Introduction
Hepatic ischemia-reperfusion injury (IRI) is the
phenomenon in which hepatocellular damage is aggravated after the
reperfusion of blood flow in the liver (1). Hepatic IRI occurs in numerous clinical
situations, such as liver transplantation, liver resection, trauma,
shock and hemorrhage (2,3). The incidence of nonalcoholic fatty
liver disease (NAFLD) has markedly increased and is currently
estimated at approximately 20-30% in the general population and up
to 70-80% in the obese population (4). An increasing trend of IRI in fatty
liver is predicted in the near future (3). Clinical studies and animal experiments
revealed that steatotic livers are particularly susceptible to IRI
(5). Fatty liver grafts are
associated with a primary non-function rate of 60% compared to 5%
for non-steatotic grafts and increase the risk of postoperative
morbidity and mortality after fatty liver surgery (6,7).
However, there are few effective methods to reduce the
susceptibility of fatty liver to IRI. Recent studies demonstrated
that inflammation played an important role in the process of fatty
liver IRI, including more obvious inflammatory cell infiltration
and inflammatory factor release (8-10).
Toll-like receptors (TLRs) form a part of the host
innate immune system that triggers the innate immune response and
production of inflammatory mediators (11). TLR4 is a key TLR family member that
is an essential participant in hepatic IRI. TLR4 is ubiquitously
expressed in hepatocytes and nonparenchymal cells (12,13).
TLR4 activation initiates signal transduction pathways. Then,
transcription factors such as NF-κB are activated and cytokines are
produced (14). TLR4-deficient mice
have decreased levels of inflammatory cytokines and are
significantly protected against hepatic IRI (15).
Augmenter of liver regeneration (ALR), also referred
to as hepatic stimulator substance (HSS), was first identified for
its promotion of liver regeneration after partial hepatectomy
(16). ALR plays an important role
in protecting hepatocytes against various injuries by reducing
endoplasmic reticulum stress and improving mitochondrial function
(17,18). A previous study has revealed that
ALR also reduced the inflammatory response. ALR reduced the
infiltration of CD4+ T cells and neutrophils to
alleviate normal liver IRI (19).
However, the relationship between ALR and the inflammatory response
in fatty liver IRI and the underlying mechanisms remain unclear. It
was hypothesized that ALR protects against fatty liver IRI via
inhibition of the TLR4/NF-κB pathway to reduce inflammatory cell
infiltration and inflammatory factor release.
Materials and methods
Animals and model of fatty liver
IRI
Experiments were performed on male C57BL/6 mice (age
~6-8 weeks; weight ~18-22 g) that were purchased from the Academy
of Military Sciences (Beijing, China). All animals were treated
humanely according to the Guidelines for Animal Care and Use in
Medical Research (20) and the
experimental methods are approved by The Ethics Committee of
Capital Medical University (Beijing, China; approval no.
AEEI-2016-094). A total of 30 male mice were housed at a constant
room temperature (22-25˚C), a relative humidity of 40-60% under a
12/12 h light-dark cycle with access to water and food ad
libitum. Mice were fed a methionine-choline-deficient (MCD)
diet (Beijing HFK Bioscience Co., Ltd.) for 2 weeks, and then
underwent warm liver IRI (21,22),
while the control mice were fed an isocaloric control diet.
Briefly, mice were anesthetized with 4% chloral hydrate
intraperitoneally (320 mg/kg), and the artery/portal vein blood
flow to the left and middle lobes of the liver was temporarily
interrupted using an atraumatic clip, which induced ~70% liver
ischemia. Liver ischemia was maintained for 30 min, and the clip
was loosened to allow reperfusion for 6 h. Mice in the
sham-operated group underwent the same surgical procedure without
clamping of the hepatic vessels. At the end of the experiments,
mice were sacrificed by inhalation of anesthetics in a closed
chamber (e.g. an overdose of 5% isoflurane). Mice continued to be
exposed to isoflurane until they stopped breathing for 1 min, and
then they were removed from the chamber to confirm death through
the absence of a heartbeat and nerve reflexes. Blood samples were
collected from the inferior vena cava, and liver tissues were
prepared for further analysis.
Gene transfer
Human ALR cDNA was cloned into a
replication-deficient adenoviral vector with the flag tag as
previously described (21). Three
days before IRI, mice subjected to an MCD diet were randomly
divided into 4 groups: 3 groups were injected with Adnull (control
virus, 1x109 PFU/mouse), AdALR (1x109
PFU/mouse) or normal saline (NS) via the tail vein. Sham-operated
mice served as controls.
Biochemical assays and triglyceride
determination
The activities of serological alanine
aminotransferase (ALT) and aspartate aminotransferase (AST) and the
concentrations of serological triglyceride (TG) were analyzed using
an automatic biochemical analyzer (Hitachi, Ltd.) at the Clinical
Laboratory Center of Capital Medical University. The TG content in
liver tissue was determined using a TG kit (cat. no. K622;
Biovision, Inc.) according to the manufacturer's protocol.
Enzyme-linked immunosorbent assay
(ELISA)
The levels of interleukin (IL)-1β, IL-6, and tumor
necrosis factor-α (TNF-α) in murine serum were assessed using
commercially available ELISA kits (cat. nos. MLB00C, M6000B,
MTA00B, respectively; R&D Systems, Inc.) according to the
manufacturer's protocol based on the quantitative sandwich enzyme
immunoassay technique. Briefly, 50 µl of murine serum was incubated
in ELISA plates at room temperature for 2 h. After washing with
washing buffer, 100 µl of conjugated antibody was added and
incubated at room temperature for another 2 h. The substrate
solution was added and incubated for 20 min in the dark, followed
by the addition of a stopping solution. The optical density (OD)
was measured at 450 nm, and the concentrations of the cytokines
were calculated by reference to the standard curves.
Histology and Oil Red O staining
Liver tissues from the ischemic lobes were fixed in
10% buffered formalin for 24 h at room temperature and embedded in
paraffin. For hematoxylin and eosin (H&E; 200X) staining,
sections of 5-µm thickness were stained in hematoxylin for 5 min at
room temperature, then transferred to 1% hydrochloric acid alcohol
for differentiation and washed in running water for 2 min. After
soaking in the eosin stain for 3 min at room temperature, sections
were washed in running water. The Suzuki score was used to evaluate
liver injury based on 3 indices: sinusoidal congestion, hepatocyte
necrosis and ballooning degeneration (23). To evaluate the intracellular lipid
distribution in the liver, liver samples were fixed in 4%
paraformaldehyde for 24 h at room temperature frozen in liquid
nitrogen and sectioned into 6-µm slices. Sections were stained in
Oil Red O solution for 15 min at room temperature, rinsed with 60%
isopropanol, stained with hematoxylin at room temperature for 30
sec, and images were captured using light microscopy
(magnification, x100).
