Introduction
Lower back pain affects 80-85% of adults at a
certain period during their lifetime (1,2).
Intervertebral disc degeneration (IVDD) is generally considered to
be the main factor contributing to lower back pain (2). The intervertebral disc (IVD) is
composed of annulus fibrosus, nucleus pulposus (NP) and
cartilaginous endplates. NP is the most hydrated proteoglycan
region of the IVD, comprising extracellular matrix and nucleus
pulposus cells (NPCs). These enable the NP to maintain osmotic
pressure and therefore to withstand compressive forces of the IVD.
During degeneration, there are alterations in collagen proportions,
the relative composition of proteoglycans and a greater water loss
from the NP, resulting in loss of the hydrostatic properties of the
disk (2). Several studies suggest
that apoptosis and senescence of NPCs are associated with disc
degeneration (3,4). Diabetes has been revealed to be a
clinical risk factor of IVDD (3,5).
Studies have shown that high-glucose (HG) concentrations may induce
disc cell apoptosis and senescence, as well as abnormal matrix
metabolism (4-7).
Telomere attrition is associated with this
phenomenon; when the telomere is shortened, it results in
senescence and ultimately apoptosis, which further deteriorates the
condition of the cells and the IVD (8). Studies have indicated that diabetes
accelerates telomere attrition in type 2 diabetic patients
(9,10). Studies have also demonstrated that
telomere attrition and lower telomerase expression is involved in
diverse types of diseases, including IVDD. Therefore, telomerase is
considered a potential therapeutic target for these diseases
(8,9). Telomerase reverse transcriptase
(TERT), together with the telomerase RNA component (TERC), is the
most important unit of the telomerase complex, though TERT is able
to perform its function without the presence of TERC (11). TERT activation may result in a
protective effect against degeneration and increase the resistance
of cells to glucose-mediated stress (12). Lower expression levels of TERT have
been implicated in various disorders, including degenerative
diseases (13). A study by Graham
et al (14) revealed that
TERT significantly suppresses p16 expression. A study by Chung
et al (12) on NPCs and
gene therapy considered telomerase to be a therapeutic target.
However, to the best of our knowledge, its role in HG-IVDD is
unknown.
Extracts from Astragalus roots are recognized
as traditional medicines and foods (15). Cycloastragenol (CAG) is a bioactive
molecule isolated from various species in the Astragalus
genus, which are used as a dietary ingredients (16,17).
Astragalosides are chemical compounds isolated from the dried root
of the fundamental Chinese herb Astragalus membranaceus.
Astragaloside IV (AG-IV) has numerous pharmaceutical properties,
such as anti-inflammation, anti-insulin resistance, antitumor and
neuroprotection (18-21).
CAG, AG-IV and other associated molecules have been identified as
small-molecule telomerase activators (16). Previous studies have demonstrated
that CAG and AG-IV are able to alleviate and reverse a number of
degenerative diseases, such as metabolic syndrome, arthritis and
age-related macular degeneration, by activating telomerase
(18,19,22).
However, the effects of both of these drugs on HG-IVDD are unknown.
Therefore, the present study hypothesized that they may postpone
the progression of HG-IVDD by upregulating TERT.
The present study demonstrated the expression
profile of TERT in NPCs and that CAG and AG-IV can activate TERT,
indicating its therapeutic potential in HG IVDD. To the best of our
knowledge, this is the first report to describe the link between
TERT and HG-induced IVDD, and to also introduce CAG and AG-IV as
pharmacological agonists of TERT in the treatment of IVDD.
Materials and methods
Ethics statement
The present study was carried out in accordance with
the Guide for the Care and Use of Laboratory Animals of the
National Institutes of Health (NIH Publications No. 8023, revised
1978). The animal protocol was approved by the Ethics Committee of
Wenzhou Medical University (approval no. wydw2014-0129; Wenzhou,
China).
Reagents and antibodies
CAG (CAS 84605-18-5; cat. no. ZL20170428) and
Astragaloside IV (CAS 84687-43-4; cat. no. ZL20170415) were
purchased from Nanjing Zelang Medical Technology Co., Ltd. with
≥98% purity. The monoclonal anti-p16INK4a/CDKN2 antibody
(cat. no. SAB5300498), D-glucose, T-hydro tert-butyl hydroperoxide
(TBHP) and type II collagenase were from Sigma-Aldrich (Merck
KGaA). Monoclonal anti-cleaved caspase 3 (c-C3) antibody (cat. no.
9664) was supplied by Cell Signaling Technology, Inc. Monoclonal
anti-BAX antibody (cat. no. ab232479) was acquired from Abcam.
Monoclonal anti-Bcl-2 antibody (cat. no. 625509) was obtained from
R&D Systems, Inc. Monoclonal anti-α tubulin antibody (cat. no.
66031-1-Ig) was obtained from ProteinTech Group, Inc. Polyclonal
anti-TERT antibody (cat. no. ABE2075) was purchased from
MilliporeSigma. Anti-rabbit and anti-mouse horseradish
peroxidase-labeled secondary antibodies (cat. nos. BL003A and
BL001A) were obtained from Biosharp Life Sciences. BCA kit, RIPA
lysis buffer and phenylmethylsulfonyl fluoride (PMSF) were
purchased from Beyotime Institute of Biotechnology. Femto Enhanced
Chemi-Luminescence substrate and primers for PCR were purchased
from Thermo Fisher Scientific, Inc. SYBR® Green system
(cat. no. RR420L) for reverse transcription-quantitative PCR
(RT-qPCR) was purchased from Takara Biotechnology Co., Ltd. DNeasy
animal blood and tissue kit (cat. no. 69506) was purchased from
Qiagen GmbH. DMEM and FBS for cell culture were purchased from
Gibco (Thermo Fisher Scientific, Inc.).
NPC isolation and cell culture
Gel-like tissues (nucleus pulposus) were collected
from the tail intervertebral discs of 35 male Sprague-Dawley rats
(age, 3-4 weeks; weight, 50-100 g), which were obtained from the
Animal Center of the Chinese Academy of Sciences. Rats were housed
individually at 23-25˚C with a 40-50% humidity under a 12 h
dark/light cycle with free access to standard chow. Before
collecting tissues, the rats were anesthetized with 50 mg/kg
nembutal. The NP tissues of the L1-L6 spinal column were collected
in sterile environment and then digested in 0.2% type II
collagenase for ~4 h at 37˚C. Sprague-Dawley rats were then
euthanized with the 200 mg/kg Nembutal (23). After washing with PBS twice, the
digested tissues were transferred to DMEM 1 g/l (5.5 mM) with 15%
FBS and 1% antibiotics (100 U/ml penicillin and 100 µg/ml
streptomycin) and placed in an incubator of 5% CO2 at
37˚C. When cells reached 80-90% confluence, the cells were passaged
following trypsinization with 0.25% Trypsin-EDTA, and cultured in
10-cm culture plates at a density of 2x105 cells/ml. The
first three passages cells were used in the experiments.
Treatment protocol of cell
culture
D-glucose was added to the DMEM (low glucose) and
then filtered through sterile 0.45-mm PVDF Membrane filter (Merck
KGaA) and a stock solution of 500 mM glucose concentration was
obtained. Then the stock solution was diluted in complete culture
media to obtain the desired concentrations of glucose (5.5, 12.5,
50, 100 and 200 mM). When 50-70% confluence was. reached, NPCs were
treated with different doses of glucose at 37˚C for varying amounts
of time (24, 48, 72 or 96 h).
CAG and AG-IV were dissolved in dimethyl sulfoxide
(DMSO) at concentrations of 500 mM, and then appropriately diluted
with complete culture medium (final DMSO concentration <1%). All
the relevant groups were pretreated with different drug
concentrations (1, 3, 5 and 10 mM) and then with HG (50 mM). All
the experiments were performed in triplicate.
Cell viability assay
NPCs were seeded in 96-well plates (5,000 cell/well)
and incubated at 37˚C for 72 h. Subsequently, the NPCs were treated
with DMEM (15% FBS) in increasing concentrations and times of
glucose, or with increasing CAG and AG-IV concentrations (0.01,
0.1, 0.5, 1, 3, 5, 10, 50 and 100 µM) for different time periods
(24, 48, 72 and 96 h). After washing the cells with PBS, 100 µl of
DMEM containing 10 µl of Cell Counting Kit-8 (CCK-8; Beyotime
Institute of Biotechnology) solution was added to each well
according to the manufacturer's protocol. The plate was then
incubated for ~1 h at 37˚C. The optical density of the wells was
measured at 450 nm using a microplate reader.
