Introduction
Knee osteoarthritis (KOA) is a disease characterized
by severe wear of articular cartilage and regeneration of articular
margin and subchondral bone (1).
Similar to hip OA, KOA can cause weight-bearing difficulty in the
lower limbs, eventually resulting in limb disability (2). Inflammatory response induced by
synovium tissue injury is an common cause of OA (3,4). The
main symptoms of KOA include joint pain and stiffness (5). Joint pain initially appears in joint
movement and gradually develops into persistent pain (5). The incidence and development of KOA
are associated with age, and the incidence rate is significantly
higher in women compared with men (6). According to the etiology and
pathogenesis of KOA, it is divided into two subtypes, primary and
secondary KOA (7). The pathological
process of primary KOA is gradual, and its pathogenesis remains
unclear. Secondary KOA often occurs in young adults, and common
causes include accidental trauma, deformity, insufficiency of blood
supply and joint inflammation (7).
Notably, inflammatory mediators, proteoglycans and matrix
metalloproteinases (MMPs) are all involved in the pathogenesis of
KOA (8-10).
Cyclooxygenase-2 (COX-2) is a member of the
cyclooxygenase isoenzyme family, which catalyzes the metabolism of
arachidonic acid to form prostaglandins (PGEs). PGE2, an
inflammatory mediator, can activate inositol triphosphate through
its receptors (EP1R, EP2R, EP3R or EP4R) to promote the release of
Ca2+ and inhibit Na+/Ca2+ exchange
channel to decrease Ca2+ outflow (11). Simultaneously, intracellular
Ca2+ release elevates nitrogen oxides expression, which
induces the production of reactive oxygen species (ROS) and
oxidative stress (12,13). COX-2 expression increases with the
occurrence of inflammatory reaction and oxidative stress, which in
turn increases the expression levels of the inflammatory mediators,
PGE2 and PGH2, as well as vascular endothelial growth factor, which
may be induced by PGE2(14). Thus,
the production of oxidative stress-related substances, such as
oxygen free radicals and ROS is indirectly induced, further
promoting cell damages (14,15).
In addition, COX-2 is closely associated with inflammation-related
diseases and tumors, and it is speculated that COX-2 can promote
tumor angiogenesis, tissue invasion and enhance the apoptosis
resistance of tumor cells (16-18).
However, the regulation of COX-2 remains unclear.
MicroRNA (miRNA/miR)-758 has been demonstrated to
play important regulatory roles in cell metabolism, cholesterol
outflow, atheromatous plaque formation and hepatitis C virus (HCV)
infection (19). For example,
miR-758-3p can regulate cholesterol efflux by inhibiting ABCA1
protein expression (20).
Furthermore, Mandolini et al (21) reported that miR-758-3p regulates
ABCA1 expression in human atherosclerotic plaques. Yang et
al (19) demonstrated that
miR-758-3p expression is significantly upregulated in HCV
infection. In addition, abnormal miR-758-3p expression is
associated with tumor progression (22). During chemotherapy for esophageal
cancer, chemotherapeutic drugs can inhibit miR-758-3p expression,
and the loss of control of miR-758-3p to its downstream target
genes may be the reason for drug resistance of tumors (23). The bioinformatics of the present
study predicted that miR-758-3p has the potential to regulate COX-2
expression. However, the underlying molecular mechanism by which
miR-758-3p regulates COX-2 has not yet been investigated.
The present study aimed to investigate COX-2 mRNA
and protein expression levels in synovium tissues and synovial
fluid from patients with KOA, and determine the underlying
molecular mechanism by which miR-758-3p regulates KOA via
COX-2.
Materials and methods
Patients
A total of 37 patients with KOA who received
treatment at Heze Municipal Hospital between December 2017 and July
2019 were included in the experimental group (20 men and 17 women;
age range, 30-60 years; median age, 47.6 years). A total of 29
patients with acute knee trauma who received treatment at Heze
Municipal Hospital during the same period were included in the
control group (17 men and 12 women; age range, 28-58 years; median
age, 46.9 years). All patients with KOA met the 2010 EULAR
recommendations (24) for
diagnosis. Patients with cancer, rheumatoid arthritis and other
immune diseases were excluded from the present study. Synovium
tissue samples were collected from patients with KOA during knee
replacement or knee arthroscopy, while synovium tissues were
collected from patients with knee trauma by resection during
surgery. Part of the synovium was used to culture primary cells,
while part was used for detection. Synovial fluid was respectively
collected from patients with KOA and knee trauma during surgery,
and blood cells were removed via centrifugation at 1,000 x g at 4˚C
for 10 min. All samples were stored at -80˚C until subsequent
experimentation. The present study was approved by the Ethics
Committee of Heze Municipal Hospital (Heze, China; approval no.
