Introduction
The most effective treatment for acute myocardial
infarction (AMI) is prompt reperfusion of the ischemic myocardium.
However, the restoration of blood flow to the ischemic myocardium
may result in arrhythmias, myocardial stunning, microvascular
obstruction and lethal myocardial reperfusion injury (1), known as myocardial
ischemia/reperfusion injury (I/R). Although several critical
factors that act to mediate the damaging effects of myocardial I/R
have been identified, there is as yet no effective therapeutic
strategy for the prevention of myocardial I/R injury in patients
(2).
MicroRNAs (miRs/miRNAs) are endogenous, small,
non-coding RNAs that serve crucial roles in the regulation of gene
expression via sequence-specific interaction with the
3'-untranslated region (3' UTR) of target messenger RNA (mRNA)
(3). miRNAs are highly conserved
and ubiquitously expressed in all species. There is evidence to
suggest that miRNAs regulate necrotic, apoptotic and autophagic
cardiomyocyte death by altering key signaling in myocardial I/R
injury (4,5). For example, miR-494 targets pro- and
antiapoptotic proteins to provide cardioprotective effects against
I/R-induced injury via activation of the Akt pathway (6). miR-145 is highly expressed in the
vasculature and its role in the cardiovascular system has been
extensively investigated (7-9).
A few studies have explored the role of miR-145-5p in myocardial
I/R (10-12).
However, the results of these studies were inconsistent and the
role of miR-145-5p in myocardial I/R remains elusive.
Rho-kinases (Rho-associated coiled-coil-containing
kinases; ROCK) have important roles in numerous cellular functions,
including contraction, motility, proliferation and apoptosis
(13). Additionally, these kinases
have been demonstrated to serve a role in the pathogenesis of
vasospasm, arteriosclerosis, heart failure, hypertension, pulmonary
hypertension and myocardial I/R injury (14-19).
There are two isoforms of ROCK in humans and mice: ROCK1 and
ROCK2(20). Although ROCK1 and
ROCK2 are expressed in cardiomyocytes, there are some differences
in their functions. A number of studies have indicated that ROCK1
may be a key molecule in the mediation of apoptotic signaling in
cardiomyocytes (21-24).
The present study established a
hypoxia/reoxygenation (H/R) H9c2 cell model to investigate the
effects of miR-145-5p on myocardial I/R injury. The effect of H/R
on the expression of miR-145-5p in H9c2 cardiomyocytes was
investigated. In addition, the effect of overexpressing miR-145-5p
on H/R-induced cardiomyocyte apoptosis and whether the underlying
mechanism involves the targeting of ROCK1 were examined. The
findings of the present study may reveal novel information to
assist in the therapy of myocardial I/R injury.
Materials and methods
H/R cell model
H9c2 cardiomyocytes were purchased from the American
Type Culture Collection (cat. no. CRL-1446) and cultured in DMEM
(Hyclone; Cytiva) containing glucose (25 mmol/l), 10% fetal bovine
serum (FBS; GE Healthcare) and 1% penicillin/streptomycin at 37˚C
with 5% CO2. Prior to hypoxia, the culture medium was
changed to FBS-free DMEM. Hypoxia was induced by culturing the H9c2
cardiomyocytes in FBS-free DMEM without glucose in 95%
N2 and 5% CO2. Following hypoxia treatment
for 6 h, H9c2 cardiomyocytes were cultured in normoxic conditions
(with glucose; 5% CO2) for 6 h for reoxygenation. H9c2
cardiomyocytes cultured in normal conditions served as
controls.
Cell transfection
Transfection of the H9c2 cardiomyocytes was
performed prior to hypoxia treatment. miR-145-5p mimic
(miR10000851), miR mimic negative control (miR mimic-NC;
miR1N0000001-1-5), miR-145-5p inhibitor (miR20000851) and miR
inhibitor-NC (miR2N0000001-1-5) were purchased from Guangzhou
RiboBio Co., Ltd. and transfected into H9c2 cardiomyocytes using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
H9c2 cardiomyocytes were seeded in 6-well plates (5x105
cells/well) and transfected with 100 pmol miR-145-5p mimic,
miR-145-5p inhibitor or the respective controls in 500 µl
serum-free media at 37˚C for 24 h. After 24 h of transfection,
western blotting and reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) were performed to select the most
efficient mimic and inhibitor sequences. The sequences of the
miR-145-5p mimic, miR-145-5p inhibitor and controls were as
follows: miR-145-5p mimic, 5'-GUCCAGUUUUCCCAGGAAUCCCU-3';
miR-145-5p inhibitor, 5'-AGGGAUUCCUGGGAAAACUGGAC-3'; miR mimic-NC,
5'-UUCUCCGAACGUGUCACGUTT-3'; miR inhibitor-NC,
5'-CAGUACUUUUGUGUAGUACAA-3'. The ROCK1 sequence was sub-cloned into
a pcDNA3.1 vector (Invitrogen; Thermo Fisher Scientific, Inc.) to
generate a ROCK1-expression vector (1 µg/µl) and transfected into
the H9c2 cardiomyocytes using Lipofectamine® 2000 at
37˚C for 48 h. Cells transfected with the empty pcDNA3.1 vector
were used as the negative control.
