Introduction
Liver cancer is one of the most common causes of
cancer-associated mortality worldwide, with the fifth highest
incidence and mortality rates in the United States, and a
continuously increasing mortality rate on an annual basis, despite
the advances in medical technology (1). Hepatocellular carcinoma (HCC) is the
most common type of primary liver cancer, and its occurrence has
been associated with hepatitis B virus, hepatitis C virus, smoking,
alcohol consumption, obesity, diabetes and other factors (2,3).
However, there is a lack of effective treatment strategies for HCC
due to its high metastasis and recurrence rates (4). In recent years, with in-depth research
on the molecular mechanism of HCC tumorigenesis, certain targeted
drugs for patients with HCC have been developed (5). However, their efficacy is
unsatisfactory due to the heterogeneity of tumors and the
complexity of the molecular mechanisms involved (6). Therefore, it is urgent to further
study the pathogenesis of HCC and to explore potential molecular
targets to facilitate the development of treatments for the
disease.
Assembly factor for spindle microtubules (ASPM) is
an ASP homologous gene of Drosophila melanogaster expressed
in the cytoplasm, which is crucial for the normal mitotic spindle
function of embryonic neuroblasts (7,8). A
number of studies have reported that ASPM serves an important role
in the progression of various types of cancer. For instance, Pai
et al (9) has reported that
ASPM increases the stability of the upstream regulatory factor of
the Wnt signaling pathway, enhances the Wnt-dishevelled segment
polarity protein 3-β-catenin signaling pathway and promotes the
malignant progression of prostate cancer. Different subtypes of
ASPM serve different roles in pancreatic cancer, and ASPM-II mainly
regulates Wnt signaling and tumor growth, while ASPM-III
selectively regulates the cell cycle progression of pancreatic
ductal adenocarcinoma cells (10).
In addition, it has been demonstrated in a number of studies that
ASPM can serve as a biomarker in bladder cancer (11,12),
breast cancer (13), non-small cell
lung cancer (14), prostate cancer
(15) and ovarian cancer (16,17).
Additionally, high ASPM expression is often associated with a poor
prognosis (18). Lin et al
(19) reported that ASPM expression
is upregulated in HCC, and that upregulation of ASPM increases the
invasion and metastasis of HCC. Furthermore, several studies have
suggested ASPM as a prognostic biomarker for HCC (20,21),
although the specific mechanism of action of ASPM in HCC remains
elusive.
Kinesin family member 11 (KIF11), which encodes EG5,
belongs to the kinesin-like protein family. KIF11 serves an
essential role in cell mitosis (22) and the transport of secreted proteins
(23). It has been demonstrated
that KIF11 mutation causes autosomal dominant familial exudative
vitreoretinopathy, as well as microcephaly, with or without
choroidal retinopathy, lymphedema or hypophrenia (24,25).
KIF11 acts as an oncogene in tumors and is associated with a poor
prognosis in breast cancer (26).
Additionally, it promotes cell invasion, proliferation and
self-renewal in glioblastoma (27).
The present study aimed to investigate the
expression levels of ASPM in HCC cells and to determine whether
interference with ASPM expression could affect cell proliferation,
invasion, migration and epithelial-to-mesenchymal transition (EMT).
It was further investigated whether ASPM was associated with KIF11
to promote the malignant progression of HCC, as well as the
involvement of the Wnt/β-catenin signaling pathway in this process.
The aim of the present study was to provide a novel approach to the
targeted therapy and clinical diagnosis of HCC in the future.
Materials and methods
Cell lines
The normal human liver cell line (THLE-2) and human
HCC cell lines (SK-HEP-1, Huh7, HCC-LM3 and Hep3B) were purchased
from National Infrastructure of Cell Line Resource. Each cell line
was cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Thermo Fisher Scientific, Inc.) and 100
U/ml Penicillin/Streptomycin (Thermo Fisher Scientific, Inc.) with
5% CO2 at 37˚C.
Bioinformatics analysis
Gene Expression Profiling Interactive Analysis
(GEPIA; http://gepia.cancer-pku.cn/) is a
network server for gene expression analysis based on tumor and
normal samples in The Cancer Genome Atlas and Genotype-Tissue
Expression databases (28). GEPIA
was used to detect the expression levels of ASPM and KIF11 in
patients with liver hepatocellular carcinoma (LIHC), as well as
their overall survival and disease-free survival rates, with
|log2 fold change|>1 and P<0.01 used as cut-off
values. The Human Protein Atlas (HPA; http://www.proteinatlas.org/) database is a data
resource that integrates a tissue atlas, cell atlas, pathological
atlas, brain atlas, blood atlas and metabolic atlas. The present
study analyzed the association between prognosis and ASPM and KIF11
in HCC using the pathological atlas in the HPA database. The Search
Tool for the Retrieval of Interacting Genes/Proteins (STRING;
https://www.string-db.org/) database is
an online database for the search of known protein-protein
interactions. The interactive association between ASPM and KIF11
was explored through the multi-protein retrieval of the STRING
database. The present study examined the correlation between ASPM
and KIF11 expression using the pan-cancer analysis platform
starBase (http://starbase.sysu.edu.cn/), which enables the
analysis of interactions between multiple RNAs (29).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. cDNA was synthesized
using a reverse transcription system kit (Invitrogen; Thermo Fisher
Scientific, Inc.) as follows: Priming at 25˚C for 5 min, reverse
transcription at 37˚C for 30 min and reverse transcription
inactivation at 85˚C for 5 min. For qPCR, the MiScript SYBR-Green
PCR kit (Qiagen GmbH) was used according to the manufacturer's
protocol. The thermocycling condition was as follows: Initial
denaturation at 95˚C for 3 min, 40 cycles of denaturation at 95˚C
for 10 sec and annealing at 60˚C for 30 sec and extension at 60˚C
for 30 sec. The normalization method was adopted with GADPH as the
internal reference, and the results were presented as the
differences in target gene expression quantity between the control
group and the experimental group with the use of 2-ΔΔCq
values (30). The primers used in
the present study are listed in Table
I.
 | Table ISequences of the primers used for
reverse transcription-quantitative PCR. |
Table I
Sequences of the primers used for
reverse transcription-quantitative PCR.
| Gene | Sequence |
|---|
| KIF11 | F:
5'-GATGGACGTAAGGCAGCTCA-3' |
| | R:
5'-TGTGGTGTCGTACCTGTTGG-3' |
| ASPM | F:
5'-GGAGCGAGATCCCTCCAAAAT-3' |
| | R:
5'-GGCTGTTGTCATACTTCTCATGG-3' |
| Cadherin | F:
5'-CTGTGCCCAGCCTCCATGTTTT-3' |
| | R:
5'-CTGGATAGCTGCCCATTGCAAGTTA-3' |
| Cadherin | F:
5'-CCATCAAGCCTGTGGGAATC-3' |
| | R:
5'-GCAGATCGGACCGGATACTG-3' |
| Vimentin | F:
5'-TGTCCAAATCGATGTGGATGTTTC-3' |
| | R:
5'-TTGTACCATTCTTCTGCCTCCTG-3' |
| Ki-67 | F:
5'-CCACACTGTGTCGTCGTTTG-3' |
| | R:
5'-CCGTGCGCTCATCCATTCA-3' |
| PCNA | F:
5'-ATGTTTGAGGCACGCCTGATCCAG-3' |
| | R:
5'-CTAAGATGCTTCCTCACTTCAATC-3' |
| GAPDH | F:
5'-GGAGCGAGATCCCTCCAAAAT-3' |
| | R:
5'-GGCTGTTGTCATACTTCTCATGG-3' |
Western blot analysis
The cells of each treatment group were collected and
protein was extracted on ice with precooled RIPA lysis buffer
(Beyotime Institute of Biotechnology) for 10 min. Protein was
quantified using a BCA assay kit (Abcam) and the same amount of
protein (25 µg/lane) was separated by 12% SDS-PAGE. Subsequently,
the proteins were transferred to nitrocellulose membranes at 60 V
for 120 min. The membranes were first blocked with 5% skimmed milk
for 2 h at room temperature. The membranes were then incubated with
primary antibodies at 4˚C overnight followed by HRP-conjugated goat
anti-rabbit IgG secondary antibody at room temperature for 120 min.
