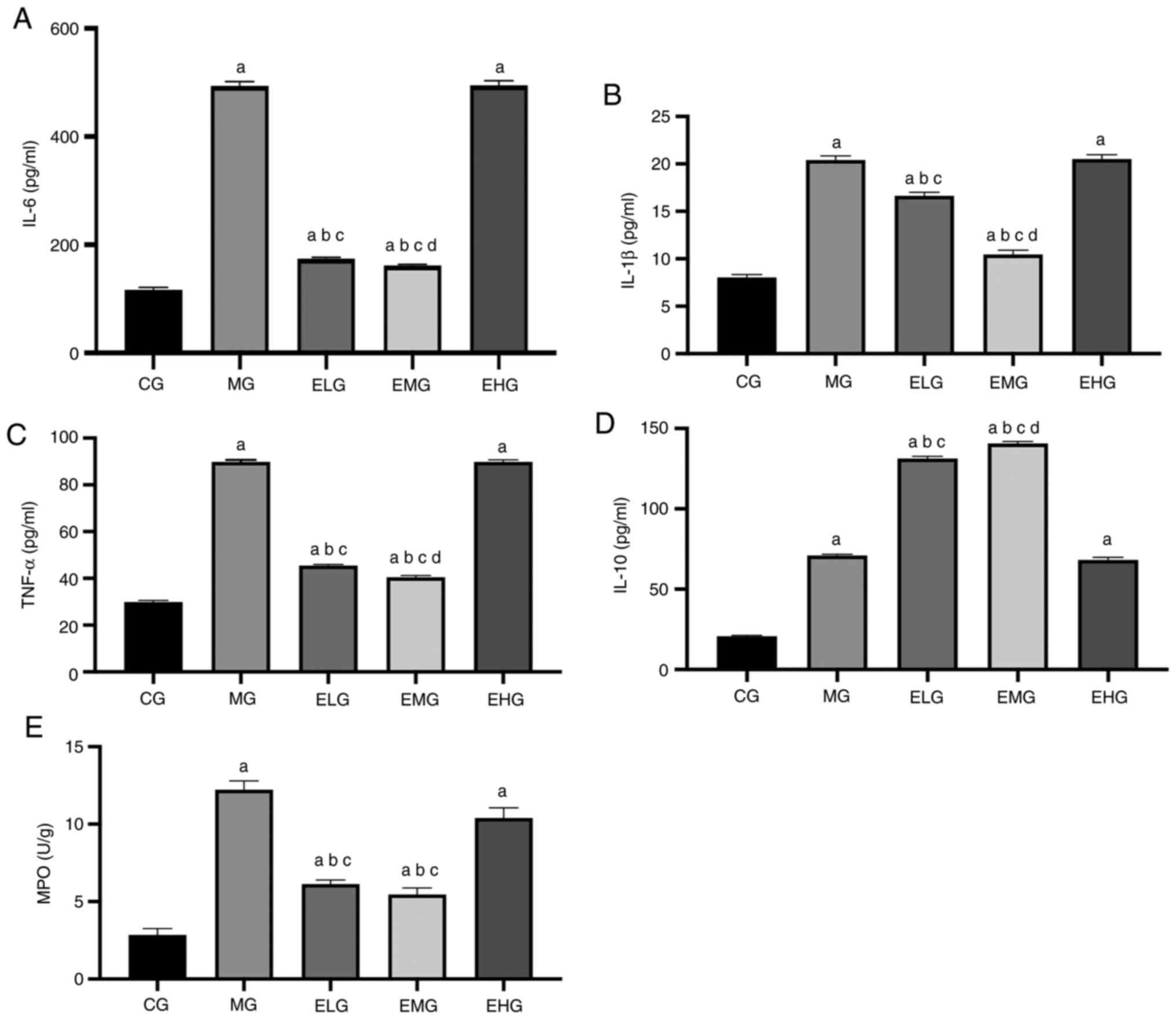
Effects of etomidate on the
inflammatory response
The levels of TNF-α, IL-6, IL-1β, IL-10 and MPO were
higher in the MG, ELG, EMG and EHG mice compared with that of the
CG mice (P<0.05). TNF-α, IL-6, IL-1β and MPO levels were lower
in the ELG and EMG mice, and the levels of the anti-inflammatory
factor IL-10 were lower in MG and EHG mice compared with that of
the ELG and EMG mice (P<0.05). Compared with the ELG mice, the
EMG mice showed lower TNF-α, IL-6, and IL-1β levels and higher
IL-10 levels (P<0.05; Fig.
