Introduction
Breast cancer (BC) is one of the most common types
of cancer that affects women worldwide (1). In China, ~304,000 cases of female BC
and 70,000 deaths were recorded in 2015(2). A number of factors are involved in
the development and promotion of breast malignancy, where the most
of which involve changes in the expression profile of microRNAs
(miRNAs/miRs) (3,4).
miRNAs are non-coding RNAs that are 21-24
nucleotides in length and post-transcriptionally regulate gene
expression by binding to the 3' untranslated regions (UTR) of their
mRNA targets (5). Previous studies
have demonstrated that miRNAs can regulate an array of signaling
pathways, thereby negatively (6)
or positively (7) affecting
tumorigenesis and various aspects of cancer progression.
Particularly in BC, Kim et al (8) revealed that the stable inhibition of
miR-155 can reduce tumor growth by decreasing glucose metabolism
through the phosphoinositide-3-kinase regulatory subunit 1-pyruvate
dehydrogenase complex kinase/AKT-forkhead box O3a-cMYC axis. Shao
et al (9) demonstrated that
the plasma level of miR-200a was associated with BC staging at the
time of surgery, where high levels of miR-210 expression was
associated with internal organ metastasis (liver, lung and brain).
Therefore, to promote the application of miRNAs in BC diagnosis,
treatment and prognosis prediction, deeper understanding into the
targets and underlying regulatory mechanisms of miRNAs is of great
importance.
miR-515-5p was first detected to be a
placenta-specific miRNA present in the maternal circulation
(10). Previously, miR-515-5p was
found to be differentially expressed in prostate cancer tissues,
hepatocellular adenomas and normal tissues (11,12).
In addition, knockdown miR-515-5p was reported to promote non-small
cell lung cancer cell survival and metastasis and miR-515-5p were
downregulated in the C-X-C motif chemokine ligand 6(13). These previous observations suggest
that miR-515-5p can be used as a biomarker for the prognosis of
patients with cancer and possibly serve as a therapeutic target for
cancer.
However, in BC, the role of miR-515-5p is unclear;
the present study therefore investigated the function of miR-515-5p
in BC.
Materials and methods
Cell lines
The human BC cell lines (MCF7, ZR-75-30, MDA-MB-231
and SKBr-3) and the normal breast cell line (MCF10A) were obtained
from the American Type Culture Collection. MCF7 cells were cultured
in modified Eagle's medium (cat. no. 30030; Thermo Fisher
Scientific, Inc.) supplemented with 1.5 g/l NaHCO3
(Merck KGaA), 10% FBS (Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 mg/ml streptomycin at 37˚C with 5%
CO2. MDA-MB-231 and MCF10A cells were cultured in DMEM
(cat. no. 11965092; Thermo Fisher Scientific, Inc.) supplemented
with 10% FBS, 100 U/ml penicillin and 100 mg/ml streptomycin at
37˚C with 5% CO2. ZR-75-30 and SKBr-3 cells were
maintained in RPMI-1640 medium (cat. no. 31870082, Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS, 100 U/ml penicillin
and 100 mg/ml streptomycin at 37˚C with 5% CO2.
Cell transfection
miR-515-5p mimic (UUCUCCAAAAGAAAGCACUUUCUG), NC
mimic (UACUGAGAGACAUAAGUUGGUC), pcDNA3.1-chromobox 4 (CBX4) and
their respective negative controls (NC mimic and Lv-NC) were
designed and obtained from Shanghai GenePharma Co., Ltd. Cell
transfection was performed on MCF7 or ZR-75-30 cells using
X-Porator H1 (cat. no. EBXP-H1, Etta Biotech Co., Ltd.), according
to the manufacturer's protocol. Briefly, the MCF7 and ZR-75-30
cells were collected and resuspended in the electroporation buffer
at 270 mOsm osmolarity, 0.1 S/m conductivity (Etta Biotech Co.,
Ltd.). The cell concentration was then adjusted to 6x105
cells/ml along with mixed 150 nM miRNA mimics and/or 1 mg/ml
plasmid. A total of 100 µl cell suspension mixed with RNAs was then
added into a 0.4 mm cuvette and used Matrix needle electrodes for
gene transfection using the operator H1. The electroporation device
was operated at a direct current square wave with a 180-V voltage,
4 nA, 500-µsec duration, 1-sec intervals and three pulses at room
temperature Following electroporation, the cells were diluted to
appropriate concentrations in MEM supplemented with 1.5 g/l
NaHCO3, 10% FBS and seeded into appropriate cell culture
plates at 37˚C with 5% CO2, for use in further assays at
24 h post-electroporation.
miRNAs and mRNA expression
Total RNA from the cells was isolated using the
TRIzol® reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocols. MicroRNA levels were
measured using TaqMan™ MicroRNA Reverse Transcription
kit (cat. no. 4366596; Thermo Fisher Scientific, Inc.). The
conditions for RT were 16˚C for 30 min, 42˚C for 30 min and 85˚C
for 5 min to terminate the reaction. TaqMan 2X Fast Universal PCR
Master Mix (cat. no. 4367846; Thermo Fisher Scientific, Inc.). The
thermocycling conditions were 95˚C for 20 sec, followed by 40
cycles of 95˚C for 1 sec and 60˚C for 20 sec. The sequence-specific
forward primers for mature miR-515-5p and the U6 internal control
were 5'-TTCTCCAAAAGAAAGCACTTTCTG-3' and 5'-CTCGCTTCGGCAGCACA-3',
respectively. The reverse primer was the universal reverse primer:
5'-GTGCAGGGTCCGAGGT-3'.
To analyze mRNA expression, both cDNA conversion and
qPCR amplification were performed using High-Capacity cDNA Reverse
Transcription kit (cat. no. 4368813) at 25˚C for 10 min, 37˚C for
120 min, and heated at 85˚C for 5 min and Fast
SYBR®-Green Master mix (cat. no. 4385614; both from
Thermo Fisher Scientific, Inc.). The thermocycling conditions were
95˚C for 20 sec, followed by 40 cycles of 95˚C for 1 sec and 60˚C
for 20 sec, respectively, according to the manufacturer's
protocols. Primers for CBX4 were forward,
5'-TGGAGTATCTGGTGAAATGGA-3' and reverse,
5'-ACGACGGGCAAAGGTAGGCAC-3'. Primer for β-actin were forward,
5'-GTGGGGCGCCCCAGGCACCAGGGC-3' and reverse,
5'-CTCCTTAATGTCACGCACGATTTC-3'.