Finally, the chemiluminescence reaction was performed using an ECL
kit (Beijing Solarbio Science & Technology Co., Ltd.), and
images were captured to observe the blots. The densitometry was
analyzed using the ImageJ software (v1.8; National Institutes of
Health). Detailed information on the antibodies is shown in
Table II. The experiment was
repeated in triplicate.
| Table IIInformation on the antibodies used
for western blotting. |
Table II
Information on the antibodies used
for western blotting.
| Antibody | Dilution | Cat. no. | Specificity | Manufacturer |
|---|
| KIF11 | 1:1,000 | ab254298 | Rabbit
monoclonal | Abcam |
| ASPM | 1:1,000 | ab238106 | Rabbit
polyclonal | Abcam |
| PCNA | 1:1,000 | ab92552 | Rabbit
monoclonal | Abcam |
| Ki-67 | 1:5,000 | ab16667 | Rabbit
monoclonal | Abcam |
| MMP2 | 1:5,000 | ab92536 | Rabbit
monoclonal | Abcam |
| MMP9 | 1:5,000 | ab76003 | Rabbit
monoclonal | Abcam |
| N-cadherin | 1:5,000 | ab76011 | Rabbit
monoclonal | Abcam |
| Vimentin | 1:5,000 | ab92547 | Rabbit
monoclonal | Abcam |
| E-cadherin | 1:10,000 | ab133597 | Rabbit
monoclonal | Abcam |
| β-catenin | 1:5,000 | ab32572 | Rabbit
monoclonal | Abcam |
| p-GSK-3β | 1:1,000 | 9336 | Rabbit
monoclonal | Cell Signaling
Technology, Inc. |
| GSK-3β | 1:1,000 | 9325 | Rabbit
monoclonal | Cell Signaling
Technology, Inc. |
| GAPDH | 1:10,000 | ab181602 | Rabbit
monoclonal | Abcam |
| IgG H&L
(HRP) | 1:10,000 | ab6721 | Goat anti-rabbit
HRP | Abcam |
Cell transfection
Silencer Select small interfering (si)RNA (si-ASPM;
5'-UGCCAUGGUGCAACUUGCU-3') and Silencer Select Negative Control No.
1 siRNA (si-NC; 5'-UUACCUCUAGUCGUCAUGU-3') were purchased from
Thermo Fisher Scientific, Inc. pEGFP1-KIF11 and pEGFP-1 as negative
control were purchased from Sangon Biotech Co., Ltd. A total of
0.8x106 cells were cultured in 35-mm culture dishes
until the logarithmic growth phase, and Lipofectamine®
3000 (Invitrogen; Thermo Fisher Scientific, Inc.) was used to
transfect 100 nM siRNA, 4 µg recombinant vector and their
respective controls into Hep3B cells for 24 h at 37˚C, according to
the product manual. Following transfection for 24 h, transfected
Hep3B cells were utilized for subsequent experimentation.
Cell counting Kit-8 (CCK-8) assay
Transfected or non-tranfected Hep3B cells were
inoculated into a 96-well plate at a density of 2,000 cells/well.
Sterile CCK-8 solution was added after 0, 24, 48 and 72 h of cell
culture. After 2 h of incubation at 37˚C, cell viability was
detected. Absorbance at a wavelength of 450 nm was measured by a
microplate reader (Molecular Devices, LLC).
Colony formation assay
Following digestion into a single-cell suspension
using trypsin, the cells were inoculated into a 6-well plate at a
density of 1,000 cells/well. After 1 week, each well was rinsed
three times with PBS at room temperature. After fixation with 4%
paraformaldehyde at room temperature for 15 min, 0.1% crystal
violet (Thermo Fisher Scientific, Inc.) was added for staining at
room temperature for 15 min. Subsequently, the clusters containing
>50 cells were considered as a colony and counted under a light
microscope (magnification, x40; Nikon Corporation).
Wound healing assay
Hep3B cells (5x105) were inoculated into
a six-well plate. At 80% confluence, a gentle scrape on the well
surface was performed with a pipette nozzle. Subsequently, the
cells were washed three times with PBS to remove free cells.
Following culture in serum-free medium at 37˚C for 24 h, cell
migration was observed at 0 and 24 h and images were captured under
a light microscope (magnification, x100; Nikon Corporation).
Transwell assay
Transwell inserts for 24-well plates (8 µm; Corning,
Inc.) were coated with prediluted Matrigel (1:8; BD Biosciences) at
37˚C for 30 min. The cells were washed twice in PBS and then
suspended in serum-free DMEM. A total of 5x105 cells in
200 µl medium were placed into the Matrigel-coated upper chamber
and the lower chamber was filled with 400 µl DMEM with 10% FBS.
Following incubation at 37˚C for 24 h, the non-invading cells in
the upper chamber were gently removed. The cells on the submembrane
surface were stained with 0.5% crystal violet for 15 min at room
temperature and observed under a light microscope (magnification,
x100; Nikon Corporation) and images were captured.
Co-immunoprecipitation (CO-IP)
assay
The cells were separated, washed twice with PBS, and
lysed in precooled cell lysis buffer for IP (cat. no. P0013;
Beyotime Institute of Biotechnology) containing protease
inhibitors. The supernatant was collected after centrifugation at
13,000 x g for 10 min at 4˚C. The supernatant of cell lysate (500
µg) were incubated at 4˚C overnight after the addition of 1 µg
KIF11 (cat. no. ab254298; Abcam) or IgG antibody (cat. no. 2729;
Cell Signaling Technology, Inc.). The pretreated 50 µg protein A
agarose beads were added to the cell lysis buffer incubated with
the antibody overnight for a slow shake, followed by incubation at
4˚C for 4 h, so that the antibody was coupled with protein A
agarose beads. After the IP reaction, agarose beads were
centrifuged at 1,000 x g for 3 min at 4˚C to the bottom of the
tube. The supernatant was then carefully absorbed, and the agarose
beads were washed three times with 1 ml lysis buffer. A total of 15
µl 2X SDS sample buffer was finally added for boiling at 100˚C for
5 min, followed by western blotting, as aforementioned.
