Introduction
Sepsis is one of the most common diseases in
patients in intensive care units and is associated with
unacceptably high mortality rates (1). The pathogenetic causes of, and
relevant therapies for sepsis are current areas of intense
research. The intestinal tract is the largest storage site of
bacteria and endotoxins in the human body (2). Gut permeability is increased in
patients with sepsis and is associated with the development of
systemic inflammatory response syndrome (SIRS) and multiple organ
dysfunction syndrome (MODS) (3-5).
The concomitant compromised proliferation and increased apoptosis
of gastrointestinal epithelial cells aggravates the dysfunction of
the mucosal epithelial barrier and exacerbates sepsis, leading to a
vicious cycle that results in severe sepsis. Therefore,
understanding the mechanism of the pathological changes in the
intestinal epithelial cells during dysfunctional proliferation and
apoptosis is key to preventing the progression of gut-origin
sepsis.
As endogenous non-coding small RNA molecules,
microRNAs (miRNAs) are widely found in plant and animal cells
(6). Their main role is to regulate
the expression of target genes at the post-transcriptional level in
eukaryotic organisms to adjust processes, including cell
proliferation, differentiation and apoptosis following different
stimuli. In recent years, attention has focused on the role of
miRNAs in the pathogenesis of sepsis. Lan et al (7) demonstrated that the expression of
miR-133a-3p was significantly higher in the sepsis group compared
with the control group in a cecal ligation and perforation mouse
model. Clinical trials indicated that serum miR-133a-3p secretion
was significantly increased in critically ill patients,
particularly patients with sepsis, and was correlated with sepsis
severity, sequential organ failure assessment score, C-reactive
protein level and procalcitonin level. The results suggested that
miR-133a-3p may be used as a marker for the diagnosis of sepsis;
however, the specific mechanism of its action remained unclear
(7). Transgelin-2 (TAGLN2) is an
actin-binding protein which mediates a variety of tumor cell
activities, including proliferation, apoptosis, invasion and
metastasis (8-12).
However, the effect of TAGLN2 on intestinal epithelial cells is
unknown. In the present study, the effects of miR-133a-3p on
proliferation and apoptosis, and the specific mechanism of its
actions, were investigated in human intestinal epithelial cells.
The results demonstrated that lipopolysaccharide (LPS) stimulation
of intestinal epithelial cells increased the expression of
miR-133a-3p and decreased the expression of TAGLN2. The results
suggested that miR-133a-3p is involved in the regulation of
intestinal epithelial cells in patients with sepsis. To the best of
our knowledge, the present study was the first to report an
association between miR-133a-3p and TAGLN2. The present study
provided novel insights into the regulatory effect of miR-133a-3p
on the proliferation and apoptosis of intestinal epithelial cells,
and poses a novel hypothesis that miR-133a-3p regulates the
apoptosis of intestinal epithelial cells by targeting TAGLN2.
Materials and methods
Cell culture
The epithelium was isolated by a modification of a
previously published method (13).
In brief, the intestine was divided into 2-3 mm lengths and washed
in Hank's Balanced Salt Solution with 0.5 mM DTT (Sigma-Aldrich;
Merck KGaA) at 4˚C for 5 min. The fractions were transferred to
chelating buffer (2 mM EDTA in PBS; Gibco; Thermo Fisher
Scientific, Inc.) and incubated at 4˚C for 20 min with constant
stirring prior to being transferred to a tube containing 20 ml cold
chelating buffer. The fractions were incubated at 4˚C for 10 min
with constant stirring, following which they were centrifuged at a
low speed (150-200 x g; 4˚C; 2 min). The cell pellets were
collected and resuspended with complete medium (DMEM supplemented
with 2 mM L-glutamine, 100 U/ml penicillin and 100 µg/ml
streptomycin; Gibco; Thermo Fisher Scientific, Inc.). The
intestinal epithelial FHs 74 Int cell line was obtained from the
American Type Culture Collection. The cells were maintained in
Dulbecco's modified Eagle's medium (DMEM; HyClone), supplemented
with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc.). The cells were cultured at 37˚C in
a humidified incubator with 5% CO2 and were treated with
1 µg/ml LPS (Sigma-Aldrich; Merck KGaA).
Cell transfection
miR-133a mimics, negative control (NC) mimics,
miR-133a inhibitor, NC inhibitor and TAGLN2-small interfering RNAs
(siRNAs) designed and constructed by Shanghai GenePharma Co., Ltd.
were transfected into cells (40 nM each) using Lipofectamine 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocols. The sequences were as follows:
miR-133a mimics, 5'-UUUGGUCCCCUUCAACCAGCUG-3'; NC mimics,
5'-UUUUCCGAACGUUCACGUTT-3'; miR-133a inhibitor,
5'-CAGCUGGUUGAAGGGGACCAAA-3'; NC inhibitor,
5'-CAGUACUUUUGUGUAGUACAA-3'; TAGLN2 siRNA,
5'-GCAAGAACGUGAUCGGGUU-3'; NC siRNA, 5'-AGUACUGCUUACGAUACGG-3'. The
transfection concentration was determined according to the
manufacturer's instructions. Following incubation for 48 h, the
cells were subjected to subsequent experiments. The efficiency of
the transfections was determined using reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
Cell proliferation assay
To determine the effects of miR-133-a-3p and
TAGLN2-siRNA transfection on intestinal epithelial cell
proliferation, the proliferation of transfected cells was assessed
using a Cell Counting kit-8 (CCK-8; Biomol), according to the
manufacturer's protocols. Cells were plated at 2 x103
cells per well in 96-well plates. At the indicated time points, the
cells were treated with 10 µl CCK-8 solution and incubated in the
dark for another 2 h. The absorbance was measured at a wavelength
of 450 nm. Cell proliferation was also examined under a light
microscope (Olympus Corporation). A total of four regions of
interest (ROI) were randomly selected in each well and the cell
density was examined and compared between groups. Representative
images of ROI were provided.