Western blotting
NPCs were lysed on ice-cold RIPA lysis buffer with 1
mM PMSF. Protein concentrations of samples were measured using the
BCA protein assay kit. Samples with equal amounts of protein (40
µg) were then separated by 12-15% SDS-PAGE and transferred to a
PVDF membrane (MilliporeSigma). Membranes were blocked with 5%
non-fat milk for 2 h at room temperature. The bands were probed
with primary antibodies specific to p16 (1:500), c-C3 (1:1,000),
BAX (1:1,000), Bcl-2 (1:1,000), α-Tubulin (1:1,000) and TERT
(1:1,000) overnight at 4˚C, before being incubated with respective
anti-rabbit (1:1,000) and anti-mouse (1:1,000) horseradish
peroxidase-labeled secondary antibodies for 2 h at room
temperature. Blots were visualized using ECL (Thermo Fisher
Scientific, Inc.). The intensity of these bands were
semi-quantified using Image Lab 3.0 software (Bio-Rad Laboratories,
Inc.).
RT-qPCR analysis
Total RNA was isolated from NPCs using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), and 1 µg of total RNA was used to synthesize cDNA.
Quantitative PCR was performed using the SYBR® Green
system (Takara Bio, Inc.). Amplification of cDNA samples was
carried out on the CFX96 real-time PCR system (Bio-Rad
Laboratories, Inc.). The reaction steps were as follows: 95˚C for 3
min, 60˚C for 45 sec and for 39 cycles. At-last melt curve
analysis: 65˚C Then 95˚C and 5˚C for 5 sec each. The relative
quantification of the target gene was normalized to β-actin, and
target gene was compared with the control sample using the
2-ΔΔCq method (24).
The primers for TERT and β-actin are listed in Table I.
 | Table IPrimer sequences for reverse
transcription-quantitative PCR. |
Table I
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Forward primer
(5'-3') | Reverse primer
(5'-3') |
|---|
| β-actin |
CCGCGAGTACAACCTTCTTG |
TGACCCATACCCACCATCAC |
| TERT |
AGTGGTGAACTTCCCTGTGG |
CAACCGCAAGACTGACAAGA |
| Telomere |
GGTTTTTGAGGGTGAGGGTGAGGG
TGAGGGTGAGGGT |
TCCCGACTATCCCTATCCCTATCCC
TATCCCTATCCCTA |
| 36B4 |
CTCACTCCATCATCAATGGATACAA |
CAGCCAGTGGGAAGGTGTAGTCA |
DNA was extracted using the DNeasy kit from Qiagen
GmbH. After the DNA was extracted from the NPCs in six-well plates
at a seeding density of 2x105 cells/ml following the
manufacturer's protocols, then the same protocol and quantification
of qPCR data were carried out as aforementioned. The primers of
Telomere and 36B4 are listed in Table
I.
Sa-β-gal staining
NPCs were seeded in 6-well plates for 72 h at a
seeding density of 2x105 cells/ml, washed twice with PBS
and fixed with 0.2% glutaraldehyde for 10 min at room temperature.
Cells were subsequently stained with X-gal staining solution
overnight at 37˚C at pH 6.0. The next day, images were captured by
using an inverted Olympus IX71 light microscope (magnification,
x200; Olympus Corporation) and the percentage of SA-β-gal-positive
cells were quantified using Image Lab 3.0 software (Bio-Rad
Laboratories, Inc.).
Flow cytometry
Apoptosis was detected using an Annexin-V apoptosis
detection kit (Becton-Dickinson and Company) as described by the
manufacturer's protocol. NPCs were seeded at a density of
2x105 cells/ml in 6-well plates and treated with
different concentrations of glucose, CAG and AG-IV at 37˚C for 72
h. Subsequently, ~106 cells from each well were
collected, washed and re-suspended in 100 µl of binding buffer with
5 µl of Annexin-V FITC and 5 µl propidium iodide for 30 min in the
dark at room temperature. Cells were analyzed using flow cytometry
(BD Biosciences; Becton, Dickinson and Company). The total
apoptosis rate (early stage apoptosis plus late stage apoptosis) of
NPCs was calculated as the percentage of the ratio observed in the
lower-right and the upper-right quadrants.
Statistical analysis
All the experiments were performed at least three
times. Results were expressed as mean ± standard deviation.
Statistical analyses were performed using GraphPad Prism (version
7.0; GraphPad Software, Inc.). Data were analyzed by one-way
analysis of variance (ANOVA) followed by the Dunnett's multiple
tests for comparison between control and treatment groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
HG concentrations adversely affect the
viability of NPCs
To investigate HG-induced cytotoxicity in the
primary NPCs, they were treated with different concentrations of
glucose for varying amounts of time. The cytotoxic effect of
glucose on NPCs was determined at various concentrations of glucose
(5.5, 12.5, 25, 50, 100 and 200 mM) for 24-96 h using a CCK-8
assay. As presented in Fig. 1,
there were significant cytotoxic effects following HG treatment on
the NPCs, especially at higher doses (50, 100 and 200 mM glucose
for 24-96 h; P<0.05). While simulating the pathophysiological
characteristics of the chronic course of diabetes, treatment for 72
h exhibited similar effects to treatment for 96 h. Thus, treatment
using 50 mM glucose for 72 h was selected for subsequent
experiments.
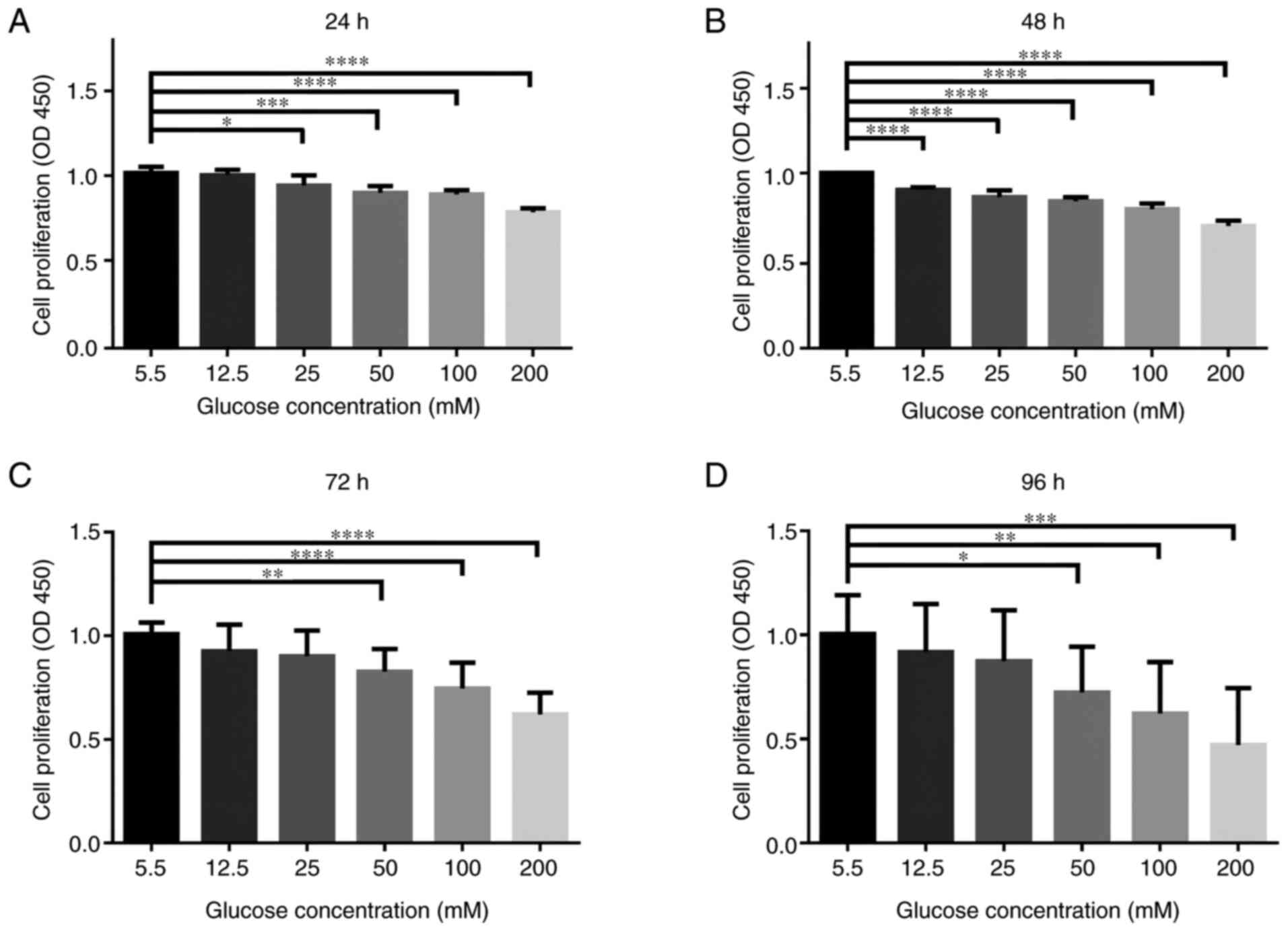 | Figure 1HG concentrations adversely affects
the cellular viability of rat NPCs. NPCs were treated with
different concentrations of glucose (5.5, 12.5, 25, 50, 100 and 200
mM) for varying durations (24, 48, 72 and 96 h) in complete DMEM
culture media. CCK-8 results of NPCs treated with different
concentrations of glucose for (A) 24 h, (B) 48 h, (C) 72 h and (D)
for 96 h. n=5/group. *P<0.05; **P<0.01;