20161226-85) and written informed consent was provided by all
patients or their families prior to the study start.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from fluid and tissue
samples using TRIzol® reagent (cat. no. R0016; Beyotime
Institute of Biotechnology), according to the manufacturer's
instructions. The purity of RNA was assessed by measuring A260/A280
on a reader. According to the manufacturer's protocol, mRNA or
miRNA (2 µg) was reverse-transcribed into cDNA at 42˚C using the
TIAN Script Ⅱ cDNA First Strand Synthetic kit and miRcute miRNA
cDNA First Strand Synthetic kit, respectively (both purchased from
Tiangen Biotech Co., Ltd.). The following primer sequences were
used for qPCR using SYBR-Green (cat. no. Q121-02; Vazyme Biotech
Co., Ltd.): COX-2 forward, 5'- CAGCCATACAGCAAATCCTTG-3' and
reverse, 5'- CAAATGTGATCTGGATGTCAAC-3'; GAPDH forward,
5'-TGACCTTGCCCACAGCCTTG-3' and reverse,
5'-CATCACCATCTTCCAGGAGCG-3'; miR-758-3p forward,
5'-ACACTCCAGCTGGGTTTGTGACCTGGTCCA-3' and reverse,
5'-TGGTGTCGTGGAGTCG-3'; and U6 forward, 5'-CTCGCTTCGGCAGCACA-3' and
reverse, 5'-AACGCTTCACGAATTTGCGT-3'. The following thermocycling
conditions were used for qPCR: Initial denaturation at 95˚C for 5
min; denaturation at 95˚C for 30 sec, annealing at 55˚C for 30 sec
and elongation at 72˚C for 60 sec (35 cycles); final extension at
72˚C for 2 min. Relative expression levels were calculated using
the 2-ΔΔCq method (25).
If the Cq value was too high (>35), the dose was adjusted, and
the experiment was repeated to increase reliable. All experiments
were performed in triplicate.
Cell culture
293T cells were purchased from The Cell Bank of Type
Culture Collection of the Chinese Academy of Sciences. Cells were
maintained in DMEM/F-12 medium (Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS; Thermo Fisher
Scientific, Inc.), at 37˚C with 5% CO2.
Prior to isolating KOA synovial cells, synovium
tissues were washed with PBS containing 2% penicillin-streptomycin
(Thermo Fisher Scientific, Inc.). Tissue samples were cut into
1-mm3-thick sections and digested using 4 mg/ml type II
collagenase (Thermo Fisher Scientific, Inc.) at 37˚C for 4 h. The
solution was subsequently diluted with PBS, filtered through
200-mesh sterile sieves and centrifuged at 37˚C and 800 x g for 5
min. Cells were resuspended in DMEM supplemented with 20% FBS and
1% penicillin-streptomycin at 37˚C with 5% CO2. Synovial
cells are generally stable within 5 passages (26), thus cells at passages 3-7 were used
for subsequent experimentation.
Cells at the logarithmic phase (3x105)
were seeded into 24-well plates and maintained in antibiotic-free
DMEM/F-12 medium supplemented with 10% FBS 24 h prior to
transfection, until they reached 70% confluence. In the first vial,
1 µl miR-NC (20 pmol/µl; forward, 5'-UUCUCCGAACGUGUCACGUTT-3' and
reverse, 5'-ACGUGACACGUUCGGAGAATT-3') or miR-758-3p mimic (20
pmol/µl; forward, 5'-UUUGUGACCUGGUCCACUAACC-3' and reverse,
5'-UUAGUGGACCAGGUCACAAAUU-3') was added to 50 µl
Opti-MEM® medium (Thermo Fisher Scientific, Inc.). In
the second vial, 1 µl Lipofectamine® 2,000 reagent
(Thermo Fisher Scientific, Inc.) was added to 50 µl
Opti-MEM® medium. Following incubation at 37˚C for 5
min, the vials were combined and incubated for a further 20 min at
room temperature. The mixture was subsequently added to each well
and incubated at 37˚C for 6 h. The medium was replaced with
DMEM/F-12 medium supplemented with 10% FBS. Cells were collected
for subsequent experimentation after 48 h.
Western blotting
Total protein was extracted from tissues or cells
using E.Z.N.A total DNA/RNA/Protein kit (Omega Bio-Tek, Inc.).
Total protein was quantified using the BCA Protein Assay kit (cat.
no. ab102536; Abcam) and 50 µg protein/lane was separated by
SDS-PAGE on a 10% gel electrophoresis at 100 V. The separated
proteins were subsequently transferred onto polyvinylidene
difluoride membranes on ice (100 V for 2 h), and blocked with 5%
skimmed milk at room temperature for 1 h. The membranes were
incubated with rabbit anti-human COX-2 (1:1,000; cat. no. ab15191;
Abcam) and β-actin (1:5,000; cat. no. ab129348; Abcam) polyclonal
primary antibodies, overnight at 4˚C. Membranes were washed three
times with PBS containing Tween-20 (0.1%) for 15 min, and
subsequently incubated with goat anti-rabbit horseradish
peroxidase-conjugated secondary antibody (1:3,000; cat. no. ab6721;
Abcam) for 1 h at room temperature. Membranes were re-washed three
times with PBS containing Tween-20 (0.1%) for 15 min. Protein bands
were visualized using the enhanced chemiluminescence detection kit
(cat. no. ab65623; Abcam) and analyzed using Image lab v3.0
software (Bio-Rad Laboratories, Inc.).
ELISA
Synovial fluid was centrifuged at 1,000 x g at 4˚C
for 10 min and the supernatant was collected, which was used to
detect protein contents according to the manufacturer's protocols
(COX-2 ELISA kit; cat. no. ab267646; Abcam and Ca PGE2 ELISA kit;
cat. no. CSB-E07965h; Cusabio Technology LLC).
Bioinformatics analysis
The miRanda database (http://www.microrna.org/microrna/home.do) was used to
predict the genes that regulate COX-2 expression.