Statistical analysis
All data were processed using GraphPad Prism 8.0
(GraphPad Software, Inc.). Measurement data are presented as the
mean ± SD. Comparisons between two groups were conducted using a
unpaired Student's t-test, and one-way ANOVA with Tukey's test was
used to compare multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
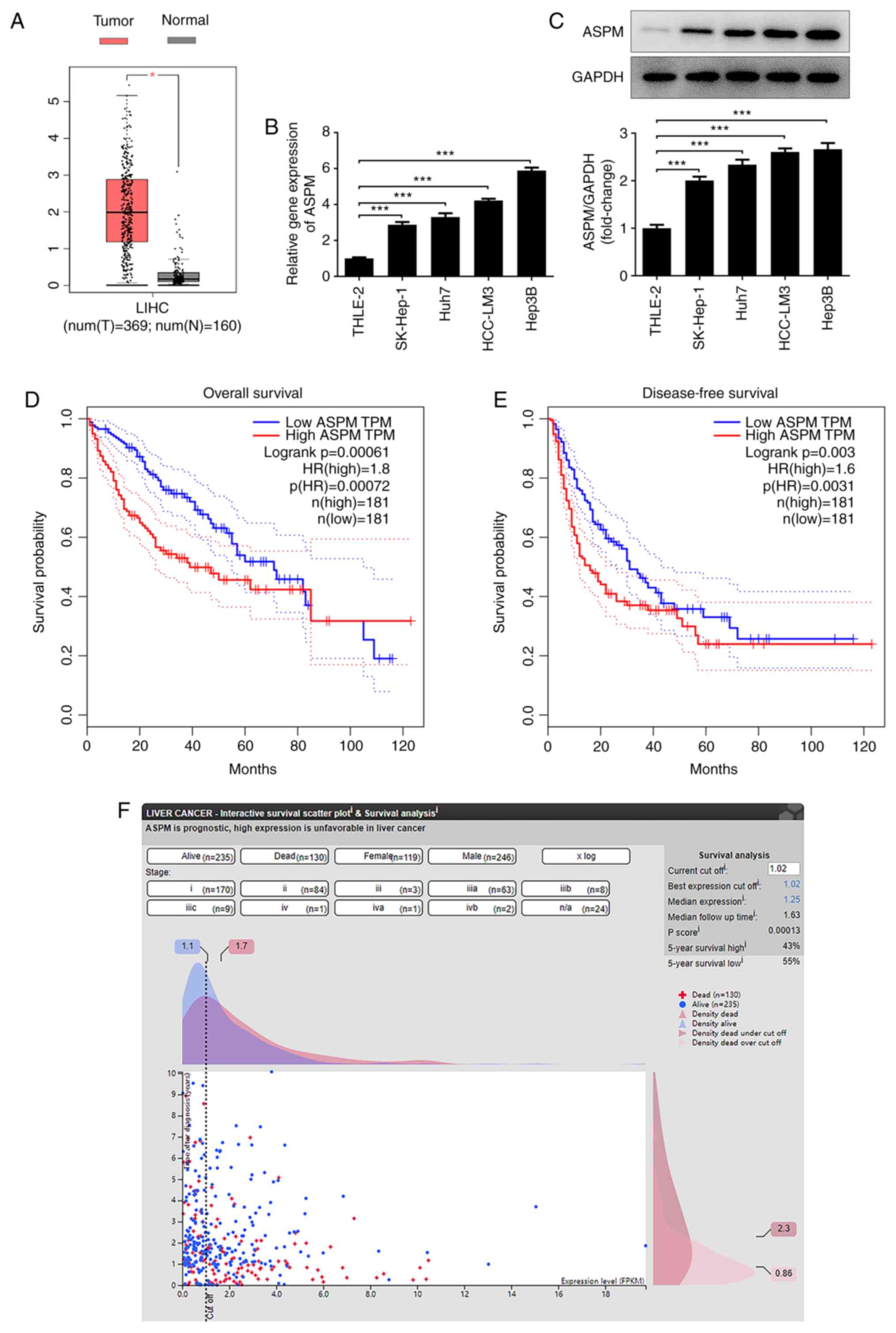
ASPM is highly expressed in HCC
tissues and cell lines
To explore the expression levels of ASPM in HCC and
its clinical significance, the GEPIA database was used for the
detection of the expression levels of ASPM in HCC and normal liver
tissues, and to analyze the overall survival and disease-free
survival rates. A total of 160 normal samples and 369 tumor samples
were obtained from the LIHC dataset of GEPIA2, with |Log2 fold
change|>1 and P<0.01 as cut-off values. Higher ASPM
expression was observed in HCC tissues compared with in normal
tissues (Fig. 1A). In addition, the
expression levels of ASPM in normal hepatocytes and HCC cell lines
were detected by RT-qPCR and western blotting, and the results
revealed significantly higher expression levels of ASPM in HCC cell
lines compared with in normal hepatocytes (Fig. 1B and C). Furthermore, Hep3B cells were selected
for further experiments, as they exhibited the highest expression
levels of ASPM. To evaluate of the significance of ASPM in clinical
studies of HCC, the present study revealed through GEPIA database
analysis that the overall survival rate was markedly different
between the low and high ASPM groups before 80 months, while there
was no difference observed after 80 weeks. In terms of the
disease-free survival rate, there was no difference at ~50 months.
In general, the overall and disease-free survival of patients with
high ASPM expression were markedly lower than those of patients
with low ASPM expression (Fig. 1D
and E). Furthermore, in HPA
database analysis, most patients with low ASPM expression were
alive within 10 years after diagnosis. In patients with high ASPM
expression, a large proportion of deaths had already occurred
within 1 year after diagnosis. This indicated that ASPM was closely
associated with prognosis (Fig.
1F), revealing that ASPM may serve as an independent prognostic