Extraction of total RNA and
RT-qPCR
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. The RNA
concentration and purity were quantified using a
NanoDrop™ ND-2000 spectrophotometer (Thermo Fisher
Scientific, Inc.). The final concentration of RNA was adjusted to
200 ng/µl in all samples. The specific stem-loop primer of
miR-145-5p was used in RT-qPCR and the U6 gene was used as an
endogenous control when quantifying miR-145-5p; β-actin served as
the endogenous control for ROCK1. RT of the RNA was conducted using
a RevertAid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. qPCR
was performed with Fast Start Universal SYBR Green I kit (Roche
Diagnostics). Fold-changes were calculated using the
2-ΔΔCq method (25) and
each data was performed in triplicate. The thermocycling conditions
were as follows: Initial denaturation at 95˚C for 30 sec, followed
by 40 cycles at 95˚C for 5 sec and 60˚C for 30 sec. The primer
sequences used for RT-qPCR are listed in Table I.
 | Table IPrimers used in quantitative PCR. |
Table I
Primers used in quantitative PCR.
| Primer name | Forward | Reverse |
|---|
| miR-145-5p |
ACACTCCAGCTGGGGTCCAGTTTTCCCAGGA |
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGT
TGAGAGGGATTC |
| ROCK1 |
GGAAACGCTCCGAGACACTG |
CTGTTCTCACTGGGATTTGCTG |
| β-actin |
TGCTATGTTGCCCTAGACTTCG |
GTTGGCATAGAGGTCTTTACGG |
| U6 |
CTCGCTTCGGCAGCACA |
AACGCTTCACGAATTTGCGT |
Western blotting
Cells were harvested and lysed with lysis buffer
(Beyotime Institute of Biotechnology). The protein concentration of
the cell lysate was determined using a BCA assay kit (Beyotime
Institute of Biotechnology) according to the manufacturer's
instructions. Protein samples (50 µg/lane) were subjected to 12%
SDS-PAGE and transferred to PVDF membranes (GE Healthcare). The
membranes were then blocked with 5% defatted milk in TBST (0.5%
Tween-20 in TBS) for at 37˚C 1 h and incubated with primary
antibodies at 4˚C overnight. The primary antibodies used were as
follows: ROCK1 (1:1,000; 158 kDa; cat. no. ab219587; Abcam),
pro-caspase-3 (1:1,000; 32 kDa; cat. no. ab90437; Abcam), cleaved
caspase-3 (1:1,000; 17 kDa; cat. no. ab49822; Abcam) and β-actin
(1:1,000; 42 kDa; cat. no. ab8227; Abcam). Subsequently, the
membranes were washed in TBST and then incubated with
HRP-conjugated secondary antibody (1:10,000; cat. no. ab205718;
Abcam) at room temperature for 2 h. Immunoreactive bands were
visualized with the SuperSignal West Pico enhanced chemiluminescent
substrate (Pierce; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Band intensities were quantified using
Quantity One software v4.6.6 (Bio-Rad Laboratories, Inc.).
Flow cytometric analysis
To detect cell apoptosis, the cardiomyocytes were
digested with 0.25% trypsin at 37˚C for 5 min, washed and
double-stained using an Annexin V-FITC/PI Apoptosis
Staining/Detection kit (Nanjing KeyGen Biotech Co., Ltd.). The
percentage of apoptotic cells was detected using a CytoFLEX flow
cytometer (Beckman Coulter, Inc.). Data were analyzed with FlowJo
software (10.0.7; TreeStar).
Dual luciferase reporter gene
assay
TargetScan online bioinformatics software
(http://www.targetscan.org/vert_72/)
was used to predict target genes that may be regulated by
miR-145-5p, and ROCK1 was identified as a target. H9c2
cardiomyocytes were transfected with the wild-type or mutant ROCK1
reporter plasmids (Shanghai GenePharma, Inc.) together with either
miR-145-5p mimic or miR mimic-NC using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.). The resulting
firefly and Renilla luciferase activities were measured 24 h
post-transfection using the Dual-Glo™ Luciferase Assay
System, according to the manufacturer's instructions (Promega
Corporation) The firefly luciferase activity was normalized to
Renilla luciferase activity.
Statistical analysis
All experiments were performed at least six times,
and representative data are shown. All descriptive variables are
expressed as the mean ± SD. The Shapiro-Wilk test was used for
testing normality. Comparisons between two groups were conducted
using an unpaired Student's t-test when the variables were normally
distributed, and Mann-Whitney U test when they were abnormally
distributed. Differences among multiple groups were analyzed using
one-way ANOVA followed by Tukey's post hoc tests. Statistical
analyses were performed using SPSS 20.0 (IBM Corp.). P<0.05 was
considered to indicate a statistically significant difference.