Dual-luciferase reporter assay
Based on bioinformatics analysis, wild-type (WT) and
mutant regions of miR-758-3p in the 3'-UTR of COX-2 mRNA were
chemically synthesized in vitro. The ends were joined with
Spe-1 and HindIII restriction sites, and subsequently cloned into
pMIR-REPORT luciferase reporter plasmids (Thermo Fisher Scientific,
Inc.). Plasmids (1 µg) with WT or mutant 3'-UTR sequences were
co-transfected into 293T cells with agomiR-758-3p (100 nM; forward,
5'-UUUGUGACCUGGUCCACUAACC-3' and reverse,
5'-UUAGUGGACCAGGUCACAAAUU-3') or agomiR-negative control
(agomiR-NC; 100 nM; forward, 5'-UUCUCCGAACGUGUCACGUTT-3' and
reverse, 5'-ACGUGACACGUUCGGAGAATT-3'; both purchased from Sangon
Biotech Co., Ltd.) using Exfect Transfection Reagent (Vazyme
Biotech Co., Ltd.). Following incubation at 37˚C for 24 h, cells
were lysed using the dual-luciferase reporter assay kit (cat. no.
E1980; Promega Corporation), according to the manufacturer's
instructions, and luciferase activities were detected using the
GloMax 20/20 luminometer (Promega Corporation). Firefly luciferase
activity was normalized to Renilla luciferase activity.
MTT assay
Cell proliferation was assessed via the MTT assay.
Following transfection, cells were seeded into 96-well plates at a
density of 2x103 cells/well. Each condition was assessed
in triplicate wells. At 24, 48 and 72 h post-transfection, 20 µl
MTT reagent (5 g/l; cat. no. C0009; Beyotime Institute of
Biotechnology) was added to each well and incubated for 4 h at
37˚C. Following the MTT incubation, the purple formazan crystals
were dissolved using DMSO (150 µl/well) and proliferation was
subsequently analyzed at a wavelength of 490 nm, using a microplate
reader (Bio-Rad Laboratories, Inc.).
Statistical analysis
Statistical analysis was performed using SPSS 18.0
software (SPSS, Inc.). Each experiment was repeated three times.
Data are presented as the mean ± standard deviation. Unpaired
Student's t-test was used to compare differences between two
groups, while one-way ANOVA followed by Bonferroni post hoc test
were used to compare differences between multiple groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
COX-2 expression is higher in patients
with KOA than those with acute knee trauma
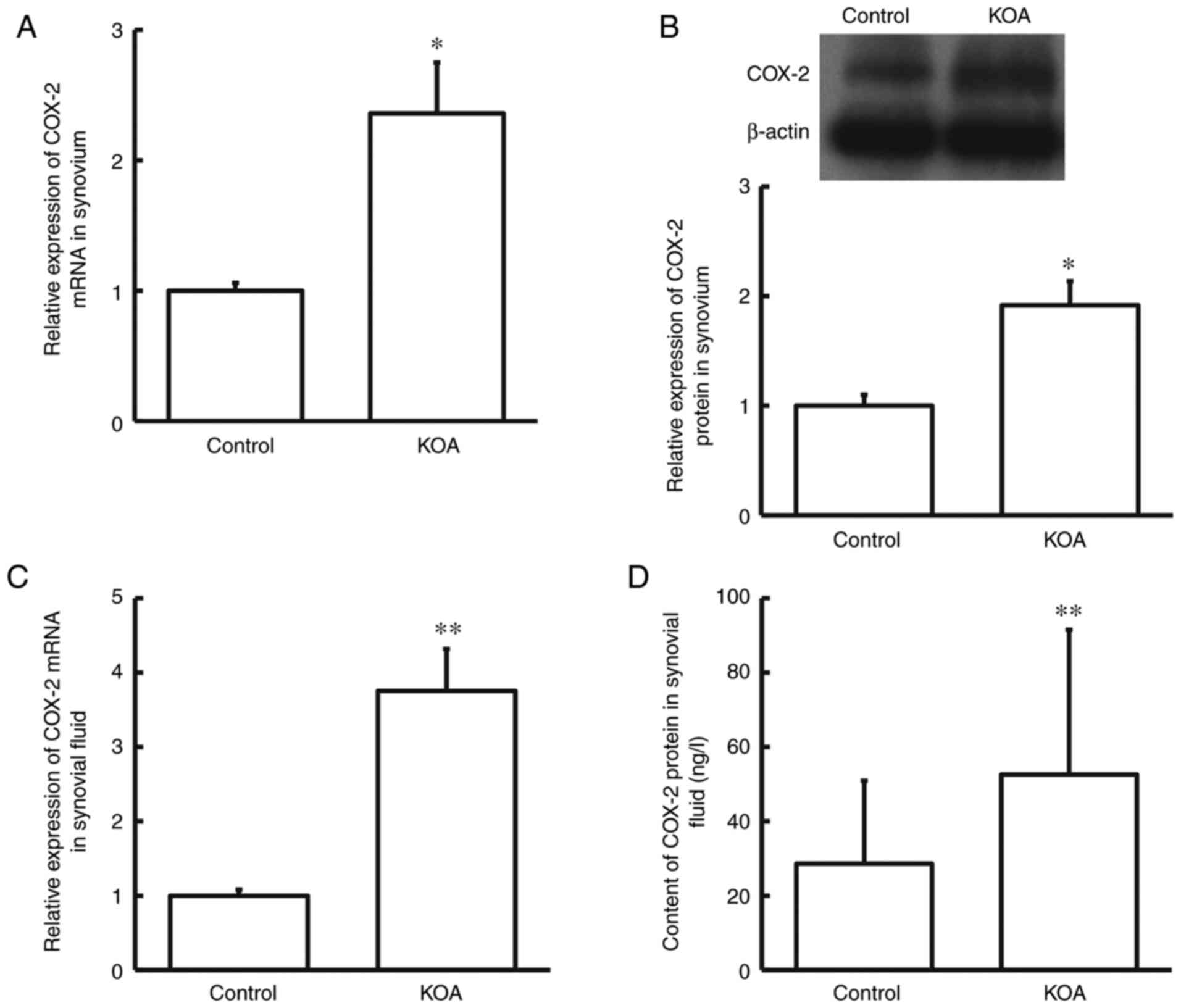
RT-qPCR analysis was performed to detect COX-2 mRNA
expression in synovium tissues and synovial fluid. In addition,
western blotting and ELISA were performed to detect COX-2 protein
expression in synovium tissues and synovial fluid, respectively.