factor for patients with HCC.
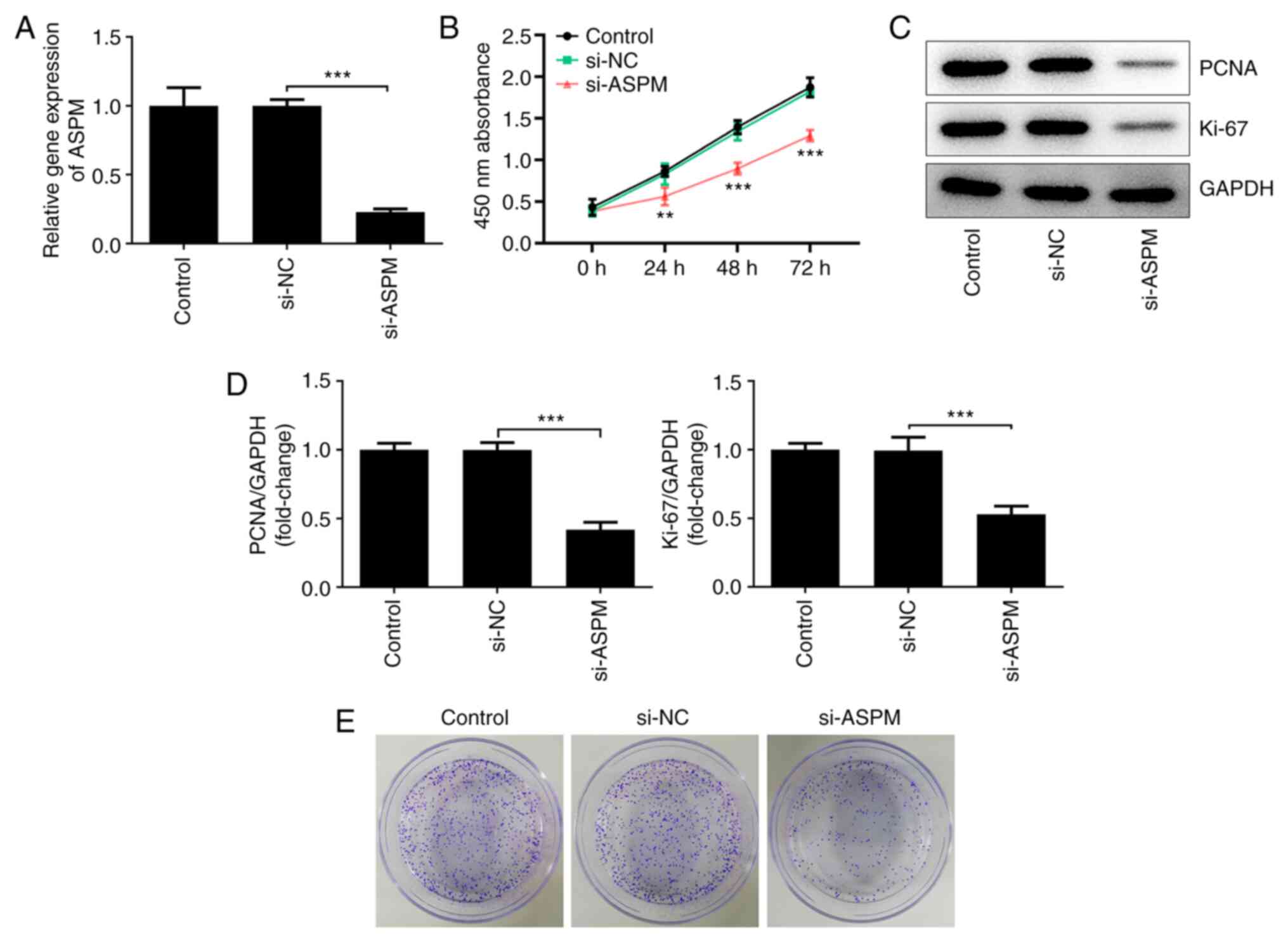
Interference with ASPM inhibits the
proliferation of HCC cells
To further investigate the biological functions of
ASPM in HCC cell lines, the present study interfered with ASPM
expression in Hep3B cells and detected the expression levels of
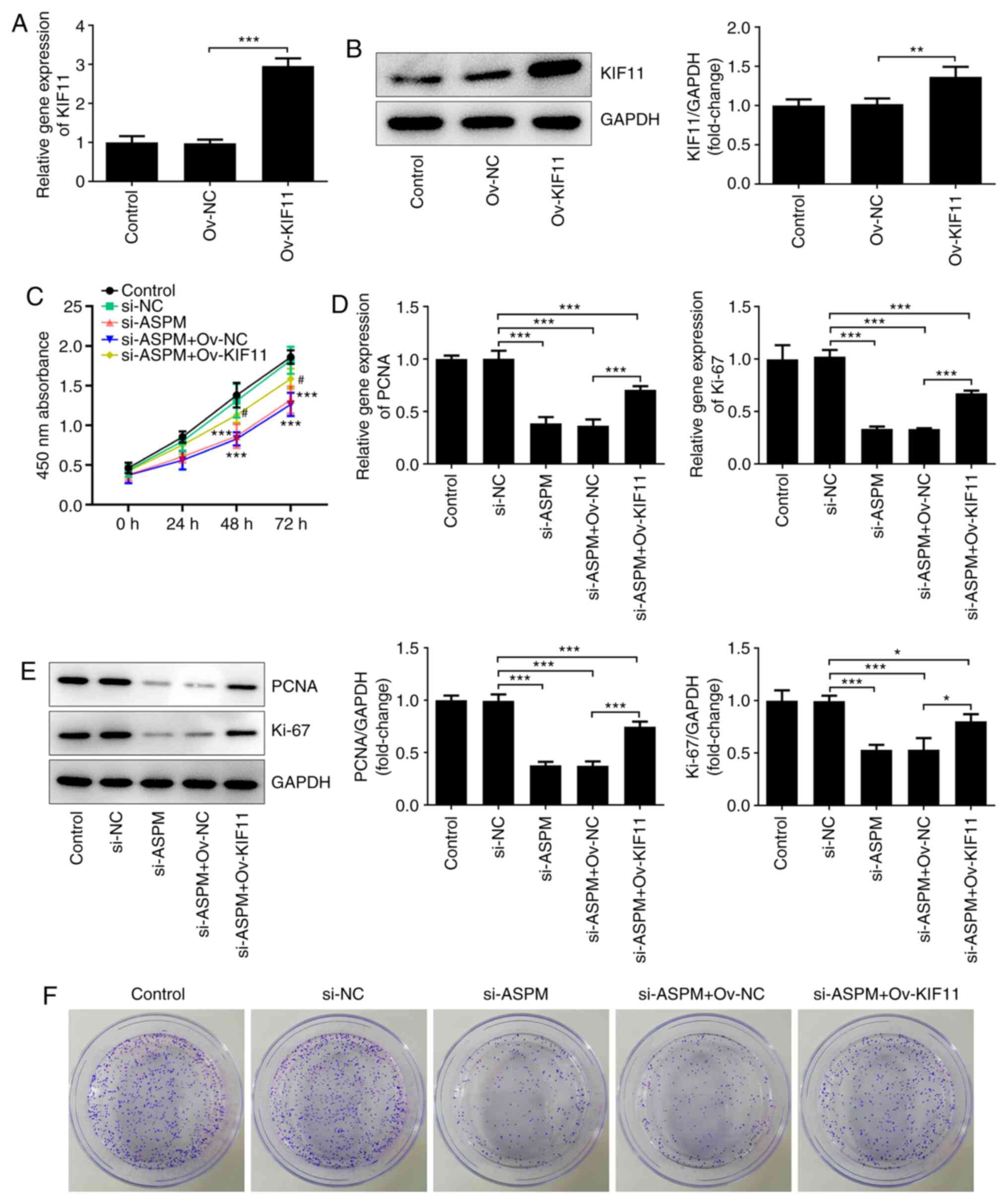
ASPM after interference using RT-qPCR (Fig. 2A). Cell viability was detected using
a CCK-8 assay, and it was revealed that the number of viable Hep3B
cells was decreased significantly following ASPM-knockdown compared
with the si-NC group (Fig. 2B).
Western blotting was used to detect the expression levels of
proliferation-associated proteins, namely proliferating cell
nuclear antigen (PCNA) and Ki-67. Compared with the si-NC group,
the expression levels of PCNA and Ki-67 were decreased
significantly after interference with ASPM expression (Fig. 2C and D). In addition, using a colony formation
assay, it was demonstrated that interference with ASPM expression
markedly inhibited the colony formation ability of Hep3B cells
(Fig. 2E). The aforementioned
experiments collectively indicated that interference with ASPM
expression inhibited the proliferation of HCC cells.
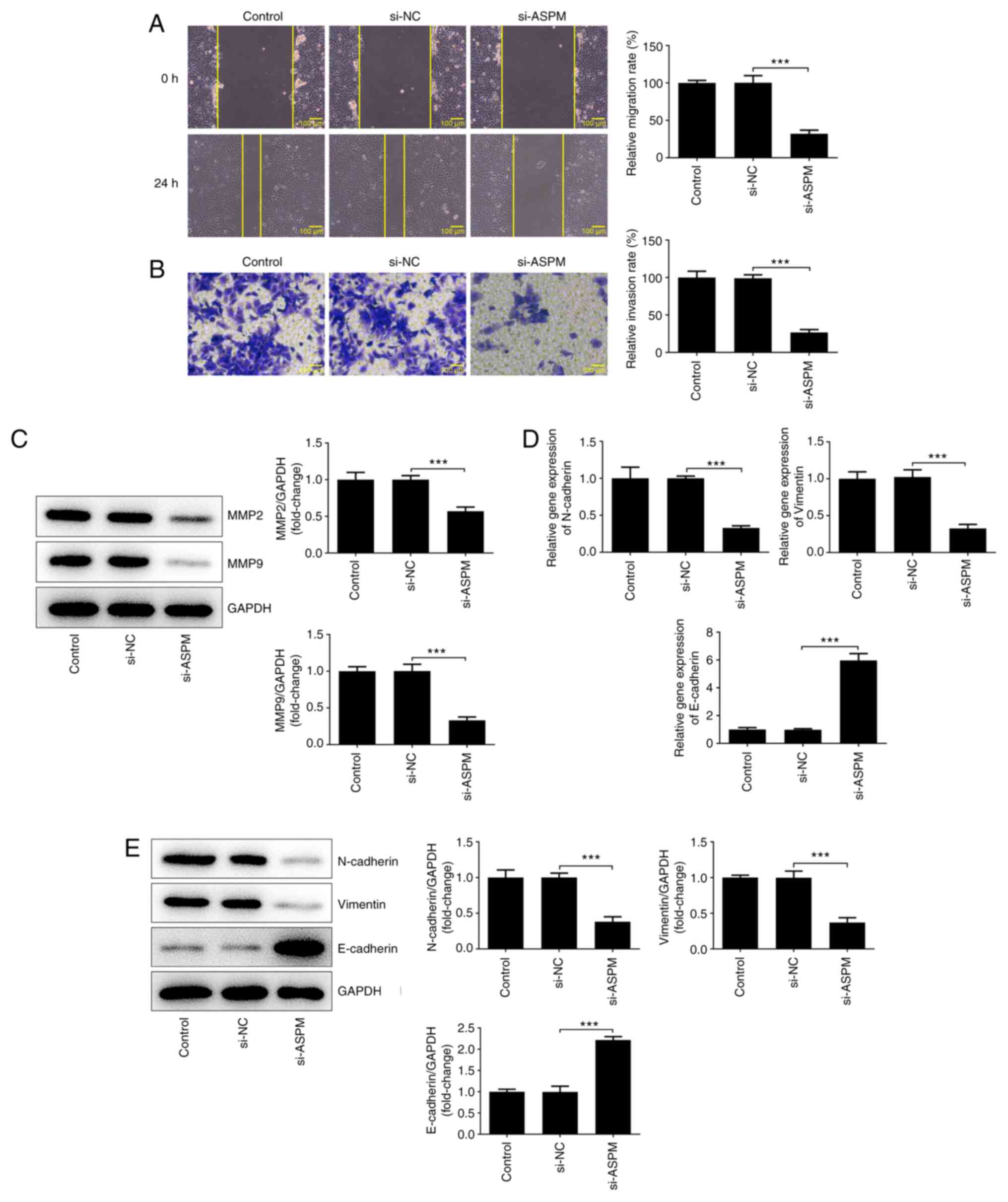
Interference with ASPM inhibits the
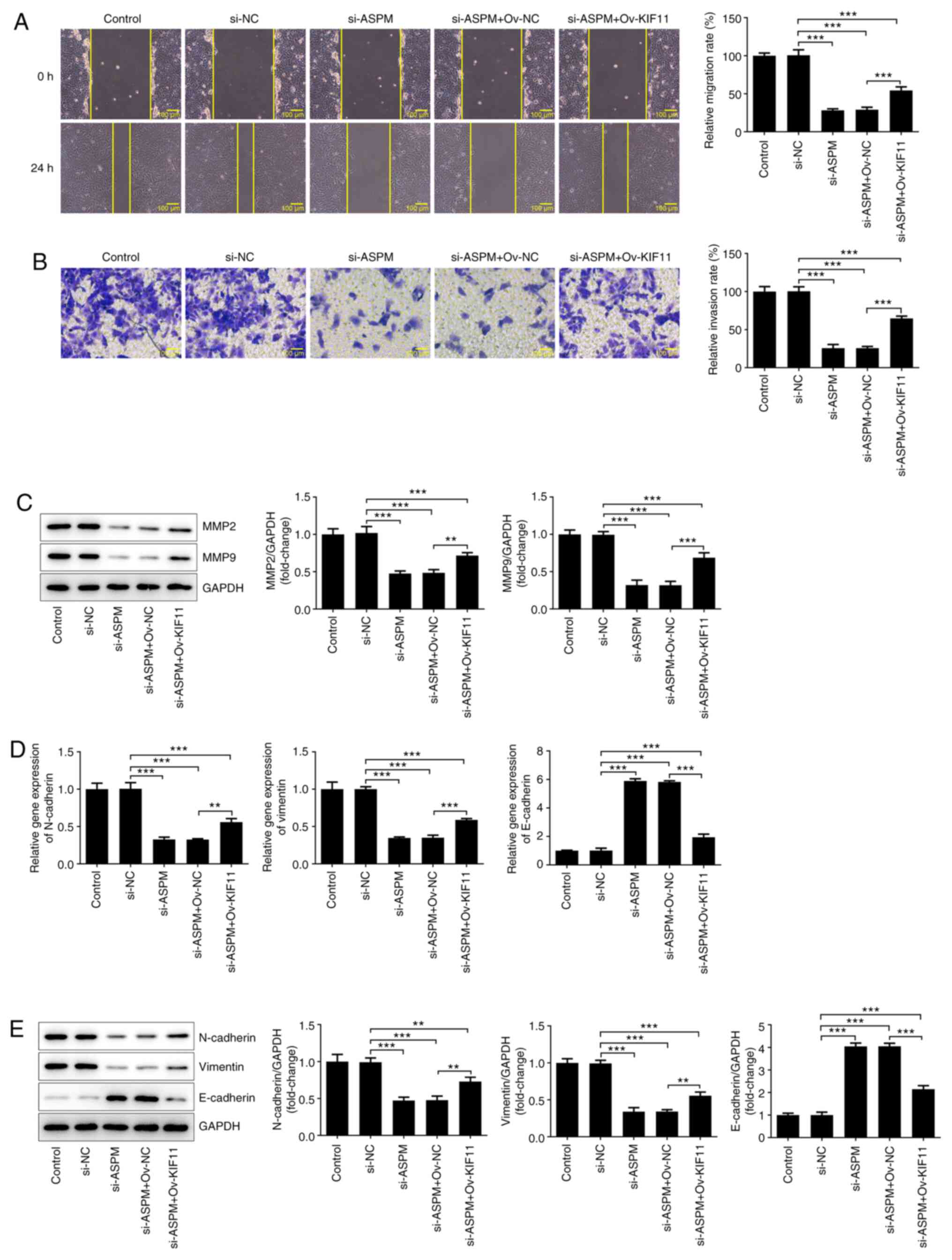
migration, invasion and EMT of HCC cells
The high metastatic propensity of HCC serves a key
role in the malignant progression of HCC (19). Therefore, the effects of ASPM on the
migration, invasion and EMT of HCC cells were examined. The results
of the wound healing assay revealed a significant decrease in the
migratory ability of the si-ASPM group compared with that of the
si-NC group (Fig. 3A). The
Transwell assay demonstrated a significant decrease in the invasive
ability of HCC cells after interference with ASPM expression
(Fig. 3B). MMP2 and MMP9 both serve
an important role in the invasion and metastasis of tumor cells
(31). Therefore, their protein
expression levels were detected by western blotting, and it was
revealed that the expression levels of these two proteins were
significantly downregulated after ASPM-knockdown (Fig. 3C). The invasion and metastasis of
tumors are often accompanied by the occurrence of EMT. Therefore,
