Introduction
Diabetic nephropathy (DN) is a microvascular
complication of diabetes that represents the leading cause of
mortality among patients with type 1 diabetes mellitus (DM) and the
second leading cause of mortality among those with type 2 DM. DN is
also the primary cause of end-stage renal disease (ESRD) (1,2).
Therefore, studying the mechanism of DN pathogenesis and
controlling its progression is urgently required.
The pathogenesis of DN is complex and includes
hemodynamic changes, glucose and lipid metabolism disorders,
autophagy, inflammation and growth factor disorders (3-5).
Mesangial cells are important inherent cells in the glomerulus
which, together with the extracellular matrix (ECM) they secrete,
constitute the glomerular mesangial region and play an important
role in supporting the capillary loop (6,7). In
glomerular diseases, mesangial cells are one of the most important
effector cells involved in the disease process. Mesangial cells can
promote cell proliferation and ECM deposition by auto-activation
and secretion of several bioactive molecules to affect the function
of other inherent cells in the glomeruli through paracrine
mechanisms, in turn participating in the process of glomerular
injury or repair and ultimately promoting the occurrence and
development of glomerulosclerosis (8,9).
Therefore, exploring the mechanism of abnormal mesangial cell
function in diabetes will provide a reliable theoretical basis for
developing a more effective therapeutic approach.
Yes-associated protein 1 (YAP1) is an oncogenic
transcriptional co-regulator and the effector molecule of a variety
of signaling pathways. The expression level of YAP1 was found to be
higher in activated fibroblasts. Moreover, YAP1 may promote the
activation of fibroblasts and participate in the secretion of ECM
by regulating the expression of activation-associated genes,
including growth factors (10,11).
Podocyte injury has been reported to stimulate the activity of
transcriptional coactivator YAP and the expression of the YAP
target gene in a rat glomerular disease model; in addition, YAP
overexpression increased the levels of ECM-associated proteins,
which may lead to fibrosis (12).
In a different study, Chen et al (13) found that YAP1 knockdown inhibited
the deposition of ECM and ultimately improved pulmonary fibrosis
in vitro and in vivo. In addition, YAP1 has been
proven to promote tumorigenesis in vivo, in vitro and
in human specimens. YAP1 may activate the transcription of the
inflammatory cytokine IL-1β and induce the development of
inflammation-associated gastric cancer; therefore, YAP1 may
activate inflammatory cytokines to further aggravate the
inflammatory response (14).
However, the effect of YAP1 on a glomerular mesangial cell (GMC)
model of DN treated with high glucose has not yet been studied in
detail.
In the present study, the regulatory effect of YAP1
on mesangial cells treated with high glucose and the potential
underlying mechanism were explored. The results revealed that YAP1
promoted inflammation and ECM deposition by activating the
NF-κB/JMJD3 signaling pathway in high glucose-induced GMCs. The
findings of the present study may provide a new research direction
for the diagnosis and treatment of DN.
Materials and methods
Cell culture and treatment
The HBZY-1 renal mesangial cell line was purchased
from the American Type Culture Collection. HBZY-1 cells were
cultured in 5.5 mmol/l DMEM (Cytiva) containing 10% fetal bovine
serum (Cytiva) in a humidified atmosphere of 95% O2 and
5% CO2 at 37˚C. Cells were not used beyond passage 3.
HBZY-1 cells cultured in 5.5 mmol/l serum-free DMEM normal-glucose
media were used as the normal control group (NG) and cells cultured
in 30 mmol/l serum-free DMEM high-glucose media were used as the
high-glucose group (HG). Mannitol was used as a control to
eliminate the effect of osmotic pressure. The model cells were
treated with 30 mmol/l serum-free DMEM for 0, 6, 12, 24 and 48
h.
Cell transfection
The YAP1 overexpression vector pcDNA-TRIM31
(Oe-YAP1) and empty control vector pcDNA (Oe-NC) were constructed
by Shanghai GenePharma Co., Ltd. Small interfering RNA (siRNA)
against YAP1 (si-YAP1) and siRNA negative control (si-NC) were
synthesized by Shanghai GenePharma Co., Ltd. A final concentration
of 100 nM plasmids were transfected into cells using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
transfection reagent. After 48 h transfection, the transfection
efficiency was detected and cells were collected for subsequent
experiments.
Cell counting kit-8 (CCK-8) assay
Cells were inoculated into 96-well plates
(5x103 cells/well) during the logarithmic growth phase
and 3 repeated wells were set. CCK-8 solution (10 µl) was added to
each well, and cells were incubated at 37˚C. The absorbance value
in each well was detected at 450 nm at 72 h using a microplate
reader (BioTek Instruments, Inc.).
Enzyme-linked immunosorbent assay
(ELISA)
The levels of tumor necrosis factor (TNF)-α,
interleukin (IL)-1β and IL-6 in each group were determined by ELISA
performed using respective ELISA detection kits (cat. no. EK0526,
EK0393 and EK0412; Wuhan Boster Biological Technology, Ltd.),
according to the manufacturer's instructions.
Immunofluorescence
Immunofluorescence was used to detect the expression
level of collagen IV in HBZY-1 cells. The glass slides containing
HBZY-1 cells were washed by PBS twice, and then fixed with 4%
paraformaldehyde for 10 min at room temperature. Next, cells were
permeabilized with 0.5% Triton X-100 for 10 min at room temperature
and sealed with 5% BSA for 30 min at room temperature. Primary
antibody against collagen IV (cat. no. Ab6586; dilution, 1:1,000;
Abcam) was added to the slide and incubated overnight at 4˚C.
Finally, the nucleus was stained with DAPI solution at room
temperature for 10 min, and the stained sections were examined.
Images were captured at x200 magnification under a light microscope
(Olympus Corporation).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.). RNA
was then reverse-transcribed into cDNA using the QuantiTect Reverse
Transcription kit (Qiagen AB) and the reaction was incubated at
25˚C for 5 min, 42˚C for 30 min, 85˚C for 5 min and then kept at
4˚C for 5 min. The PCR reaction was performed using the ABI 7500
Real Time PCR Detection System (ABI) following the Quantitect SYBR
Green PCR kit protocols (Qiagen AB). The relative expression of
target genes was calculated using the 2-ΔΔCq method
(15). GAPDH was used as the
internal control. The primer sequences were as follows: Yap1
forward, 5'-GCAACTCCAACCAGCAGCAACA-3'; Yap1 reverse,
5'-CGCAGCCTCTCCTTCTCCATCTG-3'; monocyte chemoattractant protein-1
(MCP-1) forward, 5'-ATAGCAGCCACCTTCATTCG-3'; MCP-1 reverse,
5'-TTCCCCAAGTCTCTGTATCT-3'; β-actin forward,
5'-TACATGGCTGGGGTGTTGAA-3'; and β-actin reverse
5'-AAGAGAGGCATCCTCACCCT-3'.
