Introduction
Diabetic nephropathy (DN) is a common complication
of diabetes and is the main cause of end-stage renal disease in
China and worldwide (1). DN is
characterized by persistent proteinuria, increase of protein
glomerular filtration and abnormal renal function. The early
pathological changes include the accumulation of extracellular
matrix (ECM) components and renal hypertrophy and the late
pathological changes comprise the development of glomerulosclerosis
and tubulointerstitial fibrosis (2-4).
The abnormal proliferation of mesangial cells (MCs) may partially
account for the increase of mesangial matrix and glomerulosclerosis
(5,6). Therefore, reducing the damage of
mesangial cells may serve a notable role during the treatment of
DN.
A number of studies have indicated that oxidative
stress plays a considerable role in the occurrence and development
of DN (6,7). Recent studies have demonstrated that
the stimulation with various factors, such as high glucose (HG),
hydrogen peroxide, angiotensin II, aldosterone and inflammation
leads to an increase of oxidative stress in MCs, thereby promoting
the occurrence of glomerulosclerosis (8,9).
Therefore, decreasing or terminating oxidative stress injury of
glomerular mesangial cells could improve early diabetic
nephropathy. Hyperglycemia induces the increase of reactive oxygen
species (ROS) production in glomerular mesangial cells, enhances
the activation of transcriptional activator protein-1 (AP-1) and
NF-κB and promotes the expression of inflammatory factors, such as
intercellular adhesion molecule and monocyte chemoattractant
protein-1, thereby aggravating the local inflammatory response and
oxidative stress (10). It was
hypothesized that angiotensin is an important mediator of oxidative
stress and angiotensin receptor blockers could reduce the level of
oxidative stress in patients with diabetes and alleviate renal
injury (11).
Ellagic acid (EA) is a natural compound that exists
in various soft fruits, nuts and other plant tissues. It has been
demonstrated that EA has antioxidant, anticancer,
anti-atherosclerotic and anti-inflammatory effects in multiple
diseases, including ulcerative colitis, colon carcinoma,
hepatopathy, Alzheimer's disease and Diabetes (12-14).
EA has been indicated to inhibit attenuated renal dysfunction and
oxidative stress in high fat diet/low dose streptozotocin
(HFD/STZ)-induced type 2 diabetic Wistar albino rats by inhibiting
the renal NF-κB activation (15).
Furthermore, EA can reduce ROS production induced by HG in human
aortic endothelial cells by promoting ERK1/2 phosphorylation and
increasing the expression of NADPH oxidase 4 (NOX4). However,
whether EA could improve the damage of glomerular mesangial cells
induced by HG and the associated mechanism is still unclear. The
present study investigated the protective effect of EA on the
injury of glomerular mesangial cells induced by HG and further
explored the implicated molecular mechanism.
Materials and methods
Cell culture and treatment
EA, D-glucose and the PI3K inhibitor (LY294002) were
purchased from Sigma-Aldrich; Merck KGaA. Rat MCs (HBZY-1) were
purchased from the China Center for Type Culture Collection, cat.
no. GDC0124 (http://cctcc.whu.edu.cn/portal/no_center/cell_detail/id/101.html).
MCs were cultured in low glucose or HG DMEM containing 10% fetal
bovine serum (both Gibco; Thermo Fisher Scientific, Inc.). The
concentrations of D-glucose and streptomycin/penicillin (Gibco;
Thermo Fisher Scientific, Inc.) were 5.5 mM (or 30 mM) and 100
U/ml, respectively. Cells were incubated in a constant temperature
incubator with 5% CO2 at 37˚C. When the cells grew to
80% confluency, they were treated as follows: i) Normal control
group (NC), MCs were cultured in DMEM with 5.5 mM glucose; ii) HG
group, MCs were cultured in DMEM with 30 mM glucose; iii) HG and
low concentration EA intervention group (L-EA), MCs were cultured
in DMEM with 30 mM glucose and treated with 10 µM EA; iv) HG and
medium concentration EA intervention group (M-EA), MCs were
cultured in DMEM medium with 30 mM glucose and treated with 20 µM
EA; v) HG and high concentration EA intervention group (H-EA), MCs
were cultured in DMEM with 30 mM glucose and treated with 40 µM EA;
and vi) HG and PI3K inhibitor (LY294002) group, MCs were cultured
in DMEM with 30 mM glucose and treated with 20 µM LY294002.
Overall, MCs were cultured in DMEM with 5.5 mM glucose or DMEM with
30 mM glucose plus EA (10, 20 or 40 µM) or DMEM with 30 mM glucose
and 20 µM LY294002 at 37˚C for 24 h (16,17).
To detect the cytotoxic effect of EA on MCs,
different concentrations of EA (0, 1, 5, 10, 20, 40 or 80 µM ) or
20 µM LY294002 were incubated with MCs under low glucose conditions
at 37˚C for 24 h.
To examine whether EA mediated protection of MCs
following HG treatment via the PI3K/Akt signaling pathway,
HG-stimulated MCs were treated with EA (40 µM) and/or the PI3K
agonist 740Y-P (10 µM, MedChemExpress) at 37˚C for 24 h (18).
MTT assay
At a density of 1x105 cells/ml, MCs in
the logarithmic growth phase were inoculated into a 96-well plate
(4x103 cells/well) and subsequently cultured in a 5%
CO2 incubator at 37˚C for 24 h. After the cells were
treated as aforementioned for 24 h, 25 µl MTT (5 mg/ml) solution
was added into each well. After incubation at 37˚C for 4 h, 100 µl
dimethyl sulfoxide was added into each well. After 2 min
oscillation of the plate, the absorbance value was measured at 570
nm using a microplate reader (BioTek Instruments, Inc.).
5-Ethynyl-2'-deoxyuridine (EdU)
assay
MCs, treated as aforementioned, were incubated in
96-well plates at 4x103 cells/well in a 5%
CO2 incubator at 37˚C for 24 h. Subsequently, 1 ml 4%
paraformaldehyde was used for fixation at room temperature for 15
min and then cells were washed with PBS containing 3% BSA
(Sigma-Aldrich; Merck KGaA). Cells were subsequently incubated with
1 ml 0.5% Triton X-100 at room temperature for 20 min. After being
washed with PBS containing 3% BSA, cells in each group were treated
with 10 µM EdU for 30 min at room temperature. After washing with
PBS, DAPI (5 µg/ml) was added to the cells in the dark for 5 min at
room temperature. The cells were observed under an inverted
fluorescence microscope (Leica DMI3000-B; Leica Microsystems GmbH)
and the number of positive cells was counted from five random
fields in three wells using ImageJ 1.48v software (National
Institutes of Health).
Measurement of ROS
The intracellular ROS of MCs were detected using a
ROS detection kit (cat. no. S0033S; Beyotime Institute of
Biotechnology) according to the manufacturer's instructions.
Briefly, 2',7'-dichlorodihydrofluorescein diacetate (DCFH-DA) was
diluted with serum-free DMEM at a ratio of 1:1,000 to a final
concentration of 10 µM. The cells were collected and suspended in
the diluted DCFH-DA at a concentration of 1x108/ml,
incubated at 37˚C for 20 min and then washed with serum-free DMEM
three times. A fluorescence microscope (Leica DMI3000-B; Leica
Microsystems GmbH) was used to observe and capture images. The
fluorescence intensity was detected using a microplate reader
(BioTek Instruments, Inc.), at an excitation wavelength of 488 nm
and an emission wavelength of 525 nm.