Results
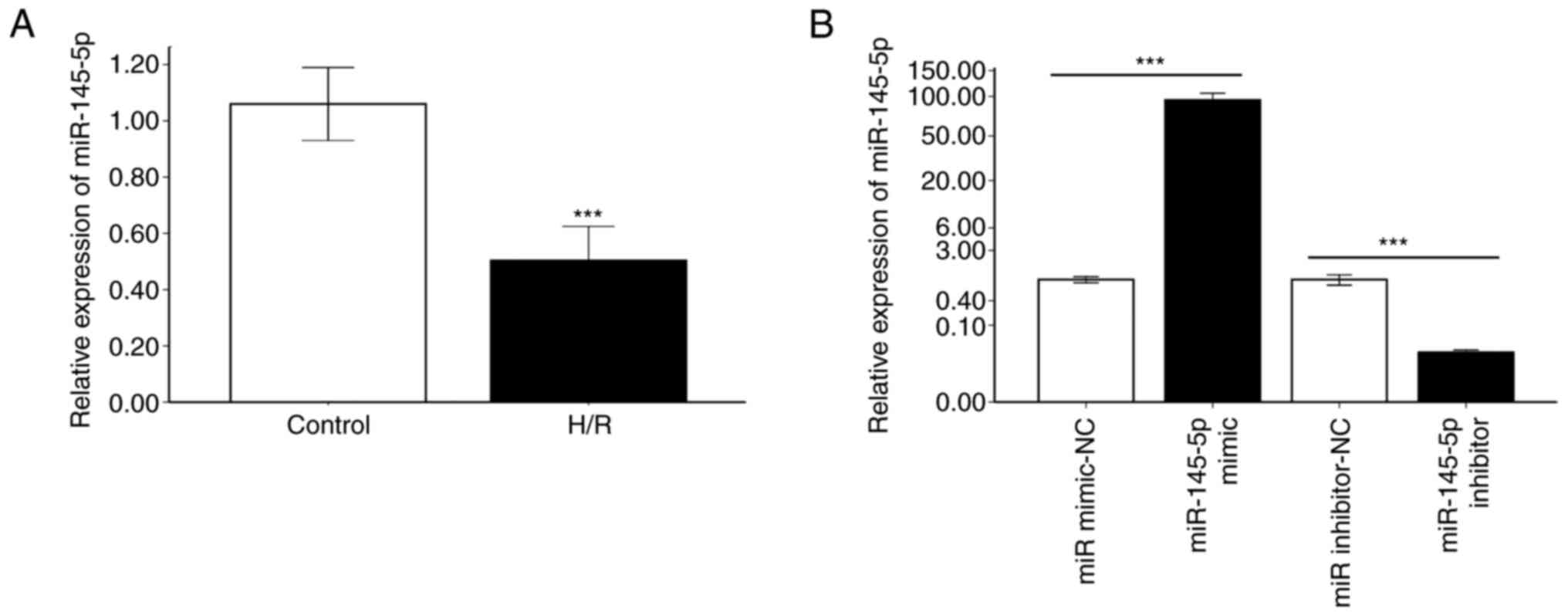
miR-145-5p expression is downregulated
in cardiomyocytes following H/R
To assess the changes of miR-145-5p expression in
cardiomyocytes subjected to H/R, RT-qPCR was performed on H9c2
cells. miR-145-5p expression in H9c2 cardiomyocytes after H/R was
significantly downregulated compared with that in cells cultured
under normal conditions (Fig. 1A;
P<0.001). miR-145-5p mimic and miR-145-5p inhibitor were then
used to transfect H9c2 cardiomyocytes. RT-qPCR analysis revealed
that miR-145-5p expression was significantly increased after
transfection with miR-145-5p mimic compared with miR mimic-NC,
indicating that the miR-145-5p mimic significantly induced
miR-145-5p expression in the H9c2 cardiomyocytes (Fig. 1B; P<0.001). In addition,
transfection with miR-145-5p inhibitor resulted in lower levels of
miR-145-5p expression compared with those in cells transfected with
the miR inhibitor NC (Fig. 1B;
P<0.001). These results confirm the successful transfection of
the H9c2 cells with miR-145-5p mimic and inhibitor.
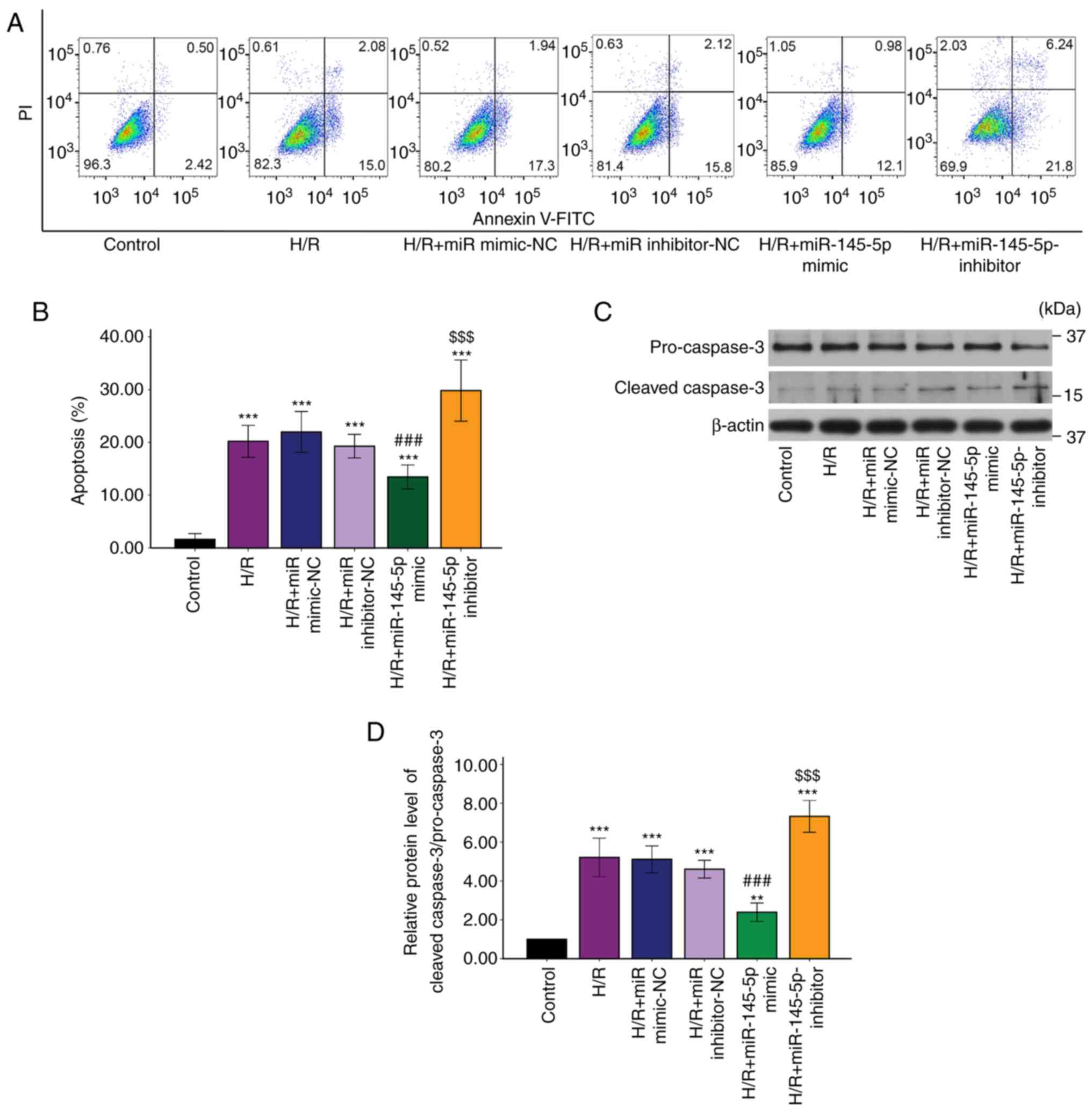
Upregulation of miR-145-5p reduces
H/R-induced cardiomyocyte apoptosis
The proportion of apoptotic cells following H/R
injury was determined by flow cytometry with Annexin V-FITC/PI
staining (Fig. 2A). The H9c2
cardiomyocytes in the H/R group exhibited a significant increase in