Immunohistochemistry
Immunohistochemical staining was performed as
previously described (21).
Briefly, the liver tissue sections (5-mm) were dewaxed, hydrated
and pretreated using the heat-induced antigen retrieval technique.
Sections were treated sequentially with a 3%
H2O2 solution and 5% BSA to block endogenous
peroxidase activity and nonspecific antibody binding, respectively.
Primary antibodies directed against ALR (1:500; cat. no.
11293-1-AP; Proteintech Group, Inc.), mouse EGF-like
module-containing mucin-like hormone receptor-like 1 (F4/80; 1:200;
cat. no. ab6640; Abcam), myeloperoxidase (MPO; 1:200; cat. no.
ab90810; Abcam), proliferating cell nuclear antigen (PCNA; 1:500;
cat. no. 13110; Cell Signaling Technology, Inc.), TLR4 (1:100; cat.
no. sc-293072; Santa Cruz Biotechnology, Inc.) and NF-κB p65
(1:200; cat. no. 8242; Cell Signaling Technology, Inc.) were
incubated with the sections overnight at 4˚C, followed by
HRP-conjugated IgG incubation (1:200; cat. no. SA00001-2;
ProteinTech Group, Inc.) Sections were visualized with
diaminobenzidine (DAB) and counterstained with hematoxylin. The
positively stained areas and positive cells were evaluated using
Image-Pro Plus 6.0 software (Media Cybernetics Inc.). Staining was
quantified in 10 randomly selected x200 high-power fields per
tissue sample.
To detect cellular apoptosis in liver tissue, a
terminal deoxynucleotidyltransferase (dUTP)-mediated nick
end-labeling (TUNEL) assay was performed using an In Situ
Cell Death/Apoptosis Detection kit (cat. no. 11684817910; Roche
Diagnostics, GmbH) according to the manufacturer's protocol.
Briefly, the liver tissue sections were dewaxed in an incubator at
60˚C for 20 min followed by deparaffinization in xylene for 10 min,
and then were soaked in 100, 95, 90, 80 and 70% gradient alcohol
for 5 min each at room temperature for rehydration. Slides were
incubated with Proteinase K working solution for 20 min at room
temperature, followed by incubation with TUNEL reaction mixture for
60 min at 37˚C in the dark. Next, Converter-POD antibody (1:500)
from aforementioned TUNEL kit was used for further reaction for 30
min at 37˚C, followed by the addition of DAB substrate for 10 min
at room temperature. The sections were then counterstained with
hematoxylin at room temperature for 2 min and mounted using
glycerol. The results were scored semi-quantitatively by averaging
the numbers of TUNEL-positive cells per high-power field
(magnification, x200) for 10 fields per tissue sample with ImageJ
1.5 software (National Institutes of Health).
Cytoplasmic and nuclear protein
extraction
Cytoplasmic and nuclear protein extraction was
performed using a commercial kit (NT-032; Invent Biotechnologies,
Inc.) according to the manufacturer's instructions. Briefly, 20-30
mg liver tissue was grinded with a pestle in 250 µl buffer A for
approximately 1 min. After centrifugation at 14,000 x g for 5 min
at 4˚C, a total of 200 µl of supernatant was collected and stored
as the cytosolic fraction. The pellet was grinded 60-100 times with
the pestle, followed by the addition of 0.8 ml buffer A and grinded
a few more times. After incubation on ice for 8 min, a total of 0.7
ml of supernatant was transferred to a fresh tube and centrifuged
at 500 x g for 2 min at 4˚C. Then the pellet was resuspended in 0.5
ml buffer B by vortex for 20 sec and incubated on ice for 5 min.
After centrifugation at 2,000 x g for 2 min at 4˚C, nuclear
extracts were collected for further analysis.
Western blot analysis
Immunoblotting was performed as previously described
(17). Briefly, liver tissues were
homogenized, and samples containing 50 µg of protein were loaded
and transferred to PVDF membranes. The following primary antibodies
(all at 1:1,000) were used for immunoblotting: Flag tag (cat. no.
F9291; Sigma-Aldrich; Merck KGaA), ALR, TLR4 and NF-κB p65.
Antibodies against glyceraldehyde 3-phosphate dehydrogenase (GAPDH;
1:5,000; 10494-1-AP; ProteinTech Group, Inc.) and β-actin (1:5,000;
cat. no. 20536-1-AP; ProteinTech Group, Inc.) were used as the
loading controls. Membranes were then incubated with horseradish
peroxidase (HRP)-conjugated goat anti-rabbit IgG secondary
antibodies (1:5,000; cat. no. SA00001-2; ProteinTech Group, Inc.)
for 1 h at room temperature. The relative densities of the protein
bands were quantitatively determined using ImageJ 1.5 software
(National Institutes of Health).
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was extracted from liver tissue using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. A total of 2 µg of
RNA was reversely transcribed to cDNA using a cDNA synthesis kit
(cat. no. 6215A; Takara Bio, Inc.) according to the manufacturer's
instructions. Quantitative PCR was performed (ABI7500) using
SYBR-Green mix (Qiagen GmbH). The thermocycling conditions were as
follows: 95˚C For 5 min for pre-denaturation; 40 cycles of 95˚C for
15 sec, 60˚C for 1 min and 72˚C for 45 sec. 18S rRNA was amplified
in parallel as an internal control. The 2-ΔΔCq method
was used to determine the relative mRNA level (24). The primers used are mentioned in
Table SI.
Statistical analysis
The experimental results are expressed as the means
± standard deviations (SDs) of three independent experiments. For
multiple comparisons, one-way analysis of variance (ANOVA) followed
by Tukey's post hoc test was performed using GraphPad Prism 6
software (GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
ALR expression is downregulated during
IRI in steatotic liver
After 2 weeks of MCD diet feeding, mice were
subjected to hepatic IRI, i.e., 30 min of ischemia followed by 6 h
of reperfusion. The expression of endogenous ALR was evaluated
using western blot analysis. The results demonstrated that IRI
decreased the expression of ALR (Fig.