Cell viability assay
Cell Counting Kit-8 (CCK-8; cat. no. C0037, Beyotime
Institute of Biotechnology) colorimetric assay was used to measure
cell viability. Briefly, following transfection, MCF7 or ZR-75-30
cells were seeded into 96-well plates with 1x104
cells/well. After 0, 24, 48 and 72 h cultured at 37˚C with 5%
CO2, the supernatant was removed before 100 µl MEM or
DMEM containing 10 µl CCK-8 was added into each well for 3 h of
incubation at 37˚C. A plate reader (Thermo Multiskan MK3
spectrophotometer; Thermo Fisher Scientific, Inc.) was used to
measure absorbance at 450 nm. The optical density value was
determined and used to construct a growth curve to assess cell
viability.
Colony formation assay
MCF7 or ZR-75-30 cells were seeded into 10-cm dishes
(1x103 cells/plate) following transfection. The cells
were then maintained in complete culture medium (MEM or RPMI-1640,
respectively) for another 21 days at 37˚C with 5% CO2.
Finally, the cells were fixed in 4% paraformaldehyde at 4˚C for 30
min and stained with Giemsa dye at 25˚C for 30 min. Images of cells
were captured by a camera, and the number of clones visible by eye
was calculated.
TUNEL assay
After 24 h of transfection, MCF7 or ZR-75-30 cells
were fixed with 4% paraformaldehyde and apoptosis was detected
using a TUNEL assay, as recommended in the ApopTag® Plus
Peroxidase In Situ Apoptosis kit (cat. no. S7101; Merck
KGaA). Briefly, the cells were fixed with 4% paraformaldehyde for
30 min at room temperature and permeabilized with 0.1% Triton X-100
in 0.1% sodium citrate for 2 min at 4˚C. The cells were then
incubated with 1% TdT enzyme in a humidified atmosphere at 37˚C for
90 min. Subsequently, they were stained with DAPI at a final
concentration of 4 µg/ml at 4˚C for 10 min. Finally, the cells were
observed under a fluorescence microscope (200x, CKX53; Olympus). An
average of 10 random fields with 100-200 nuclei per field was
analyzed.
Flow cytometry
The effect of miR-515-5p on the cell cycle was
measured by flow cytometry, as previously described (14). Briefly, MCF7 or ZR-75-30 cells were
harvested and fixed with 4% paraformaldehyde for 30 min at 4˚C. The
cells were then incubated with RNase (50 µg/ml) and PI (50 µg/ml;
Thermo Fisher Scientific, Inc.) for 30 min at room temperature.
Next, cell cycle profile was measured using a flow cytometry system
(BD Accuri C6; BD Biosciences) and the relative ratios of the
G0/G1, S and G2/M phases were
analyzed by FlowJo V10 software (FlowJo LLC).
Wound healing assay
Wound healing assay was performed to study the
migration of MCF7 and ZR-75-30 cells as previously described
(15). Briefly, 5x105
MCF7 or ZR-75-30 cells were seeded into each well of a six-well
plate. Following transfection for 24 h, a wound was made using a
100-µl pipette tip. Following culture in MEM supplemented with 1.5
g/l NaHCO3, 10% FBS, 100 U/ml penicillin and 100 mg/ml
streptomycin at 37˚C with 5% CO2 for another 48 h, the
wound recovery area was photographed under a light microscope (200x
magnification; CKX41, Olympus) at 0 and 48 h, and evaluated by
ImageJ software (version 1.8.0; NIH).
Transwell assay
MCF7 or ZR-75-30 cells were seeded into the upper
chamber with 5x105 cells containing MEM or DMEM. By
contrast, 10% FBS and complete culture medium were added to the
lower chamber. The cells were then cultured for ~48 h at 37˚C with
5% CO2. Next, the cells in the lower chamber were fixed
with 4% paraformaldehyde for 30 min at 4˚C and stained with 5%
crystal violet at 4˚C for 30 min. Finally, cells were photographed
under a light microscope (x100 magnification; cat. no. CKX41;
Olympus) and counted in five randomly selected microscope
fields.
Dual-luciferase reporter assay
The complementary sequences between miR-515-5p and
CBX4 were predicted using Starbase (starbase.sysu.edu.cn/), MCF7 or ZR-75-30 cells were
co-transfected with 0.24 µg of the CBX4-wild-type (WT) or
CBX4-mutant (Mut) 3'UTR and pmiRGLO Dual-Luciferase Vector were
designed and obtained from Shanghai GenePharma Co., Ltd. together
with 40 nM miR-515-5p mimic or NC mimic using cell electroporation
system operator H1, following the manufacturer's protocols. In
addition, 0.05 µg Renilla luciferase expression plasmid was
transfected into the cells as a reference control. Subsequently, 48
h after transfection, the cells were collected to measure Firefly
and Renilla luciferase activities using the Dual-luciferase
reporter assay system (cat. no. E1910; Promega Corporation). The
relative luciferase activity was calculated as the ratio of Firefly
and Renilla luciferase activities.
Western blot analysis
MCF7 or ZR-75-30 cells were harvested using RIPA
lysis buffer (P0013B; Beyotime Institute of Biotechnology) and
quantified by BCA assay. The total protein (40 µg) was then
separated on 12% SDS-PAGE and transferred onto PVDF membranes (EMD
Millipore) using a MiniGenie blotting system (Bio-Rad Laboratories,
Inc.). Next, the membranes were blocked with TBS-1% Tween-20 (TBST)
containing 1% skim milk powder for 1 h at room temperature and then
incubated with rabbit monoclonal antibodies (Cell Signaling
Technology, Inc.) against human prostaglandin-endoperoxide synthase
2 (Cox-2; cat. no. 12282T; dilution, 1:1,000), MMP2 (cat. no.
40994S; dilution, 1:1,000), MMP9 (cat. no. 13667S; dilution,
1:1,000), cyclin D1 (cat. no. 55506S; dilution, 1:1,000) cyclin
dependent kinase 2 (CDK2; cat. no. 2546S; dilution, 1:1,000),
proliferating cell nuclear antigen (PCNA; cat. no. 13110S;
dilution, 1:1,000), β-actin (cat. no. 4970T; dilution, 1:1,000) and
CBX4 (cat. no. 30559; dilution, 1:2,000) at 4˚C overnight.
Following washing with TBST, the membranes were incubated with goat
anti-rabbit secondary antibodies (cat. no. 5127; dilution,
1:10,000; Cell Signaling Technology, Inc.) for 1 h at room
temperature. Finally, the protein brands were visualized with an
enhanced chemiluminescence system (P0018FS; Beyotime Institute of
Biotechnology) and semi-quantified with ImageJ software (version
1.46; National Institutes of Health).
Statistical analysis
All experiments were performed in triplicate.
GraphPad Prism (version 6.01 for Windows; GraphPad Software, Inc.)
statistical software was used to perform the statistical analysis.