Flow cytometry (FCM)
To investigate the effect of miR-133a-3p and
TAGLN2-siRNA transfection on the apoptosis of intestinal epithelial
cells, an Annexin V-FITC kit (NeoBioscience Technology Co., Ltd.)
was used according to the manufacturer's protocols. The cells were
harvested, washed with PBS and suspended in the binding buffer at
1x106 cells/ml. The Annexin V and FITC were added to the
cells and gently mixed. The cells were then incubated for 15 min at
room temperature in the dark. Binding buffer (Invitrogen; Thermo
Fisher Scientific, Inc.) was added to each tube prior to analysis
using a flow cytometer (BD FACSDiva; BD Biosciences) and FlowJo
version 10 software (FlowJo LLC) according to the manufacturer's
instructions.
RT-qPCR
Total RNA was extracted from the cells using either
an miRNAeasy Mini kit (Qiagen, Inc.) or a TRIzol RNA Purification
kit (Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocols. qPCR was performed using a SYBR Green
qPCR kit (Takara Bio, Inc.). A reaction mixture (20 µl) containing
total RNA (1 µg) was transcribed to cDNA at 55˚C for 15 min and
85˚C for 2 min using HiScript II Q RT SuperMix for qPCR (Vazyme
Biotech Co., Ltd.). miRNA qPCR was performed using All-in-One miRNA
RT-qPCR (GeneCopoeia, Inc.). The reaction system was established
according to the manufacturer's protocols using the following
primers: miR-133a-3p forward, 5'-UUUGGUCCCCUUCAACCAGCUG-3' and
reverse, 5'-UAAACCAAGGUAAAAUGGUCGA-3'; TAGLN2 forward,
5'-CGCTTGAACGCTCCCCG-3' and reverse, 5'-TTCTGGAAGTTCTCGCGTCC-3';
and GAPDH forward, 5'-CACTCCTCCACCTTTGA-3' and reverse,
5'-CCACCACCCTGTTGCTG-3'. The conditions were set as follows:
Pre-denaturation at 95˚C for 10 min; 40 cycles of denaturation at
95˚C for 10 sec, annealing at 60˚C for 20 sec and extension at 72˚C
for 35 sec. The 2-ΔΔCq method was used to measure the
relative mRNA expression and GAPDH was used as the control
(14).
Western blot analysis
The cells were washed with PBS and lysed in RIPA
lysis buffer (Beyotime Institute of Biotechnology). The protein
content was determined with a BCA protein assay kit (Beyotime
Institute of Biotechnology). A total of 50 µg protein from each
treatment group was loaded and separated using 10% SDS-PAGE and
transferred onto PVDF membranes. The membranes were blocked with 5%
skimmed milk at room temperature for 1-2 h, and were incubated with
primary antibodies against Bax (cat. no. ab32503), Bcl2 (cat. no.
ab182858), TAGLN2 (cat. no. ab121146) and GAPDH (cat. no. ab8245;
all from Abcam; all 1:1,000) at 4˚C overnight. The membranes were
washed three times with TBST and incubated with an anti-rabbit
(cat. no. ab97051) or anti-mouse HRP-conjugated secondary antibody
(cat. no. ab6728; both from Abcam; both 1:10,000) at room
temperature for 2 h. The target proteins were visualized using an
ECL detection system (Invitrogen; Thermo Fisher Scientific, Inc.).
Protein expression was quantified with Image-Pro Plus 6.0 (Media
Cybernetics, Inc.).
Luciferase reporter assay and
bioinformatics analysis
To detect the effect of miR-133a-3p on the activity
of the TAGLN2 3'-untranslated region (UTR), the PCR amplification
product of the TAGLN2 3'-UTR fragment was integrated into the pGL3
plasmid XbaI restriction site, and a wild-type (WT) pGL3-WT-TAGLN2
eukaryotic expression vector was constructed. The GeneTailor
Site-Directed Mutagenesis system (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to mutate the miR-133p-3p binding region
in TAGLN2 3'-UTR to generate a pGL3-mut-TAGLN2 plasmid. A total of
48 h after transfection using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), the cells were washed twice with
PBS and cell lysates were prepared. The lysates were transferred
into white opaque 96-well microplates. A Dual Luciferase Reporter
system (Promega Corporation) was used according to the
manufacturer's protocols, and the luciferase activity was measured
in a microplate reader after 10 min and was normalized to the
Renilla luciferase activity. The experiments were repeated
at least three times. Bioinformatics analysis was conducted using
miRanda version 3.3a (https://anaconda.org/bioconda/miranda).