1A), which indicated that ALR is involved in IRI.
Adenovirus-mediated gene transfer of ALR was performed in mice 3
days before IRI. Exogenously delivered ALR was expressed
efficiently within the liver, as determined by western blotting
(Fig. 1B) and immunostaining
(Fig. 1C). Mice fed the MCD diet
exhibited elevated TG levels in liver tissues and mild-to-moderate
hepatic steatosis, as evidenced by Oil Red O staining (P<0.01;
Fig. 1E and F). Adenovirus-mediated ALR gene transfer
did not alter TG levels in the blood (Fig. 1D).
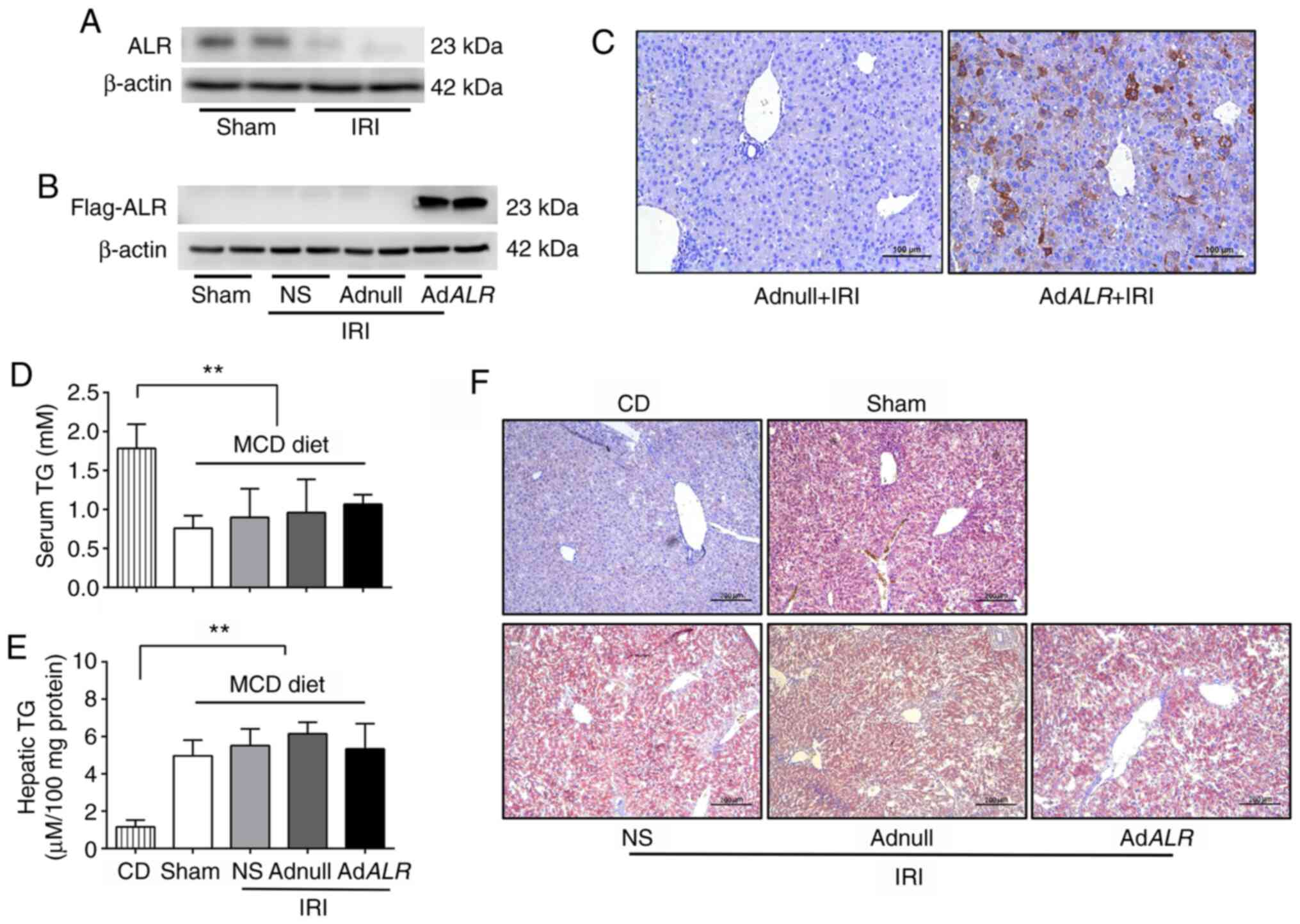 | Figure 1Expression of ALR is downregulated
and related to IRI in steatotic liver. C57BL/6 mice were fed an MCD
diet for 2 weeks, and control mice were fed an isocaloric control
diet. Mice fed the MCD diet were divided into 4 groups.
Sham-operated mice served as controls, and the other 3 groups
received normal saline, Adnull (control virus) or AdALR gene
transfer 3 days before being subjected to IRI, i.e., 30 min of
ischemia followed by 6 h of reperfusion. (A) Expression of
endogenous ALR in the liver was detected using western blotting. (B
and C) Expression of exogenously delivered ALR was detected using
western blotting and immunohistochemistry. Scale bars, 10 µm. (D)
Serum TG levels in mice. (E) Liver tissue TG levels in mice. (F)
Oil Red O staining of liver sections. Scale bars, 200 µm.
**P<0.01; n=6 per group. ALR, augmenter of liver
regeneration; IRI, ischemia-reperfusion injury; MCD,
methionine-choline-deficient; CD, isocaloric control diet; NS,
normal saline; TG, triglyceride. |
ALR expression maintains liver
function and attenuates IRI in steatotic liver
Hepatocellular function was evaluated by measuring
serum ALT and AST levels. These levels were significantly decreased
following IRI in the AdALR group compared with the Adnull
group (serum ALT: 395±42 vs. 1163±138 U/l, P<0.01; serum AST:
539±69 vs. 1214±145 U/l, P<0.01) (Fig. 2A and B). The liver histology was associated with
the serum indices. IRI significantly induced hepatic damage, as
indicated by the severe sinusoidal congestion, hepatocellular
necrosis and cytoplasmic vacuolization. However, hepatic damage was
significantly reduced in the AdALR group compared with the
Adnull group and the NS group (Fig.