One-way-ANOVA (parametric) followed by Tukey's post hoc test was
used to compare the significant differences between groups.
P<0.01 was considered to indicate a statistically significant
difference.
Results
miR-515-5p inhibits cell proliferation
of MCF7 and ZR-75-30 cells
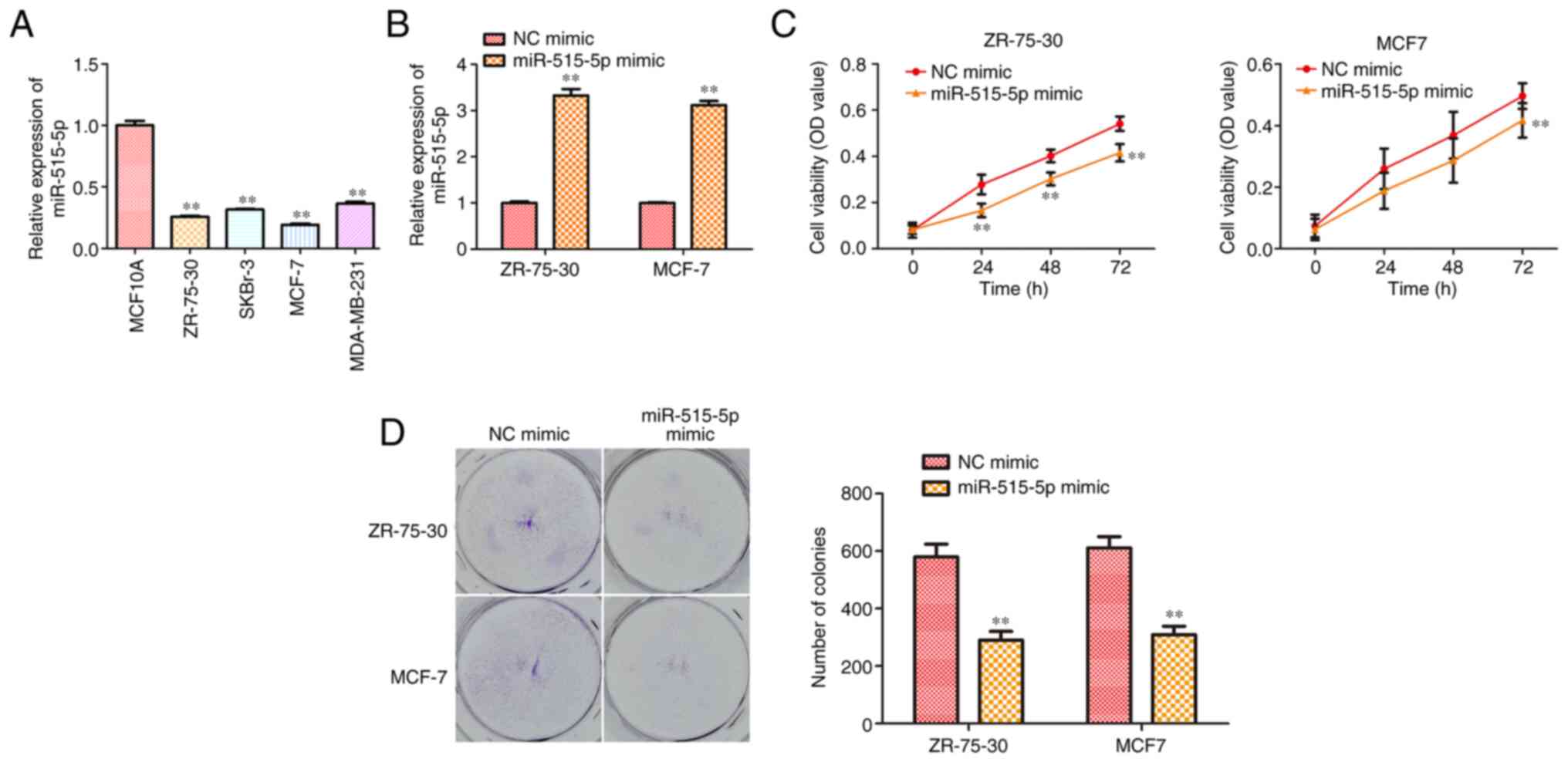
To confirm if miR-515-5p participates in BC
progression, its expression level in BC cell lines MCF7, ZR-75-30,
MDA-MB-231, SKBr-3 and the normal breast cell line MCF10A was
compared using RT-qPCR. The results showed that the expression of
miR-515-5p in human BC lines was significantly lower compared with
that in the normal breast cell line (P<0.01; Fig. 1A).
To evaluate the possible effects of miR-515-5p
expression on BC progression, MCF7 and ZR-75-30 cells were
transfected with miR-515-5p or NC mimics. RT-qPCR results showed
that transfection with the miR-515-5p mimic could significantly
upregulate miR-515-5p expression in MCF7 and ZR-75-30 cells
compared with that in cells transfected with the NC mimic
(P<0.01; Fig. 1B). In addition,
CCK-8 assay data showed that miR-515-5p mimic transfection could
significantly reduce the viability of MCF7 and ZR-75-30 cells
compared with that in cells transfected with the NC mimic
(P<0.01; Fig. 1C). Colony
formation assay results revealed that transfection with the
miR-515-5p mimic significantly reduced the proliferation of MCF7
and ZR-75-30 cells (P<0.01; Fig.
1D). In conclusion, these data suggest that miR-515-5p
overexpression could inhibit BC cell proliferation.
miR-515-5p facilitates cell apoptosis
and induces cell cycle arrest at the G0/G1
phase in MCF7 and ZR-75-30 cells
The regulation of apoptosis and cell cycle arrest
has been found to play an important role in cell proliferation
(13). To reveal the mechanism of
miR-515-5p in BC cell proliferation, TUNEL and flow cytometric
assays were performed to detect the potential effects of miR-515-5p
on cell apoptosis and cell cycle progression. First, results from
the TUNEL assay indicated that miR-515-5p mimic transfection could
significantly facilitate MCF7 and ZR-75-30 cell apoptosis compared
with that in cells transfected with the NC mimic (P<0.05 and
P<0.01; Fig. 2A). Next, data
from the flow cytometric assay suggest that the miR-515-5p mimic
significantly increased the percentage of cells at the
G0/G1 phase and decreased the ratio of cells
at G2/M phase in both MCF7 and ZR-75-30 cells
(P<0.01; Fig. 2B). Finally,
western blot analysis was performed to assess the expression of
cell cycle-related proteins. The results showed that, miR-515-5p
mimic transfection significantly reduced the expression levels of
cyclin D1, CDK2 and increased the expression level of p21 proteins
compared with those in cells transfected with the NC mimic
(P<0.05 and P<0.01; Fig.