The results demonstrated that COX-2 mRNA and protein expression
levels were significantly higher in synovium tissues from patients
with KOA compared with the control group (P<0.05; Fig. 1A and B). Similarly, COX-2 mRNA and protein
expression levels were significantly higher in synovial fluid from
patients with KOA compared with the control group (P<0.01;
Fig. 1C and D). Taken together, these results suggest
that COX-2 may play a regulatory role in the pathological process
of KOA.
miR-758-3p expression is lower in
patients with KOA than those with acute knee trauma
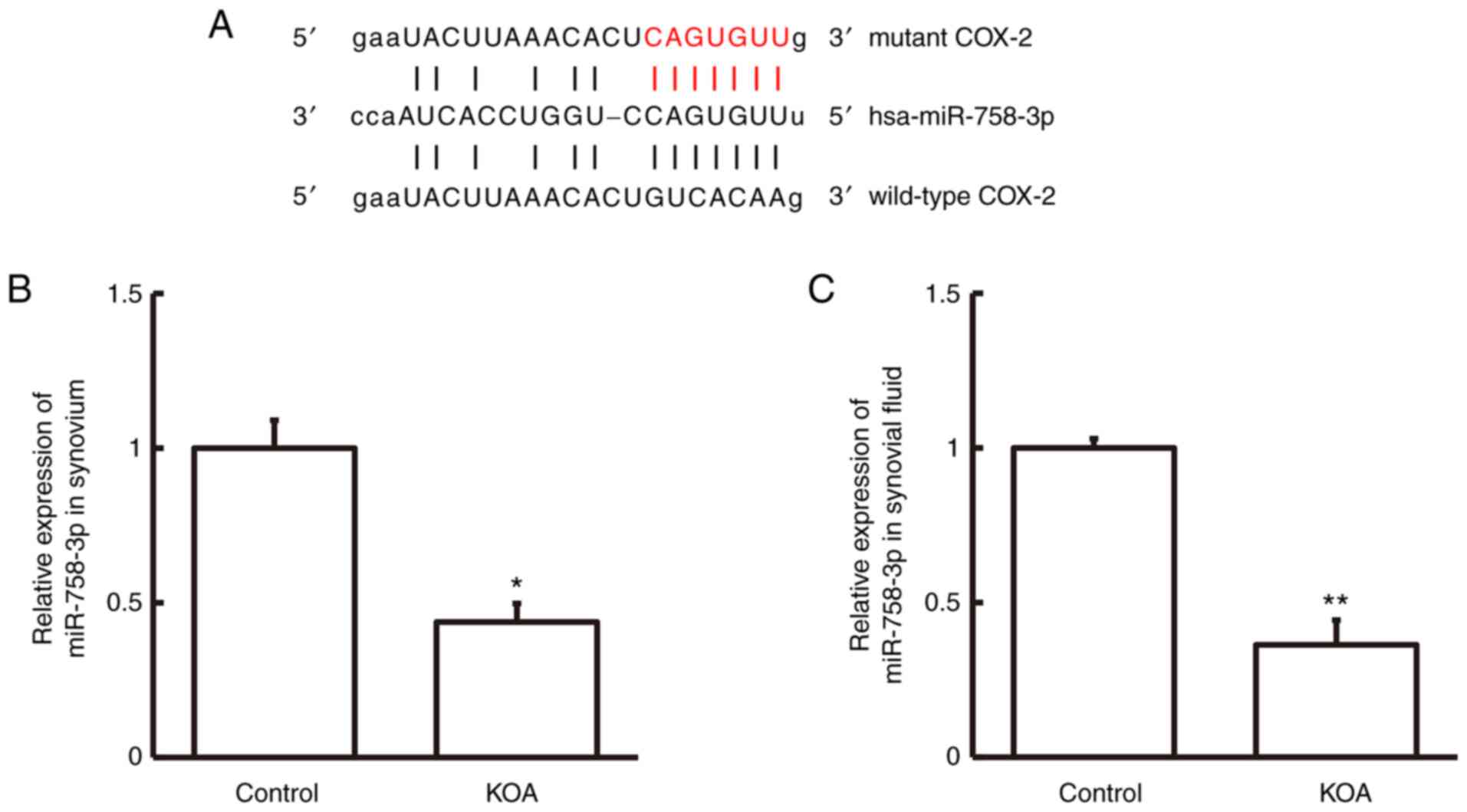
Bioinformatics analysis was performed to predict the
genes that regulate COX-2 expression. The results demonstrated that
miR-758-3p can potentially interact with COX-2 (Fig. 2A). RT-qPCR analysis was performed to
detect miR-758-3p expression in synovium tissues and synovial
fluid. The results demonstrated that miR-758-3p expression was
significantly lower in synovium tissues and synovial fluid from
patients with KOA compared with the control group (P<0.05 and
P<0.01; Fig. 2B and C, respectively). Collectively, these
results suggest that miR-758-3p expression is downregulated in
patients with KOA.