5).
Discussion
Among the clinical anesthetics available, etomidate
is frequently administered to critically ill patients owing to its
reliable sedative efficacy and stable hemodynamic effects. Besides,
etomidate seems to exert organo-protective effects by decreasing
oxidative stress. A study has shown that etomidate alleviated
rabbit spinal cord ischemia-reperfusion injury caused by aortic
occlusion by decreasing oxidative stress (21). Zhao et al (9) found that etomidate can protect retinal
ganglion cells in adult rats by activating the antioxidative stress
response, significantly decreasing the levels of nitric oxide and
MDA in the retina, and increasing the level of glutathione (GSH).
Another study showed that etomidate can alleviate the inflammatory
response and oxidative stress in rats with myocardial
ischemia-reperfusion injury, characterized by the increase in SOD
and GSH levels and the decrease in MDA levels in myocardial tissue,
as well as the decrease in iNOS levels and increase in IL-10 levels
in serum and myocardial tissue (7).
In clinical studies, the application of etomidate to maintain
sedation during lower limb fracture surgery helped effectively
maintain the serum SOD activity after the fracture was complicated
with ischemia-reperfusion injury, inhibited the release of
inflammatory factors, and decreased the incidence of postoperative
complications caused by anesthesia (6). In the present study, a HALI mouse
model was established, and etomidate was administered at different
concentrations (0.3, 3 and 10 mg·kg-1) to investigate
the potential lung-protective effects. Since the unit body surface
area of mice is approximately nine times that of humans, the usual
clinical induction dose of etomidate (0.3 mg·kg-1) was
converted to 2.7 mg·kg-1 for mice. As indicated in
available literature, etomidate is administered to mice at doses
ranging from 0.3 to 30 mg·kg-1 (22-24).
Studies have shown that the sleep duration in mice injected with
etomidate is dose-dependent but can last for 6-10 min at 0.3
mg·kg-1, and recovery is rapid and complete without any
signs of hangover (25). Therefore,
0.3 mg·kg-1 was selected as the minimum dose. In some
studies, etomidate at a maximum dose of 10 mg·kg-1
occasionally caused death in mice (26). Therefore, it was decided to use 0.3,
3 and 10 mg·kg-1 of etomidate as the low, moderate, and
high doses based on evidences from a relevant study (22). In addition, in order to avoid the
impact of different liquid volume on lung tissue and according to a
previous study (18), the same drug
administration volume of 10 ml·kg-1 was adopted. The
results showed that 0.3 and 3 mg·kg-1 etomidate
attenuated the pathological changes caused by HALI, inhibited the
activation of pro-inflammatory factors, increased the levels of
anti-inflammatory factors, inhibited lipid peroxidation of lung
tissue, and enhanced the activities of antioxidant enzymes.
Notably, 3 mg·kg-1 etomidate exerted a stronger
protective effect. Additionally, etomidate upregulated the
expression of Nrf2/HO-1 mRNA and promoted the synthesis and
secretion of Nrf2/HO-1 proteins. However, at the high dose (10
mg·kg-1), etomidate failed to attenuate lung tissue
injury. It is speculated that etomidate inhibits hyperoxia-induced
inflammation and oxidative stress in a dose-dependent manner in
HALI, which may be associated with the upregulation of the
Nrf2/HO-1 pathway. These findings support the rational clinical use
of etomidate.
Notably, the inflammatory cascade and oxidative
stress play central roles in the development of HALI (27). In mice, the prolonged inhalation of
oxygen at high concentrations leads to the large-scale recruitment
of inflammatory cells in the lungs, particularly the recruitment
and activation of neutrophils, promoting the release of
inflammatory factors and oxidative stress mediators (28-30),
which is a marker of neutrophil infiltration and activation in
tissues. While the monocyte-macrophage system releases various
pro-inflammatory factors in vivo, monocytes and B cells also
release IL-10 to inhibit the expression of multiple
pro-inflammatory cytokines, chemokines, and chemokine receptors,
and suppress the inflammatory response (31). In the present study, mice that
continuously inhaled oxygen at high concentrations for 72 h showed
elevated levels of TNF-α, IL-6 and IL-1β in the BALF, as well as
increased MPO levels in lung tissues. Although there is no clear
clinical method to treat HALI, some basic studies have pointed out
that dexmedetomidine, exogenous IL-10 and activated protein C can
all decrease HALI (32-35).
However, the new findings indicate that at low and moderate doses,
etomidate decreased the infiltration of neutrophils, lowered the
levels of MPO as well as pro-inflammatory factors, and promoted the
release of IL-10, thereby reversing the imbalance in the
inflammatory state and promoting the recovery of lung injury.