2C). These results suggest that miR-515-5p can promote cell
apoptosis whilst inducing cell cycle arrest at the
G0/G1 phase in BC.
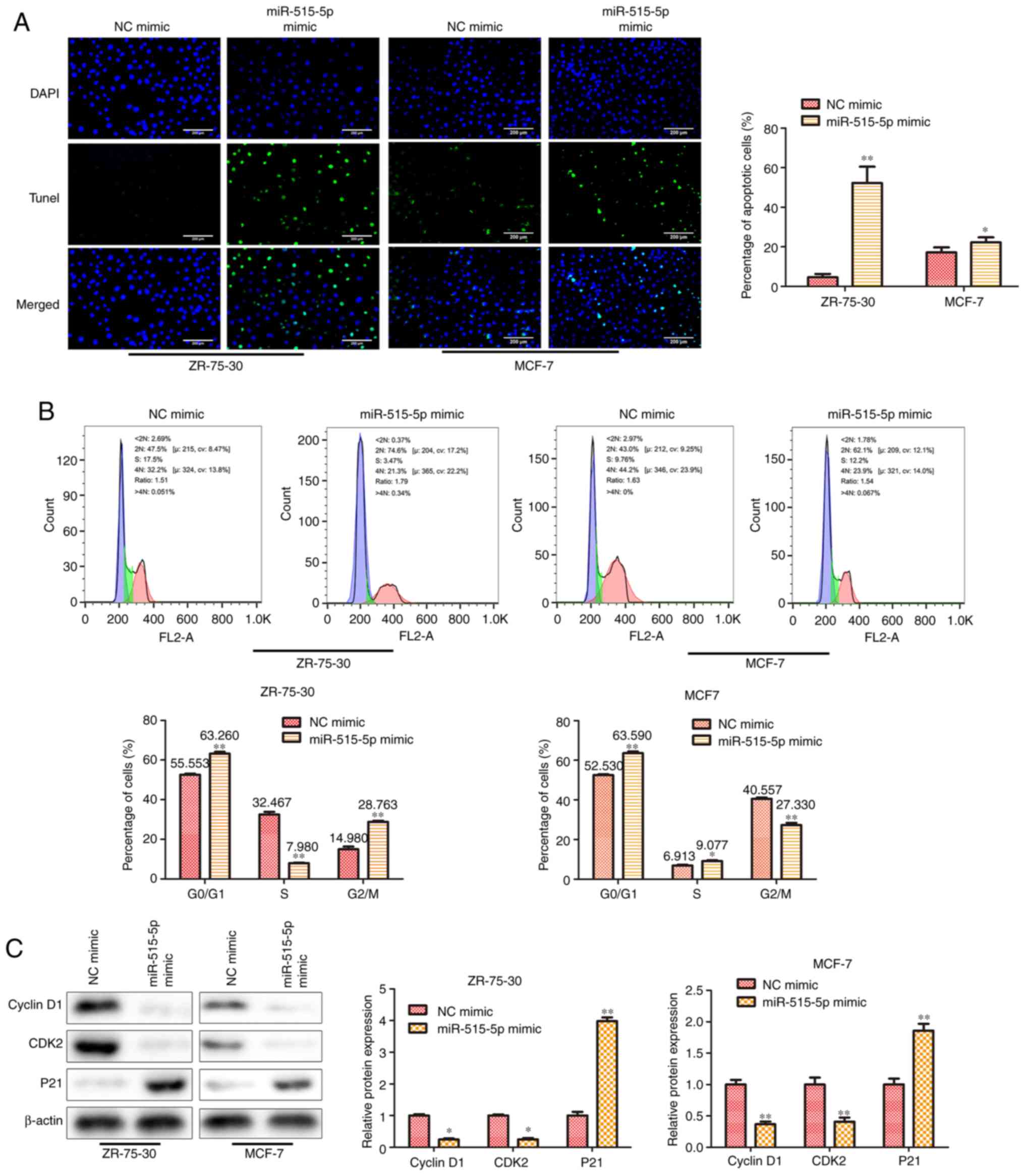 | Figure 2miR-515-5p overexpression facilitates
cell apoptosis and induces cell cycle arrest at the
G0/G1 phase in MCF7 and ZR-75-30 cells. MCF7
and ZR-75-30 cells were transfected with miR-515-5p mimic or the NC
mimic. (A) TUNEL assay was performed to assess cell apoptosis.
Scale bars, 200 µm. (B) Flow cytometric assay was performed to
assess cell cycle progression. (C) Western blot analysis was used
to assess the expression of cyclin D1, CDK2 and p21 proteins. The
data are presented as the mean ± SD. **P<0.01 and
*P<0.05 vs. NC mimic. miR, miRNA; NC, negative
control; CDK2, cyclin dependent kinase 2; PCNA, proliferating cell
nuclear antigen; P21, cyclin-dependent kinase inhibitor; <2N,
G0 phase; S, S phase; 4N, G2/M. |
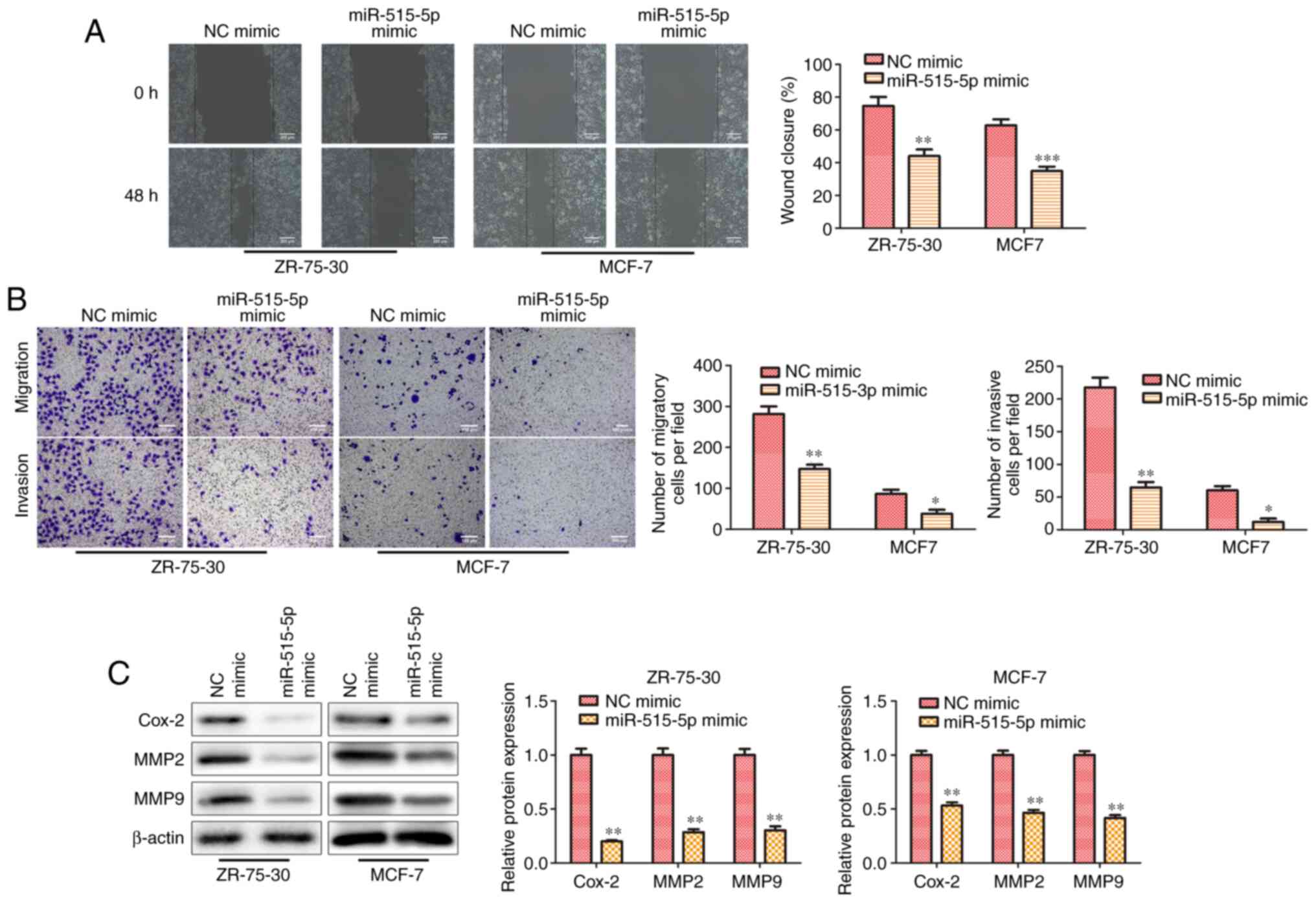
miR-515-5p inhibits cell migration of
MCF7 and ZR-75-30 cells
To further investigate the effect of miR-515-5p on
the migration and invasion in BC cells, wound healing and Transwell
assays were performed. Results from the wound healing assay
indicated that miR-515-5p mimic transfection significantly
decreased the wound closing abilities of MCF7 and ZR-75-30 cells
compared with those in cells transfected with the NC mimic