Western blot analysis
Total protein from the cells was extracted using
RIPA lysis buffer (Thermo Fisher Scientific, Inc.). The protein
concentration was determined using BCA protein quantitative kit
(Haimen Biyuntian Biological Technology Co., Ltd.). Next, protein
(30 µg per lane) was separated using 12% SDS-PAGE and then
transferred to a PVDF membrane. These membranes were blocked with
5% skimmed milk for 1 h at room temperature and incubated with
primary antibodies against YAP1 (cat. no. ab52771; dilution,
1:1,000), proliferating cell nuclear antigen (PCNA; cat. no. ab29;
dilution, 1:1,000), MCP-1 (cat. no. ab214819; dilution, 1:1,000),
collagen IV (cat. no. ab6586; dilution, 1:1,000), α-smooth muscle
actin (α-SMA; cat. no. ab240678; dilution, 1:1,000), fibronectin
(FN; cat. no. ab6328; dilution, 1:1,000), connective tissue growth
factor (CTGF; cat. no. ab209780; dilution, 1:1,000), TGF-β1 (cat.
no. ab215715; dilution, 1:1,000), phosphorylated inhibitor of NF-κB
(p-IκBα; cat. no. ab225650; dilution, 1:1,000), p-p65 (cat. no.
ab31624; dilution, 1:1,000), p-65 (cat. no. ab32536; dilution,
1:1,000), Jumonji domain-containing protein D3 (JMJD3; cat. no.
ab38113; dilution, 1:1,000) and GAPDH (cat. no. ab9485; dilution,
1:1,000) overnight at 4˚C (all from Abcam). Subsequently, these
membranes were incubated with the corresponding mouse anti-rabbit
and goat anti-mouse IgG secondary antibodies conjugated to
horseradish peroxidase (sc-2357 and sc-2005, respectively;
dilution, 1:5,000; Santa Cruz Biotechnology, Inc.) for 2 h at room
temperature. Immunoreactive protein bands were visualized using an
ECL kit (EMD Millipore) and quantified using densitometry
(QuantityOne 4.5.0 software; Bio-Rad Laboratories, Inc.). GAPDH was
used as an internal control.
Statistical analysis
Experimental data were analyzed using the SPSS 20.0
software (IBM Corp.), and the results are expressed as the mean ±
SEM. Unpaired, two-tailed Student's t-test was performed for
comparisons between two groups. One-way analysis of variance test
followed by Dunnett's post hoc test was performed for comparisons
among multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
YAP1 is upregulated in HBZY-1 cells
treated with high glucose
In order to study the role and mechanism of action
of YAP1 in a cell model of DN treated with high glucose, cells were
treated with 30 mmol/l serum-free DMEM high-glucose media. As shown
in Fig. 1A and B, the RT-qPCR and western blot analysis
results showed that YAP1 expression was increased in a
time-dependent manner. YAP1 expression peaked at 48 h. Therefore,
the cell treatment time was set as 48 h. In addition, YAP1
expression in cells was decreased following transfection with YAP1
siRNA. As shown in Fig. 1C,
compared with the HG + si-NC group, YAP1 expression was the lowest
in the HG + si-YAP1-1 group. Thus, si-YAP1-1 was used for the
subsequent experiments.
 | Figure 1YAP1 is upregulated in HBZY-1 cells
treated with high glucose. (A) RT-qPCR was used to measure the
expression of YAP1 in HBZY-1 cells treated with 30 mmol/l
serum-free DMEM medium for 0, 6, 12, 24 and 48 h, respectively. (B)
Western blotting was used to measure the expression of YAP1 in
HBZY-1 cells treated with 30 mmol/l serum-free DMEM medium for 0,
6, 12, 24 and 48 h, respectively. (C) Transfection efficiency of
YAP1 small interfering RNA was detected by RT-qPCR.
***P<0.001 vs. HG 0 h; ###P<0.001 vs.
HG + si-NC. YAP1, Yes-associated protein 1; RT-qPCR, reverse
transcription-quantitative PCR; si-, small interfering RNA; NC,
negative control; HG, high-glucose group; NG, normal control group;
MA, mannitol. |
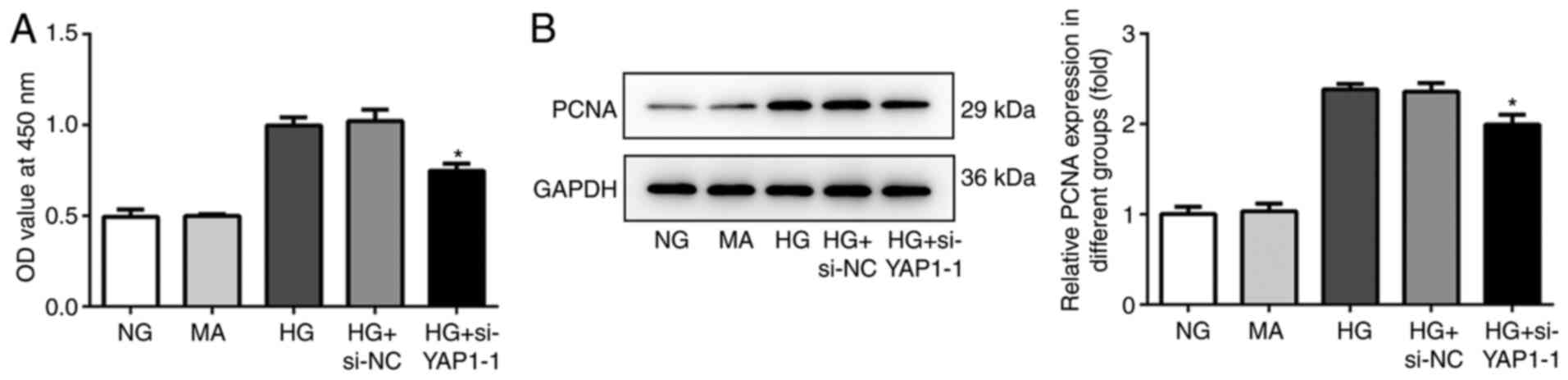
YAP1 knockdown inhibits the
proliferation of HBZY-1 cells treated with high glucose
CCK-8 assay and western blot analysis were used to
determine the effect of YAP1 on cell proliferation. The CCK-8
results showed that the cell viability in the HG + si-YAP1-1 group
was significantly lower compared with that in the HG + si-NC group,
indicating that YAP1 knockdown significantly decreased the
proliferation of HBZY-1 cells treated with high glucose (Fig. 2A). In addition, western blot
analysis results showed that YAP1 knockdown significantly decreased
the expression of proliferation-associated protein PCNA (Fig. 2B).
YAP1 knockdown suppresses the
inflammation of HBZY-1 cells treated with high glucose
Next, the expression level of inflammatory cytokines
was detected by ELISA. As shown in Fig.