Detection of malondialdehyde (MDA)
content and superoxide dismutase (SOD) activity
MDA content was detected using a commercial kit
(cat. no. S0131S; Beyotime Institute of Biotechnology) and was
performed in accordance with the manufacturer's instructions. MCs
were lysed for 30 min on ice using RIPA lysis buffer (Beyotime
Institute of Biotechnology) and centrifuged at 12,000 rpm for 10
min. Subsequently, 100 µl supernatant was obtained and mixed with
200 µl MDA working solution. Following that, the solution was
heated at 100˚C for 15 min, cooled to room temperature and
centrifuged at 1,000 x g for 10 min at room temperature.
Subsequently, 200 µl supernatant/MDA mixture was added into 96-well
plates and the absorbance value was determined at 532 nm using a
microplate reader (BioTek Instruments, Inc.). SOD activity was
detected using total SOD activity detection kit (WST-8 method; cat.
no. S0101S; Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. After the cells were lysed for 30 min
on ice using SOD sample preparation solution, 160 µl WTS-8/enzyme
working solution and 20 µl reaction working solution were mixed
with 20 µl supernatant and then incubated at 37˚C for 30 min. After
this incubation, the absorbance value at 450 nm was detected using
a microplate reader.
ELISA
MCs were inoculated into 12-well plates at a density
of 9x105 cells/well. They were incubated at 37˚C in 5%
CO2 for 24 h. MCs were treated as aforementioned and
cultured at 37˚C for 24 h. The cell culture medium was collected
and centrifuged at 1,000 x g for 10 min at 4˚C and the supernatant
was subsequently collected. The levels of IL-6 (cat. no. SR6000B),
IL-1β (cat. no. SRLB00), TNF-α (cat. no. SRTA00), fibronectin (FN)
(cat. no. ab108850), tissue inhibitor of metalloproteinases 1
(TIMP-1; cat. no. SRTM100) and MMP-9 (cat. no. DY8174-05) were
detected using commercial ELISA kits (R&D Systems, Inc. or
Abcam) according to the manufacturer's instructions. Detection was
carried out using a microplate reader (BioTek Instruments, Inc.) at
450 nm.
Western blotting
Nuclear proteins were obtained using NE-PER™ Nuclear
and Cytoplasmic Extraction Reagents kit (cat. no. 78833; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Total protein of MCs from each group was extracted
using RIPA lysis buffer (Beyotime Institute of Biotechnology) and
the protein concentration was measured with BCA protein assay kit
(Beyotime Institute of Biotechnology). The same amount of protein
(50 µg) sample per lane was electrophoresed on a 12% SDS-PAGE and
then transferred onto PVDF membranes. The membranes were blocked
using 5% skimmed milk powder for 1 h at room temperature. Following
blocking, the primary antibodies were added and the membranes were
incubated overnight at 4˚C. After being washed three times with
TBST containing 0.1% Tween-20, the membranes were incubated with
HRP-conjugated secondary antibodies at room temperature for 2 h.
After the membranes were washed three times with TBST, ECL reagent
(Bio-Rad Laboratories, Inc.) was used to visualize the bands.
Densitometry was performed by ImageJ 1.48v software (National
Institutes of Health). GAPDH was used as internal reference. The
antibodies were obtained from Abcam as follows: Anti-PI3K (cat. no.
ab191606; 1:1,000), anti-phosphorylated (p)-PI3K (cat. no.
ab182651; 1:1,000), anti-AKT (cat. no. ab179463; 1:1,000),
anti-p-AKT (cat. no. ab81283; 1:1,000), anti-FOXO3a (cat. no.
ab109629; 1:1,000), anti-SOD2 (cat. no. ab13533; 1:1,000),
anti-p-NF-κB (cat. no. ab76302; 1:1,000), anti-NF-κB (cat. no.
ab16502; 1:1,000), anti-lamin B1 (cat. no. ab16048; 1:1,000),
anti-GAPDH (cat. no. ab181602; 1:2,000).
Statistical analysis
SPSS v13.0 (SPSS, Inc.) and GraphPad Prism v6.01
software (GraphPad Software, Inc.) was used for statistical
analysis and presentation of the data. The data are presented as
the mean ± standard deviation. The comparison between multiple
groups was conducted using one-way ANOVA followed by Tukey's post
hoc test. All experiments were performed in triplicate. P<0.05
was considered to indicate a statistically significant
difference.
Results
Effects of EA on the viability and
proliferation of MCs induced by HG
The chemical formula of EA is presented in Fig. 1A. Different concentrations of EA (0,
1, 5, 10, 20, 40 or 80 µM) or 20 µM LY294002 were incubated with
MCs under low glucose conditions for 24 h. An MTT assay was used to
detect the possible effect of EA toxicity on MCs. The results
revealed that the survival rate of MCs exhibited no significant
change, indicating that EA in this concentration range had no
effect on the viability of MCs (Fig.
1B). Furthermore, the effect of EA on the viability of MCs
induced by HG was examined. The MTT assay results indicated that
compared with the NC group, the viability of MCs in the HG group
was significantly increased (Fig.
1C). Compared with the HG group, EA groups (10, 20 and 40 µM)
and the LY294002 group could significantly inhibit the increase of
cell viability induced by HG and 40 µM EA treatment caused the
greatest decrease (Fig. 1C).
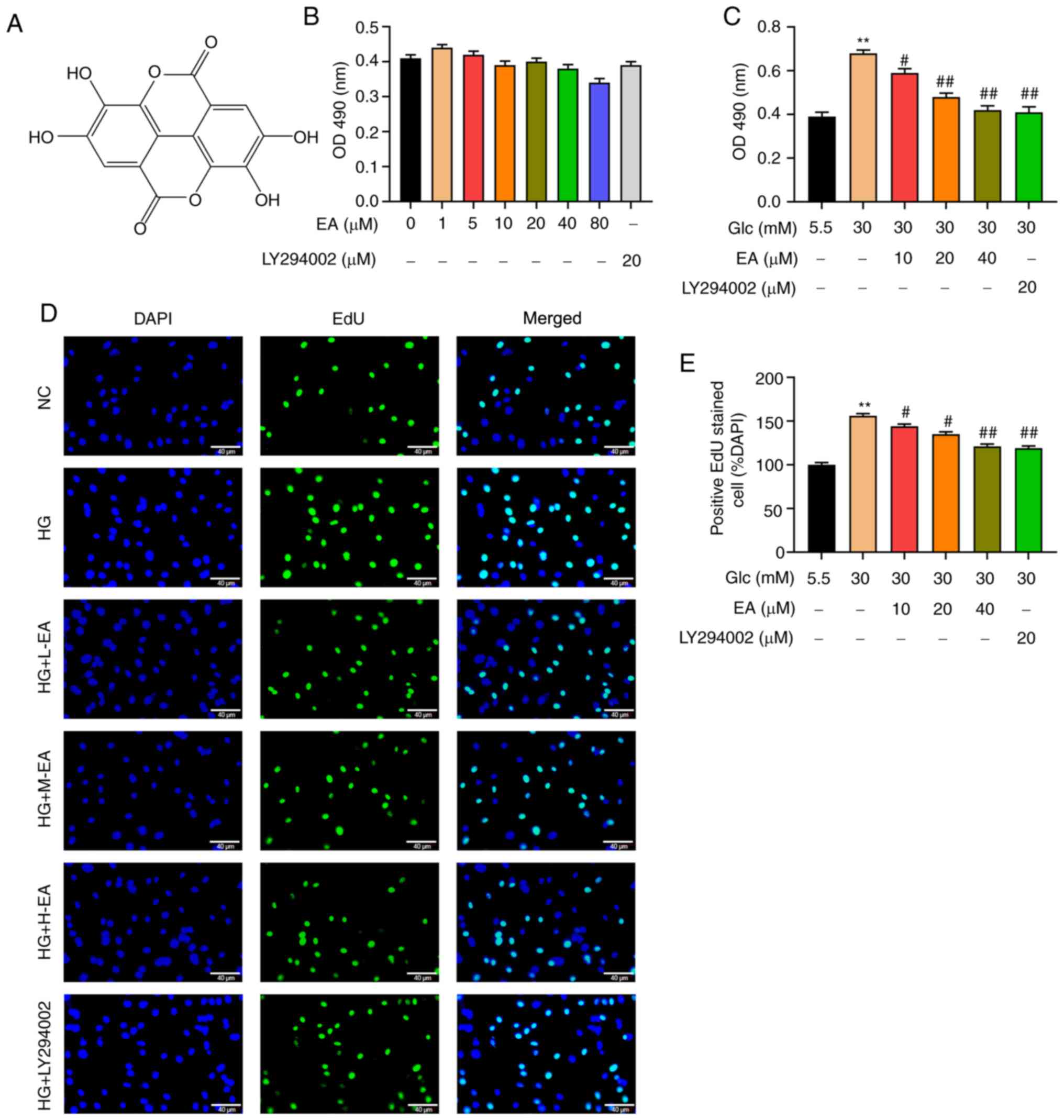 | Figure 1EA inhibits the proliferation of MCs
induced by HG. (A) Chemical structure of EA. (B) Cytotoxicity of EA
(1-80 µM) and LY294002 (20 µM) was detected via MTT assay in MCs
treated with 5.5 mM glucose. (C) Effect of EA (10, 20 and 40 µM)
and LY294002 (20 µM) on MC viability was detected using MTT assay.