the present study examined the expression levels of
N-cadherin/vimentin and E-cadherin using RT-qPCR and western
blotting, and the results revealed that the expression levels of
N-cadherin/vimentin were decreased, while those of E-cadherin were
elevated following ASPM-knockdown (Fig.
3D and E). The aforementioned
experimental results indicated that interference with ASPM
expression inhibited the migration, invasion and EMT of HCC
cells.
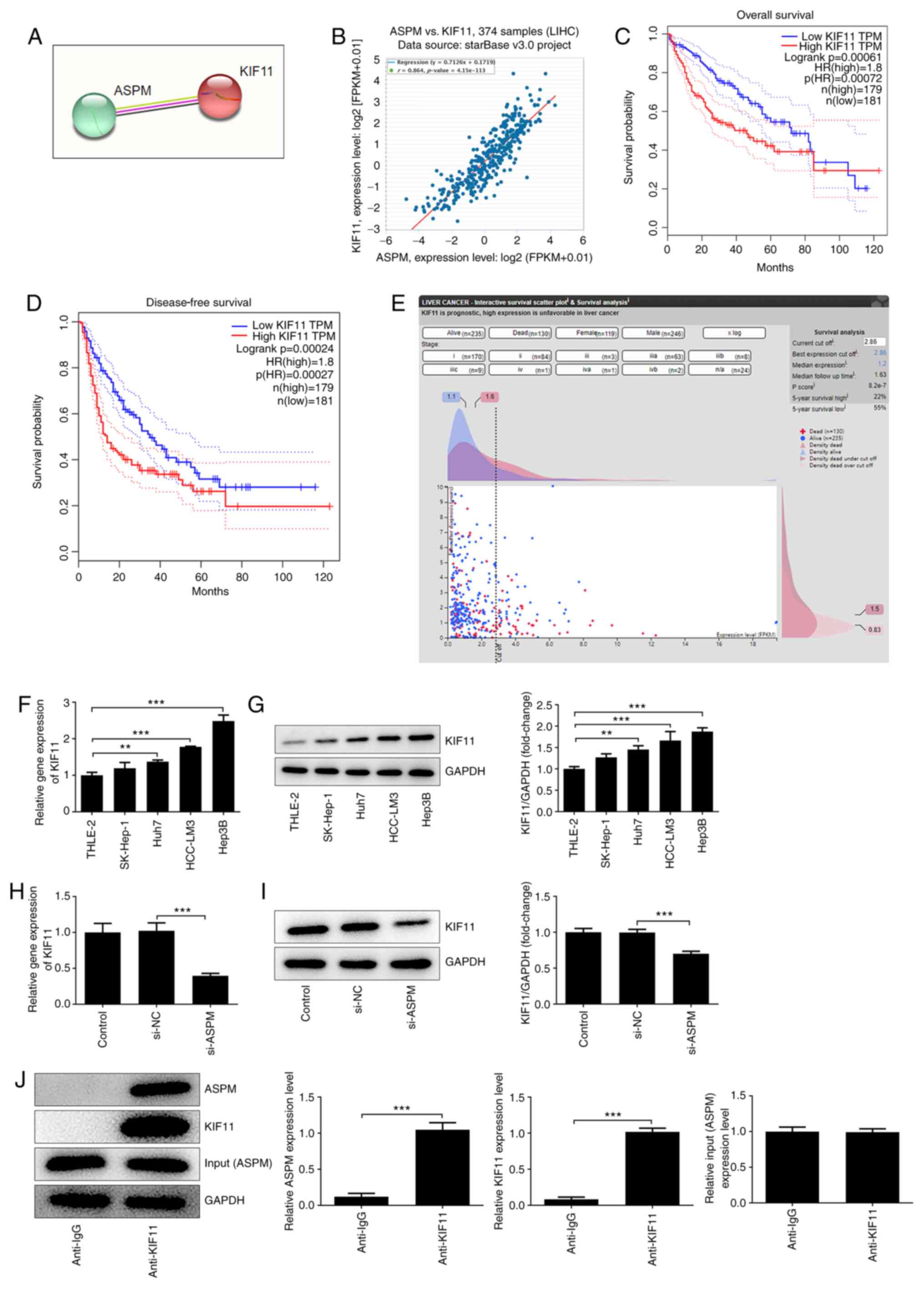
Interference with ASPM inhibits KIF11
expression in HCC cells
To further explore the mechanism of ASPM acting on
HCC, STRING and starBase were used to predict the possible
interaction between ASPM and KIF11 (Fig. 4A and B). The starBase database indicated an R
value of 0.864 between ASPM and KIF11, suggesting a distinct
positive correlation between the two (Fig. 4B). Additionally, GEPIA database
analysis suggested that the overall and disease-free survival of
patients with high KIF11 expression were significantly lower than
those of patients with low KIF11 expression (Fig. 4C and D). Additionally, the HPA database analysis
revealed that KIF11 expression was closely associated with the
prognosis of patients (Fig. 4E),
and may serve as an independent prognostic factor analogous to ASPM
for patients with HCC. These results indicated that there exists a
strong association between ASPM and KIF11, both of which are
closely associated with the development of HCC. To further study
the association between ASPM and KIF11, RT-qPCR and western
blotting were performed to detect the expression levels of KIF11 in
normal liver cells and HCC cells after interference with ASPM
expression. The results demonstrated that KIF11 expression was
significantly higher in HCC cell lines compared with in normal
liver cells (Fig. 4F and G), whereas it was significantly lower
after interference with ASPM expression (Fig. 4H and I). CO-IP was conducted to verify whether
ASPM and KIF11 interacted with each other, and the results revealed
that endogenous ASPM and endogenous KIF11 co-precipitated in Hep3B
cells (Fig. 4J). The aforementioned
experiments indicated an interaction between ASPM and KIF11, and an
inhibitory effect of ASPM-knockdown on KIF11 expression in HCC cell
lines.