3A, the ELISA results showed that YAP1 knockdown significantly
decreased the expression of the pro-inflammatory factors TNF-α,
IL-1β and IL-6. In addition, MCP-1, as an important inflammatory
mediator, was detected by RT-qPCR and western blot analyses. As
shown in Fig. 3B and C, YAP1 knockdown significantly decreased
MCP-1 expression and then enhanced the inflammation of HBZY-1 cells
treated with high glucose compared with the control group.
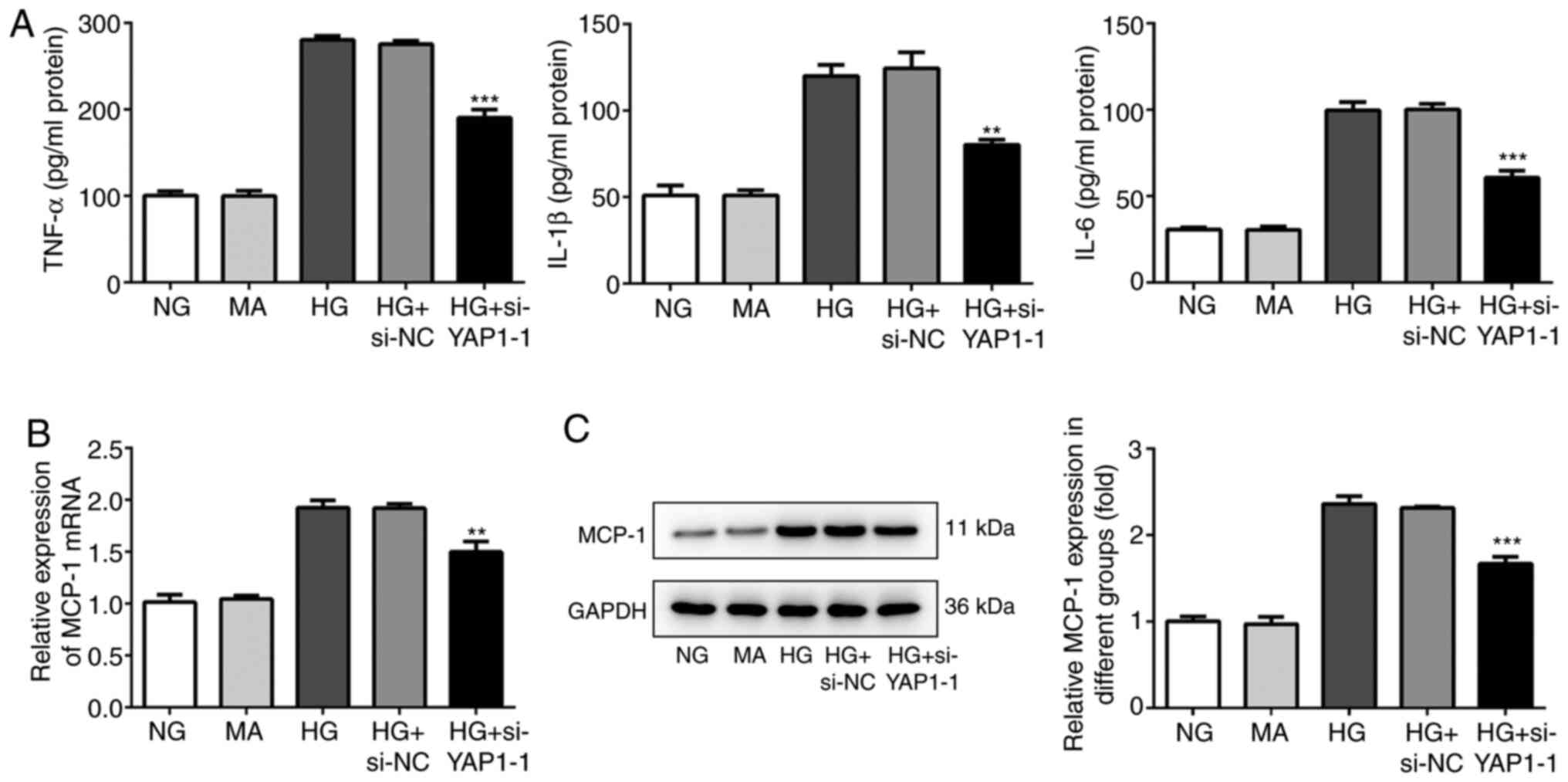 | Figure 3Knockdown of YAP1 suppresses the
inflammation of HBZY-1 cells induced by high glucose. (A) Effect of
YAP1 knockdown on the expressions of pro-inflammatory factors
(TNF-α, IL-1β and IL-6) was detected by enzyme-linked immunosorbent
assay. (B) Effect of YAP1 knockdown on the expression of MCP-1 was
detected by reverse transcription-quantitative PCR. (C) Effect of
YAP1 knockdown on the expression of MCP-1 was detected by western
blotting. **P<0.01 and ***P<0.001 vs.
HG + si-NC. YAP1, Yes-associated protein 1; si-, small interfering
RNA; NC, negative control; HG, high-glucose group; NG, normal
control group; MA, mannitol; TNF, tumor necrosis factor; IL,
interleukin; MCP-1, monocyte chemoattractant protein-1. |
YAP1 knockdown inhibits ECM deposition
and fibrosis in HBZY-1 cells treated with high glucose
Since the YAP1 inhibitor could significantly inhibit
the deposition of ECM and pulmonary fibrosis (11), the effect of YAP1 on ECM deposition
and fibrosis in HBZY-1 cells treated with high glucose was analyzed
by western blot analysis and immunofluorescence. As shown in
Fig. 4A, compared with the control
group, YAP1 knockdown significantly decreased the expression of
Leydig cell markers, including collagen IV, α-SMA and FN. The
expression of collagen IV was determined by immunofluorescence, and
the results showed that YAP1 knockdown significantly decreased the
fluorescence intensity of collagen IV (Fig. 4B and C). In addition, compared with the control
group, YAP1 knockdown significantly decreased the expression of the
fibrogenic factors CTGF and TGF-β1 (Fig. 4D). In general, YAP1 knockdown could
significantly enhance ECM deposition and fibrosis in HBZY-1 cells
treated with high glucose.