2C). The Suzuki score was used for histological evaluation of
hepatic injury, and these scores also confirmed the protective role
of ALR against hepatic IRI (P<0.05; Fig. 2D). A TUNEL assay was performed to
evaluate cellular apoptosis. Few TUNEL-positive cells were observed
in the sham group, but significantly increased numbers were
observed in the Adnull and NS groups after hepatic IRI. Conversely,
the number of TUNEL-positive hepatocytes was decreased in the
AdALR group compared with the NS and Adnull groups, which
indicated that ALR expression protected hepatocytes against
IRI-induced cell death (P<0.01; Fig.
2E and F). The hepatocyte
proliferation is demonstrated in Fig.
2G. The number of the proliferating cells in the
ALR-transfected mice were increased compared with the Adnull and NS
mice, which indicated that ALR enhanced cell growth in the liver
after IRI (P<0.01; Fig. 2G and
H).
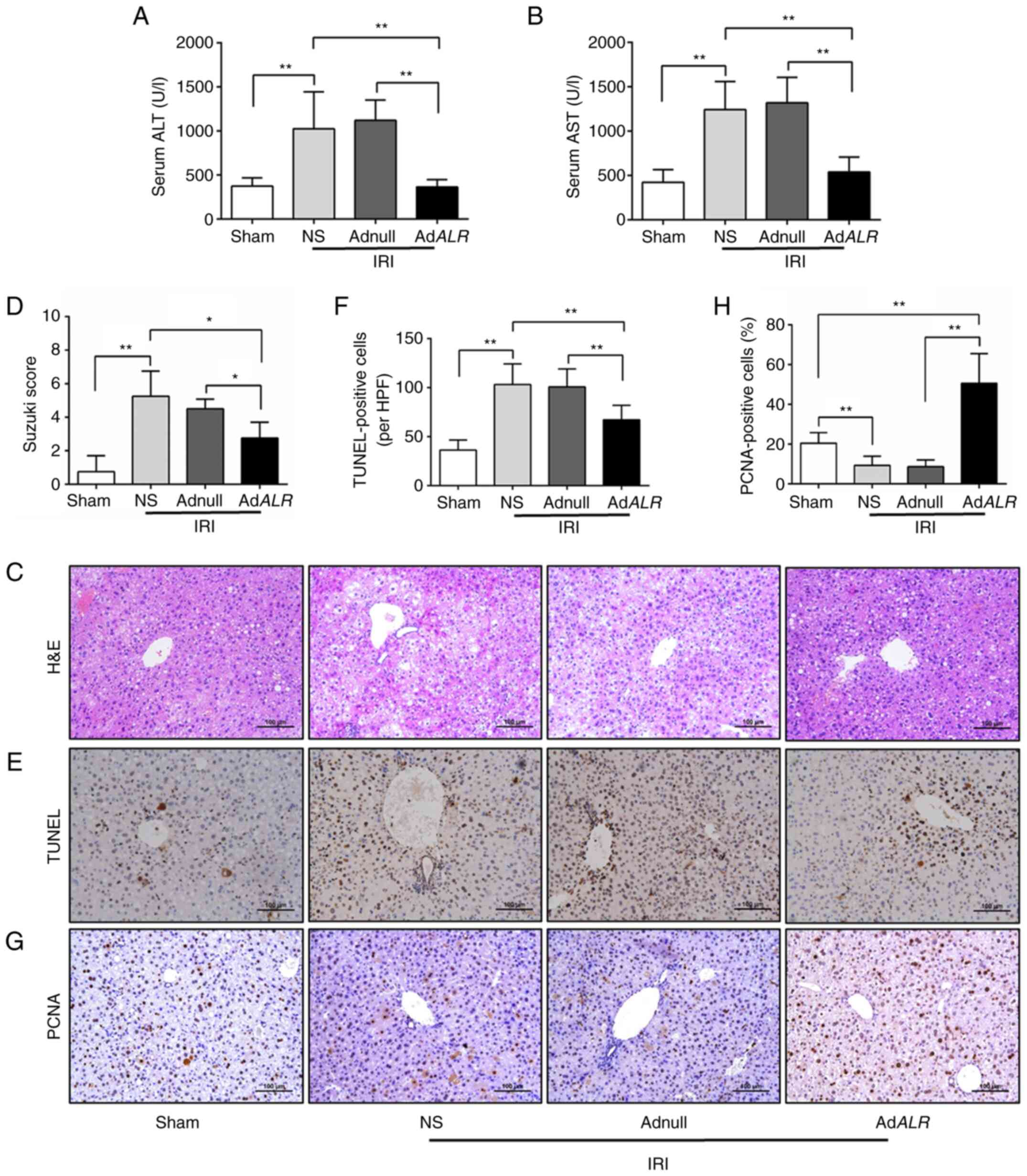 | Figure 2ALR administration decreases liver
injury and cellular apoptosis in steatotic livers after IRI.