apoptosis compared with those in the control group (Fig. 2B; P<0.001). Transfection with
miR-145-5p mimic significantly decreased the H/R-induced apoptosis
rate compared with that in the H/R + miR mimic-NC group (Fig. 1B; P<0.01), and transfection with
miR-145-5p inhibitor significantly increased the apoptosis rate
compared with that in the H/R + miR inhibitor-NC group. The
activation of caspase-3 is a hallmark of apoptotic cell death
(26). Therefore, caspase-3
cleavage in the cells was investigated. Western blotting revealed
that transfection of the H9c2 cells with miR-145-5p mimic
significantly reduced the H/R-induced increase in caspase-3
cleavage compared with that in the H/R + miR mimic-NC group. In
addition, caspase-3 cleavage in the miR-145-5p inhibitor group was
significantly increased compared with that in the H/R + miR
inhibitor NC group (Fig. 2C and
D). The aforementioned findings
indicate that miR-145-5p suppressed H/R-induced cardiomyocyte
apoptosis.
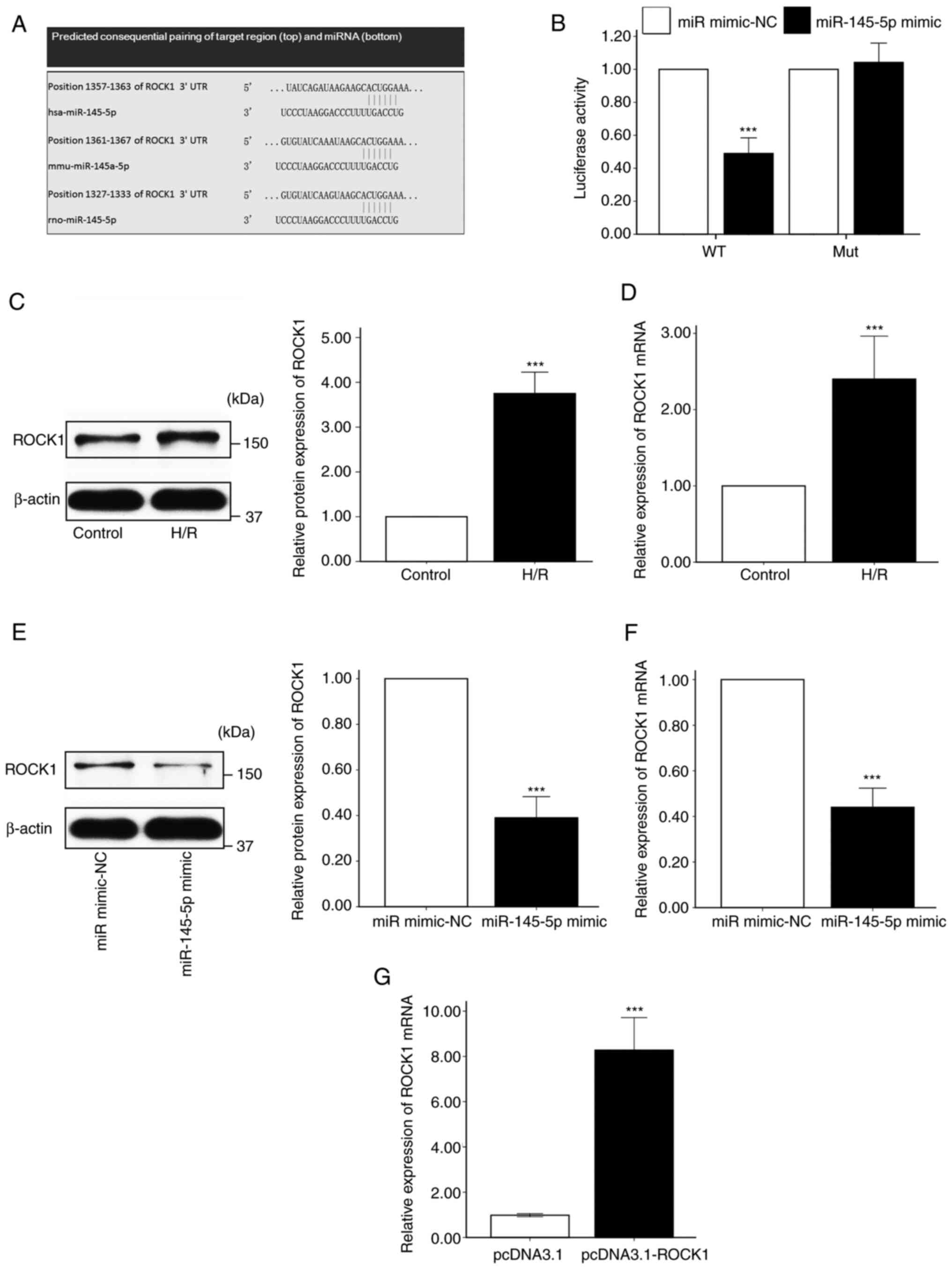
miR-145-5p directly targets ROCK1 and
negatively regulates its expression
TargetScan was used to predict target genes that may
be regulated by miR-145-5p, and ROCK1 was revealed as a possible
target (Fig. 3A). The potential
regulation of ROCK1 by miR-145-5p was investigated using a
luciferase reporter assay. The results demonstrated that luciferase
activity was significantly repressed in the presence of miR-145-5p