Statistical analysis
Statistical analysis was performed using SPSS 20.0
software (IBM Corp.). The results are expressed as the mean ±
standard error of the mean of at least three independent
experiments. Data were analyzed using one-way analysis of variance
followed by Dunnett's post hoc test for multiple comparisons or
Student's t-test for comparisons between two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-133a-3p is upregulated and TAGLN2
is downregulated in LPS-treated intestinal epithelial cells
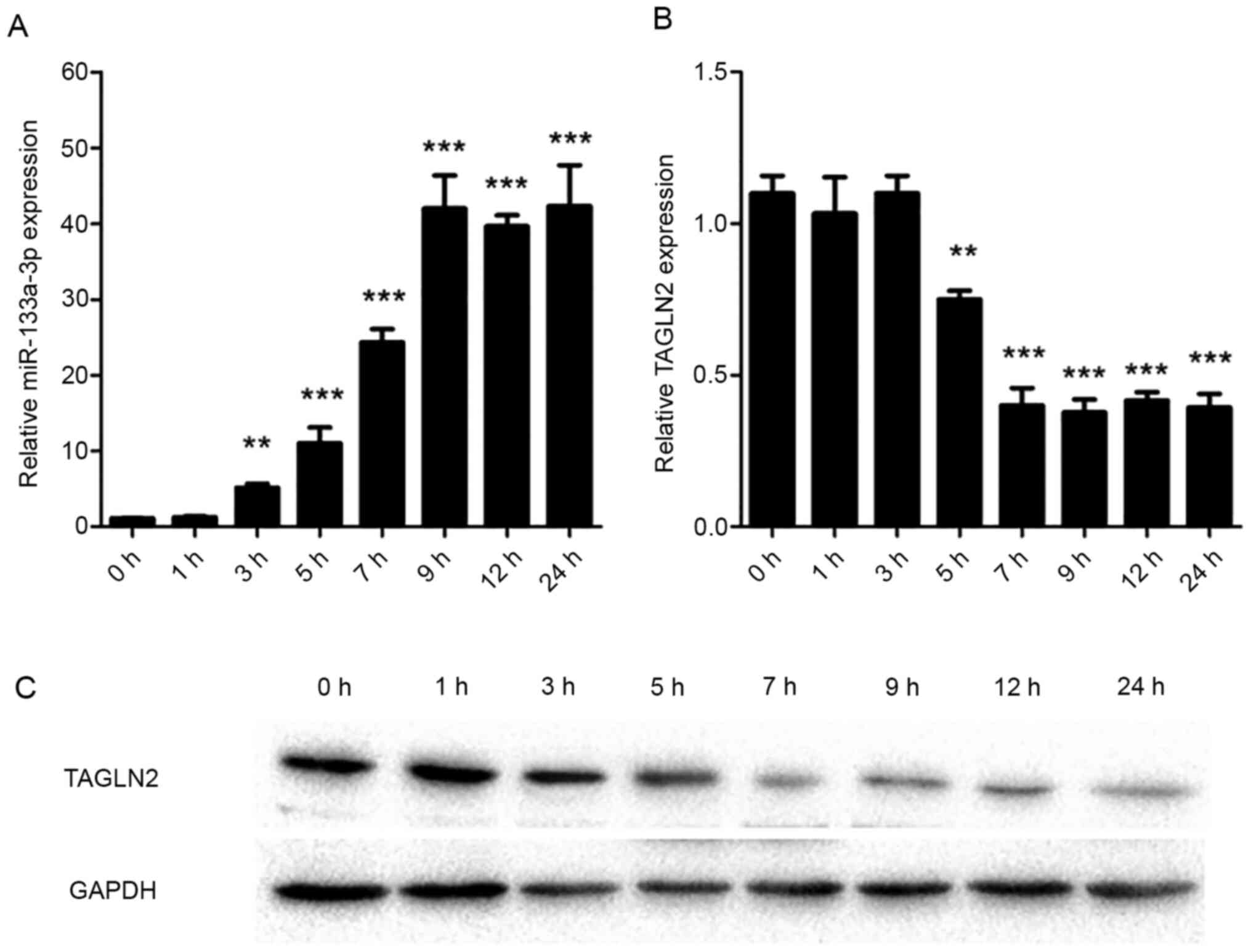
qPCR was used to assess the expression levels of
miR-133a-3p and TAGLN2 in intestinal epithelial cells. The results
of the present study demonstrated that miR-133a-3p increased 3 h
after LPS treatment (P=0.007) and reached a peak 9 h (P<0.001)
after treatment (Fig. 1A). By
contrast, the expression of TAGLN2 was negatively correlated with
the expression of miR-133a-3p. Expression of TAGLN2 began to
decrease 5 h after the treatment (P=0.008), and the expression was
lowest at 7 h (P<0.001; Fig.
1B). Western blot analysis results, consistent with the RT-qPCR