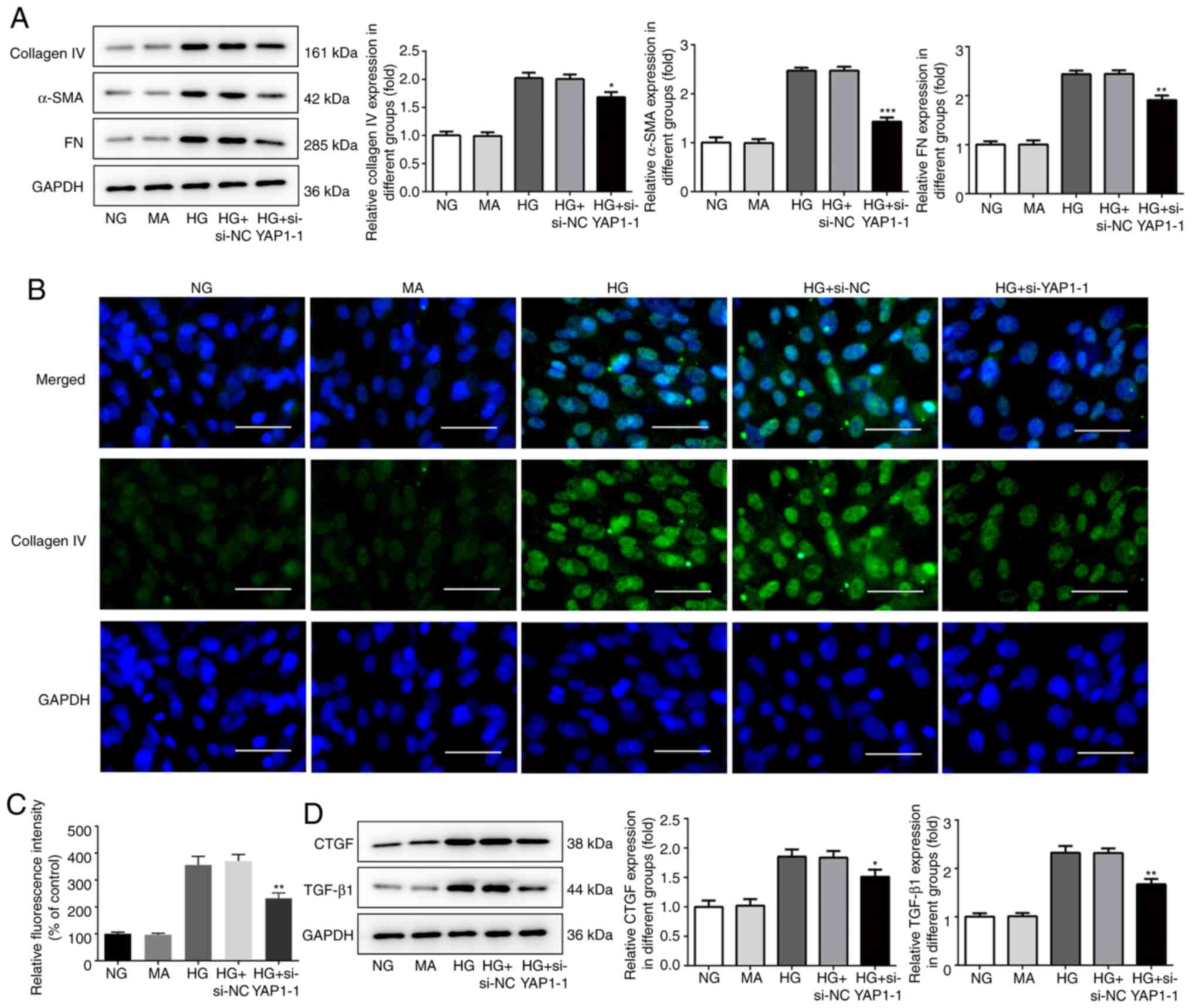 | Figure 4Knockdown of YAP1 inhibits the
deposition of extracellular matrix and fibrosis in HBZY-1 cells
induced by high glucose. (A) Effect of YAP1 knockdown on the
expressions of collagen IV, α-SMA and FN was detected by western
blotting. (B and C) Effect of YAP1 knockdown on the expressions of
collagen IV was detected by immunofluorescence. Scale bars, 50 µm.
(D) Effect of YAP1 knockdown on the expressions of fibrogenic
factors CTGF and TGF-β1 was detected by western blotting.
*P<0.05, **P<0.01 and
***P<0.001 vs. HG + si-NC. YAP1, Yes-associated
protein 1; si-, small interfering RNA; NC, negative control; HG,
high-glucose group; NG, normal control group; MA, mannitol; SMA,
smooth muscle actin; FN, fibronectin; CTGF, connective tissue
growth factor; TGF, transforming growth factor. |
YAP1 knockdown inhibits the
NF-κB/JMJD3 signaling pathway in HBZY-1 cells treated with high
glucose
To further study the mechanism of YAP1 on the HBZY-1
cell injury induced by high glucose, the effect of YAP1 on the
activation of the NF-κB/JMJD3 signaling pathway was detected by
western blot analysis. As shown in Fig.
5A and B, YAP1 knockdown
significantly inhibited the phosphorylation levels of IκBα and p65,
and decreased JMJD3 expression. These results suggested that YAP1
knockdown significantly suppressed the activation of the
NF-κB/JMJD3 signaling pathway.
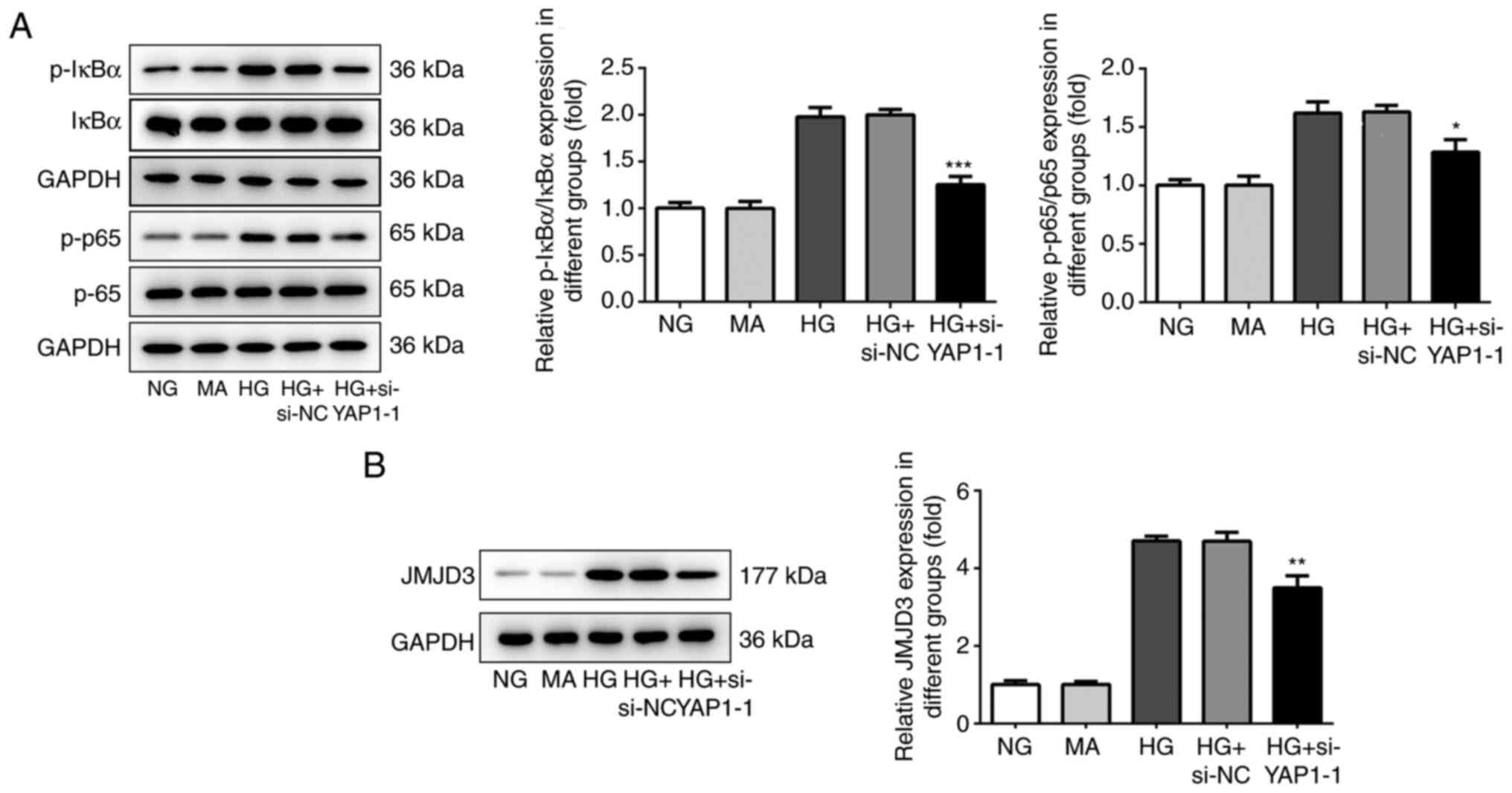 | Figure 5Knockdown of YAP1 inhibited
NF-κB/JMJD3 signal pathway in HBZY-1 cells induced by high glucose.