Reportedly, the alveolar epithelium, basement
membrane and capillary endothelial cells are destroyed in the
inflammatory state, which leads to the loss of alveolar-capillary
barrier integrity, increases the permeability of alveolar
capillaries and alveolar epithelial cells, and promotes the influx
of macromolecules such as albumin into the alveolar space, which
consequently leads to pulmonary edema, hyaline membrane formation
and alveolar wall thickening (36).
Furthermore, ROS generated by activated neutrophils and monocytes
during inflammation has been shown to affect biofilm stability
(37). Pulmonary edema is
positively associated with the inflammatory status. In the present
study, the degree of lung edema was evaluated by measuring the W/D
ratio, whereas the severity of lung histopathological changes was
quantified based on the LIS value. In HALI mice, the lung tissue
sections showed severe pathological changes, such as inflammatory
cell infiltration, alveolar congestion, and alveolar wall
thickening. Compared with that of the CG mice, the lung W/D ratio
was higher in both MG and EHG mice, which was consistent with the
trend observed for the LIS values. Conversely, at low and moderate
doses, etomidate effectively suppressed morphological damage to
lung tissues and decreased pulmonary edema, inflammatory
infiltrates and lung injury in mice. It was speculated that
etomidate may decelerate the process of lung injury by inhibiting
the infiltration of inflammatory cells into lung tissues and
stabilizing the permeability of the lung epithelial cell membrane
and microvascular membrane.
Oxidative stress also plays a critical role in ALI
pathology. ROS is a by-product of aerobic metabolism, and a
hyperoxic environment can induce ROS production (38). It should be noted that there is no
clear answer to the limit of oxygen concentration and duration of
lung damage. At present, it is clinically believed that when the
inhaled oxygen concentration is >60%, most of the nitrogen in
the alveoli is replaced by oxygen, and the oxygen in the alveoli
quickly diffuses into the blood. If the air in the corresponding
alveoli is not replenished in time due to poor respiratory tract or
other reasons, collapse may occur, causing atelectasis, and the
dose-dependent manner increases the patient's 7-day postoperative
respiratory complications and 30-day mortality (39). Notably, exogenous and endogenous
ROS-mediated oxidative stress is a key factor in initiating the
expression of inflammatory mediators and inducing cellular damage
(40). Under hyperoxic conditions,
the major sites of ROS production are the mitochondria and
nicotinamide adenine dinucleotide oxidase (41). A significant increase in ROS levels
can trigger an oxidative stress response that eventually causes
oxidative damage to cellular macromolecules such as DNA, lipids,
and proteins, leading to DNA double-strand breaks, chromosomal
alterations, and other functional changes (42). The resulting state of
oxidative/hyperoxic stress consequently activates the inflammatory
response in lung tissues (43,44).
The infiltration of neutrophils and monocytes into the pulmonary
circulation and lung interstitium can further increase the levels
of ROS in the lung, thereby aggravating cell injury (45,46).
In addition, ROS can also react with various unsaturated fatty
acids and cholesterol on the cell membrane, leading to oxidative
damage and apoptosis (47). As a
metabolite of ROS, the MDA levels reflect the extent of lipid
peroxidation, protein denaturation and impaired endothelial
integrity (48,49). ROS elimination is primarily achieved
through multiple antioxidant enzymes, including SOD and CAT, which
are important endogenous antioxidant enzymes that maintain the
dynamic balance between oxidation-reduction reactions (50). In order to assess the status of
oxidative stress in lung tissues, the levels of MDA and antioxidant
enzymes were measured in lung tissues. The HALI model showed
increased MDA and MPO levels. The ELG and EMG mice showed decreased
MDA levels and enhanced SOD and CAT activities compared with that
of the MG mice. This suggests that at low and moderate doses,
etomidate can inhibit lipid peroxidation in the lung tissue of HALI
mice and significantly promote the activities of antioxidant
enzymes, thereby decreasing lung injury. It is believed that the
therapeutic effect of etomidate on hyperoxia-induced lung injury
can be partly attributed to the maintenance of the oxidative and
antioxidative stress balance.
Nrf2 is the primary regulator of the antioxidative
response against hyperoxia-induced oxidative stress. Nrf2
stimulates the expression of multiple genes, most of which encode
antioxidant/detoxification enzymes (51), thereby regulating immune stress and
antioxidative and anti-inflammatory responses under pathological
conditions (52,53). The protective effect of Nrf2 on lung
injury induced by oxidative stress was confirmed in adult
Nrf2-knockout mice (12,54-56)
Under physiological conditions, Nrf2 exists in the cytoplasm as an
inactive complex of Kelch-like ECH-associated protein 1 (KEAP-1).