results, demonstrated a decrease in TAGLN2 levels (Fig. 1C).
miR-133a-3p inhibits the proliferation
of intestinal epithelial cells
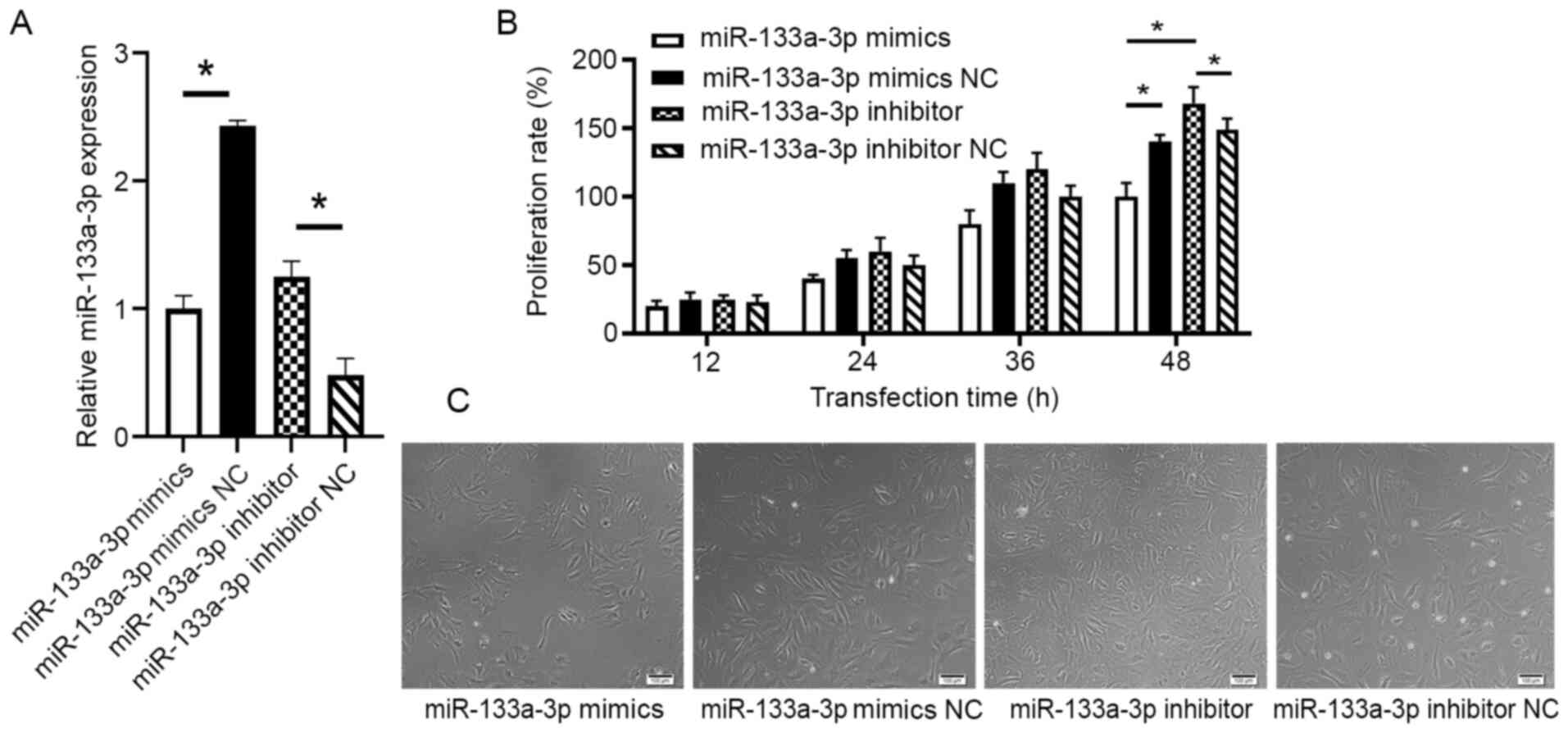
To detect the effect of miR-133a-3p on the
proliferation of intestinal epithelial cells, FHs 74 Int cells were
transfected with either miR-133a mimics, NC mimics, miR-133a
inhibitor or NC inhibitor. The efficiency of the transfection was
tested with RT-qPCR (P<0.05; Fig.
2A). The results demonstrated that while proliferation,
measured using the CCK-8 assay, gradually increased following 24 h
of culture, the proliferation rate of the mimics transfection group
was markedly lower than that of the mimics NC group following 48 h
of culture (P=0.013). The proliferation rate of the cells
transfected with miR-133a inhibitor was significantly higher than
that of the NC inhibitor group following 48 h of culture (P=0.03;
Fig. 2B). The differences in
proliferation seen in the four treatment groups were also confirmed
using microscopy (Fig. 2C).
miR-133a-3p promotes the apoptosis of
FHs 74 Int cells
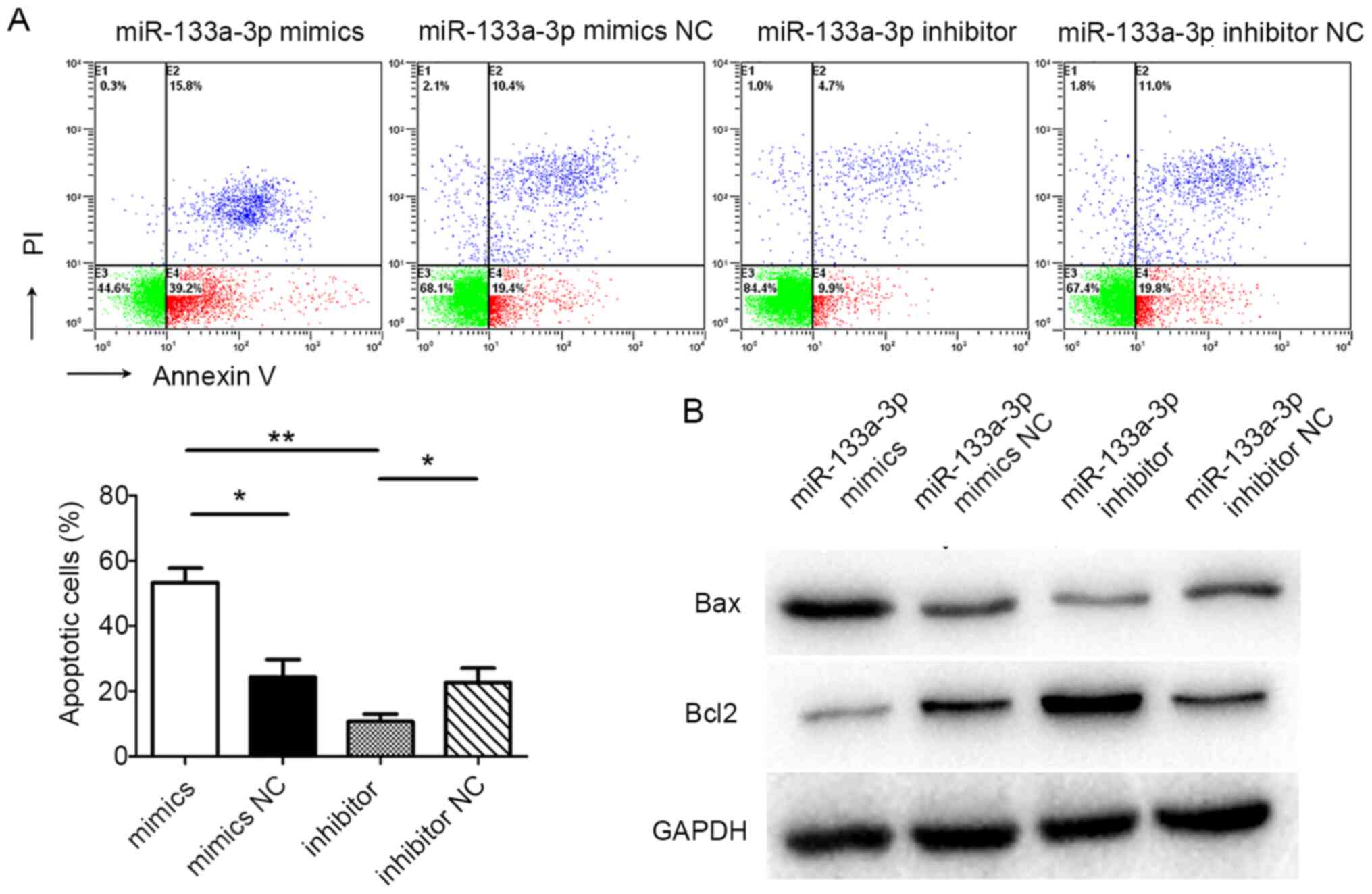
Flow cytometry was used to detect cell apoptosis.