Effect of EA (10, 20 and 40 µM) and LY294002 (20 µM) on MC
proliferation was (D) detected by EdU assay and (E) quantified.
Scale bar, 40 µm. **P<0.01 vs. NC group;
#P<0.05 and ##P<0.01 vs. HG group.
normal control (NC) group refers to 5.5 mM glucose treated cells.
EA, ellagic acid; MCs, mesangial cells; OD, optical density; HG,
high glucose; H-EA, high concentration EA intervention group; M-EA,
medium concentration EA intervention group; L-EA, low concentration
EA intervention group; NC, normal control; Glc, glucose; EdU,
5-ethynyl-2'-deoxyuridine. |
To detect the effect of EA on MC proliferation, EdU
assay was performed. As demonstrated in Fig. 1D and E, the number of positive cells in the HG
group was significantly increased compared with the NC group,
indicating that HG could significantly increase the proliferation
of MCs. Furthermore, compared with the HG group, EA groups (10, 20
and 40 µM) and the LY294002 group could significantly attenuate the
cell hyperproliferation induced by HG and 40 µM EA treatment
resulted in the greatest decrease.
Effects of EA on ROS, MDA and SOD in
MCs following HG treatment
Following EA treatment, the levels of ROS in MCs
were detected using a fluorescence microscope and the levels of SOD
and MDA were measured. The content of ROS in MCs was detected using
a DCFH-DA probe. The results revealed that compared with the NC
group, the fluorescence intensity of MCs in the HG group was
significantly increased, indicating that HG significantly increased
the level of ROS (Fig. 2A and
B). In addition, compared with the
HG group, the fluorescence intensity of MCs in the EA groups (10,
20 and 40 µM) and LY294002 group decreased significantly,
indicating that ROS level in each group was significantly
decreased. Moreover, the fluorescence intensity was attenuated with
the increase of EA concentration, which indicated that EA
alleviated oxidative stress in a concentration-dependent manner
(Fig. 2A and B).
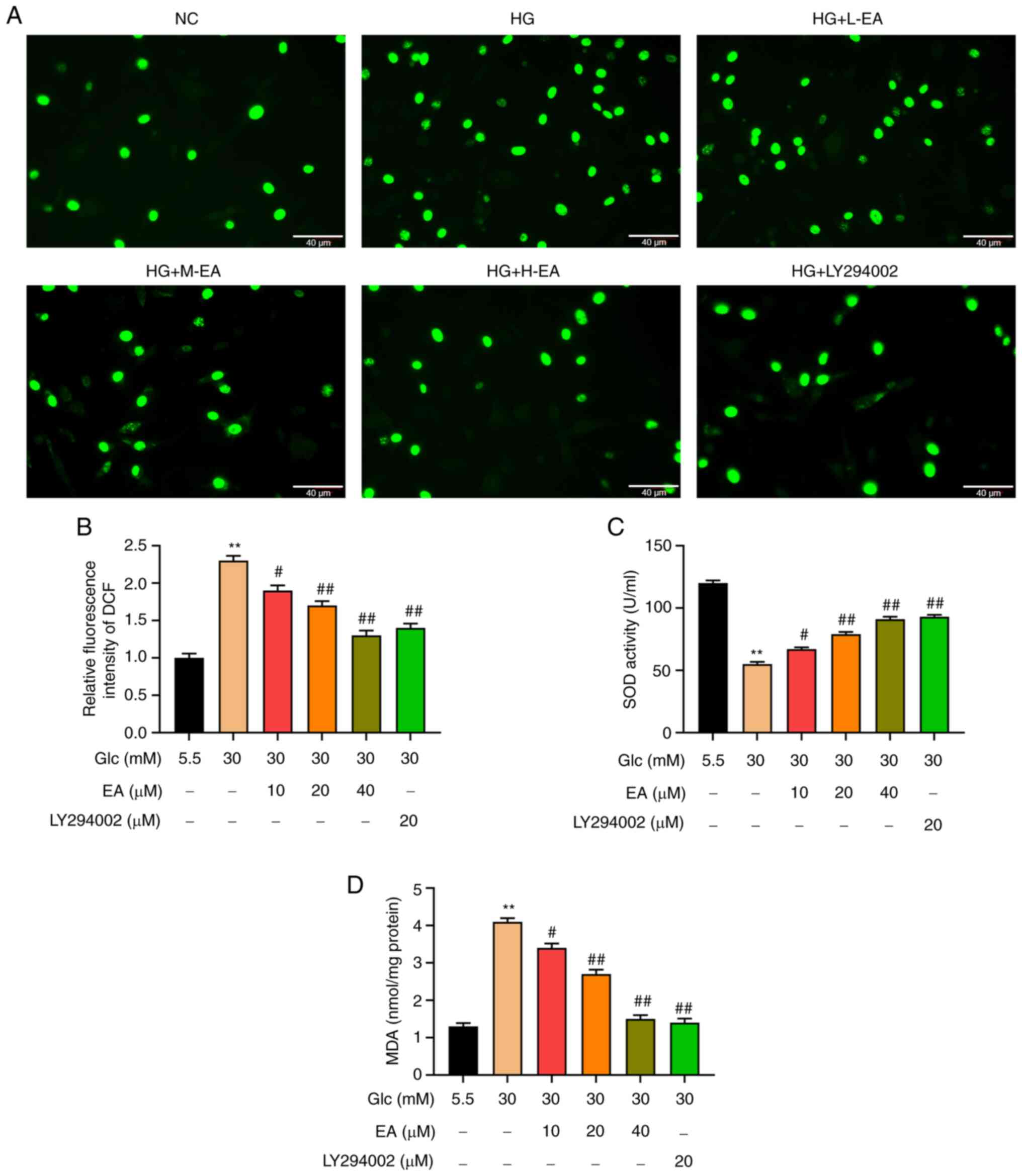 | Figure 2Antioxidant effect of EA on MCs
treated with HG. Effect of EA (10, 20 and 40 µM) and LY294002 (20
µM) on (A and B) the levels of reactive oxygen species, (C) SOD
activity and (D) MDA content in MCs induced by HG (30 mM). Scale
bar, 40 µm. **P<0.01 vs. NC group;
#P<0.05 and ##P<0.01 vs. HG group. NC
group refers to 5.5 mM glucose treated cells. EA, ellagic acid;
MCs, mesangial cells; HG, high glucose; H-EA, high concentration EA
intervention group; M-EA, medium concentration EA intervention
group; L-EA, low concentration EA intervention group; NC, normal
control; SOD, superoxide dismutase; MDA, malondialdehyde; DCF,
dichlorofluorescein; Glc, glucose. |
MDA content and SOD activity was detected in MCs
cells using commercial kits. The results indicated that the
activity of SOD in the HG group was significantly lower compared
with the NC group and that the MDA level was higher (Fig. 2C and D). Compared with the HG group, the
activity of SOD in the EA groups (10, 20 and 40 µM) and the
LY294002 group was significantly increased and MDA level was
decreased. The effect of EA on MDA content and SOD activity was
enhanced with the increase of EA concentration.