mimic compared with the miR mimic-NC in cells transfected with the
wild-type ROCK1 3' UTR, while the miR-145-5p mimic exhibited no
inhibitory effects on luciferase activity in cells transfected with
the mutant ROCK1 3' UTR (Fig. 3B).
In addition, western blot and RT-qPCR analysis revealed that the
protein and mRNA levels of ROCK1 were significantly upregulated in
H9c2 cells following H/R compared with those in H9c2 cells
maintained under normal conditions (Fig. 3C and D). Furthermore, the transfection of H9c2
cells with miR-145-5p mimic significantly downregulated ROCK1
protein and mRNA expression (Fig.
3E and F). These results
indicate that miR-145-5p specifically targeted ROCK1.
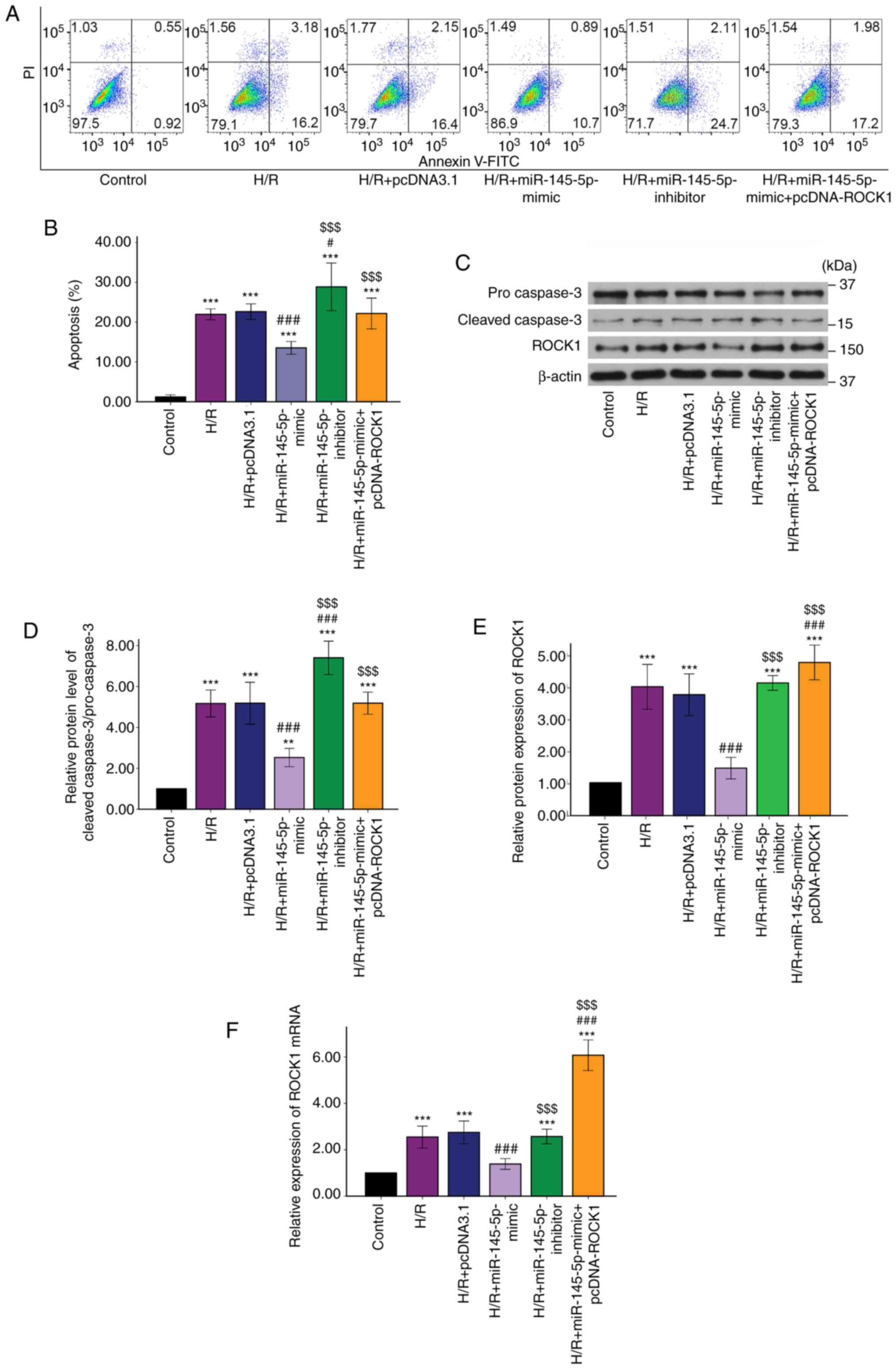
miR-145-5p protects H9c2 cells
subjected to H/R by targeting ROCK1
To investigate the mechanism underlying the effects
of miR-145-5p 1, further experiments were conducted using cells
transfected with a ROCK1 overexpression vector (Fig. 3G). Annexin V-FITC/PI double staining
demonstrated that the H9c2 cardiomyocyte apoptosis rate
significantly decreased following transfection with miR-145-5p
mimic and increased following transfection with miR-145-5p
inhibitor. However, the apoptosis rate of H9c2 cardiomyocytes
transfected with miR-145-5p mimic + pcDNA-ROCK1 was significantly
increased compared with that of the H9c2 cardiomyocytes transfected
with miR-145-5p mimic alone (Fig.