The flow cytometric results suggested that the apoptotic rate of
cells in the miR-133a-3p mimics group was significantly higher than
in the NC mimics group (P=0.044). Furthermore, the apoptotic rate
of cells in the miR-133a-3p inhibitor group was markedly lower than
in the NC inhibitor group (P=0.026). In addition, the apoptotic
rate of cells in the miR-133a-3p mimics group was significantly
higher than in the miR-133a-3p inhibitor group (P=0.007; Fig. 3A). Western blotting was used to
assess the levels of apoptotic markers in FHs 74 Int cells
following transfection. The amount of the pro-apoptotic protein Bax
in miR-133a-3p mimic-transfected cells was significantly higher
than in the control group, while in the miR-133a-3p inhibitor
group, Bax levels were decreased compared with the control group.
The levels of the anti-apoptotic protein Bcl2 was inversely
correlated with the levels of Bax (Fig.
3B). These results suggested that miR-133a-3p promoted the
apoptosis of FHs 74 Int cells.
miR-133a-3p regulates FHs 74 Int cells
by targeting TAGLN2
Bioinformatics analysis demonstrated that the 3'-UTR
of TAGLN2 mRNA had complementary paired sequences with miR-133a-3p
(15), suggesting a binding site
between miR-133a-3p and TAGLN2 mRNA (Fig. 4A). Results from the dual-luciferase
reporter assay indicated that, in the TAGLN2-WT-transfected group,
luciferase activity was significantly lower in the miR-133a-3p
mimic-transfected cells than in the NC cells (P=0.004; Fig. 4B). Furthermore, no significant
difference in luciferase activity was identified between the
miR-133a-3p mimics group and the NC group in the TAGLN2 mutant
(Mut) (P=0.84) and empty vector control co-transfected group
(P=0.93). These results suggested that miR-133a-3p may directly
bind TAGLN2 mRNA, thereby affecting TAGLN2 expression. Western blot
analysis revealed that transfection with miR-133a-3p mimics
markedly decreased TAGLN2 at the protein level, while miR-133a-3p
inhibitor transfection significantly increased TAGLN2 protein
expression (Fig. 4C).
To investigate whether TAGLN2 served a role in the
effect of miR-133a-3p on intestinal epithelial cell proliferation
and apoptosis, TAGLN2 expression was downregulated using TAGLN2
siRNA, prior to a CCK-8 assay and flow cytometry being conducted to
assess proliferation and apoptosis, respectively. The results
suggested that, compared with the control group, the TAGLN2 siRNA
group had a markedly lower proliferation rate (P=0.029; Fig. 4D) and a higher rate of apoptosis
(P=0.013; Fig. 4E). Western blot
analysis demonstrated that the TAGLN2 siRNA group had a lower Bcl2
level and a higher Bax level than the control group (Fig. 4F).
Discussion
Sepsis is a SIRS caused by infection. It is often
caused by severe infection following trauma, burns or a weakened
immune response. Severe cases may develop into septic shock,
disseminated intravascular coagulation and multi-organ failure
(16,17), which is associated with a high
mortality rate. Increased awareness of sepsis and more targeted
research into the syndrome have led to an improved understanding of
the disease (18,19). The pathogenesis of sepsis is
associated with the complex interaction between the infectious
agent, host immune system and coagulation system (17,20,21).
Despite great advances in the understanding of the pathogenesis,
sepsis remains a health concern, contributing toward more than 5
million deaths worldwide (22).
Gut injury is a key characteristic of sepsis and
gut-origin sepsis has been recognized as the start of SIRS and MODS
in critically ill patients (4). The
overgrowth of pathogenic microbiome in the intestinal tract and
intestinal barrier failure are critical processes in the
development of gut-origin sepsis. Translocation of enteric
bacteria, pathogen-associated molecular patterns (PAMPs) and LPS
from the intestinal lumen damage the intestinal epithelial cells
and exacerbate intestinal barrier failure (5,23-25).
Multiple dysregulated processes caused by the compromised gut
integrity occur during sepsis, including inhibition of
proliferation, activation of apoptosis and increased intestinal
permeability. The gut epithelium consists of a single layer of
columnar cells which is continuously renewed. Proliferating cells
are restricted to a particular region while cells that migrate to
the villus tip die by apoptosis or are exfoliated (26,27).