miR-758-3p interacts with the 3'-UTR
of COX-2 to regulate its expression
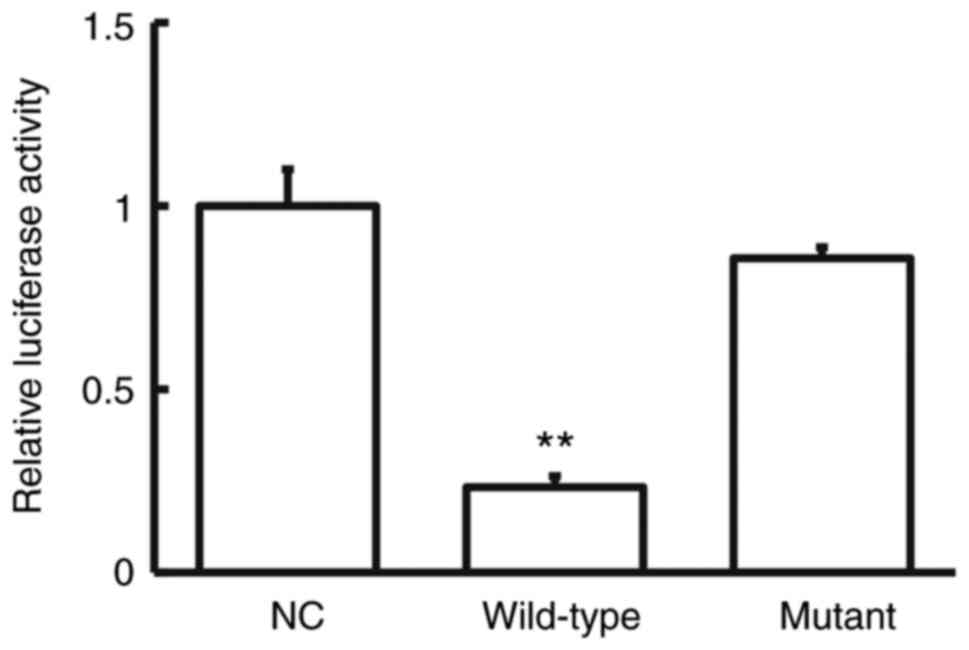
The dual-luciferase reporter assay was performed to
confirm the interaction between COX-2 mRNA and miR-758-3p. As
presented in Fig. 3, the luciferase
activity of 293T cells co-transfected with agomiR-758-3p and
pMIR-REPORT was significantly lower in the wild-type group compared
with the NC group (P<0.05). Conversely, no significant
difference was observed in the luciferase activity of cells
co-transfected with agomiR-758-3p and pMIR-REPORT between the
mutant and NC groups (P>0.05; Fig.
3). Taken together, these results suggest that miR-758-3p
interacts with the 3'-UTR of COX-2 to regulate its expression.
Overexpression of miR-758-3p inhibits
the expression and release of COX-2, as well as the proliferation
of human KOA synovial cells
To investigate how miR-758-3p regulates COX-2
expression, KOA synovial cells were transfected with agomiR-758-3p.
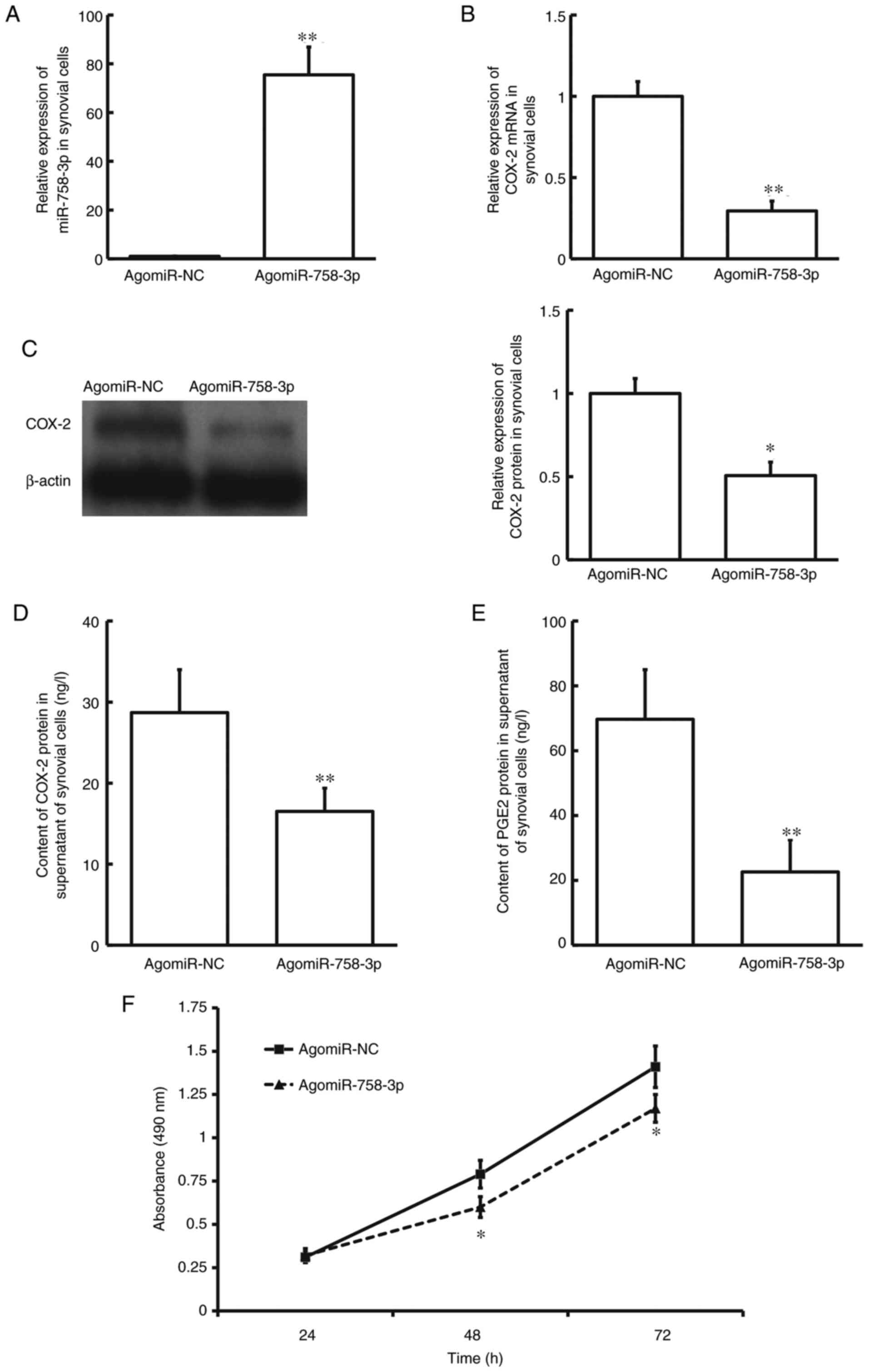
The results demonstrated that miR-758-3p expression was
significantly higher in cells transfected with agomiR-758-3p
compared with the agomiR-NC group (P<0.01; Fig. 4A). Conversely, COX-2 mRNA and
protein expression levels were significantly lower in the
agomiR-758-3p group compared with the agomiR-NC group (P<0.01
and P<0.05; Fig. 4B and C, respectively). ELISA indicated that
COX-2 protein expression was significantly lower in the supernatant
of cells transfected with agomiR-758-3p compared with the agomiR-NC
group (P<0.01; Fig. 4D). COX-2
is also known as prostaglandin-endoperoxide synthase-2 (PTGS-2).
PGE2, which is biosynthesized from arachidonic acid by COXs
(PTGSs), is a physiologically active lipid (27). Notably, PGE2 protein expression was
significantly lower in the supernatant of cells transfected with
agomiR-758-3p compared with the agomiR-NC group (P<0.01;
Fig. 4E). The results of the MTT
assay demonstrated that cells transfected with agomiR-758-3p
proliferated at a slower rate at 48 and 72 h compared with the
agomiR-NC group (P<0.05; Fig.
4F). Collectively, these results suggest that overexpression of
miR-758-3p inhibits the expression and release of COX-2, as well as
the proliferation of human KOA synovial cells.
Discussion
KOA is a chronic degenerative disease of the bone
and joints. The main pathological changes of KOA are articular
cartilage degeneration and hyperosteogeny (28). The main clinical symptoms of KOA
include pain, swelling, limited movement, friction sound and
deformity of knee joints (5). The