(P<0.01; Fig. 3A), suggesting
that miR-515-5p overexpression could inhibit cell migration.
Similarly, Transwell assay results also demonstrated that
miR-515-5p mimic transfection significantly inhibited cell invasion
in MCF7 and ZR-75-30 cells compared with that in cells transfected
with the NC mimic (P<0.05 and P<0.01; Fig. 3B). Subsequently western blot
analysis showed that miR-515-5p mimic transfection decreased the
expression levels of migration-related proteins MMP2, MMP9 and
Cox-2 compared with those in cells transfected with the NC mimic
(P<0.01; Fig. 3C).
Collectively, these results suggest that miR-515-5p overexpression
decreased the migratory and invasive abilities of BC cells in
vitro.
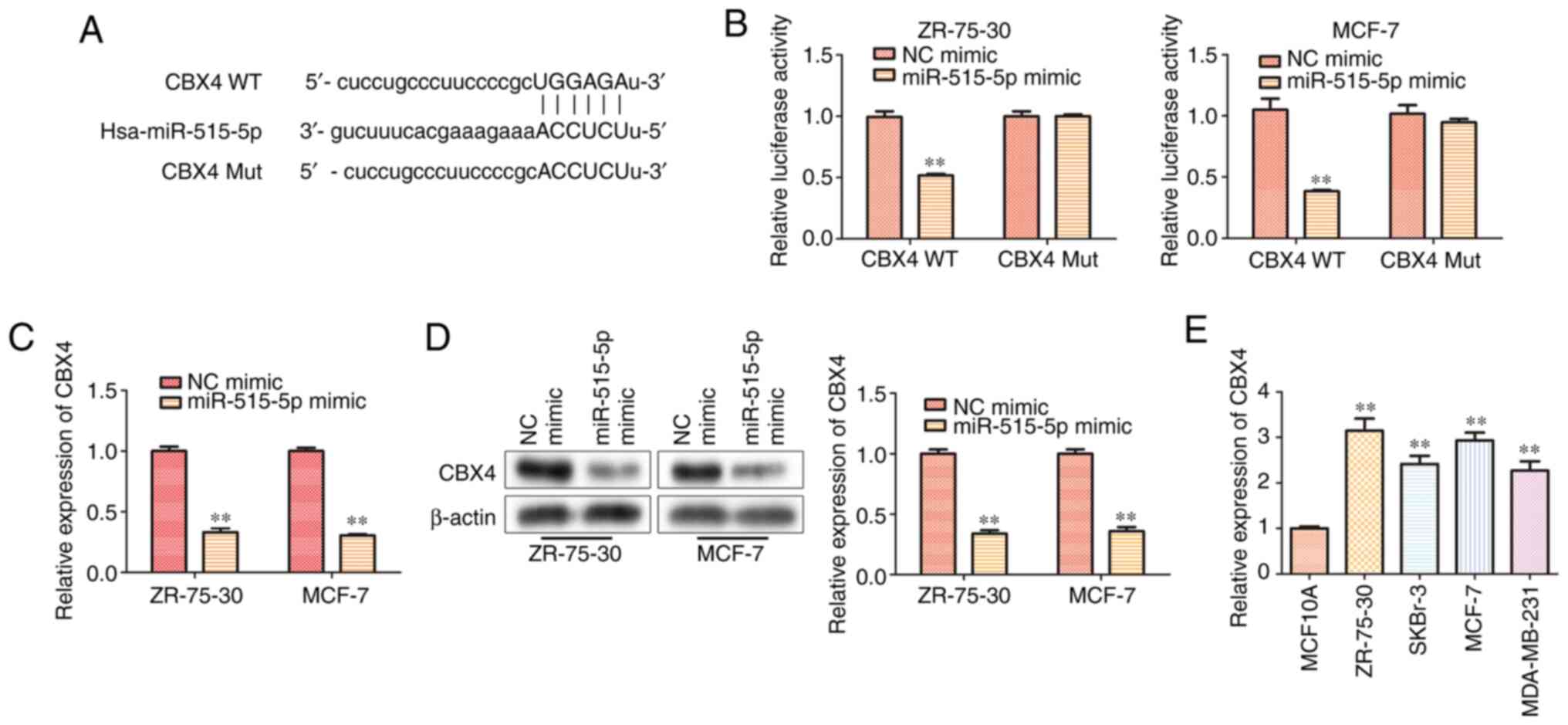
miR-515-5p inhibits CBX4 expression by
directly binding to the 3'-UTR of CBX4
To explore the underlying mechanism of miR-515-5p in
regulating BC progression, StarBase was used to screen for
miR-515-5p target genes, where and CBX4 was found to harbor
complementary binding sites for miR-515-5p (Fig. 4A). Therefore, a dual-luciferase
reporter assay was used to verify this interaction. Consistent with
bioinformatics analysis, the luciferase reporter assay results
indicated that the miR-515-5p mimic could significantly
downregulate the luciferase activity of CBX4 WT but not that of
CBX4 Mut (P<0.01; Fig. 4B).