The number of functional epithelial cells is therefore dependent on
the balance between cell proliferation and apoptosis.
miRNAs serve a critical role in regulating growth,
maintaining homeostasis and participating in pathophysiological
disease processes, and have become an area of intense research in
recent years. miRNAs are also present and involved in the
development of sepsis and may either be critical in the
pathogenesis of sepsis or key to immune regulation and therapy for
sepsis. The first miRNA to be detected in the serum of patients
with sepsis was miR-150, and it was suggested to be an early
biomarker of sepsis (28). Abnormal
expression of miR-146a and miR-223 have also been identified in the
serum of patients with sepsis; these, along with miR-122 and
miR-21, have been suggested as biomarkers for the diagnosis of
sepsis (29,30). A previous study also confirmed that
miR-499 serves a significant role in the diagnosis of myocardial
injury in sepsis (31). A number of
previous studies have also focused on miR-133a, and its role in
cell differentiation, skeletal muscle proliferation, cancer
development and fibrosis has been confirmed (32-36).
The results of the present study demonstrated that
the expression of miR-133a-3p was significantly increased in
LPS-challenged intestinal epithelial cells. Therefore, we
hypothesized that miR-133a-3p was associated with sepsis and that
effective regulation of miR-133a-3p would be beneficial in the
treatment of sepsis. Following transfection with either miR-133a
mimics, an miR-133a inhibitor or a NC, cell proliferation was
determined using a CCK-8 assay and apoptosis was assessed using
FCM. The results of the present study demonstrated that miR-133a-3p
inhibited the proliferation and promoted the apoptosis of
intestinal epithelial cells. Western blot analysis showed that
miR-133a-3p increased the level of the pro-apoptotic protein, Bax,
and decreased the level of the anti-apoptotic protein, Bcl2. These
results indicated that miR-133a-3p had a regulatory effect on the
proliferation and apoptosis of intestinal epithelial cells. The
present study further investigated the specific regulatory
molecular mechanism.
TAGLN2 is an actin-binding protein, which
participates in the process of fiber polymerization and
depolymerization by interacting with actin fibers. It serves an
important role in the physiological processes of stabilizing and
regulating the cytoskeleton (8,37).
Recent studies of TAGLN2 focused on its role in a variety of
malignant tumors and its effect on apoptosis (9,38).
TAGLN2 expression increased significantly in maxillary sinus
squamous carcinoma, and it mediated the proliferation, apoptosis,
invasion and migration of cancer cells (12). The current study showed that
following LPS treatment, the expression of TAGLN2 mRNA and the
amount of TAGLN2 protein was significantly downregulated in
intestinal epithelial cells and was negatively correlated with
miR-133a-3p. Based on the aforementioned findings, we hypothesized
that miR-133a-3p regulated the proliferative and apoptotic activity
of intestinal epithelial cells by targeting the expression of
TAGLN2. However, whether TAGLN2 is the direct target of miR-133a-3p
remains to be clarified. With the construction of WT and Mut TAGLN2
3'-UTR dual-luciferase reporter plasmids, it was revealed that,
compared with the control group, luciferase activity was
significantly decreased in the miR-133a-3p mimics and TAGLN2 3'-UTR
WT co-transfected group. These results further confirmed that the
expression of TAGLN2 was directly regulated by miR-133a-3p. Changes
in the expression of TAGLN2 also affected the levels of apoptotic
Bax and Bcl2 proteins as detected by western blot analysis,
indicating the role of TAGLN2 in the regulation of intestinal
epithelial cells by miR-133a-3p.
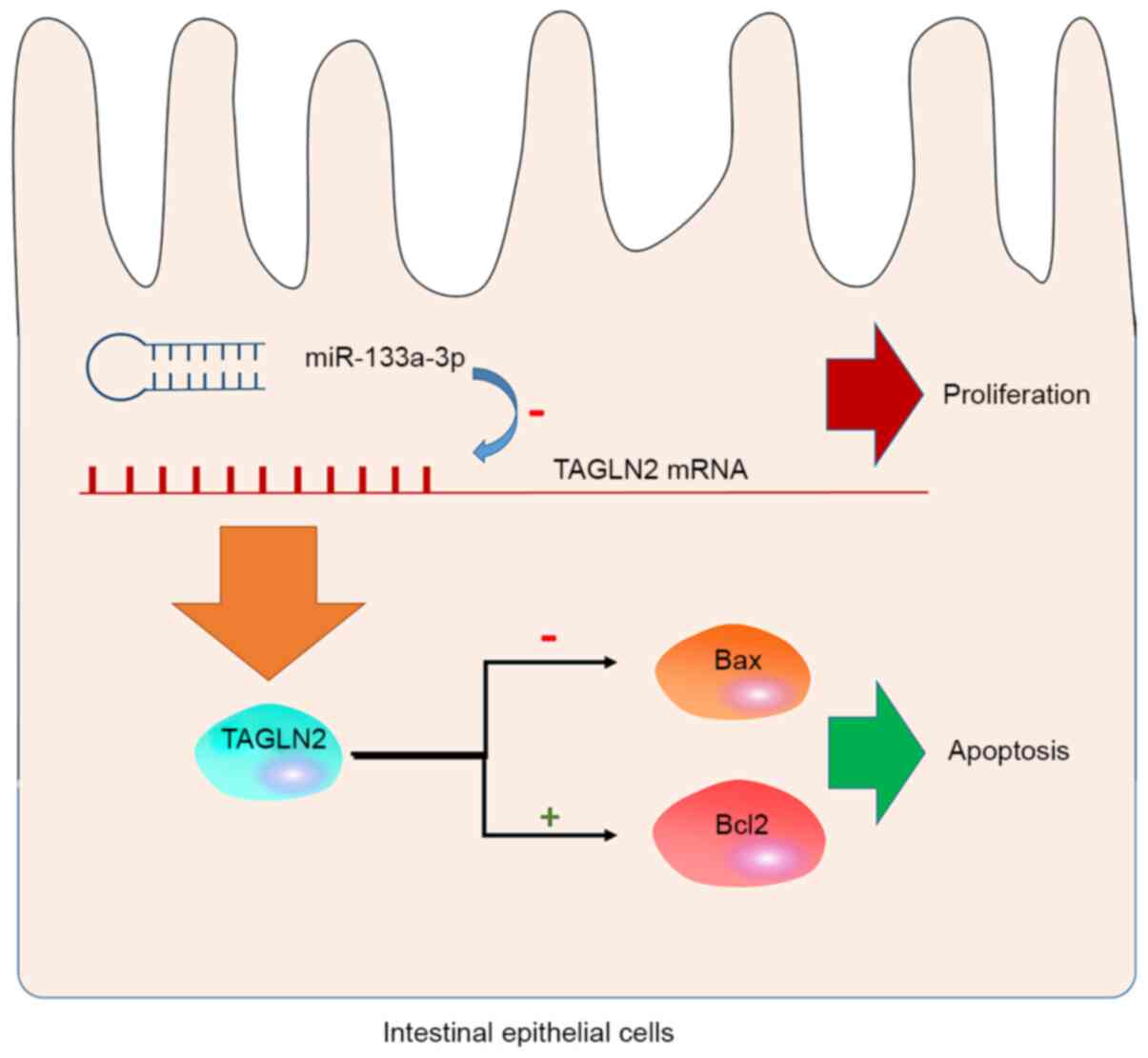
In summary, the present study revealed, for the
first time, that miR-133a-3p serves a regulatory role in the
proliferation and apoptosis of intestinal epithelial cells. The
regulatory effects of miR-133a-3p on intestinal epithelial cells is
associated with its inhibitory effects on TAGLN2 expression. In the