Interference with ASPM inhibits the
proliferation of HCC cells via KIF11
To verify that ASPM exerted an effect via KIF11, the
KIF11 overexpression vector (Ov-KIF11) was first constructed. The
results of RT-qPCR and western blotting demonstrated that the mRNA
and protein expression levels of KIF11 were significantly increased
after transfection with the KIF11 overexpression vector (Fig. 5A and B). In addition, si-ASPM, Ov-KIF11 and
their respective NCs were co-transfected into Hep3B cells, and
their proliferation levels were then detected using a CCK-8 assay.
The results revealed that interference with ASPM expression
significantly inhibited the proliferative ability of HCC cells,
while this effect was reversed by simultaneous transfection of
si-ASPM and Ov-KIF11 (Fig. 5C).
RT-qPCR and western blotting revealed that the simultaneous
transfection of si-ASPM and Ov-KIF11 restored the mRNA and protein
expression levels of proliferation-associated genes PCNA and Ki-67
(Fig. 5D and E), and this trend was consistent with the
results of the CCK-8 assay. Furthermore, the colony formation assay
demonstrated that simultaneous transfection of si-ASPM and Ov-KIF11
reversed the inhibitory effect of si-ASPM on the colony formation
capacity of Hep3B cells (Fig. 5F).
Overall, the aforementioned experimental results indicated that
ASPM-knockdown inhibited the proliferation and colony formation
ability of HCC cells via KIF11.
Interference with ASPM inhibits the
migration, invasion and EMT of HCC cells via KIF11
To further determine the mechanism, si-ASPM,
Ov-KIF11 and their respective NCs were co-transfected into Hep3B
cells. Wound healing and Transwell assays were utilized to detect
the changes in cell invasion and migration. The results
demonstrated that simultaneous transfection of si-ASPM and Ov-KIF11
reversed the inhibitory effect of si-ASPM on the migration and
invasion of Hep3B cells (Fig. 6A
and B). The detection of MMP2 and
MMP9 expression by western blotting demonstrated a significant
decrease in the expression levels of MMP2 and MMP9 after
interference with ASPM expression, whereas these expression levels
were partly restored after simultaneous transfection of si-ASPM and
Ov-KIF11 (Fig. 6C). The expression
levels of EMT-associated proteins were detected by RT-qPCR and
western blotting, revealing that, compared with the si-ASPM group,
simultaneous transfection of si-ASPM and Ov-KIF11 restored the
expression levels of N-cadherin and vimentin to some degree, in
addition to inhibiting E-cadherin expression (Fig. 6D and E). These experiments suggested that
interference with ASPM inhibited the migration, invasion and EMT of
HCC cells via KIF11.
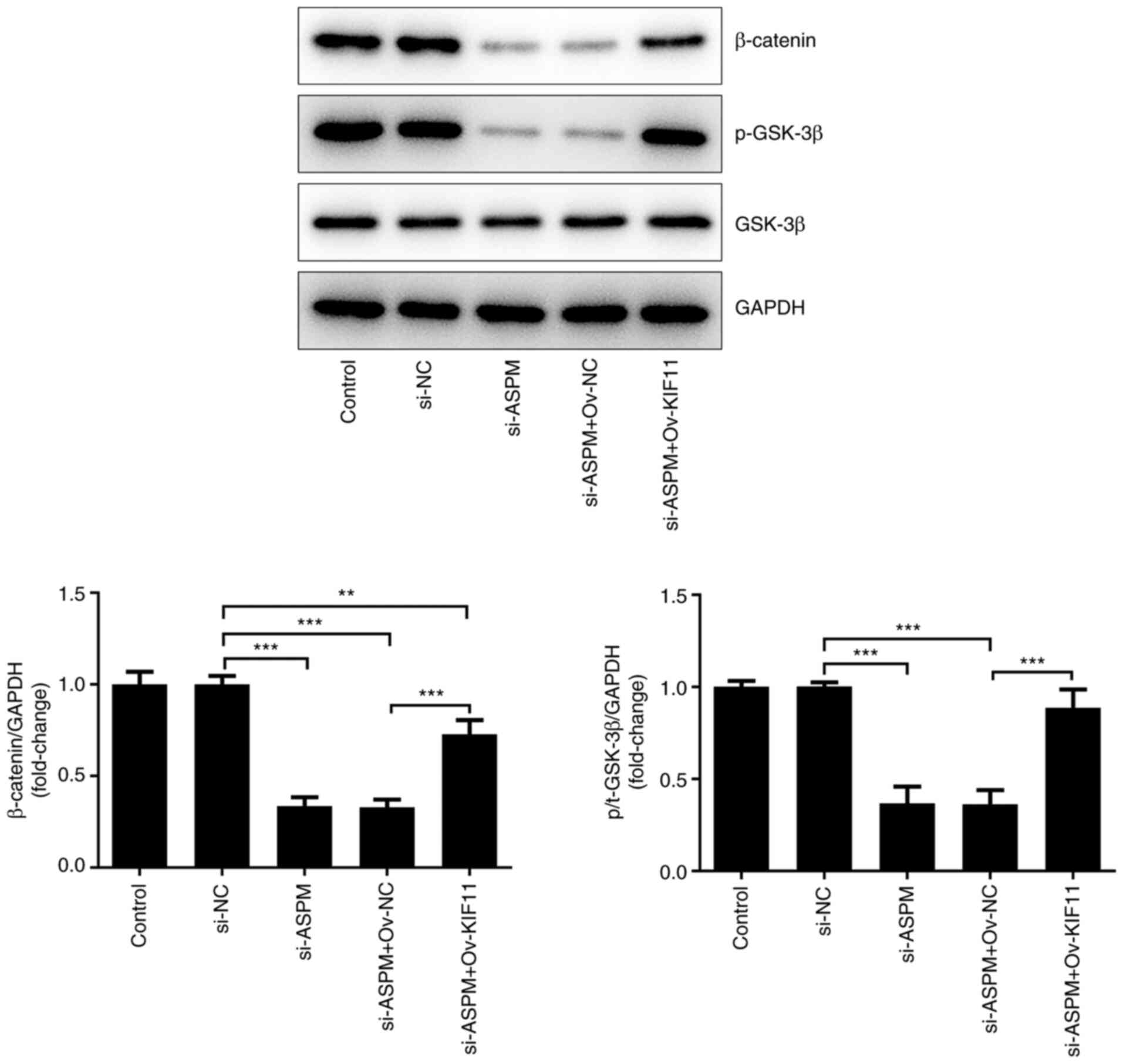
ASPM is involved in the Wnt/β-catenin
signaling pathway by regulating KIF11
A previous study has reported the ability of ASPM to
promote glioma growth through the Wnt/β-catenin signaling pathway
(32). In addition, Pei et
al (33) revealed that
silencing of KIF11 markedly decreased the self-renewal ability of
breast cancer cells by β-catenin. Based on these findings, it was
hypothesized that ASPM could regulate the occurrence and
development of HCC via the KIF11-mediated Wnt/β-catenin signaling
pathway. To verify the aforementioned hypothesis, the expression
levels of proteins associated with the Wnt/β-catenin signaling
pathway, namely β-catenin, phosphorylated (p-)GSK-3β and GSK-3β,
were detected after co-transfection of si-ASPM, Ov-KIF11 and their
respective NCs into Hep3B cells. The results demonstrated that the
levels of β-catenin and p-GSK-3 were significantly decreased after
interference with ASPM, indicating that interference with ASPM
expression silenced the Wnt/β-catenin signaling pathway.