***P<0.001; ****P<0.0001 vs. 5.5 mM.
NPCs, nucleus pulposus cells. HG, high glucose (50 mM); CCK-8, Cell
Counting Kit-8; OD, optical density. |
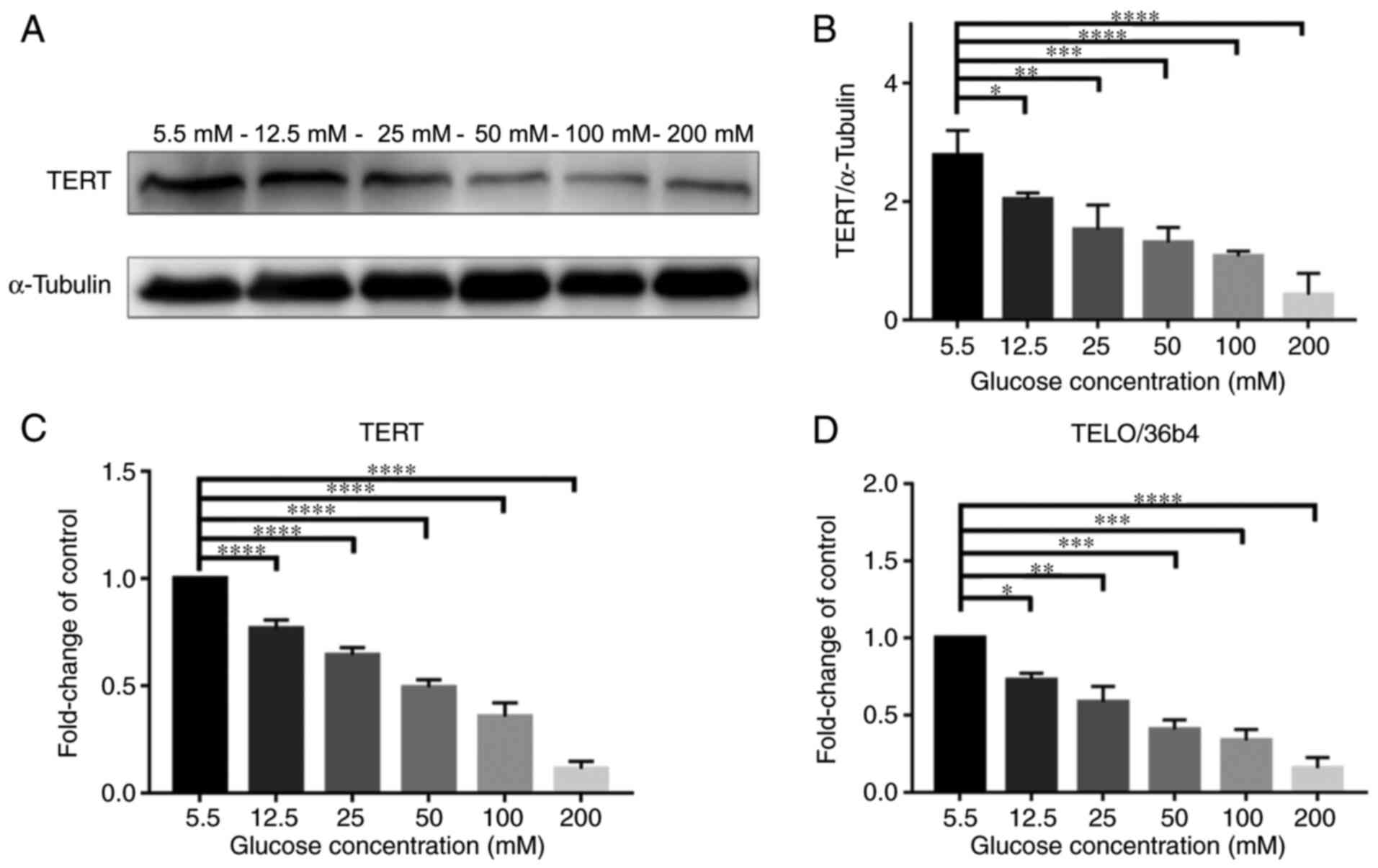
HG concentrations have adverse effects
on TERT expression and telomere length in NPCs
Fig. 2A presents
western blotting of TERT protein expression levels in NPCs when
treated with glucose at various concentrations (5.5, 12.5, 25, 50,
100 and 200 mM) for 72 h; the expression level of TERT notably
decreased with higher concentrations of glucose. Fig. 2B demonstrated that TERT protein
expression levels were significantly decreased at higher
concentrations of glucose in a dose-dependent manner
(P<0.05).
Meanwhile, as presented in Fig. 2C, the expression of mRNA TERT in
RT-qPCR was also decreased in a dose-dependent manner
(P<0.0001). In addition, as presented in Fig. 2D, the telomere length, detected
using the qPCR method by Cawthon (25), also decreased in a dose-dependent
manner (P<0.05). Telomere length was observed using qPCR, using
the Cawthon method, which is the standard method to check for the
telomere length. These results demonstrated that high levels of
glucose inhibited the expression of TERT and shortened the telomere
length.
HG concentrations promote apoptosis
and senescence of NPCs
Fig. 3A-D presents
the expression levels of apoptosis-related proteins (c-C3, BAX and
Bcl-2) in NPCs detected by western blotting after treatment with
various concentrations of glucose (5.5, 12.5, 25, 50, 100 and 200
mM) for 72 h. The expression levels of c-C3 and BAX proteins were
significantly increased in a glucose dose-dependent manner; while
Bcl-2 expression was significantly decreased (P<0.05).