intestinal epithelial cells, miR-133a-3p inhibited the expression
of TAGLN2 on the mRNA level and caused a decrease in the production
of TAGLN2 protein. This effect inhibited the anti-apoptotic effect
of TAGLN2 by downregulating Bax and upregulating Bcl2, leading to
increased apoptosis in the intestinal epithelial cells (Fig. 5). This conclusion is based on in
vitro experiments and would require confirmation using mouse
models in the future. Nevertheless, the present study identified a
novel mechanism for the regulation of intestinal epithelial cells
and provided a hypothesis on which to base target interventions for
the clinical treatment of sepsis.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the General Project
of Shaanxi Province Key Research and Development Plan (grant no.
S2017-ZDYF-YBXM-0353).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XT and LL designed the research and confirmed the
authenticity of the raw data. GF and JW performed the experiments.
QH analyzed the data. XT and CZ drafted the manuscript and analyzed
the data. BQ and JW collected and interpreted the data. LL revised
the final manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Goulden R, Hoyle MC, Monis J, Railton D,
Riley V, Martin P, Martina R and Nsutebu E: qSOFA, SIRS and NEWS
for predicting inhospital mortality and ICU admission in emergency
admissions treated as sepsis. Emerg Med J. 35:345–349.
2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Sittipo P, Lobionda S, Lee YK and Maynard
CL: Intestinal microbiota and the immune system in metabolic
diseases. J Microbiol. 56:154–162. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Frazier TH, DiBaise JK and McClain CJ: Gut
microbiota, intestinal permeability, obesity-induced inflammation,
and liver injury. JPEN J Parenter Enteral Nutr. 35 (5
Suppl):14S–20S. 2011.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Assimakopoulos SF, Triantos C, Thomopoulos
K, Fligou F, Maroulis I, Marangos M and Gogos CA: Gut-origin sepsis
in the critically ill patient: Pathophysiology and treatment.
Infection. 46:751–760. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Vaishnavi C: Translocation of gut flora
and its role in sepsis. Indian J Med Microbiol. 31:334–342.
2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lan C, Shi X, Guo N, Pei H and Zhang H:
Value of serum miR-155-5p and miR-133a-3p expression for the
diagnosis and prognosis evaluation of sepsis. Zhonghua Wei Zhong
Bing Ji Jiu Yi Xue. 28:694–698. 2016.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
8
|
Jeon BN, Kim HR, Chung YS, Na BR, Park H,
Hong C, Fatima Y, Oh H, Kim CH and Jun CD: Actin stabilizer TAGLN2
potentiates adoptive T cell therapy by boosting the inside-out
costimulation via lymphocyte function-associated antigen-1.
Oncoimmunology. 7(e1500674)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Pei J, Li P, Zhang ZY, Zhang HL, Gao YH,
Wang DY, Zheng Y, Xu X and Cui JZ: Effect of TAGLN2 in the
regulation of meningioma tumorigenesis and development. Eur Rev Med
Pharmacol Sci. 22:307–313. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhang H, Jiang M, Liu Q, Han Z, Zhao Y and
Ji S: miR-145-5p inhibits the proliferation and migration of
bladder cancer cells by targeting TAGLN2. Oncol Lett. 16:6355–6360.
2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Leung WK, Ching AK, Chan AW, Poon TC, Mian
H, Wong AS, To KF and Wong N: A novel interplay between oncogenic
PFTK1 protein kinase and tumor suppressor TAGLN2 in the control of
liver cancer cell motility. Oncogene. 30:4464–4475. 2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Meng T, Liu L, Hao R, Chen S and Dong Y:
Transgelin-2: A potential oncogenic factor. Tumour Biol.
39(1010428317702650)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Sato T and Clevers H: Primary mouse small
intestinal epithelial cell cultures. Methods Mol Biol. 945:319–328.
2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Arocho A, Chen B, Ladanyi M and Pan Q:
Validation of the 2-DeltaDeltaCt calculation as an alternate method
of data analysis for quantitative PCR of BCR-ABL P210 transcripts.