Both RT-qPCR (Fig. 4C) and western
blot analysis (Fig. 4D) results
demonstrated that miR-515-5p mimic transfection could significantly
decrease the mRNA and protein levels of CBX4 compared with those in
cells transfected with the NC mimic (P<0.01). To confirm if CBX4
participates in BC progression, its expression level in BC cell
lines MCF7, ZR-75-30, MDA-MB-231, SKBr-3 and the normal breast cell
line MCF10A was compared using RT-qPCR. The results showed that the
expression of CBX4 in human BC lines was significantly higher
compared with that in the normal breast cell line (P<0.01;
Fig. 4E). These data suggest that
miR-515-5p could inhibit the gene expression of CBX4 by directly
binding to the 3'-UTR of CBX4 in BC cells.
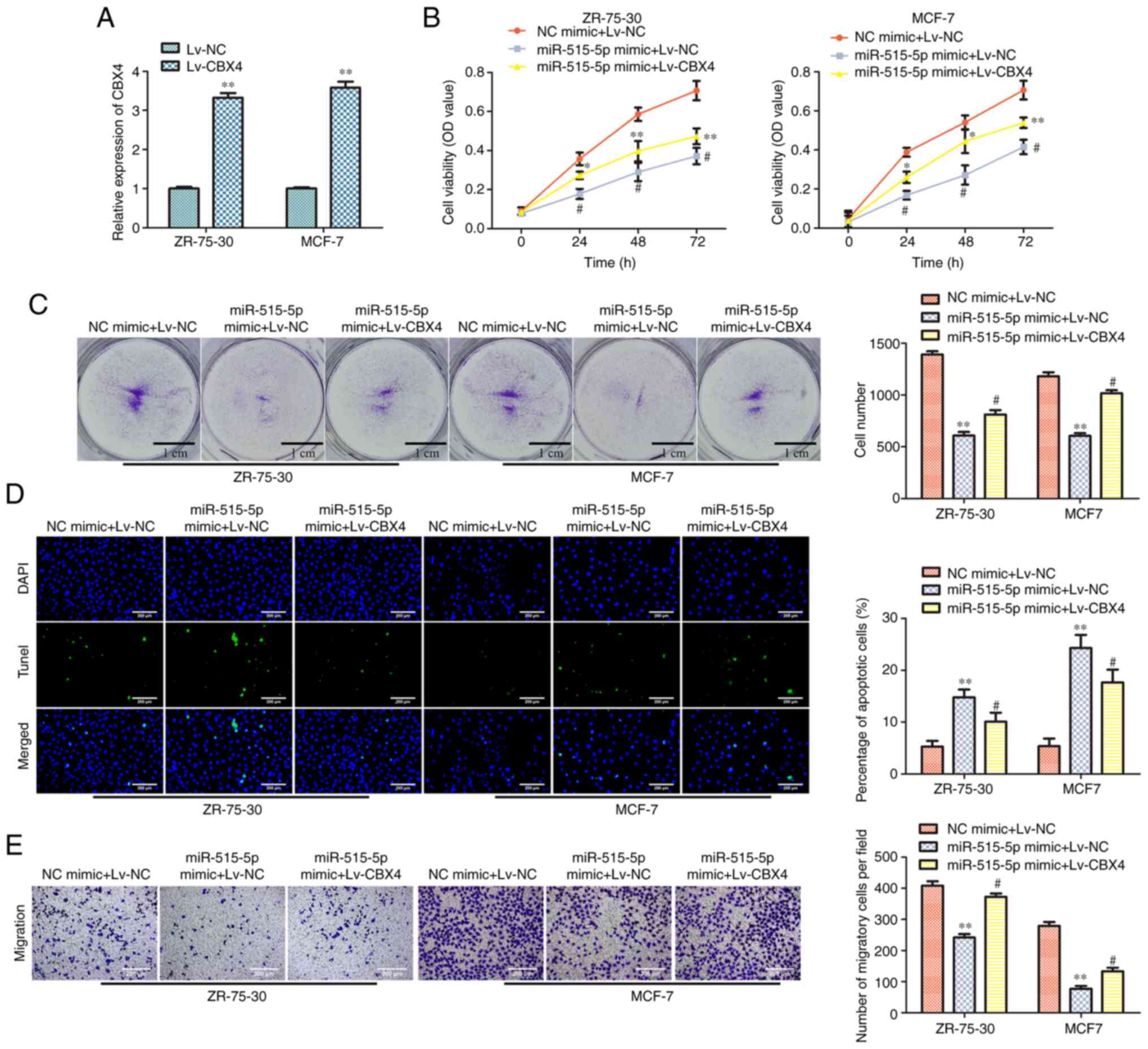
Overexpression of CBX4 reverses the
effects of miR-515-5p mimic in MCF7 and ZR-75-30 cells
To confirm the association between miR-515-5p and
CBX4, MDA-MB-157 cells were co-transfected with miR-515-5p or NC
mimics and Lv-CBX4 or Lv-NC. RT-qPCR data showed that Lv-CBX4
transfection could significantly upregulate the expression of CBX4
in MCF7 and ZR-75-30 cells compared with that in cells transfected
with the Lv-NC (P<0.01; Fig.
5A). To test the effect of the overexpression of CBX4 in BC
cells viability, CCK-8 assay was conducted. The results showed that
co-transfection with miR-515-5p mimic and Lv-CBX4 could
significantly increase the viability of MCF7 and ZR-75-30 cells
compared with that in cells co-transfected with the miR-515-5p
mimic and Lv-NC (P<0.05; Fig.