C57BL/6 mice were fed a methionine-choline-deficient diet for 2
weeks then subjected to IRI, i.e., 30 min of ischemia followed by 6
h of reperfusion. (A and B) Serum levels of the enzymes ALT and
AST. (C and D) H&E staining of liver tissue and histological
evaluation of liver damage using the Suzuki score. (E and F) TUNEL
assay of apoptosis in fatty livers after IRI and number of
TUNEL-positive cells was analyzed. (G and H) Immunohistochemical
staining of PCNA was performed, and number of PCNA-positive cells
was analyzed. Scale bars, 100 µm. *P<0.05 and
**P<0.01; n=6 per group. ALR, augmenter of liver
regeneration; IRI, ischemia-reperfusion injury; ALT, alanine
aminotransferase; AST, aspartate aminotransferase; H&E,
hematoxylin-eosin; TUNEL, terminal deoxynucleotidyl transferase
dUTP nick end labeling; NS, normal saline. |
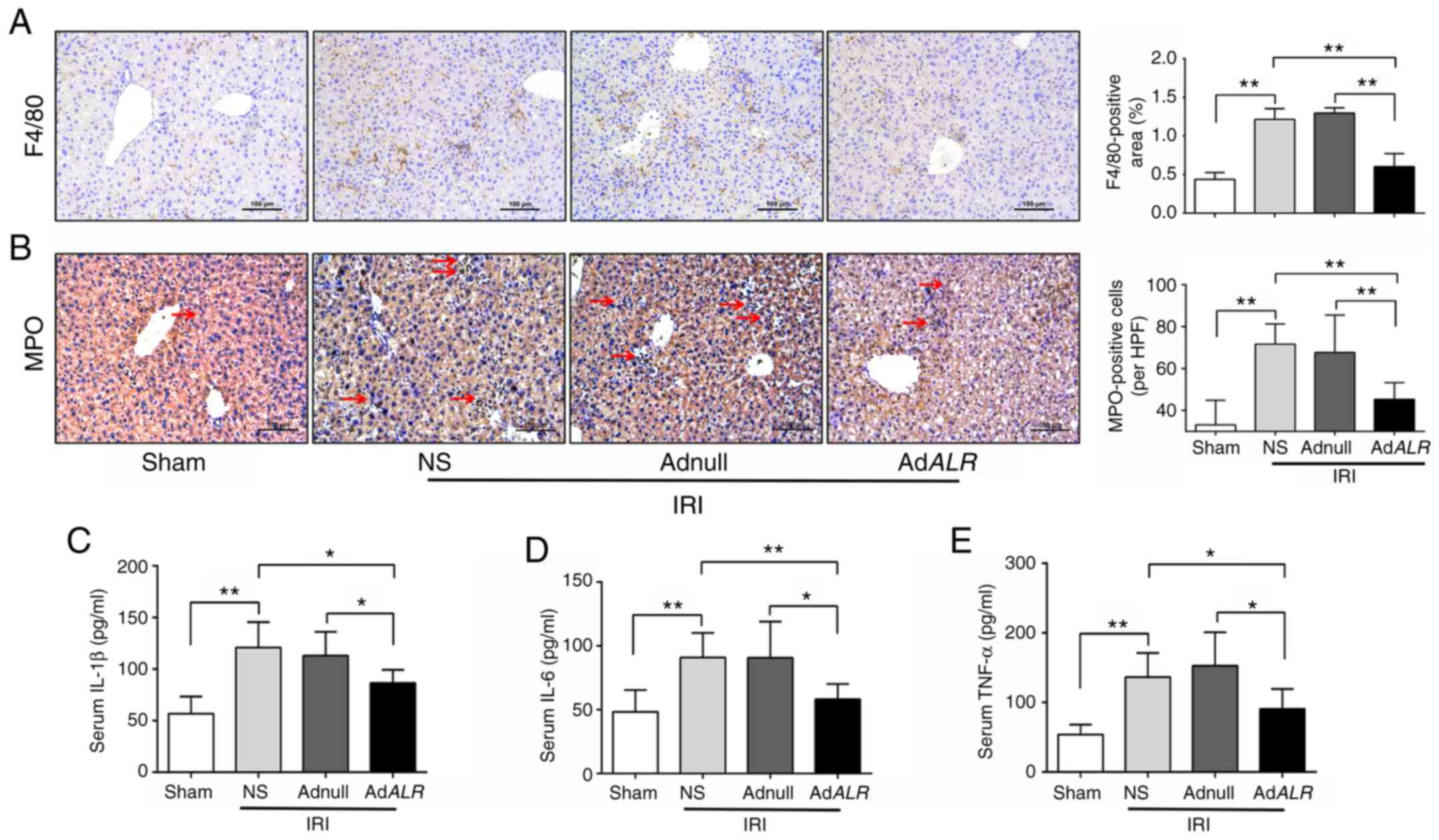
ALR expression decreases the
infiltration of inflammatory cells and the production of
inflammatory mediators
Kupffer cell (KCs) activation, as indicated by F4/80
staining, was the early inflammatory response during hepatic IRI.
The F4/80-positive area was significantly increased in the IRI
groups, and the activation of KCs was decreased in AdALR
mice compared with Adnull mice after IRI (P<0.01; Fig. 3A). MPO is abundantly expressed in
neutrophils and is used as a marker of neutrophil infiltration
(25). The number of MPO-positive
cells was increased following IRI, and neutrophil infiltration in
IRI-induced fatty livers was effectively suppressed in the
AdALR group compared with the Adnull and NS groups
(P<0.01; Fig. 3B). Inflammatory
cytokines play key roles in the pathophysiology of hepatic IRI
(26). The serum levels of IL-1β,
IL-6, and TNF-α were assessed using ELISA (Fig. 3C-E). After IRI, serum IL-1β, IL-6
and TNF-α levels were significantly increased in the IRI groups
compared with the sham-operated group but were significantly
decreased in the AdALR group compared with the Adnull and NS
groups (P<0.05).
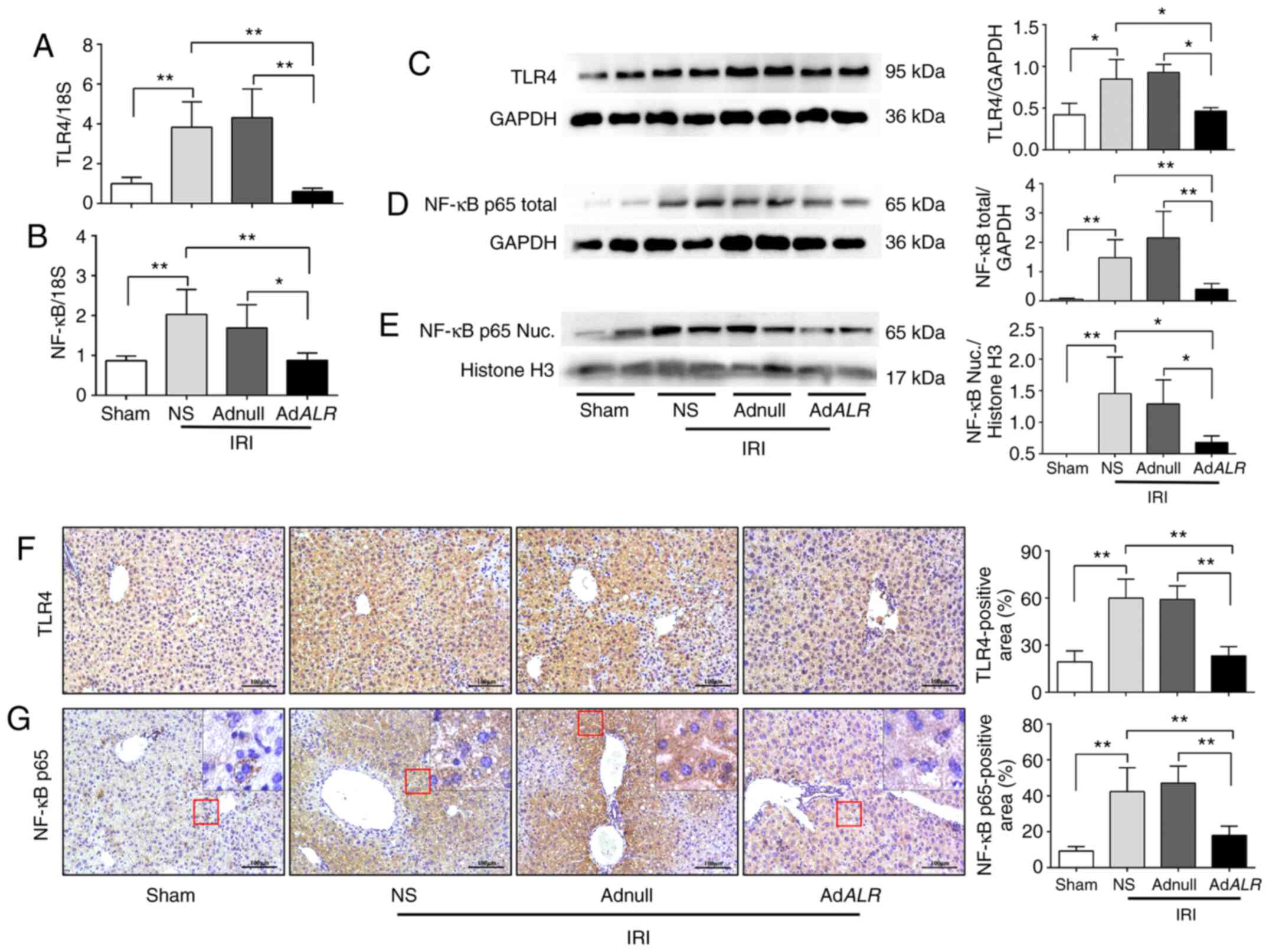
ALR modulates the TLR4/NF-κB signaling
pathway during IRI in steatotic liver
TLR4 is ubiquitously expressed in the liver, and it
initiates the sterile inflammatory response following IRI. TLR4
activation eventually leads to the activation and nuclear
translocation of NF-κB (27). To
further understand the inflammation-regulatory role of ALR, RT-qPCR
was used to observe the expression of TLR4 and NF-κB p65 in hepatic
IRI. The mRNA levels of TLR4 and NF-κB p65 were significantly
increased in the NS and Adnull groups after IRI compared with the
sham-operated group. However, the mRNA levels of TLR4 and NF-κB in
the AdALR group were significantly decreased compared with
the NS and Adnull groups (P<0.01 and P<0.05; Fig. 4A and B). ALR expression inhibited the expression
of TLR4 and NF-κB p65 after IRI, as demonstrated by western blot
analysis (P<0.05 and P<0.01, respectively; Fig. 4C and D). The nuclear fraction was extracted to
detect the nuclear distribution of NF-κB. The results revealed that
the levels of nuclear NF-κB p65 were significantly increased in the
NS and Adnull groups compared with the sham-operated group.
However, ALR transfection decreased the nuclear NF-κB p65
(P<0.05; Fig. 4E) compared with
the NS and Adnull groups. Immunohistochemistry was also used to
detect the expression of TLR4 and NF-κB during hepatic IRI. After
IRI, the expression levels of these proteins were significantly
increased in the NS and Adnull groups compared with the