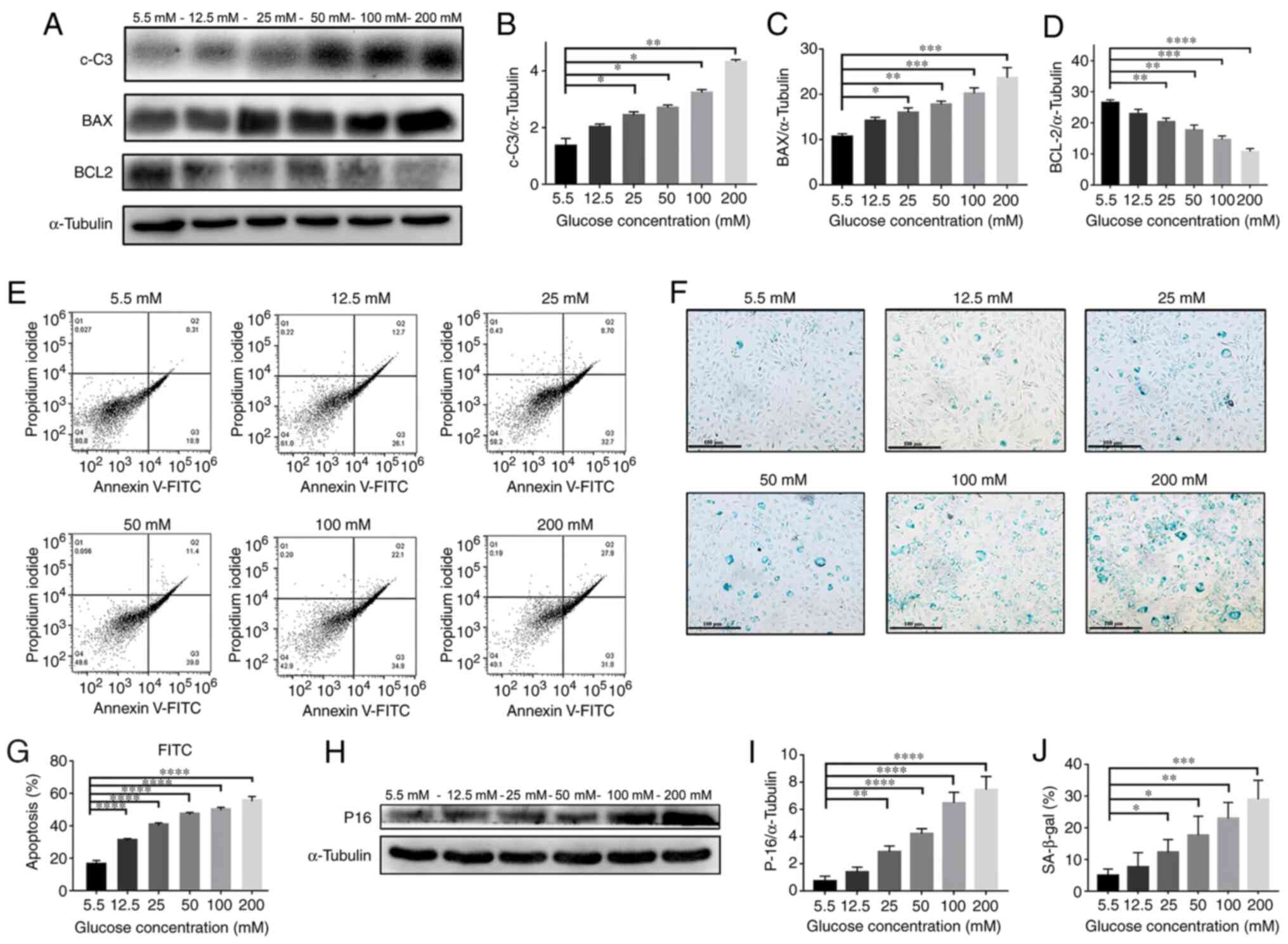 | Figure 3HG induces NPC apoptosis and
senescence. (A) Western blotting results of protein content and
quantification of (B) c-C3, (C) BAX and (D) BCL-2 in NPCs treated
with different concentrations (5.5, 12.5, 25, 50, 100 and 200 mM)
of glucose for 72 h. (E) Flow cytometry plots and (G) quantified
results of NPCs treated with different concentrations of glucose
for 72 h. (H) p16 western blotting results and (I) quantification
of protein contents of NPCs treated with different concentrations
of glucose for 72 h. (F) SA-β-gal staining assay was performed and
(J) quantified on NPCs treated with different concentrations of
glucose for 72 h (magnification, x100; scale bar, 100 µm).
*P<0.05; **P<0.01;
***P<0.001; ****P<0.0001 vs. 5.5 mM.
NPCs, nucleus pulposus cells; HG, high-glucose (50 mM); LG,
low-glucose (5.5 mM); c-C3, cleaved caspase 3. |
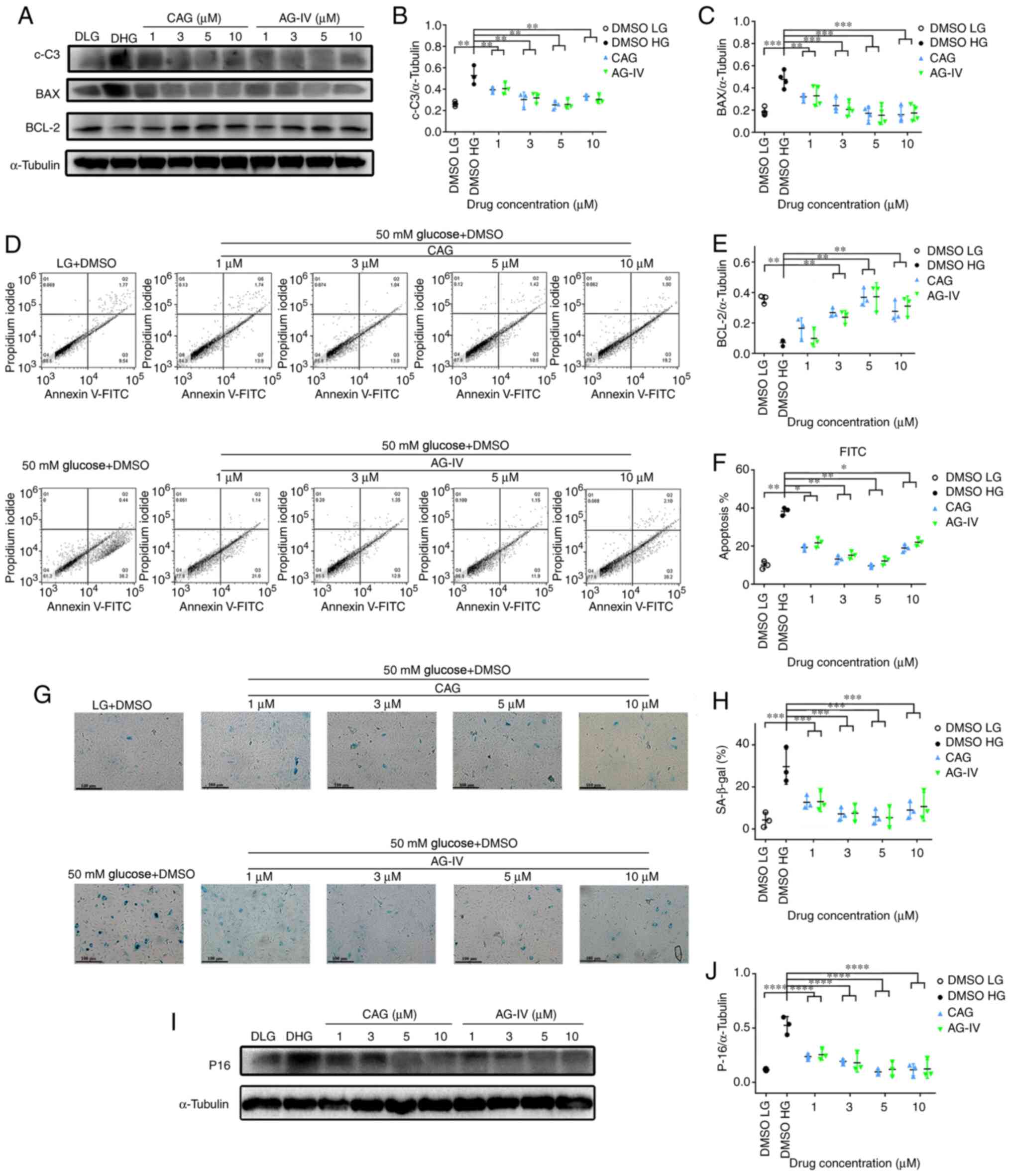
Moreover, as presented in Fig. 3E and G, the percentage of apoptosis as detected
by Annexin-V was significantly increased in a glucose
dose-dependent manner (P<0.0001). The results of the western
blotting in Fig. 3H and I also demonstrated that the expression
level of p16 protein was increased in a dose-dependent manner
(P<0.01). In addition, the β-Gal staining in Fig. 3F and J indicates a significantly higher
percentage of positively stained cells in a glucose
concentration-dependent manner (P<0.05) These results
demonstrated that high levels of glucose significantly promoted the
cellular apoptosis and senescence of NPCs.