incidence of KOA continues to increase with age and obesity, and
thus has attracted great interest in clinical practice (29).
Currently, the treatment strategies for KOA lack
efficiency. Western medicine is classified into drug treatment and
surgical treatment (30). COX-2 is
a membrane binding protein, and its expression in cartilages is
elevated following impacts by physical and chemical factors, such
as trauma, inflammatory mediators and cytokines (31). COX-2 can promote the synthesis of
PGE2, which induces the production of nitric oxide (32). PGE2 is a multifunctional
inflammatory mediator that increases the production of MMPs and
catabolic substances, affects the structure and function of the
joint, and accelerates the destruction of joint structure (33). Thus, it can be used as a target for
KOA therapy.
The results of the present study demonstrated that
COX-2 expression was abnormally high in synovium tissues and
synovial fluid from patients with KOA, suggesting that COX-2 plays
an important biological role in KOA. However, the regulation of
COX-2 in KOA remains unclear. Bioinformatics analysis revealed that
miR-758-3p regulates COX-2 expression. miRNAs regulate mRNAs by
inhibiting translation (34). It
has been reported that miR-758-3p can target HMGB3 and inhibit the
proliferation and metastasis of cervical cancer cells via the
Wnt/β-catenin signaling pathway (35). Similarly, miR-758-3p targets HMGA1
to inhibit the malignant phenotype of osteosarcoma via the
Wnt/β-catenin signaling pathway (36). miR-758-3p also inhibits the
malignant progression of retinoblastoma by directly targeting
PAX6(37). In addition, miR-758-3p
also inhibits lung cancer cells (38,39).
Based on these findings, it was speculated that miR-758-3p is
closely associated with inflammation.
The results of the present study demonstrated that
miR-758-3p expression was significantly lower in synovium tissues
and synovial fluid from patients with KOA compared with the control
group. Considering the abnormally high COX-2 expression, it was
speculated that downregulated miR-758-3p expression induces
upregulated COX-2 expression, which was confirmed via the
dual-luciferase reporter assay. Synovial cells were subsequently
transfected with agomiR-758-3p and the MTT assay was performed to
assess cell proliferation. The results demonstrated that
overexpression of miR-758-3p decreased COX-2 mRNA and protein
expression levels, as well as the secretion of COX-2. In addition,
overexpression of miR-758-3p decreased the proliferative ability of
the cells.