Effects of EA on the expression level
of inflammatory factors and ECM molecules in MCs induced by HG
In the present study, the levels of FN, MMP-9,
TIMP-1, IL-1β, IL-6 and TNF-α in MCs were detected by ELISA. The
results revealed that the expression levels of IL-6, IL-1β and
TNF-α were significantly increased in the HG group, but were
inhibited by a dose-dependent EA treatment and LY294002 (Fig. 3A-C). Furthermore, the levels of ECM
components were detected. The results demonstrated that compared
with the NC group, the FN levels in the HG group were significantly
increased, which indicated that HG resulted in increased ECM
synthesis (Fig. 3D). In addition,
the levels of MMP-9 and TIMP-1 were significantly increased, but
the ratio of MMP-9/TIMP-1 was significantly decreased compared with
the NC group, indicating that HG induced the degradation of ECM in
MCs (Fig. 3E-G). Moreover, the
increase of FN, MMP-9 and TIMP-1 contents and the decrease of
MMP-9/TIMP-1 ratio induced by HG were partially alleviated by a
dose-dependent treatment with EA and LY294002.
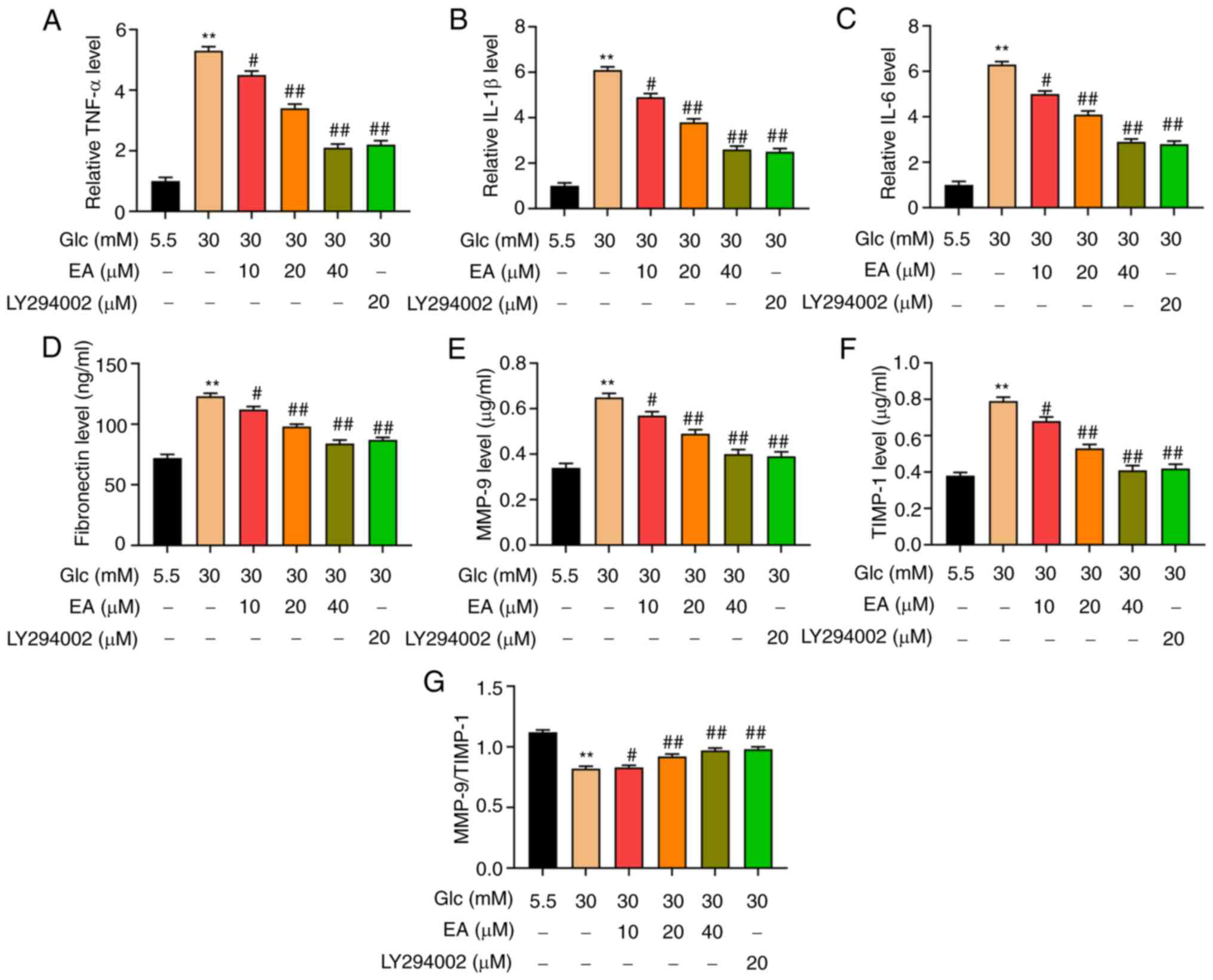 | Figure 3EA inhibits the secretion of
inflammatory factors and the production of ECM in MCs induced by
HG. Effect of EA (10, 20 and 40 µM) and LY294002 (20 µM) on
inflammatory factors (A) TNF-α, (B) IL-1β and (C) IL-6 was detected
in MCs induced by HG (30 mM). Effect of EA (10, 20 and 40 µM) and
LY294002 (20 µM) on ECM components (D) fibronectin, (E) MMP-9 and
(F) TIMP-1 was detected in MCs induced by HG (30 mM). (G)
MMP-9/TIMP-1 ratio was analyzed. **P<0.01 vs. NC
group; #P<0.05 vs. and ##P<0.01 vs. HG
group. NC group refers to 5.5 mM glucose treated cells. EA, ellagic
acid; ECM, extracellular matrix; MCs, mesangial cells; HG, high
glucose; Glc, glucose; TIMP-1, tissue inhibitor of
metalloproteinases 1. |
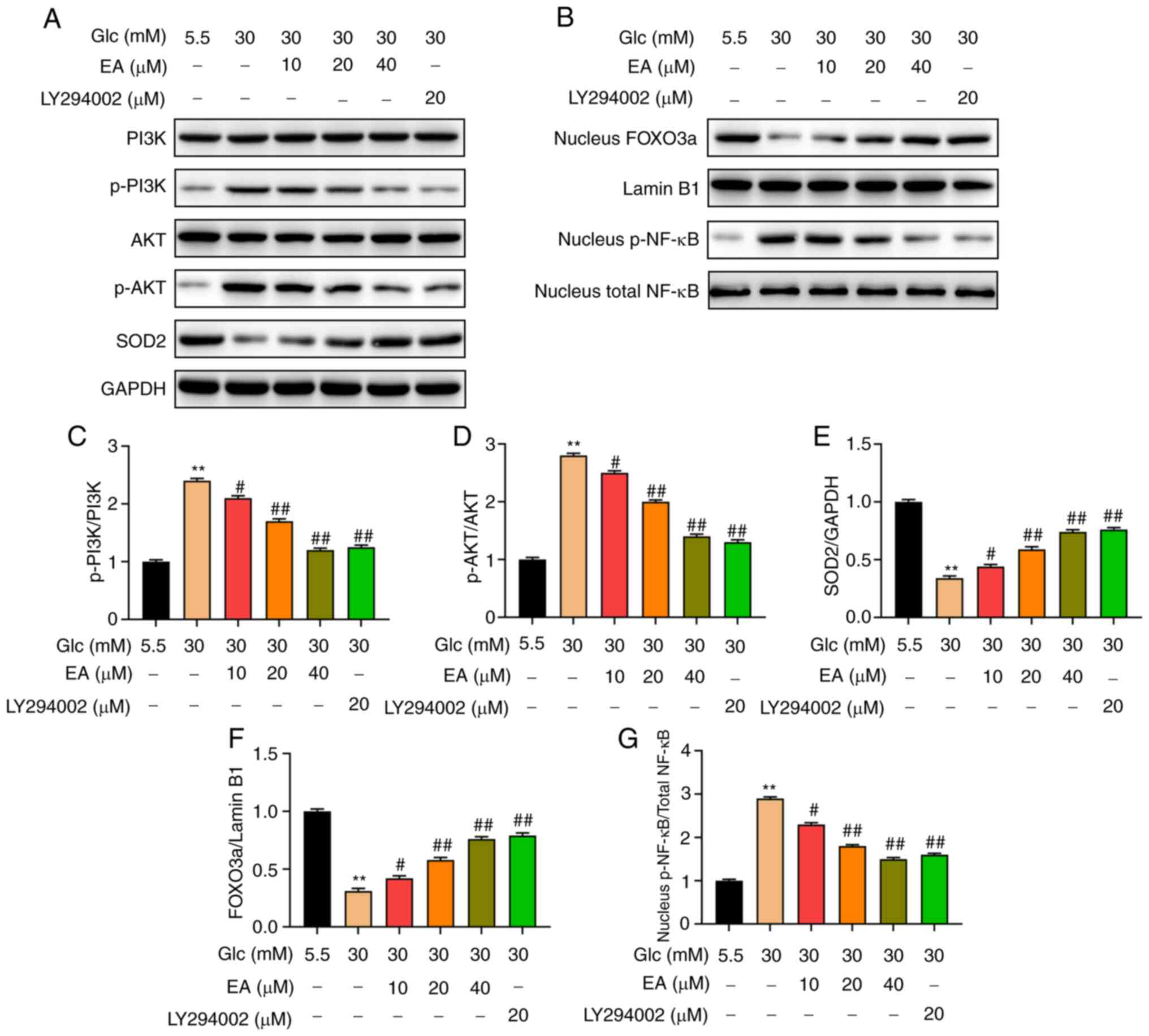
Effect of EA on the PI3K/Akt signaling
pathway and p-NF-κB and FOXO3a expression levels in the nuclei of
MCs following HG treatment
FOXO3a and NF-κB are downstream transcription
factors of the PI3K/Akt signaling pathway (19,20).