Under oxidative stress, Nrf2 is released from KEAP-1 and
translocates to the nucleus, following which it binds to specific
antioxidant response elements and initiates the transcription of
genes encoding proteins with cytoprotective properties, of which
HO-1 is the most prominent (57).
HO-1 is a cytoprotective enzyme with important
physiological roles in the anti-inflammatory response,
antioxidation, cell proliferation and angiogenesis regulation, and
can be induced by different stimuli associated with inflammation
and oxidative stress, including ROS, heme, cytokines, endotoxins
and heavy metals (58,59). Previous studies have shown that the
inhibition of HO-1 activity enhances the inflammatory response,
whereas the induction of HO-1 enzyme activity effectively
suppresses the production of inflammatory mediators (60,61).
It was observed that the levels of proteins associated with the
Nrf2/HO-1 pathway were mildly elevated in the lung tissue of the
HALI model, which was attributed to the activation of the Nrf2/HO-1
pathway in response to hyperoxic stimulation; during lung injury,
the production and utilization of Nrf2 and HO-1 are balanced, with
seemingly unchanged Nrf2 and HO-1 levels, which may be attributed
to self-protective mechanisms. However, this is insufficient to
prevent ALI development. Furthermore, following the administration
of etomidate at different doses, a significant increase in Nrf2 and
HO-1 mRNA expression was observed in the ELG and EMG mice compared
with that of the CG mice. The aforementioned finding, combined with
the western blotting results, suggested that etomidate may activate
the Nrf2/HO-1 pathway, upregulate Nrf2/HO-1 gene expression and
promote Nrf2/HO-1 protein synthesis and secretion. Studies have
shown that the activation of the Nrf2/HO-1 pathway can effectively
suppress inflammatory responses by inhibiting the expression of
pro-inflammatory mediators (13,14).
Activation of the Nrf2/HO-1 pathway exerts a protective effect on
the integrity of alveolar epithelial cells and microvascular
membranes, preventing the infiltration of inflammatory cells.
Therefore, the activation of the Nrf2/HO-1 pathway plays an
important role in the protective effects exerted by etomidate.
Based on the findings of the present study,
etomidate appears to exert an organo-protective effect,
particularly a lung-protective effect, in addition to its
anesthetic effect. Anesthesia remains a rather unexplored subject
thus far, and there is no clear and systematic conclusion on the
mechanism of action of general anesthetics. In addition to
anesthetic effects, there have been several breakthroughs in the
alternative properties of general anesthetics, such as organ
protection, anti-neuropsychiatric effects and antitumor effects.
Similarly, even etomidate, which has been used in clinical practice
for several years now, is still being assessed for its potential
organo-protective effects beyond its anesthetic effects. In fact,
several scholars have explored the additional effects of
anesthesia; for example, propofol has been found to prevent
cerebral ischemia-reperfusion injury (62), dexmedetomidine has been shown to
alleviate spinal cord injury (63),
and sevoflurane has been shown to exert anti-inflammatory effects
and decrease lung injury in patients (64). In the present study, the alternative
uses of etomidate in addition to its anesthetic effect was
investigated, and both the point of initiation and preliminary
findings of the present study were significant.; this may be one of
the organo-protective effects of etomidate.
However, the present study has the following
limitations. Firstly, in principle, the recognized drug that is
most effective and safest for the treatment of HALI should be
selected as the positive control. Unfortunately, there is no
definite, effective and clinically recognized drug for the
treatment of HALI at present, so the present study is consistent
with other associated studies (65,66)
without using a positive control. However, a positive control
should be selected as a reference for further studies in the
future. Secondly, the present findings revealed that the reversal
of lung injury induced by etomidate administration at low and
moderate doses (0.3 and 3 mg·kg-1) was not observed upon
administration at a high dose (10 mg·kg-1). Thus,
etomidate may only exert lung-protective effects within a certain
dose range. Further exploration of the optimal etomidate dose
required for lung-protective effects is intended in the future.
In summary, the present study showed that etomidate
can effectively decrease lung injury in a HALI model within a
specific dose range, which may involve the inhibition of
inflammatory responses and improvement in antioxidant capacity, and
the potential mechanism may involve the upregulation of the
Nrf2/HO-1 pathway.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The data that support the findings of this study are
available from Shanxi Provincial People's Hospital, Taiyuan,
Shanxi, China but restrictions apply to the availability of these
data, which were used under license for the current study, and so
are not publicly available. Data are however available from the
authors upon reasonable request and with permission of Shanxi
Provincial People's Hospital, Taiyuan, Shanxi, China.