The present study has a limitation in the small
number of patients, which will be overcome in the future by
performing studies with a larger number of patients. COX-2 is an
inducible enzyme of inflammatory cytokines, including interleukin
(IL)-1 and tumor necrosis factor-α (TNF-α). Notably, IL-1 and TNF-α
have been detected in synovial fluid from patients with KOA
(40). However, the present study
failed to investigate whether COX-2 expression was affected by IL-1
or TNF-α, in addition to miR-758-3p.
In conclusion, the results of the present study
demonstrated that miR-758-3p regulates COX-2 transcription factor
by directly targeting and changing COX-2 protein expression, which
in turn affects the proliferation of synovial cells. Taken
together, these results suggest that miR-758-3p, in joint fluid,
may be used as a diagnostic biomarker for patients with KOA.
Acknowledgements
The authors of the present study would like to thank
Dr Bo Zhang from Heze Municipal Hospital (Heze, China) for
providing suggestions and instructions.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL and XH designed the present study. ZL, JS and TL
performed the experiments. ZL, JS and XH analyzed the data. ZL and
XH interpreted the results and drafted the initial manuscript. ZL,
JS and XH confirm the authenticity of all the raw data. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Heze Municipal Hospital (approval no. 20161226-85;
Heze, China) and written informed consent was provided by all
patients or their families prior to the study start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Szychlinska MA, Trovato FM, Di Rosa M,
Malaguarnera L, Puzzo L, Leonardi R, Castrogiovanni P and Musumeci
G: Co-expression and co-localization of cartilage glycoproteins
CHI3L1 and lubricin in osteoarthritic cartilage: Morphological,
immunohistochemical and gene expression profiles. Int J Mol Sci.
17(359)2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Musumeci G: Effects of exercise on
physical limitations and fatigue in rheumatic diseases. World J
Orthop. 6:762–769. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Castrogiovanni P, Di Rosa M, Ravalli S,
Castorina A, Guglielmino C, Imbesi R, Vecchio M, Drago F,
Szychlinska MA and Musumeci G: Moderate physical activity as a
prevention method for knee osteoarthritis and the role of
synoviocytes as biological key. Int J Mol Sci.
20(511)2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Di Rosa M, Castrogiovanni P and Musumeci
G: The synovium theory: Can exercise prevent knee osteoarthritis?
The role of ̔mechanokines’, a possible biological key. J Funct
Morphol Kinesiol. 4(11)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Abbasi J: Can exercise prevent knee
osteoarthritis? JAMA. 318:2169–2171. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Koh YG, Jo SB, Kwon OR, Suh DS, Lee SW,
Park SH and Choi YJ: Mesenchymal stem cell injections improve
symptoms of knee osteoarthritis. Arthroscopy. 29:748–755.
2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yuan C, Pan Z, Zhao K, Li J, Sheng Z, Yao
X, Liu H, Zhang X, Yang Y, Yu D, et al: Classification of four
distinct osteoarthritis subtypes with a knee joint tissue
transcriptome atlas. Bone Res. 8(38)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Geng R, Xu Y, Hu W and Zhao H: The
association between MMP-1 gene rs1799750 polymorphism and knee
osteoarthritis risk. Biosci Rep. 38(BSR20181257)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Matsuzaki T, Alvarez-Garcia O, Mokuda S,
Nagira K, Olmer M, Gamini R, Miyata K, Akasaki Y, Su AI, Asahara H
and Lotz MK: FoxO transcription factors modulate autophagy and
proteoglycan 4 in cartilage homeostasis and osteoarthritis. Sci
Transl Med. 10(eaan0746)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Abramson SB: Nitric oxide in inflammation
and pain associated with osteoarthritis. Arthritis Res Ther. 10
(Suppl 2)(S2)2008.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Shi H, Sun X, Kong A, Ma H, Xie Y, Cheng
D, Wong CKC, Zhou Y and Gu J: Cadmium induces
epithelial-mesenchymal transition and migration of renal cancer
cells by increasing PGE2 through a cAMP/PKA-COX2 dependent
mechanism. Ecotoxicol Environ Saf. 207(111480)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Riba A, Deres L, Sumegi B, Toth K,
Szabados E and Halmosi R: Cardioprotective effect of resveratrol in
a postinfarction heart failure model. Oxid Med Cell Longev.
2017(6819281)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ogata S, Kubota Y, Satoh S, Ito S,
Takeuchi H, Ashizuka M and Shirasuna K: Ca2+ stimulates
COX-2 expression through calcium-sensing receptor in fibroblasts.
Biochem Biophys Res Commun. 351:808–814. 2006.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Salvado MD, Alfranca A, Escolano A,
Haeggström JZ and Redondo JM: COX-2 limits prostanoid production in
activated HUVECs and is a source of PGH2 for transcellular
metabolism to PGE2 by tumor cells. Arterioscler Thromb Vasc Biol.
29:1131–1137. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Cianchi F, Cortesini C, Schiavone N, Perna
F, Magnelli L, Fanti E, Bani D, Messerini L, Fabbroni V, Perigli G,
et al: The role of cyclooxygenase-2 in mediating the effects of
histamine on cell proliferation and vascular endothelial growth
factor production in colorectal cancer. Clin Cancer Res.
11:6807–6815. 2005.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zheng Y, Comaills V, Burr R, Boulay G,
Miyamoto DT, Wittner BS, Emmons E, Sil S, Koulopoulos MW, Broderick
KT, et al: COX-2 mediates tumor-stromal prolactin signaling to
initiate tumorigenesis. Proc Natl Acad Sci USA. 116:5223–5232.