sham-operated group but were significantly decreased in the
AdALR group (P<0.01; Fig.
4F and G) compared with the NS
and Adnull groups. These results indicated that ALR suppressed the
TLR4 inflammatory signaling cascade during hepatic IRI.
Discussion
The present study evaluated the effects of ALR on
the inflammatory response during fatty liver IRI. The key findings
are summarized as follows: i) ALR expression was reduced in
steatotic livers during IRI; ii) adenovirus-mediated ALR
transfection maintained liver function and alleviated IRI in
steatotic livers by suppressing inflammatory cell inflation and
reducing inflammatory cytokine production; and iii) ALR played a
regulatory role in the inflammatory response during fatty liver IRI
via inhibition of the TLR4/NF-κB signaling pathway.
Although ALR was initially reported as a mitogenic
factor that promoted liver regeneration after partial hepatectomy
in rats, substantial evidence has revealed that ALR was a survival
factor that protected the liver in the clinical context against
damaging agents, such as ethanol, carbon tetrachloride,
d-galactosamine and acetaminophen-induced liver injury (18,28).
Our previous research revealed that ALR alleviated IRI in fatty
livers by promoting mitochondrial function, reducing oxidative
stress and enhancing antioxidant activity (21). Previous studies reported that ALR
played an important role in the formation of fatty liver. Decreased
ALR expression promoted fat accumulation in the liver of mice, and
the ALR level in the serum of patients with steatohepatitis was
also significantly reduced (29,30).
However, the regulatory effects of ALR on the inflammatory response
in fatty liver IRI are not clear.
The expression of ALR after IRI in fatty livers was
initially examined and it was revealed that IRI decreased ALR
expression, which indicated that ALR depletion during fatty liver
IRI was associated with aggravated liver injury.
Adenovirus-mediated ALR transfection was used to detect the gain of
function of ALR in fatty liver IRI. Our results indicated that ALR
maintained liver function and alleviated liver injury, as indicated
by the decreased ALT and AST levels, preserved hepatic structure,
reduced apoptosis and increased hepatocyte proliferation.
Inflammation plays pivotal roles in hepatic IRI,
especially in steatotic livers (31,32).
The cytosolic components and cell fragments released from the
damaged liver trigger the production of IL-1β, IL-6 and TNF-α,
which are key proinflammatory mediators of hepatic IRI (22). KCs are liver-resident macrophages
that become activated and release inflammatory mediators during the
early phase of IRI. Neutrophils and monocytes migrate to and
accumulate in the injured liver at later phases, which contribute
to the additional, prolonged injury. An anti-inflammatory strategy
of modulating the exaggerated inflammatory response to protect the
liver against IRI was reported in numerous studies (33,34).
ALR expression reduced the serum levels of
inflammatory cytokines, such as IL-1β, IL-6 and TNF-α, in the
present study. Activation of KCs and neutrophil recruitment, as
evaluated by staining for F4/80 and MPO, respectively, were
inhibited by ALR expression in fatty liver IRI. Consistent with our
results, Khandoga et al used intravital fluorescence
microscopy and observed that ALR expression decreased the number of
rolling and firmly adherent leukocytes in post-sinusoidal venules
in normal hepatic IRI (19).