5B). Colony formation assay results showed that Lv-CBX4
co-transfection could promote the colony formation of MCF7 and
ZR-75-30 cells transfected with miR-515-5p mimic, as compared with
that of cells transfected with the Lv-NC and the miR-515 mimic
(P<0.05; Fig. 5C). TUNEL assay
results indicated that Lv-CBX4 co-transfection could inhibit the
apoptosis of MCF7 and ZR-75-30 cells transfected with the
miR-515-5p mimic compared with that in cell co-transfected with
Lv-NC and miR-515 (P<0.05 and P<0.01; Fig. 5D). Finally, Transwell assay data
revealed that Lv-CBX4 co-transfection could promote the migration
of MCF7 and ZR-75-30 cells transfected with the miR-515-5p mimic
compared with that in cells co-transfected with Lv-NC and
miR-515-5p mimic (P<0.05 and P<0.01; Fig. 5E). These results suggest that the
overexpression of CBX4 can partially reverse the effects of
miR-515-5p overexpression on BC cell downregulation.
Discussion
BC is the leading cause of cancer-related mortality
in women worldwide (1). Since BC
is a multifactorial disease, unraveling the molecular changes
associated with this type of cancer can potentially provide
preventative and therapeutic strategies for this disease (3,16).
In BC, a number of miRNAs, such as miR-148b, miR-376c, miR-409-3p
and miR-801 have been identified as potential therapeutic targets
(17,18). In the present study, miR-515-5p
expression was found to be downregulated in BC cell lines. Since
the expression of miR-515-5p was the lowest in MCF7 and ZR-75-30
cells out of all BC cell lines tested, these two cell lines were
selected to be models for further study. It was found that
miR-515-5p overexpression inhibited the proliferation, migration
and invasion of MCF7 and ZR-75-30 cells by suppressing the
expression of CBX4. This preliminary study indicated that
miR-515-5p may be a promising candidate for BC treatment.
miRNAs serve important roles in the onset,
progression and metastasis of cancer by regulating the stability of
target mRNAs or by inhibiting their translation (19). For example, miR-107 could inhibit
autophagy, proliferation and migration in BC cells (MDA-MB-231,
MDA-MB-453) by targeting high mobility group box 1 protein
(20). There are numerous studies
on the association between miR-515-5p and cancer. For instance, low
miR-515-5p levels were found to be correlated with poor survival in
samples from patients with gastric cancer on The Cancer Genome
Atlas database (21). In addition,
high miR-515-5p expression was correlated with increased survival
of patients with BC, such that miR-515-5p can control cancer cell
migration by regulating microtubule affinity regulating kinase
4(22). In BC, miR-515-5p was
significantly downregulated in estrogen receptor (ER)-positive
compared with that in ER-negative BC (23). Furthermore, miR-515-5p was
previously found to partly modulate the proliferation of BC cells
by regulating the Wnt pathway (23).
The present data showed that increasing the
expression of miR-515-5p inhibited the proliferation of MCF7 and
ZR-75-30 cells by facilitating cell apoptosis whilst arresting the
cell cycle at the G0/G1 phase. Western blot
analysis confirmed that the expression of cyclin D1, CDK2 that
involved in G0/G1 phase were decreased and
p21 proteins were increased by transfection with the miR-515-5p
mimic. These findings were also supported by the observations the
migration and invasion-associated protein expression of Cox-2, MMP2
and MMP9 were decreased by miR-515-5p mimic transfection.
Subsequently, CBX4 was predicted by StarBase to be a direct target
of miR-515-5p, which was further validated by luciferase report
assays to demonstrate that miR-515-5p interacts directly with the
3'UTR of CBX4.
CBX4, also known as hPC2 or NBP16, is a canonical
component of the polycomb repressive complex 1, which is associated
with a variety of tumors (24).
CBX4 is directly involved in the DNA damage response pathway and
regulates DNA modification, stability and terminal repair (24). CBX4 has been found to promote
proliferation and metastasis in lung cancer (25) and hepatocellular carcinoma
(26). CBX4 has also been reported
to mediate miR-129-5p-induced inhibition of cell proliferation in
BC (27). The present results
suggest that miR-515-5p overexpression can decrease the expression
of CBX4 in MCF7 and ZR-75-30 cells by directly interacting with the
3'UTR of CBX4. Additionally, overexpression of CBX4 reversed the
effects of miR-515-5p on the proliferation, migration and invasion
of MCF7 and ZR-75-30 cells. CBX4 has been reported to be involved
in the small ubiquitin-like modifier (SUMO) modification of DNA
methyltransferase 3A, PR domain containing 16, B cell-specific
moloney murine leukemia virus integration site 1 and
hypoxia-induced factor-1α, as a SUMO E3 ligase (24,28-30).
Therefore, miR-515-5p could mediate the SUMO modification states of
proteins in breast cancer cells by targeting CBX4.
In conclusion, the present study demonstrated the
function of miR-515-5p in MCF7 and ZR-75-30 cells as a breast
cancer model. Specifically, miR-515-5p could regulate the cell
viability and proliferation of BC cells by targeting CBX4, thereby
inhibiting the migration and invasion of BC cells. The present
study promoted the understanding of the mechanism via which
miR-515-5p regulates the progression and metastasis of BC.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by The Tianjin
Municipal Education Commission Scientific Research Plan Project
(grant no. 2020KJ145).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PYT and LJW designed the study and performed the
experiments. PYT and YSW analyzed the data. LJW and PYT confirm the
authenticity of all the raw data. All authors read and approved the
final version of this manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
DeSantis CE, Ma J, Gaudet MM, Newman LA,
Miller KD, Goding Sauer A, Jemal A and Siegel RL: Breast cancer
statistics, 2019. CA Cancer J Clin. 69:438–451. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhang ML, Peng P, Wu CX, Gong YM, Zhang
SW, Chen WQ and Bao PP: Report of breast cancer incidence and
mortality in China registry regions, 2008-2012. Zhonghua Zhong Liu
Za Zhi. 41:315–320. 2019.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
3
|
Wu H, Wang Q, Zhong H, Li L, Zhang Q,
Huang Q and Yu Z: Differentially expressed microRNAs in exosomes of
patients with breast cancer revealed by next-generation sequencing.
Oncol Rep. 43:240–250. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Rossing M, Sørensen CS, Ejlertsen B and
Nielsen FC: Whole genome sequencing of breast cancer. APMIS.
127:303–315. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Lytle JR, Yario TA and Steitz JA: Target
mRNAs are repressed as efficiently by microRNA-binding sites in the
5' UTR as in the 3' UTR. Proc Natl Acad Sci USA. 104:9667–9672.