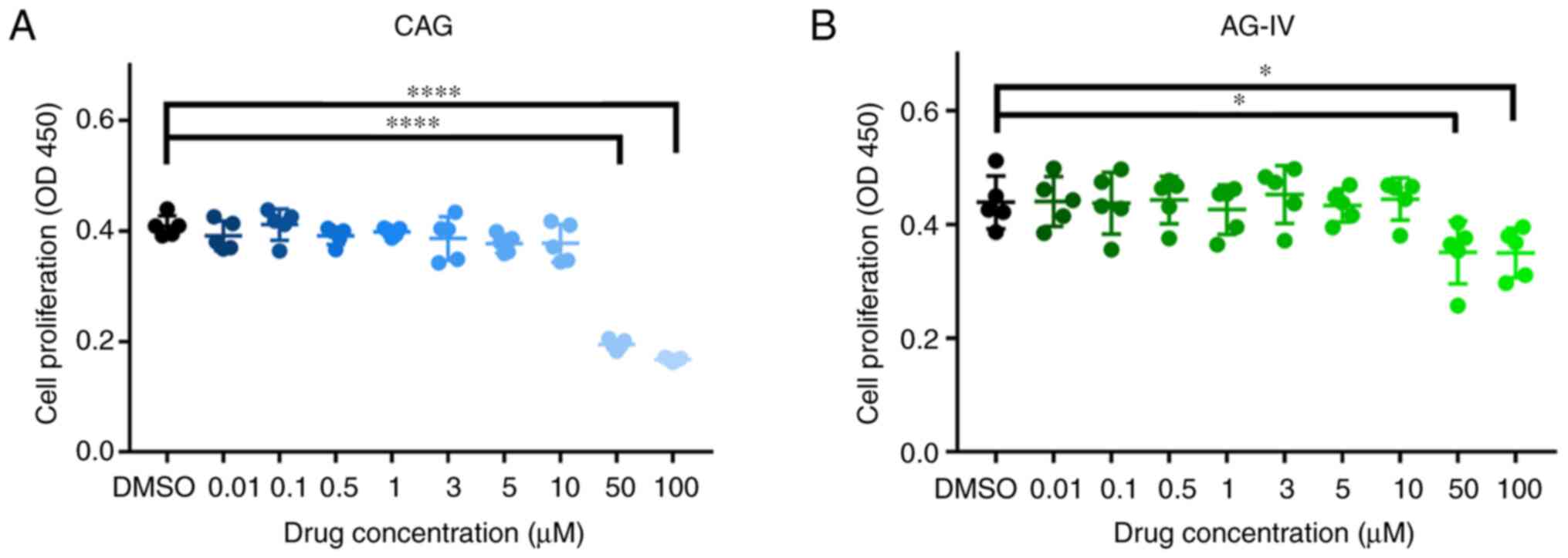
Cytotoxicity of CAG and AG-IV on
NPCs
NPCs were treated with different doses of CAG and
AG-IV and the cytotoxic effects of these drugs were determined at
various concentrations (0.01, 0.1, 0.5, 1, 3, 5, 10, 50 and 100 mM)
for 24 h using the CCK-8 assay. As presented in Fig. 4A and B, there were significant cytotoxic
effects at high doses (50 and 100 mM) of both drugs on the NPCs
(P<0.05). For subsequent experiments, the concentrations of 1,
3, 5 and 10 mM for 24 h were selected as the experimental
concentrations.
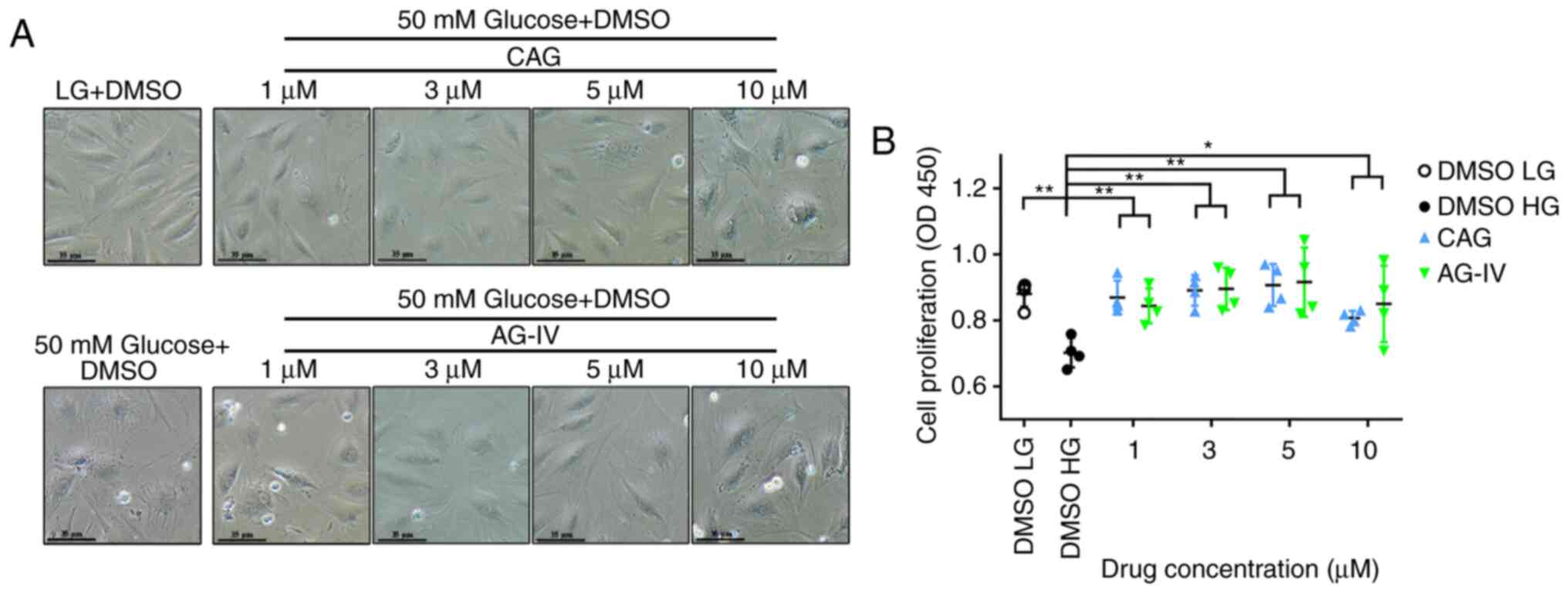
CAG and AG-IV improve cell morphology
and viability in HG conditions
NPCs seeded in 24-well plates were pretreated with
CAG or AG-IV at different concentrations (1, 3, 5 or 10 mM) for 24
h, and then serum starved for 24 h and treated with a HG
concentration of 50 mM for 72 h. Compared with the low glucose (LG,
5.5 mM) group, the NPCs differed in size and shape (bigger cells)
and the cell cytoplasm seemed coarse when treated with HG (50 mM).
However, treatment with CAG or AG-IV could prevent this phenomenon
at concentrations of 1, 3 and 5 mM (Fig. 5A).
The cellular viability was determined after 72 h of
HG treatment in pretreated NPCs using the CCK-8 assay. As presented
in Fig. 5B, there is a significant
difference between the untreated group (DMSO HG) and treated groups
(CAG and AG-IV). After treating with HG, the cell viability was
decreased compared with the DMSO LG group (P<0.001); however,
CAG and AG-IV demonstrated a significant protective effect against
HG (P<0.05; Fig. 5B). These
results demonstrated that CAG and AG-IV pretreatment resulted in
improved morphology of the HG-treated NPCs and significantly
increased their viability.