2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Hashemi Goradel N, Najafi M, Salehi E,
Farhood B and Mortezaee K: Cyclooxygenase-2 in cancer: A review. J
Cell Physio. 234:5683–5699. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yu T, Lao X and Zheng H: Influencing COX-2
activity by COX related pathways in inflammation and cancer. Mini
Rev Med Chem. 16:1230–1243. 2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yang Q, Fu S and Wang J: Hepatitis C virus
infection decreases the expression of Toll-like receptors 3 and 7
via upregulation of miR-758. Arch Virol. 159:2997–3003.
2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Yao Y, Li Q, Gao P, Wang W, Chen L, Zhang
J and Xu Y: Glucagon-like peptide-1 contributes to increases ABCA1
expression by downregulating miR-758 to regulate cholesterol
homeostasis. Biochem Biophys Res Commun. 497:652–658.
2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Mandolini C, Santovito D, Marcantonio P,
Buttitta F, Bucci M, Ucchino S, Mezzetti A and Cipollone F:
Identification of microRNAs 758 and 33b as potential modulators of
ABCA1 expression in human atherosclerotic plaques. Nutr Metab
Cardiovasc Dis. 25:202–209. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chen J, Xu Z, Yu C, Wu Z, Yin Z, Fang F
and Chen B: miR-758-3p regulates papillary thyroid cancer cell
proliferation and migration by targeting TAB1. Pharmazie.
74:235–238. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Hummel R, Wang T, Watson DI, Michael MZ,
Van der Hoek M, Haier J and Hussey DJ: Chemotherapy-induced
modification of microRNA expression in esophageal cancer. Oncol
Rep. 26:1011–1017. 2011.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Carrión-Barberà I, Salman-Monte TC,
Vílchez-Oya F and Monfort J: Neuropsychiatric involvement in
systemic lupus erythematosus: A review. Autoimmun Rev.
20(102780)2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Manferdini C, Paolella F, Gabusi E,
Silvestri Y, Gambari L, Cattini L, Filardo G, Fleury-Cappellesso S
and Lisignoli G: From osteoarthritic synovium to synovial-derived
cells characterization: Synovial macrophages are key effector
cells. Arthritis Res Ther. 18(83)2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Joseph T, Zalenskaya IA, Sawyer LC,
Chandra N and Doncel GF: Seminal plasma induces
prostaglandin-endoperoxide synthase (PTGS) 2 expression in
immortalized human vaginal cells: Involvement of semen
prostaglandin E2 in PTGS2 upregulation. Biol Reprod.
88(13)2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wu Y, Lin Z, Yan Z, Wang Z, Fu X and Yu K:
Sinomenine contributes to the inhibition of the inflammatory
response and the improvement of osteoarthritis in mouse-cartilage
cells by acting on the Nrf2/HO-1 and NF-κB signaling pathways. Int
Immunopharmacol. 75(105715)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Roos EM and Arden NK: Strategies for the
prevention of knee osteoarthritis. Nat Rev Rheumatol. 12:92–101.
2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Price AJ, Alvand A, Troelsen A, Katz JN,
Hooper G, Gray A, Carr A and Beard D: Knee replacement. Lancet.
392:1672–1682. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Amin AR, Dave M, Attur M and Abramson SB:
COX-2, NO, and cartilage damage and repair. Curr Rheumatol Rep.
2:447–453. 2000.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Tsuge K, Inazumi T, Shimamoto A and
Sugimoto Y: Molecular mechanisms underlying prostaglandin
E2-exacerbated inflammation and immune diseases. Int Immunol.
31:597–606. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Echizen K, Hirose O, Maeda Y and Oshima M:
Inflammation in gastric cancer: Interplay of the
COX-2/prostaglandin E2 and Toll-like receptor/MyD88 pathways.
Cancer Sci. 107:391–397. 2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Fabian MR, Sonenberg N and Filipowicz W:
Regulation of mRNA translation and stability by microRNAs. Annu Rev
Biochem. 79:351–379. 2010.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Song T, Hou X and Lin B: MicroRNA-758
inhibits cervical cancer cell proliferation and metastasis by
targeting HMGB3 through the WNT/β-catenin signaling pathway. Oncol
Lett. 18:1786–1792. 2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Ren J, Yang M, Xu F and Chen J:
MicroRNA-758 inhibits the malignant phenotype of osteosarcoma cells
by directly targeting HMGA1 and deactivating the Wnt/β-catenin
pathway. Am J Cancer Res. 9:36–52. 2019.PubMed/NCBI
|
|
37
|
Li J and You X: MicroRNA-758 inhibits
malignant progression of retinoblastoma by directly targeting PAX6.
Oncol Rep. 40:1777–1786. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zhang Y and Zhao F: MicroRNA-758 inhibits
tumorous behavior in tongue squamous cell carcinoma by directly
targeting metadherin. Mol Med Rep. 19:1883–1890. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhou GH, Lu YY, Xie JL, Gao ZK, Wu XB, Yao
WS and Gu WG: Overexpression of miR-758 inhibited proliferation,
migration, invasion, and promoted apoptosis of non-small cell lung
cancer cells by negatively regulating HMGB. Biosci Rep.
39(BSR20180855)2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Lin Z, Fu C, Yan Z, Wu Y, Zhan J, Lou Z,
Liao X and Pan J: The protective effect of hesperetin in
osteoarthritis: An in vitro and in vivo study. Food Funct.
11:2654–2666. 2020.PubMed/NCBI View Article : Google Scholar
|