Additionally, the levels of β-catenin and p-GSK-3β were restored
compared with the si-ASPM + Ov-NC group after simultaneous
transfection of si-ASPM and Ov-KIF11 (Fig. 7). The aforementioned experiments
indicated that ASPM may be involved in the Wnt/β-catenin signaling
pathway through the regulation of KIF11 and thereby may regulate
the malignant progression of HCC.
Discussion
HCC ranks among the top five most common malignant
tumors worldwide (34). Due to the
insidiousness of this disease, patients are often diagnosed with
advanced HCC at their initial examination in the hospital,
resulting in a poor prognosis (35). At present, chemotherapy and
immunotherapy are comparatively the best options for the treatment
of HCC (36). Therefore, it is
particularly important to perform more in-depth research on the
pathogenesis of HCC, with the aim of facilitating the development
of immunotherapy for the treatment of HCC.
ASPM is highly expressed in various types of cancer,
including breast (13), bladder
(12) and ovarian cancer (16). The present study revealed that ASPM
is highly expressed in liver cancer tissues and cells, which was
consistent with the findings of other studies (19-21).
In vitro experiments revealed that interference with ASPM
expression inhibited the proliferation, migration, invasion and EMT
of HCC cells, suggesting that the upregulation of ASPM in HCC may
promote the malignant progression of HCC. In addition, the present
study further revealed that KIF11 and ASPM are associated using
STRING and starBase database analysis, suggesting that KIF11 and
ASPM may interact directly.
KIF11 acts as an oncogene in tumors and is
associated with a poor prognosis in breast cancer (26). Additionally, it promotes cell
invasion, proliferation and self-renewal in glioblastoma (27). A number of studies have reported
that KIF11 may serve as a biological diagnostic and prognostic
marker for numerous types of tumor, including oral (37), non-small cell lung (14,38),
ovarian (39) and bladder cancer
(40). Liu et al (41) observed high KIF11 expression in HCC
tissues, which is strongly associated with liver cirrhosis and
tumor stages, and can be considered as a biomarker for poor
prognosis of HCC. The present study revealed that KIF11 was highly
expressed in HCC through bioinformatics analysis, and that it was
associated with HCC development, which was consistent with the
study by Liu et al (41).
The CO-IP assay demonstrated that ASPM could directly bind to
KIF11, and overexpression of KIF11 reversed the inhibitory effect
of ASPM-knockdown on proliferation, migration, invasion and EMT of
HCC cells to some extent.
The Wnt/β-catenin signaling pathway is a highly
conserved and tightly controlled molecular mechanism, which has
been identified to be abnormally activated in HCC and to exert a
major influence over the occurrence and development of this type of
cancer (42,43). β-catenin is an important protein
molecule in the Wnt signaling pathway, and its expression levels in
HCC tissues directly affect the activation degree of the
Wnt/β-catenin signaling pathway (44). At present, a number of studies have
validated the effectiveness of Wnt/β-catenin antagonists in
dampening the malignant progression of HCC (45,46).
The development of novel drugs targeting the Wnt/β-catenin
signaling pathway has now become one of the main focuses of
research for the treatment of HCC. It was revealed that the levels
of β-catenin and p-GSK-3β were decreased after interference with
ASPM expression, silencing the Wnt/β-catenin signaling pathway.
However, when KIF11 was overexpressed at the same time, the
β-catenin and p-GSK-3β levels were restored, which indicated that
KIF11 was involved in the Wnt/β-catenin signaling pathway.
Additionally, the current results demonstrated that ASPM regulated
the involvement of KIF11 in the Wnt/β-catenin signaling pathway to
regulate the malignant progression of HCC.
In conclusion, the present study identified that
ASPM was highly expressed in HCC tissues, and that ASPM-knockdown
inhibited the proliferation, migration, invasion and EMT of HCC
cells. Additionally, ASPM was involved in the Wnt/β-catenin
signaling pathway through KIF11, thereby promoting the
proliferation, migration, invasion and EMT of HCC cells.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by The National Natural
Science Foundation of China (grant no. 81871260).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BW designed and performed the experiments, and made
considerable contributions to the manuscript writing. CH performed
the experiments and analyzed the data. LK conceived the study,
guided the experiments and revised the manuscript. BW and CH
confirmed the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Miller K and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Anwanwan D, Singh S, Singh S, Saikam V and
Singh R: Challenges in liver cancer and possible treatment
approaches. Biochim Biophys Acta Rev Cancer.
1873(188314)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Craig AJ, von Felden J, Garcia-Lezana T,
Sarcognato S and Villanueva A: Tumour evolution in hepatocellular
carcinoma. Nat Rev Gastroenterol Hepatol. 17:139–152.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Nie J, Lin B, Zhou M, Wu L and Zheng T:
Role of ferroptosis in hepatocellular carcinoma. J Cancer Res Clin
Oncol. 144:2329–2337. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Greten TF, Lai CW, Li G and
Staveley-O'Carroll KF: Targeted and immune-based therapies for
hepatocellular carcinoma. Gastroenterology. 156:510–524.
2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Huang F, Wang BR and Wang YG: Role of
autophagy in tumorigenesis, metastasis, targeted therapy and drug
resistance of hepatocellular carcinoma. World J Gastroenterol.
24:4643–4651. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kumar A, Blanton S, Babu M, Markandaya M
and Girimaji S: Genetic analysis of primary microcephaly in Indian
families: novel ASPM mutations. Clin Genet. 66:341–348.
2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Mayya V, Lundgren DH, Hwang SI, Rezaul K,
Wu L, Eng JK, Rodionov V and Han DK: Quantitative phosphoproteomic
analysis of T cell receptor signaling reveals system-wide
modulation of protein-protein interactions. Sci Signal.
2(ra46)2009.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Pai VC, Hsu CC, Chan TS, Liao WY, Chuu CP,
Chen WY, Li CR, Lin CY, Huang SP, Chen LT and Tsai KK: ASPM
promotes prostate cancer stemness and progression by augmenting
Wnt-Dvl-3-β-catenin signaling. Oncogene. 38:1340–1353.
2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Hsu CC, Liao WY, Chan TS, Chen WY, Lee CT,
Shan YS, Huang PJ, Hou YC, Li CR and Tsai KK: The differential
distributions of ASPM isoforms and their roles in Wnt signaling,
cell cycle progression, and pancreatic cancer prognosis. J Pathol.
249:498–508. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen Q, Hu J, Deng J, Fu B and Guo J:
Bioinformatics analysis identified key molecular changes in bladder
cancer development and recurrence. Biomed Res Int.
2019(3917982)2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Xu Z, Zhang Q, Luh F, Jin B and Liu X:
Overexpression of the ASPM gene is associated with aggressiveness
and poor outcome in bladder cancer. Oncol Lett. 17:1865–1876.