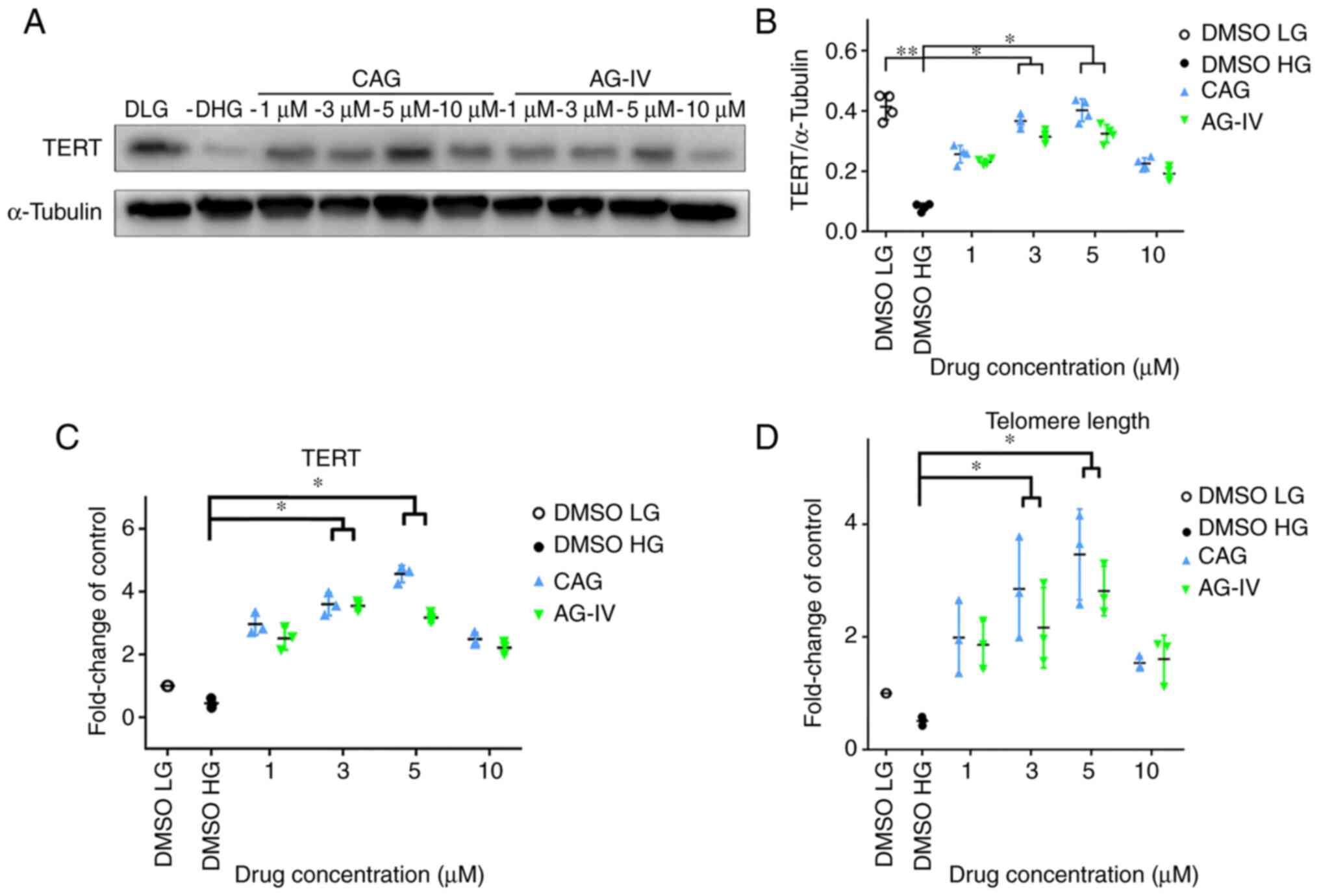
CAG and AG-IV increase TERT expression
and telomere length of NPCs in HG conditions
NPs were pretreated with CAG or AG-IV at 1, 3, 5 and
10 mM concentrations for 24 h, serum-starved for 24 h and then
treated with a HG concentration of 50 mM for 72 h. In Fig. 6A and B, western blotting demonstrated that TERT
protein expression levels were significantly decreased in HG-only
conditions compared with the LG group, but the expression recovered
in CAG and AG-IV drug treatment groups (P<0.05).
Meanwhile, as presented in Fig. 6C, the mRNA expression of TERT in
RT-qPCR was also markedly decreased in HG-only conditions compared
with the LG group, but the difference was not statistically
significant (P>0.05); however the expression levels
significantly increased in drug treatment groups at 3 and 5 mM
concentrations (P<0.05). Moreover, Fig. 6D indicates that the telomere length
detected using the qPCR method (25) was also markedly decreased in
HG-only conditions compared with the LG group, but this result was
not statistically significant. However, the telomere length
significantly increased in drug treatment groups at 3 and 5 mM
concentrations compared with the HG-only group (P<0.05).
Overall, compared with the HG group (DMSO HG), all other groups
exhibited higher TERT protein and mRNA expression levels, and
longer telomere length. These results demonstrated that TERT
expression and telomere length both increased in NPCs undergoing
HG-induced stress, following treatment with CAG and AG-IV.
CAG and AG-IV suppress apoptosis and
senescence in HG conditions
The NPCs were pretreated with CAG or AG-IV at 1, 3,
5 and 10 mM concentrations for 24 h, serum-starved for 24 h and
then treated with a HG concentration of 50 mM for 72 h. Fig. 7A presents the expression levels of
apoptosis-related proteins (c-C3, BAX and Bcl-2) as detected by
western blotting. Fig. 7B,
C and E demonstrates that expression levels of
c-C3 and BAX proteins were significantly increased, while Bcl-2 was
significantly decreased in HG-only conditions compared with the LG
group. However, while c-C3 and BAX protein levels were
significantly decreased in the drug treatment groups compared with
the HG group, while Bcl-2 was significantly increased
(P<0.01).
Meanwhile, as presented in Fig. 7D and F, the percentage of FITC (apoptosis
marker) as detected by Annexin-V was significantly increased in HG
conditions compared with the LG group, while the percentage
decreased in drug-treated groups compared with the HG group
(P<0.05). Moreover, the β-Gal staining in Fig. 7G and H shows a significantly higher percentage
of stain-positive cells in the HG group compared with the LG group;
whereas, the percentage of stain-positive cells significantly
decreased in the drug treated groups (P<0.0001). Finally, as
presented in Fig. 7I and J, the expression levels of p-16 protein
were significantly increased in the HG only group compared with the
LG condition group; whereas, the expression levels decreased in the
drug-treated groups compared with the HG group (P<0.0001). These
result demonstrated that CAG and AG-IV attenuated the cellular
apoptosis and senescence of NPCs treated with HG, and this was most
notable at concentrations of 3 or 5 mM.
Discussion
Senescence and apoptosis are considered to be the
main contributors to the pathological characteristics of IVDD and
Db-IVDD (5,6,26,27).
In senescence the cell cycle is arrested, viability is reduced and
levels of catabolic cytokines and extracellular matrix-degrading
enzymes are increased (26,28).
One of the main causes of senescence is telomere attrition, where
the telomere length is shortened to an extent that no more cell
division is possible. Though this can be avoided by the activation
of telomerase (which can elongate telomeres). TERT the main subunit
of the telomerase complex, has been indicated to significantly
suppress p16 expression (a senescence-associated protein) (14). Lower expression levels of TERT have
been implicated in a number of degenerative diseases, such as
amyotrophic lateral sclerosis and age-related macular degeneration
(13,22). CAG and AG-IV, the bioactive
constituents of the traditional Chinese herb Astragalus
membranaceus, have been identified as small-molecule TERT
activators (16,29). Previous studies have demonstrated
that CAG and AG-IV were able to alleviate and reverse degenerative
diseases by activating telomerase (22,29,30).
The present study examined the hypothesis that CAG
and AG-IV may postpone the progression of HG-IVDD by upregulating
TERT, without the disadvantages of activating TERT by viral
transfection. The results demonstrated that CAG and AG-IV protected
against HG-induced cellular senescence and apoptosis via activation
of telomerase, while at the same time increasing the telomere
length for prolonged survival and replications. Previous studies
have indicated that CAG and/or AGIV can protect or delay senescence
and apoptosis (30-32)
in different conditions, and that they are telomerase activators
(29,30); however, to the best of our
knowledge, this was the first study to examine their effect on NPCs
in HG conditions or examined their association with TERT,
senescence and apoptosis in HG conditions.
In diabetes, the main pathological factor is the
high levels of serum glucose, which leads to complications
associated with diabetes. The present study selected HG
concentrations (12.5, 25, 50, 100 and 200 mM) as the stimulant, as
previously performed by other studies to mimic the pathophysiology
of Db-IVDD in vitro (7,33,34).
The current study tested the effect of different concentrations of
glucose on cellular viability, morphology, senescence, apoptotic
markers and telomerase expression, as well as the telomere length.
Subsequently, 50 mM was selected to represent HG conditions, as
this had a significant effect under various treatment times.
A previous study demonstrated that HG concentrations
can adversely affect cellular viability (35). The present study used the CCK-8
assay to evaluate cellular viability and revealed that HG
concentrations adversely affected the cellular viability of NPCs in
a dose-dependent manner. Whereas treatment with CAG or AG-IV
improved the cellular viability, especially when treated with the
concentrations of 3 or 5 mM. It was hypothesized that this may be
associated with the activation of telomerase by CAG and AG-IV,
which in turn protects from replicative and stress-related
senescence, in addition to apoptosis of the NPCs.
It has been reported that HG concentrations and
diabetes can cause cellular senescence in NPCs (5,7). A
previous study demonstrated that TERT overexpression in human NPCs
can protect against cell cycle arrest by extending their
replicative capacity (36). It was
hypothesized that the ability of CAG (30) and AG-IV (29) to activate TERT attenuates
senescence. The present study used p16 protein levels and SA-β-gal
staining markers to measure cellular senescence, and it was
revealed that cellular senescence increased following treatment
with glucose in a dose-dependent manner in NPCs. However, by
pretreating the cells with CAG or AG-IV the cellular senescence was
attenuated in NPCs treated with HG, especially when at the
concentrations of 3 or 5 mM.