Authors' contributions
LJ and HH confirm the authenticity of all the raw
data; LJ and HH designed experiments; HH, CW and JW carried out
experiments and analyzed the data. LJ wrote the manuscript, LJ and
HH revised the manuscript. All authors approved the final
manuscript.
Ethics approval and consent to
participate
The experimental protocol was approved by the
Laboratory Animal Ethics Committee of Shanxi Provincial People's
Hospital (approval no. 2019-001-05) and is in accordance with the
‘Guide for the Care and Use of Laboratory Animals’ published by the
National Institutes of Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nyp MF, Mabry SM, Navarro A, Menden H,
Perez RE, Sampath V and Ekekezie II: Lung epithelial-specific
TRIP-1 overexpression maintains epithelial integrity during
hyperoxia exposure. Physiol Rep. 6(e13585)2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hovaguimian F, Lysakowski C, Elia N and
Tramèr MR: Effect of intraoperative high inspired oxygen fraction
on surgical site infection, postoperative nausea and vomiting, and
pulmonary function: Systematic review and meta-analysis of
randomized controlled trials. Anesthesiology. 119:303–316.
2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Allegranzi B, Zayed B, Bischoff P, Kubilay
NZ, de Jonge S, de Vries F, Gomes SM, Gans S, Wallert ED, Wu X, et
al: New WHO recommendations on intraoperative and postoperative
measures for surgical site infection prevention: An evidence-based
global perspective. Lancet Infect Dis. 16:e288–e303.
2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Edmark L, Kostova-Aherdan K, Enlund M and
Hedenstierna G: Optimal oxygen concentration during induction of
general anesthesia. Anesthesiology. 98:28–33. 2003.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ladha K, Vidal Melo MF, McLean DJ,
Wanderer JP, Grabitz SD, Kurth T and Eikermann M: Intraoperative
protective mechanical ventilation and risk of postoperative
respiratory complications: Hospital based registry study. BMJ.
351(h3646)2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Li R, Fan L, Ma F, Cao Y, Gao J, Liu H and
Li Y: Effect of etomidate on the oxidative stress response and
levels of inflammatory factors from ischemia-reperfusion injury
after tibial fracture surgery. Exp Ther Med. 13:971–975.
2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xie D, Li M, Yu K, Lu H and Chen Y:
Etomidate alleviates cardiac dysfunction, fibrosis and oxidative
stress in rats with myocardial ischemic reperfusion injury. Ann
Transl Med. 8(1181)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Djuric M, Kostic S, Nikolic Turnic T,
Stankovic S, Skrbic R, Djuric DM, Zivkovic V, Jakovljevic V and
Stevanovic P: The comparison of the effects of ketamine and
etomidate on cardiodynamics, biochemical and oxidative stress
parameters in Wistar male rats. Mol Cell Biochem. 474:125–134.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhao X, Kuang F, You YY, Wu MM and You SW:
Etomidate affects the anti-oxidant pathway to protect retinal
ganglion cells after optic nerve transection. Neural Regen Res.
14:2020–2024. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Ates O, Yucel N, Cayli SR, Altinoz E,
Yologlu S, Kocak A, Cakir CO and Turkoz Y: Neuroprotective effect
of etomidate in the central nervous system of
streptozotocin-induced diabetic rats. Neurochem Res. 31:777–783.
2006.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Cayli SR, Ates O, Karadag N, Altinoz E,
Yucel N, Yologlu S, Kocak A and Cakir CO: Neuroprotective effect of
etomidate on functional recovery in experimental spinal cord
injury. Int J Dev Neurosci. 24:233–239. 2006.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Cho HY, Jedlicka AE, Reddy SP, Kensler TW,
Yamamoto M, Zhang LY and Kleeberger SR: Role of NRF2 in protection
against hyperoxic lung injury in mice. Am J Respir Cell Mol Biol.
26:175–182. 2002.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Loboda A, Damulewicz M, Pyza E, Jozkowicz
A and Dulak J: Role of Nrf2/HO-1 system in development, oxidative
stress response and diseases: An evolutionarily conserved
mechanism. Cell Mol Life Sci. 73:3221–3247. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chen Z, Zhong H, Wei J, Lin S, Zong Z,
Gong F, Huang X, Sun J, Li P, Lin H, et al: Inhibition of Nrf2/HO-1
signaling leads to increased activation of the NLRP3 inflammasome
in osteoarthritis. Arthritis Res Ther. 21(300)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Abraham NG and Kappas A: Pharmacological
and clinical aspects of heme oxygenase. Pharmacol Rev. 60:79–127.