4A and B). Similarly,
transfection with miR-145-5p mimic + pcDNA-ROCK1 significantly
increased caspase-3 cleavage compared with that in the miR-145-5p
mimic group (Fig. 4C and D). These results demonstrate that
miR-145-5p was unable to protect H9c2 cardiomyocytes against
H/R-induced apoptosis when ROCK1 was overexpressed. The H/R
treatment induced a significant increase in the expression of ROCK1
in the cardiomyocytes at the protein and mRNA levels. Following
transfection with miR-145-5p mimic, ROCK1 protein and mRNA
expression levels were significantly decreased in the H9c2 cells
subjected to H/R. By contrast, transfection with the miR-145-5p
inhibitor resulted in ROCK1 protein and mRNA expression levels
comparable with those in the H/R group (Fig. 4E and F). In addition, when the cardiomyocytes
were co-transfected with miR-145-5p mimic and pcDNA-ROCK1 vector,
the effects of the miR-145-5p mimic on ROCK1 protein and mRNA
levels were abolished (Fig. 4E and
F). Based on these results, it was
concluded that miR-145-5p reduced H/R-induced cardiomyocyte
apoptosis by targeting ROCK1.
Discussion
miRNAs are non-coding RNAs that regulate gene
expression by inhibiting transcription or degrading mRNA (27). The expression of miRNAs undergoes
changes in numerous disease states, including myocardial I/R injury
(5). Studies have shown that some
miRNAs promote fibrosis and apoptosis while others have opposite
effects during myocardial I/R injury. The aim of the present study
was to investigate the effects of miR-145-5p on cell apoptosis in a
H9c2 cardiomyocyte H/R model. The findings confirmed that
miR-145-5p has a protective effect against H/R injury in H9c2
cardiomyocytes. Additionally, the present study demonstrated that
ROCK1 is a target of miR-145-5p.
Increasing evidence, summarized in a recent review,
has shown that miR-145-5p is associated with vascular biology and
the pathophysiology of cardiovascular disease (28). For example, a previous study
reported that miR-145-5p promoted the apoptosis of cardiomyocytes
after myocardial I/R in rats (12).
However, other studies demonstrated that miR-145-5p protected
against myocardial I/R injury in murine models via anti-apoptotic,
anti-inflammatory and autophagy pathways (10,11,29).
Hence, the role of miR-145-5p in myocardial I/R injury remains
unclear. In the present study, after subjecting H9c2 cells to H/R,
RT-qPCR was performed to detect the expression of miR-145-5p and
the results revealed that its expression in the H/R-treated cells
was significantly decreased. Following the discovery that
miR-145-5p was repressed in H9c2 cardiomyocytes subjected to H/R, a
miR-145-5p mimic and inhibitor were each transfected into H9c2
cells to assess the role of miR-145-5p in cardiomyocytes exposed to
H/R. Flow cytometry was used to detect the apoptosis rate of the
H9c2 cardiomyocytes. Transfection with miR-145-5p mimic
significantly reduced the H/R-induced apoptosis of the H9c2
cardiomyocytes. This finding suggests that miR-145-5p plays a
protective role against myocardial I/R injury.
Our previous studies indicated that ROCK has an
important role in myocardial I/R injury. One of these studies
revealed that ROCK inhibition reduced infarct size and the extent
of cardiomyocyte apoptosis induced by myocardial I/R injury in
vivo (30), while another
identified increased ROCK activity in the peripheral blood
leukocytes of patients with AMI undergoing primary percutaneous
coronary intervention, suggesting that ROCK activity is increased
in patients with myocardial I/R (31). ROCK has two isoforms, ROCK1 and
ROCK2, which are ~64% similar in sequence and have overlapping
substrates (32). However, the two
isoforms may have different roles in various types of cells and
tissues (33,34). For example, a study indicated that
ROCK1 protects the heart against pressure overload-induced heart
failure with postcapillary pulmonary hypertension while ROCK2
compromises it (34).