As reported in previous studies, diabetes and HG
concentrations can cause apoptosis (5,35).
CAG and AG-IV have been proven to have anti-apoptotic and
pro-survival properties both in vitro and in vivo in
different disease models, such as Parkinson's disease and cerebral
ischemia-reperfusion injury (30-32).
Also, as aforementioned, they are potent activators of TERT, which
is reported to have a protective effect against apoptosis (36). The current study used c-C3, BAX and
Bcl-2 proteins expression levels and FITC-Annexin-V as markers for
apoptosis. It was revealed that apoptosis was increased by glucose
treatment in a dose-dependent manner in NPCs. However, by
pretreating the cells with CAG or AG-IV, apoptosis was decreased in
NPCs with HG conditions.
Aging degenerative cells exhibit increased telomere
attrition, which is also a feature of other degenerative diseases
including osteoarthritis, neurodegeneration and diabetes (9,37,38).
Evidence has suggested that telomerase can protect cells from
telomere attrition and, thus, protects from senescence, apoptosis
and inflammation (14,39,40).
Previous studies have demonstrated that CAG and AGIV can activate
TERT, the main subunit of the telomerase complex (29,30),
which can elongate telomere ends, overcoming telomere attrition
(12,36,39).
Thus, the present study studied the anti-apoptotic and
anti-senescent effects of CAG and AG-IV with regards to telomerase
activation and telomere lengthening. The current study revealed
that at HG concentrations, TERT expression was significantly
decreased at both the mRNA and protein level, and that the telomere
length was shortened. This suggested that HG reduced TERT protein
expression and induced telomere attrition, playing a role in IVDD.
However, following CAG and AG-IV pretreatment, TERT expression and
telomere length both increased in NPCs undergoing HG-induced
stress. This suggested that CAG and AG-IV may serve a protective
role against HG-induced IVDD via an increase in TERT expression and
telomere lengthening.
It has been well characterized that morphological
changes occur in the diabetic disc both at gross and cellular
levels, and these changes are more prominent in the NP region
(41). Disc degeneration results
in a reduction in water content and increased breakdown of collagen
type II in NP (42). In addition,
the increased expression of senescence markers, such as p16 in the
NP aggravate IVDD (5,37), which is also associated with the
altered phenotype of having morphologically larger cell types.
Furthermore, previous studies have shown that HG can adversely
affect cellular proliferation and increase the expression of matrix
metalloproteinases, also resulting in IVDD (6,43).
The present study demonstrated that CAG and AG-IV pretreatment
resulted in improved morphology of the HG-treated NPCs and
significantly increased their viability. By contrast, the cells
treated with HG only demonstrated larger cell types with a coarse
and spindled appearance.
These findings may be beneficial for future research
and/or clinical trials; furthermore, they may also be used to
formulate a new type of culture media. HG culture media may
accelerate senescence in certain types of cells, while these drugs
could be helpful in delaying that. In addition, these drugs may
potentially help diabetic patients prevent the development of
Db-IVDD.
There are certain limitations to the present study.
Firstly, the in vitro HG-induced Db-IVDD model does not
perfectly reflect in vivo aspects of IVDD with diabetes, the
reason for that was the limited budget of the present study. Thus,
further in vivo studies are needed to know the effect of
these drugs. Secondly, the present study did not focus on an
in-depth study of the TERT pathway or mechanisms associated with
senescence and apoptosis; therefore, further studies are needed to
gain more in-depth knowledge of the signaling pathways involved.
Thirdly, there was no investigation carried out on the effect of
these drugs on oxidative stress, such as reactive oxygen species
(ROS), due to the HG conditions. Further studies are needed to
address this.
It is known that p53 can suppress transcription of
TERT, and that this may play a role in p53-mediated apoptosis. The
activation of NF-κB induces TERT expression, which suggests the
possibility that TERT is upregulated in response to survival
signals that activate NF-κB; including growth factors, cytokines
and stress responses. Moreover, TERT expression is induced by the
transcription factor c-Myc, a protein strongly associated with cell
immortalization and cancer. All this suggests that telomerase acts
to suppress an early step in the cell death pathway prior to the
mitochondrial step at which Bcl-2 acts. Therefore, we hypothesize
that CAG and AG-IV may inhibit p53, BAX, PUMA, Bim and ROS and, in
turn, promote controlled expression of pro-survival protein
pathways such as TERT, c-Myc, NF-κB and Bcl-2.
In summary, the present study revealed that CAG and
AG-IV serve a notable role in the prevention of HG-mediated IVDD by
attenuating senescence, including replicative senescence and
stress-induced premature senescence. These drugs may also inhibit
apoptosis in NPCs by upregulating telomerase activation and
lengthening the telomere. TERT may represent a potential target for
future therapeutic interventions, while CAG and AG-IV could be
potential therapeutic agents in attenuating HG-IVDD.
Acknowledgements
Not applicable.
Funding
Funding: The present study is supported by grants from National
Regional Natural Fund of China (grant no. 81660366), Zhejiang
Provincial Natural Science Foundation of China (grant no.
LY18H060012 and LY17H060010), Zhejiang Provincial Key Laboratory of
Orthopedics (grant no. ZJGK1802Z), Zhejiang Public Service
Technology Research Program/Social Development (grant no.
LGF18H060008), Major Scientific And Technological Project of
Medical and Health in Zhejiang Province (grant no. WKJ-ZJ-1527),
Zhejiang Provincial Project for Medical and Health Science and
Technology (grant no. 2017KY463) and Wenzhou Science and Technology
Bureau Foundation (grant no. Y20170092).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HH and JX performed the research, analyzed the data
and prepared the manuscript. QG assisted in western blotting and
β-gal staining. JD assisted in cell culture. ZJ collected nucleus
pulposus from rats. SL provided technical assistance in flow
cytometry. HZ performed statistical analysis. XW and XZ designed
the research and supervised the process of the study. HH and JX
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Wenzhou Medical University (approval no.
wydw2014-0129; Wenzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest.
References
|
1
|
Deyo RA and Mirza SK: CLINICAL PRACTICE.
Herniated lumbar intervertebral disk. N Engl J Med. 374:1763–1772.
2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Modic MT and Ross JS: Lumbar degenerative
disk disease. Radiology. 245:43–61. 2007.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kadow T, Sowa G, Vo N and Kang JD:
Molecular basis of intervertebral disc degeneration and
herniations: What are the important translational questions. Clin
Orthop Relat Res. 473:1903–1912. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Jiang L, Zhang X, Zheng X, Ru A, Ni X, Wu
Y, Tian N, Huang Y, Xue E, Wang X and Xu H: Apoptosis, senescence,
and autophagy in rat nucleus pulposus cells: Implications for
diabetic intervertebral disc degeneration. J Orthop Res.
31:692–702. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Wang L, Gao P, Zhang M, Huang Z, Zhang D,
Deng Q, Li Y, Zhao Z, Qin X, Jin D, et al: Prevalence and ethnic
pattern of diabetes and prediabetes in China in 2013. JAMA.
317:2515–2523. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Won HY, Park JB, Park EY and Riew KD:
Effect of hyperglycemia on apoptosis of notochordal cells and
intervertebral disc degeneration in diabetic rats. J Neurosurg
Spine. 11:741–748. 2009.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kong JG, Park JB, Lee D and Park EY:
Effect of high glucose on stress-induced senescence of nucleus
pulposus cells of adult rats. Asian Spine J. 9:155–161.
2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhang XU, Yang MK, Li Z, Liu C, Wu JS and
Wang J: Expression and significance of telomerase in the nucleus
pulposus tissues of degenerative lumbar discs. Biomed Rep.