2007.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ma C, Miao C, Wang C, Song F and Luo M:
PELP1 is a novel oncogene in gastric tumorigenesis and negatively
regulated by miR-15 family microRNAs. Cancer Biomark. 26:1–9.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhang C, Liu J, Tan C, Yue X, Zhao Y, Peng
J, Wang X, Laddha SV, Chan CS, Zheng S, et al: microRNA-1827
represses MDM2 to positively regulate tumor suppressor p53 and
suppress tumorigenesis. Oncotarget. 7:8783–8796. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kim S, Lee E, Jung J, Lee JW, Kim HJ, Kim
J, Yoo HJ, Lee HJ, Chae SY, Jeon SM, et al: microRNA-155 positively
regulates glucose metabolism via PIK3R1-FOXO3a-cMYC axis in breast
cancer. Oncogene. 37:2982–2991. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Shao B, Wang X, Zhang L, Li D, Liu X, Song
G, Cao H, Zhu J and Li H: Plasma microRNAs predict chemoresistance
in patients with metastatic breast cancer. Technol Cancer Res
Treat. 18(1533033819828709)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Hromadníková I, Kotlabová K, Jirásek JE
and Doucha J: Detection of placenta-specific microRNAs in maternal
circulation. Ceska Gynekol. 75:252–256. 2010.PubMed/NCBI(In Czech).
|
|
11
|
Zheng J, Sadot E, Vigidal JA, Klimstra DS,
Balachandran VP, Kingham TP, Allen PJ, D'Angelica MI, DeMatteo RP,
Jarnagin WR and Ventura A: Characterization of hepatocellular
adenoma and carcinoma using microRNA profiling and targeted gene
sequencing. PLoS One. 13(e0200776)2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang X, Zhou J, Xue D, Li Z, Liu Y and
Dong L: MiR-515-5p acts as a tumor suppressor via targeting TRIP13
in prostate cancer. Int J Biol Macromol. 129:227–232.
2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Li J, Tang Z, Wang H, Wu W, Zhou F, Ke H,
Lu W, Zhang S, Zhang Y, Yang S, et al: CXCL6 promotes non-small
cell lung cancer cell survival and metastasis via down-regulation
of miR-515-5p. Biomed Pharmacother. 97:1182–1188. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Chen Y, Du J, Wang Y, Shi H, Jiang Q, Wang
Y, Zhang H, Wei Y, Xue W, Pu Z, et al: MicroRNA-497-5p induces cell
cycle arrest of cervical cancer cells in s phase by targeting CBX4.
Onco Targets Ther. 12:10535–10545. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Rodriguez LG, Wu X and Guan JL:
Wound-healing assay. Methods Mol Biol. 294:23–29. 2005.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Das PK, Siddika MA, Asha SY, Aktar S,
Rakib MA, Khanam JA, Pillai S and Islam F: MicroRNAs, a promising
target for breast cancer stem cells. Mol Diagn Ther. 24:69–83.
2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Cuk K, Zucknick M, Heil J, Madhavan D,
Schott S, Turchinovich A, Arlt D, Rath M, Sohn C, Benner A, et al:
Circulating microRNAs in plasma as early detection markers for
breast cancer. Int J Cancer. 132:1602–1612. 2013.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Bahrami A, Aledavood A, Anvari K,
Hassanian SM, Maftouh M, Yaghobzade A, Salarzaee O, ShahidSales S
and Avan A: The prognostic and therapeutic application of microRNAs
in breast cancer: Tissue and circulating microRNAs. J Cell Physiol.
233:774–786. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hafez MM, Hassan ZK, Zekri AR, Gaber AA,
Al Rejaie SS, Sayed-Ahmed MM and Al Shabanah O: MicroRNAs and
metastasis-related gene expression in Egyptian breast cancer
patients. Asian Pac J Cancer Prev. 13:591–598. 2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ai H, Zhou W, Wang Z, Qiong G, Chen Z and
Deng S: microRNAs-107 inhibited autophagy, proliferation, and
migration of breast cancer cells by targeting HMGB1. J Cell
Biochem: Dec 2, 2018 (Epub ahead of print). doi:
10.1002/jcb.28157.
|
|
21
|
Wang D, Liu K and Chen E: LINC00511
promotes proliferation and invasion by sponging miR-515-5p in
gastric cancer. Cell Mol Biol Lett. 25(4)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Pardo OE, Castellano L, Munro CE, Hu Y,
Mauri F, Krell J, Lara R, Pinho FG, Choudhury T, Frampton AE, et
al: miR-515-5p controls cancer cell migration through MARK4
regulation. EMBO Rep. 17:570–584. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Pinho FG, Frampton AE, Nunes J, Krell J,
Alshaker H, Jacob J, Pellegrino L, Roca-Alonso L, de Giorgio A,
Harding V, et al: Downregulation of microRNA-515-5p by the estrogen
receptor modulates sphingosine kinase 1 and breast cancer cell
proliferation. Cancer Res. 73:5936–5948. 2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ismail IH, Gagné JP, Caron MC, McDonald D,
Xu Z, Masson JY, Poirier GG and Hendzel MJ: CBX4-mediated SUMO
modification regulates BMI1 recruitment at sites of DNA damage.
Nucleic Acids Res. 40:5497–5510. 2012.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hu C, Zhang Q, Tang Q, Zhou H, Liu W,
Huang J, Liu Y, Wang Q, Zhang J, Zhou M, et al: CBX4 promotes the
proliferation and metastasis via regulating BMI-1 in lung cancer. J
Cell Mol Med. 24:618–631. 2020.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Tan C, Bei C, Zhu X, Zhang Y, Qin L and
Tan S: Single nucleotide polymorphisms of CBX4 and CBX7 decrease
the risk of hepatocellular carcinoma. Biomed Res Int.
2019(6436825)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Meng R, Fang J, Yu Y, Hou LK, Chi JR, Chen
AX, Zhao Y and Cao XC: miR-129-5p suppresses breast cancer
proliferation by targeting CBX4. Neoplasma. 65:572–578.
2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chen Q, Huang L, Pan D, Zhu LJ and Wang
YX: Cbx4 sumoylates Prdm16 to regulate adipose tissue
thermogenesis. Cell Rep. 22:2860–2872. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Li B, Zhou J, Liu P, Hu J, Jin H, Shimono
Y, Takahashi M and Xu G: Polycomb protein Cbx4 promotes SUMO
modification of de novo DNA methyltransferase Dnmt3a. Biochem J.
405:369–378. 2007.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Li J, Xu Y, Long XD, Wang W, Jiao HK, Mei
Z, Yin QQ, Ma LN, Zhou AW, Wang LS, et al: Cbx4 governs HIF-1α to
potentiate angiogenesis of hepatocellular carcinoma by its SUMO E3
ligase activity. Cancer Cell. 25:118–131. 2014.PubMed/NCBI View Article : Google Scholar
|