It was hypothesized that EA improved the damage of MCs induced by
HG by regulating the PI3K/Akt signaling pathway and the nuclear
expression of FOXO3a and NF-κB. The expression levels of PI3K,
p-PI3K, Akt, p-Akt, FOXO3, SOD2 and p-NF-κB in MCs were detected
using western blotting. The results revealed that the relative
expression levels of p-PI3K and p-Akt in the HG group were
significantly higher compared with those in the NC group, while the
relative expression level of SOD2 was significantly decreased
(Fig. 4A and C-E). In addition, compared with the HG
group, the relative expression levels of p-PI3K and p-Akt in the EA
groups (10, 20 and 40 µM) and the LY294002 group were significantly
decreased and the relative expression level of SOD2 was
significantly increased. The effect of EA was concentration
dependent. Moreover, the EA groups (10, 20 and 40 µM) and the
LY294002 group reduced the level of p-NF-κB and increased the
expression of FOXO3a in the nucleus of MCs induced by HG (Fig. 4B, F
and G).
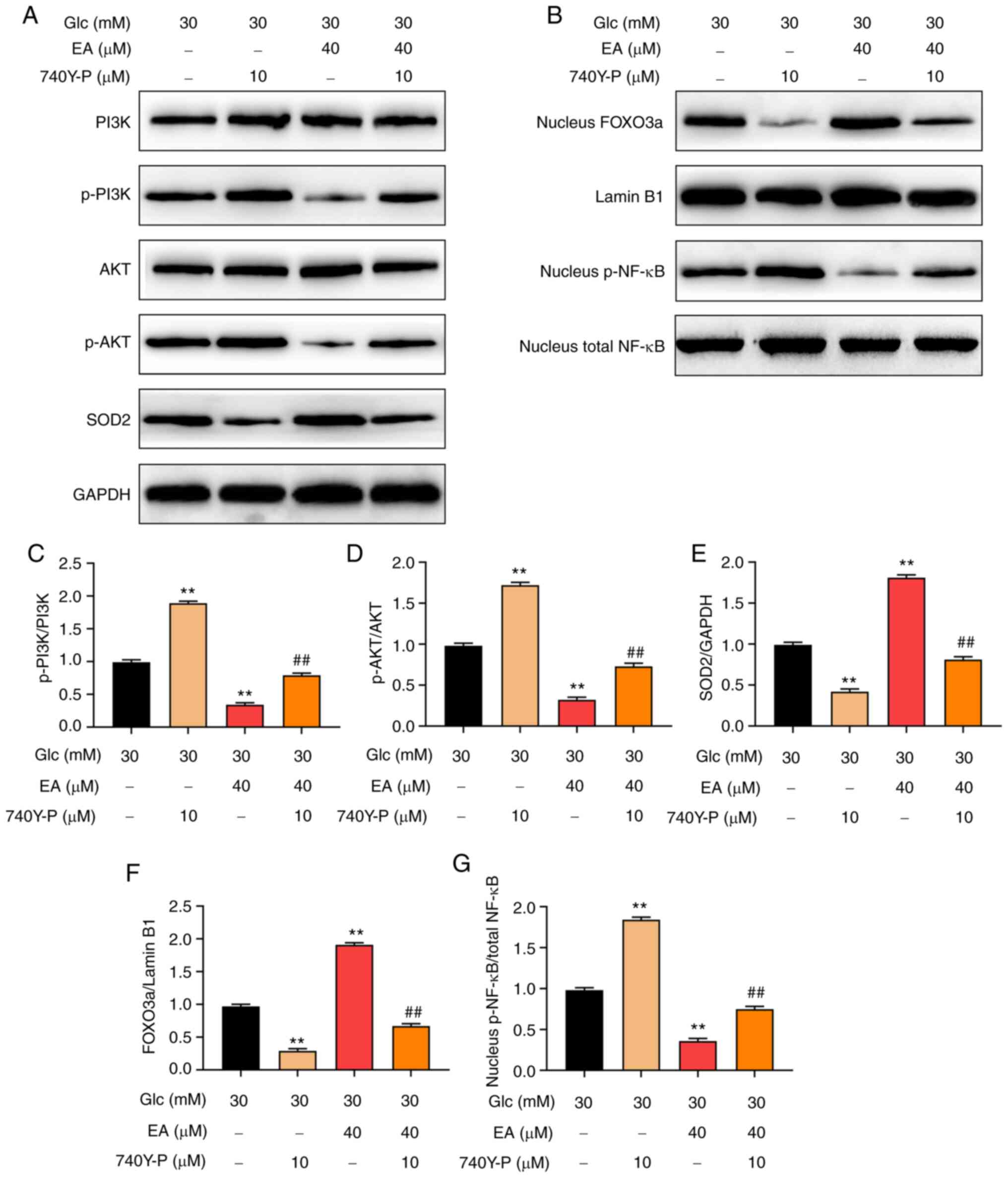
PI3K agonist 740Y-P reverses the
effects of EA on the PI3K/Akt signaling pathway and the p-NF-κB and
FOXO3a expression levels in the nuclei of MCs following HG
treatment
To investigate whether EA mediated protection of MCs
following HG treatment via regulating the PI3K/Akt signaling
pathway, HG-stimulated MCs were treated with EA (40 µM) and/or
740Y-P (10 µM) for 24 h (18). The
results revealed that 740Y-P further increased intranuclear p-NF-κB
levels and the phosphorylation of PI3K and Akt and decreased
intranuclear FOXO3a and SOD2 expression levels. However, EA
treatment exerted the opposite effect. In addition, 740Y-P could
attenuate the effect of EA on the PI3K/Akt signaling pathway,
intranuclear NF-κB, intranuclear FOXO3a and SOD2 expression level
(Fig. 5A-G). According to the
aforementioned results, PI3K inhibitor alleviated the injury of MCs
stimulated by HG. These results suggested that EA attenuated the
damage of HG-treated MCs by regulating the PI3K/Akt signaling
pathway.