TLR4 is extensively expressed in liver cells, such
as KCs and hepatocytes (13), and
it is closely related to the development of hepatic IRI (35,36).
TLR4 triggers the activation and nuclear translocation of the
transcription factor NF-κB, which culminates in the transcription
of inflammatory cytokine genes (37). Blockade of TLR4 alleviates liver IRI
by reducing neutrophil sequestration, CD4+ T cell
infiltration, and cytokine and chemokine production (36,38).
The anti-inflammatory effects of ALR were
demonstrated previously. Yan et al reported that ALR
mitigated acute renal IRI in rats by reducing the protein
expression of NF-κB, but the mechanism was not clear (39). Pan et al demonstrated that
ALR alleviated acute pancreatitis via inhibition of the TLR4/NF-κB
pathway and reduction of the release of TNF-α and cyclooxygenase-2
(COX-2) (40). These studies
indicated that ALR is closely related to TLR4. In the present
study, it was determined whether the regulatory role of ALR in
inflammation in fatty liver IRI was mediated via the TLR4/NF-κB
pathway. Our results revealed that the TLR4/NF-κB signaling pathway
was activated in fatty liver IRI, and ALR transfection provided a
protective effect by decreasing the TLR4/NF-κB signaling
pathway.
There are some limitations in the present study.
Although this study indicated that ALR inhibited the TLR4/NF-κB
signaling pathway, whether this regulation was direct or indirect
remains unclear. The manner with which ALR regulates the expression
of TLR4 and the identity of intermediate molecules that mediate the
interaction between ALR and the TLR4/NF-κB signaling pathway are
not clear. Therefore, the regulation of ALR on the TLR4/NF-κB
signaling pathway in fatty liver IRI requires further in-depth
research.
In conclusion, the present study demonstrated that
the administration of ALR effectively alleviated hepatocyte
apoptosis and preserved liver function by suppressing inflammatory
cell infiltration and reducing inflammatory cytokine production.
ALR played a role in the inflammatory response via the inhibition
of the TLR4/NF-κB signaling pathway in fatty liver IRI. Therefore,
the anti-inflammatory effect of ALR establishes a promising
strategy to protect steatotic livers against IRI.
Supplementary Material
Primer sequences of target genes for
reverse transcription-quantitative PCR.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 31371169) and the Beijing
Tongzhou District Technology Plan (grant no. KJ2019CX014-21).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding authors on reasonable
request.
Authors' contributions
JW designed and performed the experiments. JW, BX
and WL analyzed and interpreted the data. XW established the mouse
model and performed the in vivo experiments. BX and WL
designed and supervised the study, wrote and revised the
manuscript. All authors read and approved the final manuscript. JW
and WL confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
All animal studies were approved by the Ethics
Committee of Capital Medical University (Beijing, China; approval
number AEEI-2016-094).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fondevila C, Busuttil RW and
Kupiec-Weglinski JW: Hepatic ischemia/reperfusion injury--a fresh
look. Exp Mol Pathol. 74:86–93. 2003.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhai Y, Petrowsky H, Hong JC, Busuttil RW
and Kupiec-Weglinski JW: Ischaemia-reperfusion injury in liver
transplantation - from bench to bedside. Nat Rev Gastroenterol
Hepatol. 10:79–89. 2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bzowej NH: Nonalcoholic steatohepatitis:
The new frontier for liver transplantation. Curr Opin Organ
Transplant. 23:169–174. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Besutti G, Valenti L, Ligabue G, Bassi MC,
Pattacini P, Guaraldi G and Giorgi Rossi P: Accuracy of imaging
methods for steatohepatitis diagnosis in non-alcoholic fatty liver
disease patients: A systematic review. Liver Int. 39:1521–1534.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Tashiro H, Kuroda S, Mikuriya Y and Ohdan
H: Ischemia-reperfusion injury in patients with fatty liver and the
clinical impact of steatotic liver on hepatic surgery. Surg Today.
44:1611–1625. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Maurice J and Manousou P: Non-alcoholic
fatty liver disease. Clin Med (Lond). 18:245–250. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Jiménez-Castro MB, Meroño N, Mendes-Braz
M, Gracia-Sancho J, Martínez-Carreres L, Cornide-Petronio ME,
Casillas-Ramirez A, Rodés J and Peralta C: The effect of brain
death in rat steatotic and non-steatotic liver transplantation with
previous ischemic preconditioning. J Hepatol. 62:83–91.
2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kubes P and Mehal WZ: Sterile inflammation
in the liver. Gastroenterology. 143:1158–1172. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Yang F, Shang L, Wang S, Liu Y, Ren H, Zhu
W and Shi X: TNFα-mediated necroptosis aggravates
ischemia-reperfusion injury in the fatty liver by regulating the
inflammatory response. Oxid Med Cell Longev.
2019(2301903)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Li Y, Ma D, Wang Z and Yang J:
MicroRNA-155 deficiency in Kupffer cells ameliorates liver
ischemia-reperfusion injury in mice. Transplantation.
101:1600–1608. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Schwabe RF, Seki E and Brenner DA:
Toll-like receptor signaling in the liver. Gastroenterology.
130:1886–1900. 2006.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Rao J, Qian X, Li G, Pan X, Zhang C, Zhang
F, Zhai Y, Wang X and Lu L: ATF3-mediated NRF2/HO-1 signaling
regulates TLR4 innate immune responses in mouse liver
ischemia/reperfusion injury. Am J Transplant. 15:76–87.
2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhai Y, Busuttil RW and Kupiec-Weglinski
JW: Liver ischemia and reperfusion injury: New insights into
mechanisms of innate-adaptive immune-mediated tissue inflammation.
Am J Transplant. 11:1563–1569. 2011.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lv YN, Ou-Yang AJ and Fu LS: MicroRNA-27a
negatively modulates the inflammatory response in
lipopolysaccharide-stimulated microglia by targeting TLR4 and
IRAK4. Cell Mol Neurobiol. 37:195–210. 2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Jiang X, Kuang G, Gong X, Jiang R, Xie T,
Tie H, Wu S, Wang T, Wan J and Wang B: Glycyrrhetinic acid
pretreatment attenuates liver ischemia/reperfusion injury via
inhibiting TLR4 signaling cascade in mice. Int Immunopharmacol.
76(105870)2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
LaBrecque DR and Pesch LA: Preparation and
partial characterization of hepatic regenerative stimulator
substance (SS) from rat liver. J Physiol. 248:273–284.