(A) Effect of YAP1 knockdown on the phosphorylation levels of IκBα
and p65 were detected by western blotting. (B) Effect of YAP1
knockdown on the expression of JMJD3 was detected by western
blotting. *P<0.05, **P<0.01 and
***P<0.001 vs. HG + si-NC. YAP1, Yes-associated
protein 1; si-, small interfering RNA; NC, negative control; HG,
high-glucose group; NG, normal control group; MA, mannitol; NF-κB,
nuclear factor-κB; JMJD3, Jumonji domain-containing protein D3; p-,
phosphorylated-; IκBα, inhibitor of NF-κB. |
Inhibitors of the NF-κB/JMJD3
signaling pathway attenuate the promoting effects of YAP1 on the
proliferation and inflammation of HBZY-1 cells treated with high
glucose
Next, YAP1 overexpression plasmid was constructed to
increase YAP1 expression in HBZY-1 cells (Fig. 6A). NF-κB inhibitor JSH23 and JMJD3
inhibitor GSK-J4 were added to the cells to inhibit the NF-κB/JMJD3
signaling pathway. As shown in Fig.
6B, JSH23 and GSK-J4 significantly decreased the
phosphorylation levels of IκBα and p65, as well as JMJD3
expression. In addition, the cell proliferation results showed that
JSH23 and GSK-J4 could partially reverse the promoting effect of
YAP1 overexpression on cell viability and MCP-1 expression
(Fig. 6C and D). ELISA and western blot analysis results
showed that JSH23 and GSK-J4 decreased the expression of TNF-α,
IL-1β, IL-6 and MCP-1.
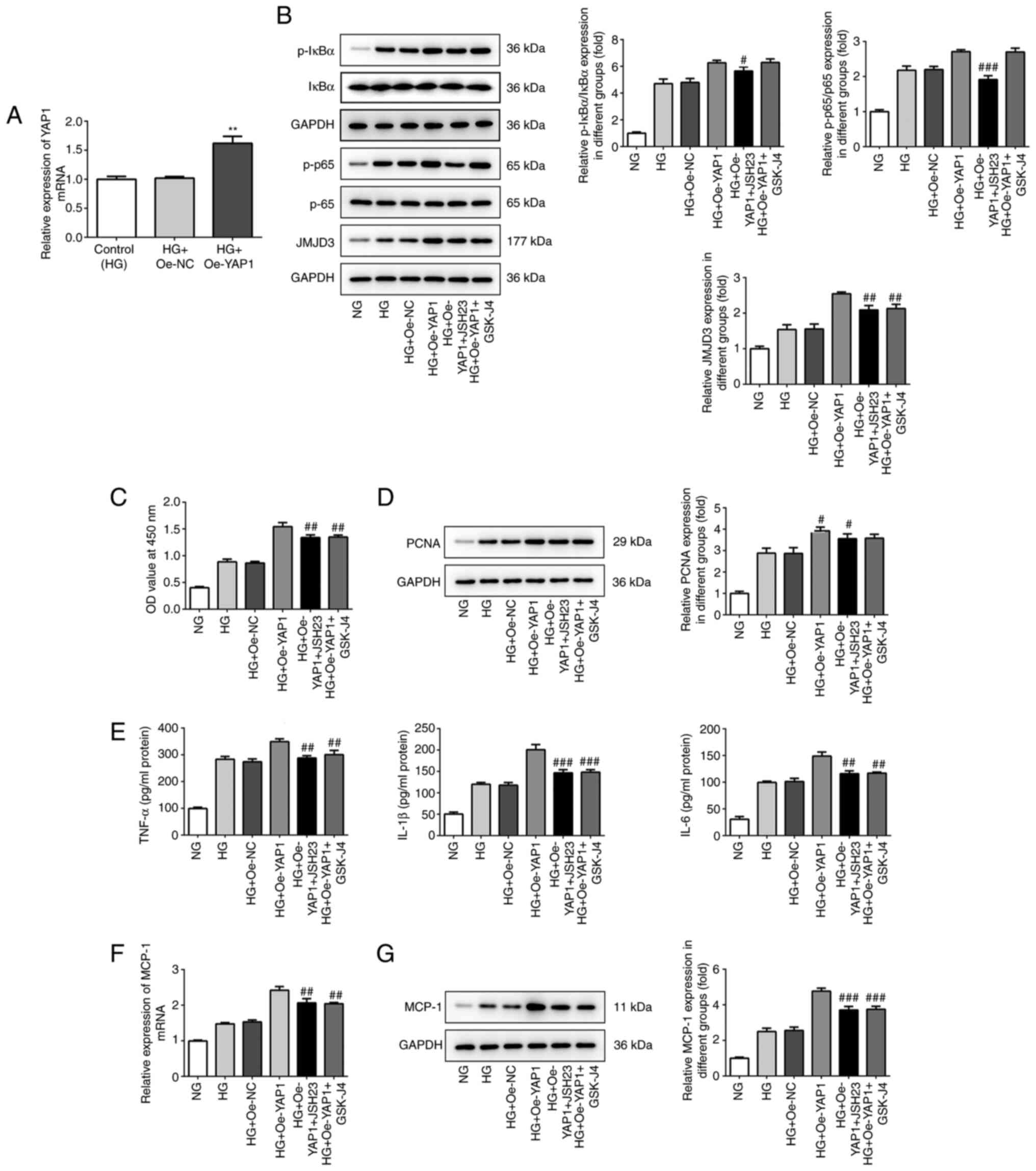 | Figure 6Inhibitors of NF-κB/JMJD3 signal
pathway attenuates the facilitatory effects of YAP1 on the
proliferation and inflammation of HBZY-1 cells induced by high
glucose. (A) The efficiency of YAP1 overexpression was determined
by RT-qPCR. (B) Effect of the inhibitors of NF-κB/JMJD3 signal
pathway on the expression levels of IκBα and p65 were detected by
western blotting. (C) Effect of the inhibitors of NF-κB/JMJD3
signal pathway on the viability of HBZY-1 cells treated with high
glucose was detected by Cell Counting Kit-8 assay. (D) Effect of
the inhibitors of NF-κB/JMJD3 signal pathway on the expression of
PCNA in the HBZY-1 cells treated with high glucose was measured by
western blotting. (E) Effect of the inhibitors of NF-κB/JMJD3
signal pathway on the expressions of pro-inflammatory factors
(TNF-α, IL-1β and IL-6) was detected by enzyme-linked immunosorbent
assay. (F) Effect of the inhibitors of NF-κB/JMJD3 signal pathway
on the expression of MCP-1 was detected by RT-qPCR. (G) Effect of
the inhibitors of NF-κB/JMJD3 signal pathway on the expression of
MCP-1 was detected by western blotting. **P<0.01 vs.