2008.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ohta S: Molecular hydrogen as a preventive
and therapeutic medical gas: Initiation, development and potential
of hydrogen medicine. Pharmacol Ther. 144:1–11. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
National Research Council (US): Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals. Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press,Washington, DC, 2011.
|
|
18
|
Paris A, Hein L, Brede M, Brand PA, Scholz
J and Tonner PH: The anesthetic effects of etomidate:
Species-specific interaction with alpha 2-adrenoceptors. Anesth
Analg. 105:1644–1649. 2007.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Patel V, Dial K, Wu J, Gauthier AG, Wu W,
Lin M, Espey MG, Thomas DD, Ashby CR Jr and Mantell LL: Dietary
antioxidants significantly attenuate hyperoxia-induced acute
inflammatory lung injury by enhancing macrophage function via
reducing the accumulation of airway HMGB1. Int J Mol Sci.
21(977)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Navidshad B, Liang JB and Jahromi MF:
Correlation coefficients between different methods of expressing
bacterial quantification using real time PCR. Int J Mol Sci.
13:2119–2132. 2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yu Q, Zhou Q, Huang H, Wang Y, Tian S and
Duan D: Protective effect of etomidate on spinal cord
ischemia-reperfusion injury induced by aortic occlusion in rabbits.
Ann Vasc Surg. 24:225–232. 2010.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Nyman Y, Fredriksson A, Lönnqvist PA and
Viberg H: Etomidate exposure in early infant mice (P10) does not
induce apoptosis or affect behaviour. Acta Anaesthesiol Scand.
60:588–596. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Besnier E, Clavier T, Tonon MC, Selim J,
Lefevre-Scelles A, Morin F, Tamion F, Dureuil B, Castel H and
Compere V: Ketamine and etomidate down-regulate the
hypothalamic-pituitary-adrenal axis in an endotoxemic mouse model.
Anesthesiology. 127:347–354. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Benkwitz C, Liao M, Laster MJ, Sonner JM,
Eger EI II and Pearce RA: Determination of the EC50 amnesic
concentration of etomidate and its diffusion profile in brain
tissue: Implications for in vitro studies. Anesthesiology.
106:114–123. 2007.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Green CJ, Knight J, Precious S and Simpkin
S: Metomidate, etomidate and fentanyl as injectable anaesthetic
agents in mice. Lab Anim. 15:171–175. 1981.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Liao M, Sonner JM, Husain SS, Miller KW,
Jurd R, Rudolph U and Eger EI II: R (+) etomidate and the
photoactivable R (+) azietomidate have comparable anesthetic
activity in wild-type mice and comparably decreased activity in
mice with a N265M point mutation in the gamma-aminobutyric acid
receptor beta3 subunit. Anesth Analg. 101:131–135, table of
contents. 2005.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Kallet RH and Matthay MA: Hyperoxic acute
lung injury. Respir Care. 58:123–141. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yuan CB, Tian L, Yang B and Zhou HY:
Isoalantolactone protects LPS-induced acute lung injury through
Nrf2 activation. Microb Pathog. 123:213–218. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Rankin SM: The bone marrow: A site of
neutrophil clearance. J Leukoc Biol. 88:241–251. 2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ho YC, Lee SS, Yang ML, Huang-Liu R, Lee
CY, Li YC and Kuan YH: Zerumbone reduced the inflammatory response
of acute lung injury in endotoxin-treated mice via Akt-NFκB
pathway. Chem Biol Interact. 271:9–14. 2017.PubMed/NCBI View Article : Google Scholar :
Cannizzaro V,
Hantos Z, Sly PD and Zosky GR: Linking lung function and
inflammatory responses in ventilator-induced lung injury. Am J
Physiol Lung Cell Mol Physiol 300, L112-120, 2011.
|
|
31
|
Zhang Q, Wu D, Yang Y, Liu T and Liu H:
Dexmedetomidine alleviates hyperoxia-induced acute lung injury via
inhibiting NLRP3 inflammasome activation. Cell Physiol Biochem.
42:1907–1919. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Li HD, Zhang QX, Mao Z, Xu XJ, Li NY and
Zhang H: Exogenous interleukin-10 attenuates hyperoxia-induced
acute lung injury in mice. Exp Physiol. 100:331–340.