Discussion
MCs participate in the early pathological changes of
DN, such as glomerular hypertrophy and ECM accumulation (5,21,22).
Hyperglycemia is the main cause of DN (23). Under normal conditions the number of
MCs is relatively stable, but under hyperglycemia abnormal
proliferation and ECM production are observed in MCs, which
eventually leads to glomerulosclerosis. Therefore, MCs are being
mainly investigated in several studies on the pathogenesis of DN
(24,25). HG-induced injury markers in MCs
include the abnormal proliferation of cells, the increase of ROS
and inflammatory factors and the accumulation of ECM (26). The present study used HG to treat
rat MCs to establish a DN model in vitro. EA is a type of
polyphenol compound, widely used in Chinese herbal medicines, such
as sanguisorba officinalis and pomegranate peel (27,28).
Numerous studies have demonstrated that EA possesses multiple
biological activities in vitro (14,29,30).
The present study explored whether EA could prevent the HG-induced
injury of MCs and its possible mechanism.
The proliferation of MCs can be abnormal due to
various pathophysiological changes and it can affect renal function
through multiple pathways. On the one hand, excessive proliferation
of MCs can induce cell adhesion factors (intercellular cell
adhesion molecule-1), ECM (MMPs, laminin and fibronectin) and other
substances (TNF-α, IL-1β and IL-6) to accumulate in the mesangial
region. On the other, excessive proliferation of MCs can promote
the release and secretion of inflammatory mediators, cytokines and
growth factors and accelerate the deterioration of renal function
(31-33).
A recent study has indicated that EA inhibited breast cancer cell
proliferation and increased apoptosis by downregulating CDK6
expression (34). In addition, EA
has been demonstrated to attenuate prostate carcinoma PC3 cell
proliferation and viability via activating caspases and inducing
apoptosis (35). The results of the
present study were consistent with these previous studies, as the
MTT and EdU assays indicated that EA could inhibit the
proliferation of MCs induced by HG in a concentration-dependent
manner. In addition, LY294002 also significantly inhibited cell
proliferation induced by HG, indicating that blocking the
activation of the PI3K/Akt signaling pathway was an effective way
to inhibit the proliferation of MCs.
Oxidative stress plays a notable role in the
occurrence and development of DN (36). HG can increase the production of ROS
or decrease the activity of antioxidant enzymes and thus increase
the production of MDA (37). It has
been previously demonstrated that EA can reduce intracellular
oxidative stress (38). In DN, the
levels of ROS are notably increased and non-enzymatic antioxidant
molecules (glutathione) and antioxidant enzyme (catalase and SOD)
activities are decreased (39).
Excessive ROS production can activate intracellular signal
transduction systems, such as PI3K/Akt, which activate
transcription factors, induce ECM protein synthesis and reduce
degradation, thereby leading to further development of DN (36,40).
EA has been indicated to decrease oxidative stress levels in
ischemic-reperfusion injury of renal tissues and cells and inhibit
NOX4 expression and phosphorylation of JAK1, JAK2 and
STAT1(41). EA can also reduce the
damage of vascular endothelial cells caused by diabetes-induced ROS
and function as a free radical scavenger by inhibiting the
ERK1/2/NOX4 signaling pathway (42). Similarly, the results of the present
study revealed that EA could significantly reduce ROS production,
improve SOD activity and decrease the MDA level in MCs treated with
HG in a concentration-dependent manner. LY294002, as a specific
inhibitor of the PI3K/Akt signaling pathway, could also increase
the activity of SOD and decrease the MDA level, thus reducing the
levels of ROS.
In DN, the balance of ECM secretion and degradation
is disrupted, which leads to the accumulation of ECM (43). DN also causes chronic inflammation,
as it has been demonstrated that the secretion of inflammatory
factors in MCs is associated with the production of ROS and
accumulation of ECM (44). The
MMP-9/TIMP-1 system serves a notable role in the degradation of
ECM, as it is the main enzyme system of ECM degradation (45). As a representative components of
ECM, FN is associated with the adhesion, migration and tissue
repair mediated by inflammatory cells in the kidney and also
participates in vascular basement membrane thickening and
glomerulosclerosis (46). DN also
causes chronic inflammation, as it has been demonstrated that the
secretion of inflammatory factors in MCs is associated with the
production of ROS and accumulation of ECM (44). In the present study, the results
demonstrated that HG could significantly increase the expression of
FN, MMP-9, TIMP-1, IL-1β, IL-6 and TNF-α, but the ratio of
MMP-9/TIMP-1 decreased significantly. Moreover, EA and LY294002
could effectively inhibit the HG-induced excessive secretion of FN,
MMP-9, TIMP-1, IL-1β, IL-6 and TNF-α and partially reverse the
MMP-9/TIMP-1 ratio to normal levels. Previous studies indicated
that EA improves renal injury in diabetic rats by inhibiting
NF-κB-induced inflammation (15).
EA can also reduce inflammatory factors in other disease models.
For instance, EA as an anti-inflammatory factor can improve
elastase-induced pneumonia and heart injury in a rat emphysema
model (47).
PI3K is a mediator between the upstream and
downstream signaling molecules activated by growth factor receptor
and G-protein-coupled receptor superfamily. PI3K can be activated
by a variety of extracellular signals (fibroblast growth factor,
vascular endothelial growth factor and Human growth factor).
Activated PI3K can promote phosphorylation of Akt, which acts as
the second messenger to mediate various cell functions (48). EA has been indicated to target the
PI3K signaling pathway, thus inhibiting the metastasis and
proliferation of endothelial cancer cells (49). Cell proliferation and the PI3K/Akt
signaling pathway are suppressed by EA in non-small cell lung
cancer (50). The dysfunction of
MCs induced by HG is associated with the activation of the PI3K/Akt
signaling pathway (51). The
present study investigated the effect of EA on the PI3K/Akt
signaling pathway in MCs induced by HG. Similarly, EA inhibited the
phosphorylation of PI3K and Akt. FOXO3a is mainly distributed in
mammalian nuclei. Phosphorylated Akt can induce the translocation
of FOXO3a from the nucleus to the cytoplasm, thereby inactivating
FOXO3a (48). A previous study has
demonstrated that upregulation of FOXO3a transcriptional level can
increase the expression of SOD2, thus reducing the production of
ROS (52). The current study
investigated whether EA could regulate intranuclear FOXO3a and SOD2
expression levels. The results revealed that EA treatment
significantly increased intranuclear FOXO3a and upregulated the
expression level of SOD2 following HG treatment. NF-κB, a
ubiquitous transcriptional regulator, is a key nuclear factor
involved in the regulation of cell proliferation and nuclear
inflammation-associated gene transcription. Activation of Akt can
promote the transcriptional activity of NF-κB (53,54).