HG + Oe-NC. #P<0.05, ##P<0.01 and
###P<0.001 vs. HG + Oe-YAP1. YAP1, Yes-associated
protein 1; RT-qPCR, reverse transcription-quantitative PCR; Oe,
overexpression vector; NC, negative control; HG, high-glucose
group; NG, normal control group; MA, mannitol; NF-κB, nuclear
factor-κB; JMJD3, Jumonji domain-containing protein D3; p-,
phosphorylated-; IκBα, inhibitor of NF-κB; TNF, tumor necrosis
factor; IL, interleukin; MCP-1, monocyte chemoattractant
protein-1. |
Inhibitors of the NF-κB/JMJD3
signaling pathway attenuate the promoting effects of YAP1 on ECM
deposition and fibrosis of high-glucose-treated HBZY-1 cells
Finally, western blot analysis and
immunofluorescence assays were performed to determine the effect of
YAP1 on ECM deposition and fibrosis of HBZY-1 cells treated with
high glucose. As shown in Fig.
7A-C, YAP1 knockdown significantly promoted the expression of
collagen IV, α-SMA and FN. Similarly, as shown in Fig. 7D, YAP1 knockdown significantly
promoted the expression of CTGF and TGF-β1. These changes could be
partially reversed by the NF-κB/JMJD3 signaling pathway inhibitors
JSH23 and GSK-J4.
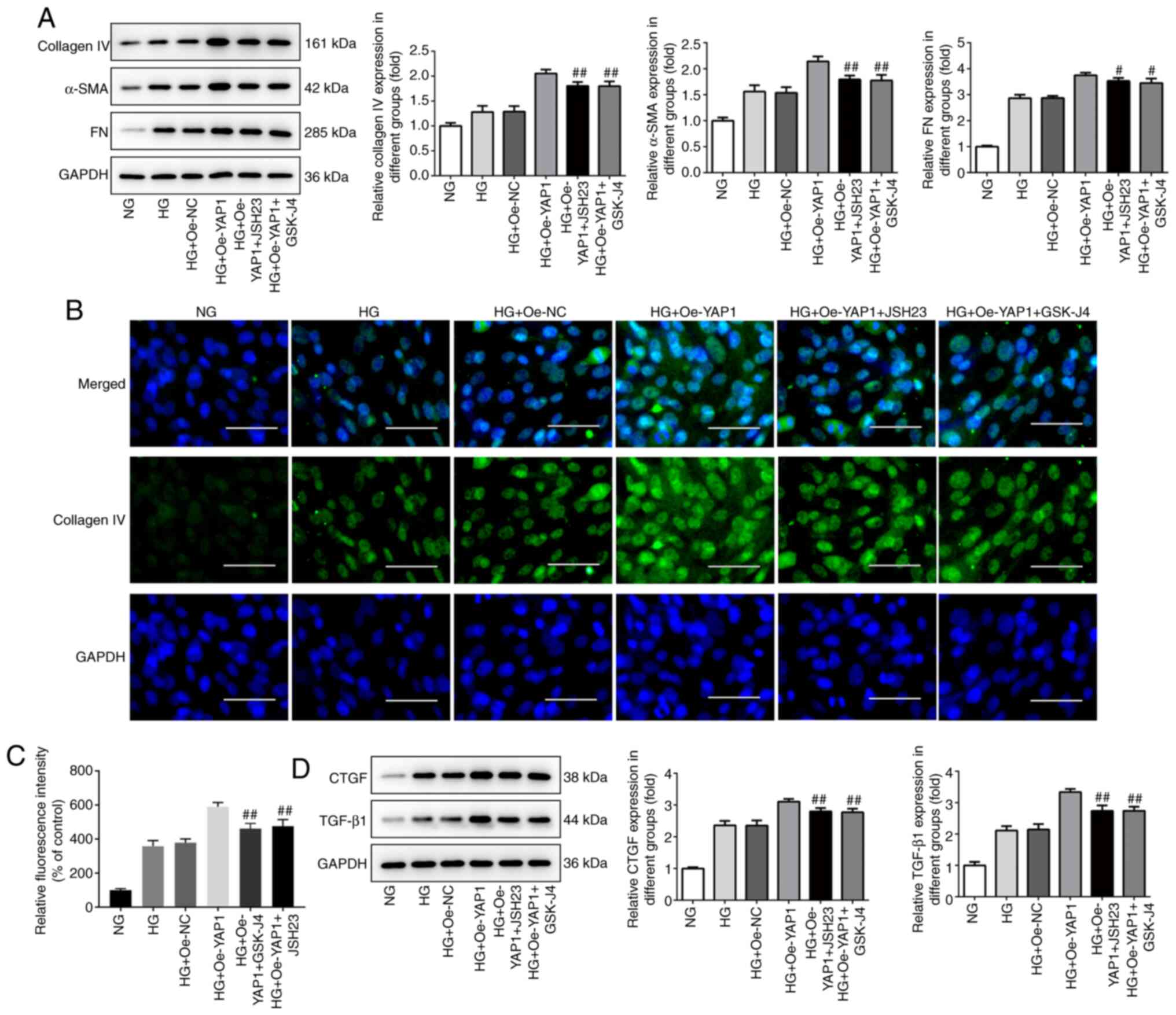 | Figure 7Inhibitors of NF-κB/JMJD3 signal
pathway attenuates the facilitatory effects of YAP1 on the
deposition of extracellular matrix and fibrosis of HBZY-1 cells
induced by high glucose. (A) Effect of the inhibitors of
NF-κB/JMJD3 signal pathway on the expression levels of collagen IV,
α-SMA and FN was detected by western blotting. (B and C) Effect of
the inhibitors of NF-κB/JMJD3 signal pathway on the expression
levels of collagen IV was detected by immunofluorescence. Scale
bars, 50 µm. (D) Effect of the inhibitors of NF-κB/JMJD3 signal
pathway on the expression levels of fibrogenic factors CTGF and
TGF-β1 was detected by western blotting. #P<0.05 and
##P<0.01 vs. HG + Oe-YAP1. YAP1, Yes-associated
protein 1; Oe, overexpression vector; NC, negative control; HG,
high-glucose group; NG, normal control group; MA, mannitol; NF-κB,
nuclear factor-κB; JMJD3, Jumonji domain-containing protein D3;
SMA, smooth muscle actin; FN, fibronectin; CTGF, connective tissue
growth factor; TGF, transforming growth factor. |
Discussion
DN is a microvascular complication of late-stage
diabetes and the main cause of ESRD (16). There is currently a lack of
sensitive indicators for the early diagnosis of DN; therefore,
identifying new diagnostic markers for DN is crucial. The results
of the present study revealed that YAP1 was upregulated in HBZY-1
cells treated with high glucose. YAP1 could promote the
proliferation, inflammation, ECM deposition and fibrosis of HBZY-1
cells by activating the NF-κB/JMJD3 pathway.