3:813–817. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Monickaraj F, Aravind S, Gokulakrishnan K,
Sathishkumar C, Prabu P, Prabu D, Mohan V and Balasubramanyam M:
Accelerated aging as evidenced by increased telomere shortening and
mitochondrial DNA depletion in patients with type 2 diabetes. Mol
Cell Biochem. 365:343–350. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Xiao F, Zheng X, Cui M, Shi G, Chen X, Li
R, Song Z, Rudolph KL, Chen B and Ju Z: Telomere
dysfunction-related serological markers are associated with type 2
diabetes. Diabetes Care. 34:2273–2278. 2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Skvortsov DA, Zvereva ME, Shpanchenko OV
and Dontsova OA: Assays for detection of telomerase activity. Acta
Naturae. 3:48–68. 2011.PubMed/NCBI
|
|
12
|
Chung SA, Wei AQ, Connor DE, Webb GC,
Molloy T, Pajic M and Diwan AD: Nucleus pulposus cellular longevity
by telomerase gene therapy. Spine (Phila Pa 1976). 32:1188–1196.
2007.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Eitan E, Tichon A, Gazit A, Gitler D,
Slavin S and Priel E: Novel telomerase-increasing compound in mouse
brain delays the onset of amyotrophic lateral sclerosis. EMBO Mol
Med. 4:313–329. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Graham MK, Principessa L, Antony L, Meeker
AK and Isaacs JT: Low p16(INK4a) expression in early passage human
prostate basal epithelial cells enables immortalization by
telomerase expression alone. Prostate. 77:374–384. 2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yu Y, Zhou L, Yang Y and Liu Y:
Cycloastragenol: An exciting novel candidate for age-associated
diseases. Exp Ther Med. 16:2175–2182. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Szabo NJ: Dietary safety of
cycloastragenol from Astragalus spp.: Subchronic toxicity
and genotoxicity studies. Food Chem Toxicol. 64:322–334.
2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Xiao WL, Motley TJ, Unachukwu UJ, Lau CB,
Jiang B, Hong F, Leung PC, Wang QF, Livingston PO, Cassileth BR and
Kennelly EJ: Chemical and genetic assessment of variability in
commercial Radix Astragali (Astragalus spp.) by ion trap
LC-MS and nuclear ribosomal DNA barcoding sequence analyses. J
Agric Food Chem. 59:1548–1556. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhang N, Wang XH, Mao SL and Zhao F:
Astragaloside IV improves metabolic syndrome and endothelium
dysfunction in fructose-fed rats. Molecules. 16:3896–3907.
2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang B and Chen MZ: Astragaloside IV
possesses antiarthritic effect by preventing interleukin
1beta-induced joint inflammation and cartilage damage. Arch Pharm
Res. 37:793–802. 2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang L, Gu W, Shi Y, Chen Y and Tan Y:
Protective effects of astragaloside IV on IL-8-treated
diaphragmatic muscle cells. Exp Ther Med. 17:519–524.
2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang J and Guo HM: Astragaloside IV
ameliorates high glucose-induced HK-2 cell apoptosis and oxidative
stress by regulating the Nrf2/ARE signaling pathway. Exp Ther Med.
17:4409–4416. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Dow CT and Harley CB: Evaluation of an
oral telomerase activator for early age-related macular
degeneration-a pilot study. Clin Ophthalmol. 10:243–249.
2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zheng G, Pan Z, Zhan Y, Tang Q, Zheng F,
Zhou Y, Wu Y, Zhou Y, Chen D, Chen J, et al: TFEB protects nucleus
pulposus cells against apoptosis and senescence via restoring
autophagic flux. Osteoarthritis Cartilage. 27:347–357.
2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cawthon RM: Telomere measurement by
quantitative PCR. Nucleic Acids Res. 30(e47)2002.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chen J, Xie JJ, Jin MY, Gu YT, Wu CC, Guo
WJ, Yan YZ, Zhang ZJ, Wang JL, Zhang XL, et al: Sirt6
overexpression suppresses senescence and apoptosis of nucleus
pulposus cells by inducing autophagy in a model of intervertebral
disc degeneration. Cell Death Dis. 9(56)2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Kletsas D: Senescent cells in the
intervertebral disc: Numbers and mechanisms. Spine J. 9:677–678.
2009.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Davalos AR, Coppe JP, Campisi J and
Desprez PY: Senescent cells as a source of inflammatory factors for
tumor progression. Cancer Metastasis Rev. 29:273–283.
2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yung LY, Lam WS, Ho MK, Hu Y, Ip FC, Pang
H, Chin AC, Harley CB, Ip NY and Wong YH: Astragaloside IV and
cycloastragenol stimulate the phosphorylation of extracellular
signal-regulated protein kinase in multiple cell types. Planta Med.
78:115–121. 2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ip FC, Ng YP, An HJ, Dai Y, Pang HH, Hu
YQ, Chin AC, Harley CB, Wong YH and Ip NY: Cycloastragenol is a
potent telomerase activator in neuronal cells: Implications for
depression management. Neurosignals. 22:52–63. 2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Huang XP, Tan H, Chen BY and Deng CQ:
Combination of total Astragalus extract and total Panax
notoginseng saponins strengthened the protective effects on
brain damage through improving energy metabolism and inhibiting
apoptosis after cerebral ischemia-reperfusion in mice. Chin J
Integr Med. 23:445–452. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhang ZG, Wu L, Wang JL, Yang JD, Zhang J,
Zhang J, Li LH, Xia Y, Yao LB, Qin HZ and Gao GD: Astragaloside IV
prevents MPP(+)-induced SH-SY5Y cell death via the inhibition of
Bax-mediated pathways and ROS production. Mol Cell Biochem.
364:209–216. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Jiang C, Liu S, Cao Y and Shan H: High
glucose induces autophagy through PPARγ-dependent pathway in human
nucleus pulposus cells. PPAR Res. 2018(8512745)2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Cheng X, Ni B, Zhang F, Hu Y and Zhao J:
High glucose-induced oxidative stress mediates apoptosis and
extracellular matrix metabolic imbalances possibly via p38 MAPK
activation in rat nucleus pulposus cells. J Diabetes Res.
2016(3765173)2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhou KL, Zhou YF, Wu K, Tian NF, Wu YS,
Wang YL, Chen DH, Zhou B, Wang XY, Xu HZ and Zhang XL: Stimulation
of autophagy promotes functional recovery in diabetic rats with
spinal cord injury. Sci Rep. 5(17130)2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Liang W, Ye D, Dai L, Shen Y and Xu J:
Overexpression of hTERT extends replicative capacity of human
nucleus pulposus cells, and protects against serum
starvation-induced apoptosis and cell cycle arrest. J Cell Biochem.
113:2112–2121. 2012.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Le Maitre CL, Freemont AJ and Hoyland JA:
Accelerated cellular senescence in degenerate intervertebral discs:
A possible role in the pathogenesis of intervertebral disc
degeneration. Arthritis Res Ther. 9(R45)2007.PubMed/NCBI View
Article : Google Scholar
|
|
38
|
Baltzis D, Meimeti E, Grammatikopoulou MG,
Roustit M, Mavrogonatou E, Kletsas D, Efraimidou S, Manes C,
Nikolouzakis TK, Tsiaoussis J, et al: Assessment of telomerase
activity in leukocytes of type 2 diabetes mellitus patients having
or not foot ulcer: Possible correlation with other clinical
parameters. Exp Ther Med. 15:3420–3424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wu J, Wang D, Ruan D, He Q, Zhang Y, Wang
C, Xin H, Xu C and Liu Y: Prolonged expansion of human nucleus
pulposus cells expressing human telomerase reverse transcriptase
mediated by lentiviral vector. J Orthop Res. 32:159–166.
2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wu J, Wang D, Zhang C, Wang C, Zhang Y,
Xin H, He Q and Ruan D: Extending the activities of human nucleus
pulposus cells with recombinant adeno-associated virus
vector-mediated human telomerase reverse transcriptase gene
transfer. Tissue Eng Part A. 17:2407–2415. 2011.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Illien-Junger S, Grosjean F, Laudier DM,
Vlassara H, Striker GE and Iatridis JC: Combined anti-inflammatory
and anti-AGE drug treatments have a protective effect on
intervertebral discs in mice with diabetes. PLoS One.
8(e64302)2013.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Antoniou J, Steffen T, Nelson F,
Winterbottom N, Hollander AP, Poole RA, Aebi M and Alini M: The
human lumbar intervertebral disc: Evidence for changes in the
biosynthesis and denaturation of the extracellular matrix with
growth, maturation, ageing, and degeneration. J Clin Invest.
98:996–1003. 1996.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Park EY and Park JB: Dose- and
time-dependent effect of high glucose concentration on viability of
notochordal cells and expression of matrix degrading and fibrotic
enzymes. Int Orthop. 37:1179–1186. 2013.PubMed/NCBI View Article : Google Scholar
|