EA improved liver injury in Wistar albino rats and inhibited NF-κB
expression (55). EA also inhibited
NF-κB and reduced renal tissue damage in diabetic nephropathy
(15). Accordingly, the present
study investigated the effect of EA on NF-κB and indicated that EA
attenuated the phosphorylation of intranuclear NF-κB in MCs induced
by HG. Furthermore, it was examined whether EA attenuated the MC
damage induced by HG via the PI3K/Akt signaling pathway. The
results indicated that the PI3K inhibitor LY294002 exerted the same
effect as EA on MCs following HG treatment and the PI3K agonist
740Y-P reversed the effect of EA on the PI3K/Akt signaling pathway
and the nuclear expression of p-NF-κB and FOXO3a.
In conclusion, the results of the present study
indicated that EA inhibited the activation of the PI3K/Akt
signaling pathway in MCs induced by HG and further regulated the
transcriptional activities of the downstream transcription factors
FOXO3 and NF-κB in the nucleus, thereby inhibiting cell
proliferation, oxidative stress, secretion of inflammatory factors
and ECM production. The current study provided an experimental
theoretical basis on the mechanism via which EA improved the injury
of MCs induced by HG. Limitations of the present study were that it
did not study the effect of PI3K agonist on the protective effect
of EA on MCs induced by high glucose and in vivo experiments
on EA improvement of DN were not performed. This is where research
could investigate further.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by Foundation for
Innovative Scientific Research of Young and Middle-Aged People in
The Second Affiliated Hospital of Harbin Medical University (grant
no. KYCX2018-16) and Heilongjiang Postdoctoral Fund (grant no.
LBH-Z18128).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL and GL contributed to study design, data
collection, statistical analysis, data interpretation and
manuscript preparation. XK, PG, YS, RD, XW, LC and RY contributed
to data collection and statistical analysis. QZ and FK contributed
to study design, data interpretation and manuscript preparation. WL
and GL confirm the authenticity of all the raw data. All authors
reviewed and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ilyas Z, Chaiban JT and Krikorian A: Novel
insights into the pathophysiology and clinical aspects of diabetic
nephropathy. Rev Endocr Metab Disord. 18:21–28. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Qi C, Mao X, Zhang Z and Wu H:
Classification and differential diagnosis of diabetic nephropathy.
J Diabetes Res. 2017(8637138)2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Wang G, Ouyang J, Li S, Wang H, Lian B,
Liu Z and Xie L: The analysis of risk factors for diabetic
nephropathy progression and the construction of a prognostic
database for chronic kidney diseases. J Transl Med.
17(264)2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Furuichi K, Shimizu M, Okada H, Narita I
and Wada T: Clinico-pathological features of kidney disease in
diabetic cases. Clin Exp Nephrol. 22:1046–1051. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Tung CW, Hsu YC, Shih YH, Chang PJ and Lin
CL: Glomerular mesangial cell and podocyte injuries in diabetic
nephropathy. Nephrology (Carlton). 23 (Suppl 4):32–37.
2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Qiao S, Liu R, Lv C, Miao Y, Yue M, Tao Y,
Wei Z, Xia Y and Dai Y: Bergenin impedes the generation of
extracellular matrix in glomerular mesangial cells and ameliorates
diabetic nephropathy in mice by inhibiting oxidative stress via the
mTOR/β-TrcP/Nrf2 pathway. Free Radic Biol Med. 145:118–135.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xu K, Guo L, Bu H and Wang H: Daphnetin
inhibits high glucose-induced extracellular matrix accumulation,
oxidative stress and inflammation in human glomerular mesangial
cells. J Pharmacol Sci. 139:91–97. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Jiang W, Wang R, Liu D, Zuo M, Zhao C,
Zhang T and Li W: Protective effects of kaempferitrin on advanced
glycation end products induce mesangial cell apoptosis and
oxidative stress. Int J Mol Sci. 19(19)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Cui FQ, Wang YF, Gao YB, Meng Y, Cai Z,
Shen C, Liu ZQ, Jiang XC and Zhao WJ: Effects of BSF on podocyte
apoptosis via regulating the ROS-mediated PI3K/AKT pathway in DN. J
Diabetes Res. 2019(9512406)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Manabe E, Handa O, Naito Y, Mizushima K,
Akagiri S, Adachi S, Takagi T, Kokura S, Maoka T and Yoshikawa T:
Astaxanthin protects mesangial cells from hyperglycemia-induced
oxidative signaling. J Cell Biochem. 103:1925–1937. 2008.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Moon JY, Tanimoto M, Gohda T, Hagiwara S,
Yamazaki T, Ohara I, Murakoshi M, Aoki T, Ishikawa Y, Lee SH, et
al: Attenuating effect of angiotensin-(1-7) on angiotensin
II-mediated NAD(P)H oxidase activation in type 2 diabetic
nephropathy of KK-A(y)/Ta mice. Am J Physiol Renal Physiol.
300:F1271–F1282. 2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Derosa G, Maffioli P and Sahebkar A:
Ellagic acid and its role in chronic diseases. Adv Exp Med Biol.
928:473–479. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zeb A: Ellagic acid in suppressing in vivo
and in vitro oxidative stresses. Mol Cell Biochem. 448:27–41.
2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ceci C, Lacal PM, Tentori L, De Martino
MG, Miano R and Graziani G: Experimental evidence of the antitumor,
antimetastatic and antiangiogenic activity of ellagic acid.
Nutrients. 10(10)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ahad A, Ganai AA, Mujeeb M and Siddiqui
WA: Ellagic acid, an NF-κB inhibitor, ameliorates renal function in
experimental diabetic nephropathy. Chem Biol Interact. 219:64–75.
2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Lee WJ, Ou HC, Hsu WC, Chou MM, Tseng JJ,
Hsu SL, Tsai KL and Sheu WH: Ellagic acid inhibits oxidized
LDL-mediated LOX-1 expression, ROS generation, and inflammation in
human endothelial cells. J Vasc Surg. 52:1290–1300. 2010.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Liu B, Lin J, Bai L, Zhou Y, Lu R, Zhang
P, Chen D, Li H, Song J, Liu X, et al: Paeoniflorin inhibits
mesangial cell proliferation and inflammatory response in rats with
mesangial proliferative glomerulonephritis through PI3K/AKT/GSK-3β
pathway. Front Pharmacol. 10(978)2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li F, Zhang Z, Wang P, Wen P, Xu Q, Wang
Y, Pan P and Ma L: ALC1 knockdown enhances cisplatin cytotoxicity
of esophageal squamous cell carcinoma cells by inhibition of
glycolysis through PI3K/Akt pathway. Life Sci.
232(116679)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhao C, Gu Y, Chen L and Su X:
Upregulation of FoxO3a expression through PI3K/Akt pathway
attenuates the progression of lupus nephritis in MRL/lpr mice. Int
Immunopharmacol. 89 (Pt A)(107027)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang J, Wang X, Vikash V, Ye Q, Wu D, Liu
Y and Dong W: ROS and ROS-mediated cellular signaling. Oxid Med
Cell Longev. 2016(4350965)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhao JH: Mesangial cells and renal
fibrosis. Adv Exp Med Biol. 1165:165–194. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Migliorini A, Ebid R, Scherbaum CR and
Anders HJ: The danger control concept in kidney disease: Mesangial
cells. J Nephrol. 26:437–449. 2013.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Cao A, Wang L, Chen X, Guo H, Chu S, Zhang
X and Peng W: Ursodeoxycholic acid ameliorated diabetic nephropathy
by attenuating hyperglycemia-mediated oxidative stress. Biol Pharm
Bull. 39:1300–1308. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Dong Z, Sun Y, Wei G, Li S and Zhao Z:
Ergosterol ameliorates diabetic nephropathy by attenuating
mesangial cell proliferation and extracellular matrix deposition
via the TGF-β1/Smad2 signaling pathway. Nutrients.