Based on a TargetScan database search and
double-luciferase reporter assay, the present study demonstrated
that miR-145-5p directly targets ROCK1. A previous study showed
that ROCK1 is targeted by miR-145-5p in laryngocarcinoma cells, and
the suppression of ROCK1 displays effects similar to those of
miR-145-5p (35). Consistent with
this, the present study detected a negative interaction between
miR-145-5p and ROCK1 expression in H9c2 cardiomyocytes. ROCK1
expression in cardiomyocytes was significantly increased by H/R
compared with that in control myocytes cultured under normal
conditions. In addition, the levels of ROCK1 mRNA and protein were
reduced in H9c2 cardiomyocytes transfected with miR-145-5p mimic
compared with H9c2 cardiomyocytes without transfection or
transfected with miR-145-5p inhibitor. These results indicate that
ROCK1 expression is negatively regulated by miR-145-5p in H9c2
cardiomyocytes. The present findings also indicate a positive
association of ROCK1 expression with apoptosis in H9c2
cardiomyocytes subjected to H/R. Elevated miR-145-5p expression may
rescue cardiomyocytes from apoptosis during H/R injury via the
inhibition of ROCK1. Notably, the overexpression of ROCK1
significantly attenuated the miR-145-5p-induced inhibition of cell
apoptosis following H/R, which further confirms the relationship
between miR-145-5p and ROCK1.
It should be noted that the experiments in the
present study were conducted only in H9c2 cardiomyocytes, and the
significance and implications of the results require verification
in another cardiac cell line or stem cell-derived cardiomyocytes.
Moreover, only in vitro experiments were performed. Further
studies with in vivo experiments are required.
In conclusion, the present study showed that
miR-145-5p is significantly downregulated in H/R, and indicated
that the overexpression of miR-145-5p reduced H/R-induced H9c2
cardiomyocyte apoptosis by targeting ROCK1. These results
demonstrate a cardioprotective effect of miR-145-5p and suggested
its potential as a therapeutic approach for the alleviation of
myocardial I/R injury.
Acknowledgements
Not applicable.
Funding
Funding: The authors gratefully acknowledge research support
provided by the Youth Foundation of the National Natural Science
Foundation of China (grant no. 81600284) and Shandong Key Research
and Development Project (grant no. 2016GSF201196).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ designed the experiments and drafted the
manuscript. CC and DLX performed the experiments and analyzed the
data. XBL and SJB analyzed and interpreted the data. JZ and CC
confirm the authenticity of all the raw data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Authors' information
ORCID of Dr Juan Zhang: 0000-0001-6241-7176.
References
|
1
|
Hausenloy DJ and Yellon DM: Myocardial
ischemia-reperfusion injury: A neglected therapeutic target. J Clin
Invest. 123:92–100. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
2
|
Davidson SM, Ferdinandy P, Andreadou I,
Bøtker HE, Heusch G, Ibáñez B, Ovize M, Schulz R, Yellon DM,
Hausenloy DJ, et al: Multitarget strategies to reduce myocardial
ischemia/reperfusion injury: JACC review topic of the week. J Am
Coll Cardiol. 73:89–99. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Gottlieb RA and Pourpirali S: Lost in
translation: miRNAs and mRNAs in ischemic preconditioning and
ischemia/reperfusion injury. J Mol Cell Cardiol. 95:70–77.
2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Siebert V, Allencherril J, Ye Y, Wehrens
XHT and Birnbaum Y: The role of non-coding RNAs in ischemic
myocardial reperfusion injury. Cardiovasc Drugs Ther. 33:489–498.
2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Wang X, Zhang X, Ren XP, Chen J, Liu H,
Yang J, Medvedovic M, Hu Z and Fan GC: MicroRNA-494 targeting both
proapoptotic and antiapoptotic proteins protects against
ischemia/reperfusion-induced cardiac injury. Circulation.
122:1308–1318. 2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Cordes KR, Sheehy NT, White MP, Berry EC,
Morton SU, Muth AN, Lee TH, Miano JM, Ivey KN and Srivastava D:
miR-145 and miR-143 regulate smooth muscle cell fate and
plasticity. Nature. 460:705–710. 2009.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Lovren F, Pan Y, Quan A, Singh KK, Shukla
PC, Gupta N, Steer BM, Ingram AJ, Gupta M, Al-Omran M, et al:
MicroRNA-145 targeted therapy reduces atherosclerosis. Circulation.
126 (Suppl 1):S81–S90. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Caruso P, Dempsie Y, Stevens HC, McDonald
RA, Long L, Lu R, White K, Mair KM, McClure JD, Southwood M, et al:
A role for miR-145 in pulmonary arterial hypertension: Evidence
from mouse models and patient samples. Circ Res. 111:290–300.
2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Liu Z, Tao B, Fan S, Pu Y, Xia H and Xu L:
MicroRNA-145 protects against myocardial ischemia reperfusion
injury via CaMKII-mediated antiapoptotic and anti-inflammatory
pathways. Oxid Med Cell Longev. 2019(8948657)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Qi Z, Li S, Su Y, Zhang J, Kang Y, Huang
Y, Jin F and Xing Q: Role of microRNA-145 in protection against
myocardial ischemia/reperfusion injury in mice by regulating
expression of GZMK with the treatment of sevoflurane. J Cell
Physiol: Mar 14, 2019 (Epub ahead of print).
|
|
12
|
Wu G, Tan J, Li J, Sun X, Du L and Tao S:
miRNA-145-5p induces apoptosis after ischemia-reperfusion by
targeting dual specificity phosphatase 6. J Cell Physiol: Mar 18,
2019 (Epub ahead of print).
|
|
13
|
Shimokawa H, Sunamura S and Satoh K:
RhoA/Rho-kinase in the cardiovascular system. Circ Res.