2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Li HD, Zhang ZR, Zhang QX, Qin ZC, He DM
and Chen JS: Treatment with exogenous hydrogen sulfide attenuates
hyperoxia-induced acute lung injury in mice. Eur J Appl Physiol.
113:1555–1563. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Husari AW, Khayat A, Awdeh H, Hatoum H,
Nasser M, Mroueh SM, Zaatari G, El-Sabban M and Dbaibo GS:
Activated protein C attenuates acute lung injury and apoptosis in a
hyperoxic animal model. Shock. 33:467–472. 2010.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Gong J, Liu H, Wu J, Qi H, Wu ZY, Shu HQ,
Li HB, Chen L, Wang YX, Li B, et al: Maresin 1 prevents
lipopolysaccharide-induced neutrophil survival and accelerates
resolution of acute lung injury. Shock. 44:371–380. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Meng X, Hu L and Li W: Baicalin
ameliorates lipopolysaccharide-induced acute lung injury in mice by
suppressing oxidative stress and inflammation via the activation of
the Nrf2-mediated HO-1 signaling pathway. Naunyn Schmiedebergs Arch
Pharmacol. 392:1421–1433. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Simonis FD, Juffermans NP and Schultz MJ:
Mechanical ventilation of the healthy lungs: Lessons learned from
recent trials. Curr Opin Crit Care. 27:55–59. 2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Staehr-Rye AK, Meyhoff CS, Scheffenbichler
FT, Vidal Melo MF, Gätke MR, Walsh JL, Ladha KS, Grabitz SD,
Nikolov MI, Kurth T, et al: High intraoperative inspiratory oxygen
fraction and risk of major respiratory complications. Br J Anaesth.
119:140–149. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Crapo JD: Oxidative stress as an initiator
of cytokine release and cell damage. Eur Respir J Suppl. 44:4s–6s.
2003.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhang X, Shan P, Sasidhar M, Chupp GL,
Flavell RA, Choi AM and Lee PJ: Reactive oxygen species and
extracellular signal-regulated kinase 1/2 mitogen-activated protein
kinase mediate hyperoxia-induced cell death in lung epithelium. Am
J Respir Cell Mol Biol. 28:305–315. 2003.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Schieber M and Chandel NS: ROS function in
redox signaling and oxidative stress. Curr Biol. 24:R453–R462.
2014.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Wiegman CH, Li F, Clarke CJ, Jazrawi E,
Kirkham P, Barnes PJ, Adcock IM and Chung KF: A comprehensive
analysis of oxidative stress in the ozone-induced lung inflammation
mouse model. Clin Sci (Lond). 126:425–440. 2014.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Villegas L, Stidham T and Nozik-Grayck E:
Oxidative stress and therapeutic development in lung diseases. J
Pulm Respir Med. 4(194)2014.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Gore A, Muralidhar M, Espey MG, Degenhardt
K and Mantell LL: Hyperoxia sensing: From molecular mechanisms to
significance in disease. J Immunotoxicol. 7:239–254.
2010.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Pagano A and Barazzone-Argiroffo C:
Alveolar cell death in hyperoxia-induced lung injury. Ann NY Acad
Sci. 1010:405–416. 2003.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Christ M, Luu B, Mejia JE, Moosbrugger I
and Bischoff P: Apoptosis induced by oxysterols in murine lymphoma
cells and in normal thymocytes. Immunology. 78:455–460.
1993.PubMed/NCBI
|
|
47
|
Yamamoto H, Yamamoto Y, Yamagami K, Kume
M, Kimoto S, Toyokuni S, Uchida K, Fukumoto M and Yamaoka Y:
Heat-shock preconditioning reduces oxidative protein denaturation
and ameliorates liver injury by carbon tetrachloride in rats. Res
Exp Med (Berl). 199:309–318. 2000.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhao X, Jin L, Shen N, Xu B, Zhang W, Zhu
H and Luo Z: Salidroside inhibits endogenous hydrogen peroxide
induced cytotoxicity of endothelial cells. Biol Pharm Bull.
36:1773–1778. 2013.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Sun Q, Wu Y, Zhao F and Wang J: Maresin 1
ameliorates lung ischemia/reperfusion injury by suppressing
oxidative stress via activation of the Nrf-2-Mediated HO-1
signaling pathway. Oxid Med Cell Longev.
2017(9634803)2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Cho HY, Jedlicka AE, Reddy SP, Zhang LY,
Kensler TW and Kleeberger SR: Linkage analysis of susceptibility to
hyperoxia. Nrf2 is a candidate gene. Am J Respir Cell Mol Biol.