11(11)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Das F, Maity S, Ghosh-Choudhury N,
Kasinath BS and Ghosh Choudhury G: Deacetylation of S6 kinase
promotes high glucose-induced glomerular mesangial cell hypertrophy
and matrix protein accumulation. J Biol Chem. 294:9440–9460.
2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lu X, Fan Q, Xu L, Li L, Yue Y, Xu Y, Su
Y, Zhang D and Wang L: Ursolic acid attenuates diabetic mesangial
cell injury through the up-regulation of autophagy via
miRNA-21/PTEN/Akt/mTOR suppression. PLoS One.
10(e0117400)2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tan YH, Shudo T, Yoshida T, Sugiyama Y, Si
JY, Tsukano C, Takemoto Y and Kakizuka A: Ellagic acid, extracted
from Sanguisorba officinalis, induces G1 arrest by
modulating PTEN activity in B16F10 melanoma cells. Genes Cells.
24:688–704. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Peng R, Wu Q, Chen J, Ghosh R and Chen X:
Isolation of ellagic acid from pomegranate peel extract by
hydrophobic interaction chromatography using graphene oxide grafted
cotton fiber adsorbent. J Sep Sci. 41:747–755. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Baradaran Rahimi V, Ghadiri M, Ramezani M
and Askari VR: Antiinflammatory and anti-cancer activities of
pomegranate and its constituent, ellagic acid: Evidence from
cellular, animal, and clinical studies. Phytother Res. 34:685–720.
2020.PubMed/NCBI View
Article : Google Scholar
|
|
30
|
Huang Q, Chai WM, Ma ZY, Deng WL, Wei QM,
Song S, Zou ZR and Peng YY: Antityrosinase mechanism of ellagic
acid in vitro and its effect on mouse melanoma cells. J Food
Biochem. 43(e12996)2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Cove-Smith A and Hendry BM: The regulation
of mesangial cell proliferation. Nephron Exp Nephrol. 108:e74–e79.
2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Kurogi Y: Mesangial cell proliferation
inhibitors for the treatment of proliferative glomerular disease.
Med Res Rev. 23:15–31. 2003.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wolf G: Molecular mechanisms of diabetic
mesangial cell hypertrophy: A proliferation of novel factors. J Am
Soc Nephrol. 13:2611–2613. 2002.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Yousuf M, Shamsi A, Khan P, Shahbaaz M,
AlAjmi MF, Hussain A, Hassan GM, Islam A, Rizwanul Haque QM and
Hassan MI: Ellagic acid controls cell proliferation and induces
apoptosis in breast cancer cells via inhibition of cyclin-dependent
kinase 6. Int J Mol Sci. 21(21)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Malik A, Afaq S, Shahid M, Akhtar K and
Assiri A: Influence of ellagic acid on prostate cancer cell
proliferation: A caspase-dependent pathway. Asian Pac J Trop Med.
4:550–555. 2011.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Jha JC, Banal C, Chow BS, Cooper ME and
Jandeleit-Dahm K: Diabetes and kidney disease: Role of oxidative
stress. Antioxid Redox Signal. 25:657–684. 2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Sztanek F, Molnárné Molnár Á and Balogh Z:
The role of oxidative stress in the development of diabetic
neuropathy. Orv Hetil. 157:1939–1946. 2016.PubMed/NCBI View Article : Google Scholar : (In Hu).
|
|
38
|
Zhou B, Li Q, Wang J, Chen P and Jiang S:
Ellagic acid attenuates streptozocin induced diabetic nephropathy
via the regulation of oxidative stress and inflammatory signaling.
Food Chem Toxicol. 123:16–27. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Giacco F and Brownlee M: Oxidative stress
and diabetic complications. Circ Res. 107:1058–1070.
2010.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Kashihara N, Haruna Y, Kondeti VK and
Kanwar YS: Oxidative stress in diabetic nephropathy. Curr Med Chem.
17:4256–4269. 2010.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Liu Q, Liang X, Liang M, Qin R, Qin F and
Wang X: Ellagic acid ameliorates renal ischemic-reperfusion injury
through NOX4/JAK/STAT signaling pathway. Inflammation. 43:298–309.
2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Rozentsvit A, Vinokur K, Samuel S, Li Y,
Gerdes AM and Carrillo-Sepulveda MA: Ellagic acid reduces high
glucose induced vascular oxidative stress through ERK1/2/NOX4
Signaling Pathway. Cell Physiol Biochem. 44:1174–1187.
2017.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Kolset SO, Reinholt FP and Jenssen T:
Diabetic nephropathy and extracellular matrix. J Histochem
Cytochem. 60:976–986. 2012.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Wada J and Makino H: Inflammation and the
pathogenesis of diabetic nephropathy. Clin Sci (Lond). 124:139–152.
2013.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Rysz J, Banach M, Stolarek RA, Pasnik J,
Cialkowska-Rysz A, Koktysz R, Piechota M and Baj Z: Serum matrix
metalloproteinases MMP-2 and MMP-9 and metalloproteinase tissue
inhibitors TIMP-1 and TIMP-2 in diabetic nephropathy. J Nephrol.
20:444–452. 2007.PubMed/NCBI
|
|
46
|
Hu C, Sun L, Xiao L, Han Y, Fu X, Xiong X,
Xu X, Liu Y, Yang S, Liu F, et al: Insights into the mechanisms
involved in the expression and regulation of extracellular matrix
proteins in diabetic nephropathy. Curr Med Chem. 22:2858–2870.
2015.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Mansouri Z, Dianat M, Radan M and Badavi
M: Ellagic acid ameliorates lung inflammation and heart oxidative
stress in elastase-induced emphysema model in rat. Inflammation.
43:1143–1156. 2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhang M and Zhang X: The role of
PI3K/AKT/FOXO signaling in psoriasis. Arch Dermatol Res. 311:83–91.
2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Wang Y, Ren F, Li B, Song Z, Chen P and
Ouyang L: Ellagic acid exerts antitumor effects via the PI3K
signaling pathway in endometrial cancer. J Cancer. 10:3303–3314.
2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Liu Q, Liang X, Niu C and Wang X: Ellagic
acid promotes A549 cell apoptosis via regulating the
phosphoinositide 3-kinase/protein kinase B pathway. Exp Ther Med.
16:347–352. 2018.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Wang EM, Fan QL, Yue Y and Xu L: Ursolic
acid attenuates high glucose-mediated mesangial cell injury by
inhibiting the phosphatidylinositol 3-kinase/Akt/mammalian target
of rapamycin (PI3K/Akt/mTOR) signaling pathway. Med Sci Monit.
24:846–854. 2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Kops GJ, Dansen TB, Polderman PE, Saarloos
I, Wirtz KW, Coffer PJ, Huang TT, Bos JL, Medema RH and Burgering
BM: Forkhead transcription factor FOXO3a protects quiescent cells
from oxidative stress. Nature. 419:316–321. 2002.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Rangan G, Wang Y and Harris D: NF-kappaB
signalling in chronic kidney disease. Front Biosci. 14:3496–3522.
2009.PubMed/NCBI View
Article : Google Scholar
|
|
54
|
Yang L, Hu X and Mo YY: Acidosis promotes
tumorigenesis by activating AKT/NF-κB signaling. Cancer Metastasis
Rev. 38:179–188. 2019.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Aslan A, Gok O, Erman O and Kuloglu T:
Ellagic acid impedes carbontetrachloride induced liver damage in
rats through suppression of NF kB, Bcl 2 and regulating Nrf 2 and
caspase pathway. Biomed Pharmacother. 105:662–669. 2018.PubMed/NCBI View Article : Google Scholar
|