118:352–366. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Shimokawa H: 2014 Williams harvey lecture:
Importance of coronary vasomotion abnormalities-from bench to
bedside. Eur Heart J. 35:3180–3193. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Kandabashi T, Shimokawa H, Miyata K,
Kunihiro I, Kawano Y, Fukata Y, Higo T, Egashira K, Takahashi S,
Kaibuchi K and Takeshita A: Inhibition of myosin phosphatase by
upregulated rho-kinase plays a key role for coronary artery spasm
in a porcine model with interleukin-1beta. Circulation.
101:1319–1323. 2000.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ikeda S, Satoh K, Kikuchi N, Miyata S,
Suzuki K, Omura J, Shimizu T, Kobayashi K, Kobayashi K, Fukumoto Y,
et al: Crucial role of rho-kinase in pressure overload-induced
right ventricular hypertrophy and dysfunction in mice. Arterioscler
Thromb Vasc Biol. 34:1260–1271. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Uehata M, Ishizaki T, Satoh H, Ono T,
Kawahara T, Morishita T, Tamakawa H, Yamagami K, Inui J, Maekawa M
and Narumiya S: Calcium sensitization of smooth muscle mediated by
a Rho-associated protein kinase in hypertension. Nature.
389:990–994. 1997.PubMed/NCBI View
Article : Google Scholar
|
|
18
|
Shimizu T, Fukumoto Y, Tanaka S, Satoh K,
Ikeda S and Shimokawa H: Crucial role of ROCK2 in vascular smooth
muscle cells for hypoxia-induced pulmonary hypertension in mice.
Arterioscler Thromb Vasc Biol. 33:2780–2791. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Xiang SY, Vanhoutte D, Del Re DP, Purcell
NH, Ling H, Banerjee I, Bossuyt J, Lang RA, Zheng Y, Matkovich SJ,
et al: RhoA protects the mouse heart against ischemia/reperfusion
injury. J Clin Invest. 121:3269–3276. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
20
|
Riento K and Ridley AJ: Rocks:
Multifunctional kinases in cell behaviour. Nat Rev Mol Cell Biol.
4:446–456. 2003.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Chang J, Xie M, Shah VR, Schneider MD,
Entman ML, Wei L and Schwartz RJ: Activation of Rho-associated
coiled-coil protein kinase 1 (ROCK-1) by caspase-3 cleavage plays
an essential role in cardiac myocyte apoptosis. Proc Natl Acad Sci
USA. 103:14495–14500. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yue X, Yang X, Lin X, Yang T, Yi X, Dai Y,
Guo J, Li T, Shi J, Wei L, et al: Rnd3 haploinsufficient mice are
predisposed to hemodynamic stress and develop apoptotic
cardiomyopathy with heart failure. Cell Death Dis.
5(e1284)2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Hartmann S, Ridley AJ and Lutz S: The
function of Rho-associated kinases ROCK1 and ROCK2 in the
pathogenesis of cardiovascular disease. Front Pharmacol.
6(276)2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Shi J, Zhang YW, Yang Y, Zhang L and Wei
L: ROCK1 plays an essential role in the transition from cardiac
hypertrophy to failure in mice. J Mol Cell Cardiol. 49:819–828.
2010.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Degterev A, Boyce M and Yuan J: A decade
of caspases. Oncogene. 22:8543–8567. 2003.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Boon RA, Jaé N, Holdt L and Dimmeler S:
Long noncoding RNAs: From clinical genetics to therapeutic targets?
J Am Coll Cardiol. 67:1214–1226. 2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Vacante F, Denby L, Sluimer JC and Baker
AH: The function of miR-143, miR-145 and the MiR-143 host gene in
cardiovascular development and disease. Vascul Pharmacol.
112:24–30. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Hu S, Cao S, Tong Z and Liu J: FGF21
protects myocardial ischemia-reperfusion injury through reduction
of miR-145-mediated autophagy. Am J Transl Res. 10:3677–3688.
2018.PubMed/NCBI
|
|
30
|
Zhang J, Li XX, Bian HJ, Liu XB, Ji XP and
Zhang Y: Inhibition of the activity of Rho-kinase reduces
cardiomyocyte apoptosis in heart ischemia/reperfusion via
suppressing JNK-mediated AIF translocation. Clin Chim Acta.
401:76–80. 2009.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhang J, Xu F, Liu XB, Bi SJ and Lu QH:
Increased Rho kinase activity in patients with heart
ischemia/reperfusion. Perfusion. 34:15–21. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Strassheim D, Gerasimovskaya E, Irwin D,
Dempsey EC, Stenmark K and Karoor V: RhoGTPase in vascular disease.
Cells. 8(551)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Shimizu T and Liao JK: Rho kinases and
cardiac remodeling. Circ J. 80:1491–1498. 2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Sunamura S, Satoh K, Kurosawa R, Ohtsuki
T, Kikuchi N, Elias-Al-Mamun M, Shimizu T, Ikeda S, Suzuki K, Satoh
T, et al: Different roles of myocardial ROCK1 and ROCK2 in cardiac
dysfunction and postcapillary pulmonary hypertension in mice. Proc
Natl Acad Sci USA. 115:E7129–E7138. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhuang S, Liu F and Wu P: Upregulation of
long noncoding RNA TUG1 contributes to the development of
laryngocarcinoma by targeting miR-145-5p/ROCK1 axis. J Cell
Biochem. 120:13392–13402. 2019.PubMed/NCBI View Article : Google Scholar
|