YAP transcriptional coactivator is a key effector of
the Hippo signaling pathway and plays an important role in
regulating cell proliferation, differentiation and tissue
homeostasis (17). YAP has been
shown to play an important role in DN. It is highly expressed in
renal tissue and associated with the levels of creatinine and blood
urea nitrogen, DN stage and pathological grade, which suggests that
it plays an important role in renal injury in type 2 DM (18). In addition, YAP overexpression has
been found to be closely associated with ECM and fibrosis (10). YAP1 knockdown could significantly
decrease the deposition of ECM and improve pulmonary fibrosis
(13). YAP1 has been shown to
promote tumor growth in both in vivo and in vitro
experiments. In combination with TEAD, YAP1 could activate the
transcription of IL-1β, a key inflammatory cytokine in gastric
cells, eventually leading to gastric cancer, since IL-1β can induce
inflammation-associated tumorigenesis. Therefore, YAP1 can play a
key role in increasing inflammation by activating inflammatory
cytokines (14). It was recently
demonstrated that YAP1 silencing may have a therapeutic effect on
diabetic retinopathy (19). Since
excessive proliferation of GMCs and ECM deposition are the main
pathological characteristics associated with the occurrence and
development of DN (20,21), it was hypothesized that YAP1 may
promote the inflammation and ECM deposition of GMCs induced by high
glucose. In the present study, YAP1 was upregulated in HBZY-1 cells
treated with high glucose. YAP1 knockdown significantly suppressed
the proliferation, inflammation, ECM deposition and fibrosis of
HBZY-1 cells treated with high glucose. These results highlighted
the importance of YAP1 in the process of DN.
To further investigate the specific regulatory
mechanism of YAP1 in DN, a large body of studies was used to screen
the signaling pathways that may interact with YAP1. JMJD3, also
known as lysine demethylase 6B, is located on human chromosome 17.
Its abnormal expression is closely associated with the occurrence
and development of various types of cancer, and it is considered to
be a tumor evaluation index with a potentially great application
value (22,23). A previous study showed that JMJD3
significantly promoted IL-1β expression in the early stage of
sepsis (24). In addition, JMJD3
may regulate neointimal proliferation following vascular injury and
promote joint destruction in rheumatoid arthritis by promoting the
proliferation and migration of fibroblast-like synoviocytes
(25). As the upstream target of
JMJD3, NF-κB regulates inflammatory gene expression in vascular
endothelial cells (26). In
addition, cystitis promotes NF-κB pathway activation and JMJD3
expression, thereby inducing proliferation of human bladder smooth
muscle cells and ECM deposition (27). It is worth noting that YAP1 was
shown to inhibit the negative regulatory factor of NF-κB USP31 and
induce excessive tumor cell proliferation. The aberrant
stabilization of YAP1 has been shown to significantly enhance the
activity of the NF-κB pathway (28). Therefore, it was hypothesized that
YAP1 may activate the NF-κB/JMJD3 signaling pathway and promote
inflammation and ECM deposition in HBZY-1 cells treated with high
glucose. In the present study, the expression of NF-κB/JMJD3
signaling pathway-associated proteins was upregulated in HBZY-1
cells treated with high glucose, and inhibitors of NF-κB/JMJD3
signaling could significantly enhance the promoting effect of YAP1
on inflammation and ECM deposition of HBZY-1 cells treated with
high glucose.
YAP-1 is a well-characterized downstream
transcriptional co-activator of Hippo pathway that interacts with
various transcription factors and modulates their transcriptional
activities. However, the association between Hippo pathway and
YAP-1-induced inflammation and deposition of extracellular matrix
in diabetic nephropathy was not study. In addition, it was further
explored how YAP-1 could potentially activate NF-κB. The
aforementioned issues will be studied in further research.
In conclusion, YAP1 may aggravate high
glucose-induced GMC inflammation and ECM deposition by activating
the NF-κB/JMJD3 signaling pathway. The results of the present study
may indicate a new direction for the identification of therapeutic
targets for DN.
Acknowledgements
Not applicable.
Funding
This work was supported by the Fundamental Research Funds for
the Provincial Universities (grant no. 2019-KYYWF-0373).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YW and ZC designed the experiments and made
considerable contributions to the manuscript writing. YW and JX
performed the experiments and analyzed the data. ZC revised the
manuscript and guided the experiments. ZC and JX confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethical approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yuan CM, Nee R, Ceckowski KA, Knight KR
and Abbott KC: Diabetic nephropathy as the cause of end-stage
kidney disease reported on the medical evidence form CMS2728 at a
single center. Clin Kidney J. 10:257–262. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Duran-Salgado MB and Rubio-Guerra AF:
Diabetic nephropathy and inflammation. World J Diabetes. 5:393–398.
2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Tessari P: Nitric oxide in the normal
kidney and in patients with diabetic nephropathy. J Nephrol.
28:257–268. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wada J and Makino H: Inflammation and the
pathogenesis of diabetic nephropathy. Clin Sci (Lond). 124:139–152.