1975.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang J, Li Y, Jiang S, Yu H and An W:
Enhanced endoplasmic reticulum SERCA activity by overexpression of
hepatic stimulator substance gene prevents hepatic cells from ER
stress-induced apoptosis. Am J Physiol Cell Physiol. 306:C279–C290.
2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Thirunavukkarasu C, Wang LF, Harvey SA,
Watkins SC, Chaillet JR, Prelich J, Starzl TE and Gandhi CR:
Augmenter of liver regeneration: An important intracellular
survival factor for hepatocytes. J Hepatol. 48:578–588.
2008.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Khandoga A, Mende K, Iskandarov E,
Rosentreter D, Schelcher C, Reifart J, Jauch KW and Thasler WE:
Augmenter of liver regeneration attenuates inflammatory response in
the postischemic mouse liver in vivo. J Surg Res. 192:187–194.
2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Care NRCU, Animals AUOL: Guide for the
Care and Use of Laboratory Animals. National Academies Press,
Washington, DC, 2011.
|
|
21
|
Weng J, Li W, Jia X and An W: Alleviation
of Ischemia-Reperfusion Injury in Liver Steatosis by Augmenter of
Liver Regeneration Is Attributed to Antioxidation and Preservation
of Mitochondria. Transplantation. 101:2340–2348. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Olthof PB, van Golen RF, Meijer B, van
Beek AA, Bennink RJ, Verheij J, van Gulik TM and Heger M: Warm
ischemia time-dependent variation in liver damage, inflammation,
and function in hepatic ischemia/reperfusion injury. Biochim
Biophys Acta Mol Basis Dis. 1863:375–385. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Suzuki S, Toledo-Pereyra LH, Rodriguez FJ
and Cejalvo D: Neutrophil infiltration as an important factor in
liver ischemia and reperfusion injury. Modulating effects of FK506
and cyclosporine. Transplantation. 55:1265–1272. 1993.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
25
|
van der Veen BS, de Winther MP and
Heeringa P: Myeloperoxidase: Molecular mechanisms of action and
their relevance to human health and disease. Antioxid Redox Signal.
11:2899–2937. 2009.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Colletti LM, Kunkel SL, Walz A, Burdick
MD, Kunkel RG, Wilke CA and Strieter RM: The role of cytokine
networks in the local liver injury following hepatic
ischemia/reperfusion in the rat. Hepatology. 23:506–514.
1996.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Nace GW, Huang H, Klune JR, Eid RE,
Rosborough BR, Korff S, Li S, Shapiro RA, Stolz DB, Sodhi CP, et
al: Cellular-specific role of toll-like receptor 4 in hepatic
ischemia-reperfusion injury in mice. Hepatology. 58:374–387.
2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liu L, Xie P, Li W, Wu Y and An W:
Augmenter of liver regeneration protects against ethanol-induced
acute liver injury by promoting autophagy. Am J Pathol.
189:552–567. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Maehara Y and Fernandez-Checa JC:
Augmenter of liver regeneration links mitochondrial function to
steatohepatitis and hepatocellular carcinoma. Gastroenterology.
148:285–288. 2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Kumar S, Verma AK, Rani R, Sharma A, Wang
J, Shah SA, Behari J, Salazar Gonzalez R, Kohli R and Gandhi CR:
Hepatic deficiency of augmenter of liver regeneration predisposes
to nonalcoholic steatohepatitis and fibrosis. Hepatology.
72:1586–1604. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Nakamura K, Zhang M, Kageyama S, Ke B,
Fujii T, Sosa RA, Reed EF, Datta N, Zarrinpar A, Busuttil RW, et
al: Macrophage heme oxygenase-1-SIRT1-p53 axis regulates sterile
inflammation in liver ischemia-reperfusion injury. J Hepatol.
67:1232–1242. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Jimenez-Castro MB, Cornide-Petronio ME,
Gracia-Sancho J and Peralta C: Inflammasome-mediated inflammation
in liver ischemia-reperfusion injury. Cells. 8(1131)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Nakamura K, Zhang M, Kageyama S, Ke B,
Fujii T, Sosa RA, Reed EF, Datta N, Zarrinpar A, Busuttil RW, et
al: Macrophage heme oxygenase-1-SIRT1-p53 axis regulates sterile
inflammation in liver ischemia-reperfusion injury. J Hepatol.
67:1232–1242. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Motiño O, Francés DE, Casanova N,
Fuertes-Agudo M, Cucarella C, Flores JM, Vallejo-Cremades MT,
Olmedilla L, Pérez Peña J, Bañares R, et al: Protective role of
hepatocyte cyclooxygenase-2 expression against liver
ischemia-reperfusion injury in mice. Hepatology. 70:650–665.
2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ben-Ari Z, Avlas O, Fallach R,
Schmilovitz-Weiss H, Chepurko Y, Pappo O and Hochhauser E: Ischemia
and reperfusion liver injury is reduced in the absence of Toll-like
receptor 4. Cell Physiol Biochem. 30:489–498. 2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Shen XD, Ke B, Zhai Y, Gao F, Tsuchihashi
S, Lassman CR, Busuttil RW and Kupiec-Weglinski JW: Absence of
toll-like receptor 4 (TLR4) signaling in the donor organ reduces
ischemia and reperfusion injury in a murine liver transplantation
model. Liver Transpl. 13:1435–1443. 2007.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Li X: IRAK4 in TLR/IL-1R signaling:
Possible clinical applications. Eur J Immunol. 38:614–618.
2008.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Jiang N, Zhang X, Zheng X, Chen D, Zhang
Y, Siu LK, Xin HB, Li R, Zhao H, Riordan N, et al: Targeted gene
silencing of TLR4 using liposomal nanoparticles for preventing
liver ischemia reperfusion injury. Am J Transplant. 11:1835–1844.
2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Yan R, Li Y, Zhang L, Xia N, Liu Q, Sun H
and Guo H: Augmenter of liver regeneration attenuates inflammation
of renal ischemia/reperfusion injury through the NF-kappa B pathway
in rats. Int Urol Nephrol. 47:861–868. 2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Pan LF, Yu L, Wang LM, He JT, Sun JL, Wang
XB, Wang H, Bai ZH, Feng H and Pei HH: Augmenter of liver
regeneration (ALR) regulates acute pancreatitis via inhibiting
HMGB1/TLR4/NF-κB signaling pathway. Am J Transl Res. 10:402–410.
2018.PubMed/NCBI
|