Diagn Mol Pathol. 15:56–61. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Li AY, Yang Q and Yang K: miR-133a
mediates the hypoxia-induced apoptosis by inhibiting TAGLN2
expression in cardiac myocytes. Mol Cell Biochem. 400:173–181.
2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Cecconi M, Evans L, Levy M and Rhodes A:
Sepsis and septic shock. Lancet. 392:75–87. 2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Salomao R, Ferreira BL, Salomao MC, Santos
SS, Azevedo LCP and Brunialti MKC: Sepsis: Evolving concepts and
challenges. Braz J Med Biol Res. 52(e8595)2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Berg D and Gerlach H: Recent advances in
understanding and managing sepsis. F1000Res 7: F1000 Faculty
Rev-1570, 2018.
|
|
19
|
Keeley A, Hine P and Nsutebu E: The
recognition and management of sepsis and septic shock: A guide for
non-intensivists. Postgrad Med J. 93:626–634. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Levi M and van der Poll T: Coagulation and
sepsis. Thromb Res. 149:38–44. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
van der Poll T, van de Veerdonk FL,
Scicluna BP and Netea MG: The immunopathology of sepsis and
potential therapeutic targets. Nat Rev Immunol. 17:407–420.
2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Rhodes A, Evans LE, Alhazzani W, Levy MM,
Antonelli M, Ferrer R, Kumar A, Sevransky JE, Sprung CL, Nunnally
ME, et al: Surviving sepsis campaign: International guidelines for
management of sepsis and septic shock: 2016. Crit Care Med.
45:486–552. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Amornphimoltham P, Yuen PST, Star RA and
Leelahavanichkul A: Gut leakage of fungal-derived inflammatory
mediators: Part of a gut-liver-kidney axis in bacterial sepsis. Dig
Dis Sci. 64:2416–2428. 2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Haak BW and Wiersinga WJ: The role of the
gut microbiota in sepsis. Lancet Gastroenterol Hepatol. 2:135–143.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Haak BW, Prescott HC and Wiersinga WJ:
Therapeutic potential of the gut microbiota in the prevention and
treatment of sepsis. Front Immunol. 9(2042)2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chopra DP, Dombkowski AA, Stemmer PM and
Parker GC: Intestinal epithelial cells in vitro. Stem Cells Dev.
19:131–142. 2010.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chelakkot C, Ghim J and Ryu SH: Mechanisms
regulating intestinal barrier integrity and its pathological
implications. Exp Mol Med. 50(103)2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Vasilescu C, Rossi S, Shimizu M, Tudor S,
Veronese A, Ferracin M, Nicoloso MS, Barbarotto E, Popa M,
Stanciulea O, et al: MicroRNA fingerprints identify miR-150 as a
plasma prognostic marker in patients with sepsis. PLoS One.
4(e7405)2009.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Essandoh K, Li Y, Huo J and Fan GC:
miRNA-mediated macrophage polarization and its potential role in
the regulation of inflammatory response. Shock. 46:122–131.
2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang JF, Yu ML, Yu G, Bian JJ, Deng XM,
Wan XJ and Zhu KM: Serum miR-146a and miR-223 as potential new
biomarkers for sepsis. Biochem Biophys Res Commun. 394:184–188.
2010.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhou R, Huang W, Fan X, Liu F, Luo L, Yuan
H, Jiang Y, Xiao H, Zhou Z, Deng C and Dang X: miR-499 released
during myocardial infarction causes endothelial injury by targeting
alpha7-nAchR. J Cell Mol Med. 23:6085–6097. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Tang Y, Pan J, Huang S, Peng X, Zou X, Luo
Y, Ren D, Zhang X, Li R, He P and Wa Q: Downregulation of
miR-133a-3p promotes prostate cancer bone metastasis via activating
PI3K/AKT signaling. J Exp Clin Cancer Res. 37(160)2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang Y, Ma J, Qiu W, Zhang J, Feng S, Zhou
X, Wang X, Jin L, Long K, Liu L, et al: Guanidinoacetic acid
regulates myogenic differentiation and muscle growth through
miR-133a-3p and miR-1a-3p Co-mediated Akt/mTOR/S6K signaling
pathway. Int J Mol Sci. 19(2837)2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
He B, Lin X, Tian F, Yu W and Qiao B:
miR-133a-3p inhibits oral squamous cell carcinoma (OSCC)
proliferation and invasion by suppressing COL1A1. J Cell Biochem.
119:338–346. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Jin LW, Ye HY, Xu XY, Zheng Y and Chen Y:
miR-133a/133b inhibits Treg differentiation in IgA nephropathy
through targeting FOXP3. Biomed Pharmacother. 101:195–200.
2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Wei P, Xie Y, Abel PW, Huang Y, Ma Q, Li
L, Hao J, Wolff DW, Wei T and Tu Y: Transforming growth factor
(TGF)-β1-induced miR-133a inhibits myofibroblast differentiation
and pulmonary fibrosis. Cell Death Dis. 10(670)2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Kim HR, Kwon MS, Lee S, Mun Y, Lee KS, Kim
CH, Na BR, Kim BNR, Piragyte I, Lee HS, et al: TAGLN2 polymerizes
G-actin in a low ionic state but blocks Arp2/3-nucleated actin
branching in physiological conditions. Sci Rep.
8(5503)2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Yoshino H, Chiyomaru T, Enokida H,
Kawakami K, Tatarano S, Nishiyama K, Nohata N, Seki N and Nakagawa
M: The tumour-suppressive function of miR-1 and miR-133a targeting
TAGLN2 in bladder cancer. Br J Cancer. 104:808–818. 2011.PubMed/NCBI View Article : Google Scholar
|