26:42–51. 2002.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Kuo PC, Yu IC, Scofield BA, Brown DA,
Curfman ET, Paraiso HC, Chang FL and Yen JH:
3H-1,2-Dithiole-3-thione as a novel therapeutic agent for the
treatment of ischemic stroke through Nrf2 defense pathway. Brain
Behav Immun. 62:180–192. 2017.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Jung KA and Kwak MK: The Nrf2 system as a
potential target for the development of indirect antioxidants.
Molecules. 15:7266–7291. 2010.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Chan K and Kan YW: Nrf2 is essential for
protection against acute pulmonary injury in mice. Proc Natl Acad
Sci USA. 96:12731–12736. 1999.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Iizuka T, Ishii Y, Itoh K, Kiwamoto T,
Kimura T, Matsuno Y, Morishima Y, Hegab AE, Homma S, Nomura A, et
al: Nrf2-deficient mice are highly susceptible to cigarette
smoke-induced emphysema. Genes Cells. 10:1113–1125. 2005.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Cho HY and Kleeberger SR: Noblesse oblige:
NRF2 functions in the airways. Am J Respir Cell Mol Biol.
50:844–847. 2014.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Hirotsu Y, Katsuoka F, Funayama R,
Nagashima T, Nishida Y, Nakayama K, Engel JD and Yamamoto M:
Nrf2-MafG heterodimers contribute globally to antioxidant and
metabolic networks. Nucleic Acids Res. 40:10228–10239.
2012.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Murakami A, Tanaka T, Lee JY, Surh YJ, Kim
HW, Kawabata K, Nakamura Y, Jiwajinda S and Ohigashi H: Zerumbone,
a sesquiterpene in subtropical ginger, suppresses skin tumor
initiation and promotion stages in ICR mice. Int J Cancer.
110:481–490. 2004.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Willis D, Moore AR and Willoughby DA: Heme
oxygenase isoform expression in cellular and antibody-mediated
models of acute inflammation in the rat. J Pathol. 190:627–634.
2000.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Li QF, Zhu YS, Jiang H, Xu H and Sun Y:
Heme oxygenase-1 mediates the anti-inflammatory effect of
isoflurane preconditioning in LPS-stimulated macrophages. Acta
Pharmacol Sin. 30:228–234. 2009.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Amata E, Pittalà V, Marrazzo A, Parenti C,
Prezzavento O, Arena E, Nabavi SM and Salerno L: Role of the
Nrf2/HO-1 axis in bronchopulmonary dysplasia and hyperoxic lung
injuries. Clin Sci (Lond). 131:1701–1712. 2017.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Hausburg MA, Banton KL, Roman PE, Salgado
F, Baek P, Waxman MJ, Tanner A II, Yoder J and Bar-Or D: Effects of
propofol on ischemia-reperfusion and traumatic brain injury. J Crit
Care. 56:281–287. 2020.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Bell MT, Puskas F, Bennett DT, Herson PS,
Quillinan N, Fullerton DA and Reece TB: Dexmedetomidine, an α-2a
adrenergic agonist, promotes ischemic tolerance in a murine model
of spinal cord ischemia-reperfusion. J Thorac Cardiovasc Surg.
147:500–506. 2014.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Araújo MN, Santos CL, Samary CS, Heil LB,
Cavalcanti VC, Cruz FF, Felix NS, Silva JD, Morales MM, Pelosi P,
et al: Sevoflurane, compared with isoflurane, minimizes lung damage
in pulmonary but not in extrapulmonary acute respiratory distress
syndrome in rats. Anesth Analg. 125:491–498. 2017.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Zhang Y, Du H, Yu X and Zhu J: Fucoidan
attenuates hyperoxia-induced lung injury in newborn rats by
mediating lung fibroblasts differentiate into myofibroblasts. Ann
Transl Med. 8(1501)2020.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Tayman C, Cekmez F, Kafa IM, Canpolat FE,
Cetinkaya M, Tonbul A, Uysal S, Tunc T and Sarici SU: Protective
effects of nigella sativa oil in hyperoxia-induced lung injury.
Arch Bronconeumol. 49:15–21. 2013.PubMed/NCBI View Article : Google Scholar : (In Spanish).
|
|
66
|
Zhang Q, Wu D, Yang Y, Liu T and Liu H:
Effects of dexmedetomidine on the protection of hyperoxia-induced
lung injury in newborn rats. Int J Clin Exp Pathol. 8:6466–6473.
2015.PubMed/NCBI
|