2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Meza Letelier CE, San Martín Ojeda CA,
Ruiz Provoste JJ and Frugone Zaror CJ: Pathophysiology of diabetic
nephropathy: A literature review. Medwave. 17(e6839)2017.PubMed/NCBI View Article : Google Scholar : (In Spanish).
|
|
6
|
Xu X, Xiao L, Xiao P, Yang S, Chen G, Liu
F, Kanwar YS and Sun L: A glimpse of matrix metalloproteinases in
diabetic nephropathy. Curr Med Chem. 21:3244–3260. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kwiatkowska E, Domanski L, Bober J,
Safranow K, Romanowski M, Pawlik A, Kwiatkowski S and Ciechanowski
K: Urinary metalloproteinases-9 and-2 and their inhibitors TIMP-1
and TIMP-2 are markers of early and long-term graft function after
renal transplantation. Kidney Blood Press Res. 41:288–297.
2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Mazanowska O, Żabińska M,
Kościelska-Kasprzak K, Kamińska D, Krajewska M, Banasik M,
Madziarska K, Zmonarski SC, Chudoba P, Biecek P, et al: Increased
plasma matrix metalloproteinase-2 (MMP-2), tissue inhibitor of
proteinase-1 (TIMP-1), TIMP-2, and urine MMP-2 concentrations
correlate with proteinuria in renal transplant recipients.
Transplant Proc. 46:2636–2639. 2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Hu K, Mars WM and Liu Y: Novel actions of
tissue-type plasminogen activator in chronic kidney disease. Front
Biosci. 13:5174–5186. 2008.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Li FJ, Surolia R, Li H, Wang Z, Liu G, Liu
RM, Mirov SB, Athar M, Thannickal VJ and Antony VB: Low-dose
cadmium exposure induces peribronchiolar fibrosis through
site-specific phosphorylation of vimentin. Am J Physiol Lung Cell
Mol Physiol. 313:L80–L91. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Grampa V, Delous M, Zaidan M, Odye G,
Thomas S, Elkhartoufi N, Filhol E, Niel O, Silbermann F, Lebreton
C, et al: Novel NEK8 mutations cause severe syndromic renal cystic
dysplasia through YAP dysregulation. PLoS Genet.
12(e1005894)2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Rinschen MM, Grahammer F, Hoppe AK, Kohli
P, Hagmann H, Kretz O, Bertsch S, Höhne M, Göbel H, Bartram MP, et
al: YAP-mediated mechanotransduction determines the podocyte's
response to damage. Sci Signal. 10(eaaf8165)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Chen Y, Zhao X, Sun J, Su W, Zhang L, Li
Y, Liu Y, Zhang L, Lu Y, Shan H and Liang H: YAP1/Twist promotes
fibroblast activation and lung fibrosis that conferred by miR-15a
loss in IPF. Cell Death Differ. 26:1832–1844. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wu Y, Shen L, Liang X, Li S, Ma L, Zheng
L, Li T, Yu H, Chan H, Chen C, et al: Helicobacter pylori-induced
YAP1 nuclear translocation promotes gastric carcinogenesis by
enhancing IL-1β expression. Cancer Med. 8:3965–3980.
2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang GL, Xia XL, Li XL, He FH and Li JL:
Identification and expression analysis of the MSP130-related-2 gene
from Hyriopsis cumingii. Genet Mol Res. 14:4903–4913.
2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Mulder S, Hamidi H, Kretzler M and Ju W:
An integrative systems biology approach for precision medicine in
diabetic kidney disease. Diabetes Obes Metab. 20 (Suppl 3):S6–S13.
2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Choi SY, Bae H, Jeong SH, Park I, Cho H,
Hong SP, Lee DH, Lee CK, Park JS, Suh SH, et al: YAP/TAZ direct
commitment and maturation of lymph node fibroblastic reticular
cells. Nat Commun. 11(519)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ma R, Ren JM, Li P, Zhou YJ, Zhou MK, Hu Z
and Xiao XY: Activated YAP causes renal damage of type 2 diabetic
nephropathy. Eur Rev Med Pharmacol Sci. 23:755–763. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Han N, Tian W, Yu N and Yu L: YAP1 is
required for the angiogenesis in retinal microvascular endothelial
cells via the inhibition of MALAT1-mediated miR-200b-3p in high
glucose-induced diabetic retinopathy. J Cell Physiol.
235:1309–1320. 2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Park MJ, Kim DI, Lim SK, Choi JH, Han HJ,
Yoon KC and Park SH: High glucose-induced O-GlcNAcylated
carbohydrate response element-binding protein (ChREBP) mediates
mesangial cell lipogenesis and fibrosis: The possible role in the
development of diabetic nephropathy. J Biol Chem. 289:13519–13530.
2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang J, Yang Q, Nie Y, Guo H, Zhang F,
Zhou X and Yin X: Tetrahydrobiopterin contributes to the
proliferation of mesangial cells and accumulation of extracellular
matrix in early-stage diabetic nephropathy. J Pharm Pharmacol.
69:182–190. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Sakaki H, Okada M, Kuramoto K, Takeda H,
Watarai H, Suzuki S, Seino S, Seino M, Ohta T, Nagase S, et al:
GSKJ4, a selective jumonji H3K27 demethylase inhibitor, effectively
targets ovarian cancer stem cells. Anticancer Res. 35:6607–6614.
2015.PubMed/NCBI
|
|
23
|
Tokunaga R, Sakamoto Y, Nakagawa S, Miyake
K, Izumi D, Kosumi K, Taki K, Higashi T, Imamura Y, Ishimoto T, et
al: The prognostic significance of histone lysine demethylase
JMJD3/KDM6B in colorectal cancer. Ann Surg Oncol. 23:678–685.
2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen Y, Liu Z, Pan T, Chen E, Mao E, Chen
Y, Tan R, Wang X, Tian R, Liu J and Qu H: JMJD3 is involved in
neutrophil membrane proteinase 3 overexpression during the
hyperinflammatory response in early sepsis. Int Immunopharmacol.
59:40–46. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Jia W, Wu W, Yang D, Xiao C, Su Z, Huang
Z, Li Z, Qin M, Huang M, Liu S, et al: Histone demethylase JMJD3
regulates fibroblast-like synoviocyte-mediated proliferation and
joint destruction in rheumatoid arthritis. FASEB J. 32:4031–4042.
2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yu S, Chen X, Xiu M, He F, Xing J, Min D
and Guo F: The regulation of Jmjd3 upon the expression of NF-κB
downstream inflammatory genes in LPS activated vascular endothelial
cells. Biochem Biophys Res Commun. 485:62–68. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lai J, Ge M, Shen S, Yang L, Jin T, Cao D,
Xu H, Zheng X, Qiu S, Wang K, et al: Activation of NFKB-JMJD3
signaling promotes bladder fibrosis via boosting bladder smooth
muscle cell proliferation and collagen accumulation. Biochim
Biophys Acta Mol Basis Dis. 1865:2403–2410. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ye S, Lawlor MA, Rivera-Reyes A, Egolf S,
Chor S, Pak K, Ciotti GE, Lee AC, Marino GE, Shah J, et al:
YAP1-mediated suppression of USP31 enhances NFκB activity to
promote sarcomagenesis. Cancer Res. 78:2705–2720. 2018.PubMed/NCBI View Article : Google Scholar
|