2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tang J, Lu M, Cui Q, Zhang D, Kong D, Liao
X, Ren J, Gong Y and Wu G: Overexpression of ASPM, CDC20, and TTK
confer a poorer prognosis in breast cancer identified by gene
co-expression network analysis. Front Oncol. 9(310)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu X, Liu X, Li J and Ren F:
Identification and integrated analysis of key biomarkers for
diagnosis and prognosis of non-small cell lung cancer. Med Sci
Monit. 25:9280–9289. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Xie JJ, Zhuo YJ, Zheng Y, Mo RJ, Liu ZZ,
Li BW, Cai ZD, Zhu XJ, Liang YX, He HC and Zhong WD: High
expression of ASPM correlates with tumor progression and predicts
poor outcome in patients with prostate cancer. Int Urol Nephrol.
49:817–823. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Alsiary R, Brüning-Richardson A, Bond J,
Morrison EE, Wilkinson N and Bell SM: Deregulation of microcephalin
and ASPM expression are correlated with epithelial ovarian cancer
progression. PLoS One. 9(e97059)2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Brüning-Richardson A, Bond J, Alsiary R,
Richardson J, Cairns DA, McCormack L, Hutson R, Burns P, Wilkinson
N, Hall GD, et al: ASPM and microcephalin expression in epithelial
ovarian cancer correlates with tumour grade and survival. Br J
Cancer. 104:1602–1610. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Gao ZY, Yu F, Jia HX, Ye Z and Yao SJ:
ASPM predicts poor prognosis and regulates cell proliferation in
bladder cancer. Kaohsiung J Med Sci. 36:1021–1029. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Lin SY, Pan HW, Liu SH, Jeng YM, Hu FC,
Peng SY, Lai PL and Hsu HC: ASPM is a novel marker for vascular
invasion, early recurrence, and poor prognosis of hepatocellular
carcinoma. Clin Cancer Res. 14:4814–4820. 2008.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Xue JM, Liu Y, Wan LH and Zhu YX:
Comprehensive analysis of differential gene expression to identify
common gene signatures in multiple cancers. Med Sci Monit.
26(e919953)2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhou Z, Li Y, Hao H, Wang Y, Zhou Z, Wang
Z and Chu X: Screening Hub genes as prognostic biomarkers of
hepatocellular carcinoma by bioinformatics analysis. Cell
Transplant. 28 (Suppl 1):76S–86S. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Rapley J, Nicolàs M, Groen A, Regué L,
Bertran MT, Caelles C, Avruch J and Roig J: The NIMA-family kinase
Nek6 phosphorylates the kinesin Eg5 at a novel site necessary for
mitotic spindle formation. J Cell Sci. 121:3912–3921.
2008.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wakana Y, Villeneuve J, van Galen J,
Cruz-Garcia D, Tagaya M and Malhotra V: Kinesin-5/Eg5 is important
for transport of CARTS from the trans-Golgi network to the cell
surface. J Cell Biol. 202:241–250. 2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Güneş N, Taşdemir E, Jeffery H, Yetik H,
Ostergaard P and Tüysüz B: A novel mutation of KIF11 in a
Child with 22q11.2 deletion syndrome associated with MCLMR. Mol
Syndromol. 9:266–270. 2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hu H, Xiao X, Li S, Jia X, Guo X and Zhang
Q: KIF11 mutations are a common cause of autosomal dominant
familial exudative vitreoretinopathy. Br J Ophthalmol. 100:278–283.
2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhou J, Chen WR, Yang LC, Wang J, Sun JY,
Zhang WW, He ZY and Wu SG: KIF11 functions as an oncogene and is
associated with poor outcomes from breast cancer. Cancer Res Treat.
51:1207–1221. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Venere M, Horbinski C, Crish JF, Jin X,
Vasanji A, Major J, Burrows AC, Chang C, Prokop J, Wu Q, et al: The
mitotic kinesin KIF11 is a driver of invasion, proliferation, and
self-renewal in glioblastoma. Sci Transl Med.
7(304ra143)2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Tang Z, Kang B, Li C, Chen T and Zhang Z:
GEPIA2: An enhanced web server for large-scale expression profiling
and interactive analysis. Nucleic Acids Res. 47:W556–W560.
2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Qin L, Liao L, Redmond A, Young L, Yuan Y,
Chen H, O'Malley BW and Xu J: The AIB1 oncogene promotes breast
cancer metastasis by activation of PEA3-mediated matrix
metalloproteinase 2 (MMP2) and MMP9 expression. Mol Cell Biol.
28:5937–5950. 2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Chen X, Huang L, Yang Y, Chen S, Sun J, Ma
C, Xie J, Song Y and Yang J: ASPM promotes glioblastoma growth by
regulating G1 restriction point progression and Wnt-β-catenin
signaling. Aging (Albany NY). 12:224–241. 2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Pei YY, Li GC, Ran J, Wan XH, Wei FX and
Wang L: Kinesin family member 11 enhances the self-renewal ability
of breast cancer cells by participating in the Wnt/β-catenin
pathway. J Breast Cancer. 22:522–532. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
De Stefano F, Chacon E, Turcios L, Marti F
and Gedaly R: Novel biomarkers in hepatocellular carcinoma. Dig
Liver Dis. 50:1115–1123. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Xie QL, Liu Y and Zhu Y: Chromosome region
maintenance 1 expression and its association with clinical
pathological features in primary carcinoma of the liver. Exp Ther
Med. 12:59–68. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Chen J, Cao J, Wang P and He X:
NT5DC2 is a novel prognostic marker in human hepatocellular
carcinoma. Oncol Lett. 20(70)2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Daigo K, Takano A, Thang PM, Yoshitake Y,
Shinohara M, Tohnai I, Murakami Y, Maegawa J and Daigo Y:
Characterization of KIF11 as a novel prognostic biomarker and
therapeutic target for oral cancer. Int J Oncol. 52:155–165.
2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Schneider MA, Christopoulos P, Muley T,
Warth A, Klingmueller U, Thomas M, Herth FJ, Dienemann H, Mueller
NS, Theis F and Meister M: AURKA, DLGAP5, TPX2, KIF11 and CKAP5:
Five specific mitosis-associated genes correlate with poor
prognosis for non-small cell lung cancer patients. Int J Oncol.
50:365–372. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Xu Z, Zhou Y, Cao Y, Dinh TL, Wan J and
Zhao M: Identification of candidate biomarkers and analysis of
prognostic values in ovarian cancer by integrated bioinformatics
analysis. Med Oncol. 33(130)2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Pan S, Zhan Y, Chen X, Wu B and Liu B:
Identification of biomarkers for controlling cancer stem cell
characteristics in bladder cancer by network analysis of
transcriptome data stemness indices. Front Oncol.
9(613)2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Liu C, Zhou N, Li J, Kong J, Guan X and
Wang X: Eg5 overexpression is predictive of poor prognosis in
hepatocellular carcinoma patients. Dis Markers.
2017(2176460)2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Perugorria MJ, Olaizola P, Labiano I,
Esparza-Baquer A, Marzioni M, Marin JJG, Bujanda L and Banales JM:
Wnt-β-catenin signalling in liver development, health and disease.
Nat Rev Gastroenterol Hepatol. 16:121–136. 2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Wang W, Smits R, Hao H and He C:
Wnt/β-catenin signaling in liver cancers. Cancers (Basel).
11(926)2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Yang G, Shen T, Yi X, Zhang Z, Tang C,
Wang L, Zhou Y and Zhou W: Crosstalk between long non-coding RNAs
and Wnt/β-catenin signalling in cancer. J Cell Mol Med.
22:2062–2070. 2018.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Shi XD, Yu XH, Wu WR, Xu XL, Wang JY, Xu
LB, Zhang R and Liu C: Dickkopf-1 expression is associated with
tumorigenity and lymphatic metastasis in human hilar
cholangiocarcinoma. Oncotarget. 7:70378–70387. 2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Yu B, Yang X, Xu Y, Yao G, Shu H, Lin B,
Hood L, Wang H, Yang S, Gu J, et al: Elevated expression of DKK1 is
associated with cytoplasmic/nuclear beta-catenin accumulation and
poor prognosis in hepatocellular carcinomas. J Hepatol. 50:948–957.
2009.PubMed/NCBI View Article : Google Scholar
|