Introduction
Diabetes mellitus (DM) is a metabolic dysfunction
disease that is caused by insufficient insulin action or
ineffective insulin production. Insulin is a naturally occurring
hormone produced by the β cells of pancreatic islets, which helps
the body utilize glucose for energy (1). Abnormal insulin levels lead to
hyperglycemia, which causes acute or chronic complications in
patients with diabetes, such as vascular disease (2). The brain is an organ of the central
nervous system (CNS) that is most responsive to changes in oxygen
and metabolites in humans and animals. It maintains neuronal
function and control over other organs of the body. Because the
nervous system and blood circulation are associated, disturbances
of cerebral blood flow (CBF) lead to changes in neural function
(2). DM has been associated with
pathological changes in the CNS, which can lead to cognitive and
affective deficits as well as increased risk of vascular
complications in the brain (3).
The circle of Willis connects the anterior and
posterior cerebral circulations. The anterior circulation of the
brain derives from the bilateral internal carotid arteries and
branches of the common carotid arteries; the posterior circulation
derives from the bilateral vertebral arteries and branches of the
subclavian arteries, which combine to form the posterior cerebral
artery (4). The anterior cerebral
circulation supplies blood to the anterior portion of the brain. It
is supplied by the internal carotid artery (ICA) and its branches:
The anterior cerebral artery (ACA) and the middle cerebral artery
(MCA) (4). The ICA supplies
oxygenated blood to most midline portions of the frontal lobe and
superior medial parietal lobe. It is the most important blood
supply of the cerebral circulation from the circle of Willis
(5). The MCA receives 80% of the
carotid blood flow, whereas the ACA receives 20%. Each branch of
the anterior circulation supplies the cortex and the basal surface
of the brain, providing blood to deep structures, such as the
anterior hypothalamus, basal ganglia and internal capsule (6).
Diabetes mellitus increases the risk of
cardiovascular and cerebrovascular diseases (7). Furthermore, it has been demonstrated
to diminish angiogenesis in peripheral vascular beds (7). VEGFA and Ang-1 are highly effective
inducers of endothelial proliferation; they also promote
hyperplasia, tortuosity and a decreased lumen diameter. High blood
glucose conditions result in brain hypoxia-induced angiogenesis
through the upregulation of hypoxia-inducible genes, such as
hypoxia-inducible factor-1 (HIF-1). The vascular inner wall affects
blood pressure and, thus, the perfusion of nutrients and blood
cells that control the delivery of oxygen and immunological
surveillance. Several factors, including inflammatory cytokines
(TNF-α and IL-1β) influence endothelial cell luminal integrity as a
result in capillaries tube regression, a critical pathological
process regulating human disease. Investigating the mechanisms and
the factors that control, maintain and regulate lumen size and
diameter will increase our understanding of pathophysiological
processes. Lumen diameter is tightly regulated by several factors
(8). For example, VEGFA and its
receptors increase lumen formation and decrease lumen diameter
through the proliferation of myointimal cells and collagen
deposition (8,9). Ang-1 stabilizes the nascent vessel
(10).
An increased expression of VEGFA is caused by local
ischemic conditions and the activation of hypoxia-induced factor
(HIF)-1α (7). Brain angiogenesis
complicates several brain functions following the release of a
number of growth factors from the endothelial cells of the brain,
including VEGFA (11). VEGFA is a
protein with increase vascular permeability activity during
angiogenesis, which regulates the formation and maintenance of
blood vessel structures and is considered crucial for the
progression of several pathology and diseases, such as cancer
(11). VEGFA serves an important
role in mediating diabetic vasculopathy and microvascular
permeability (12). As a potent
vasodilator, VEGFA is regarded as a survival factor for endothelial
cells (12). Angiopoietins also
serve important roles in vascular development and angiogenesis.
Ang-1 is a member of the angiopoietin family. Ang-1 and its
receptor, Tie-2, are important for the regulation of angiogenesis.
Tie-2 receptors are predominantly expressed on endothelial cells
(13). Furthermore, the Tie2/Ang-1
signaling pathway serves a key role in the latter stages of
improvement of vascular dysfunction, remodeling and stabilizing
vessels, suppressing plasma leakage, inhibiting vascular
inflammation and preventing endothelial cell death (13). Ang-1 serves a crucial role in
mediating reciprocal interactions between endothelium, surrounding
matrix and mesenchyme; it is also involved in vascular maturation
and stability (10). Endothelial
budding is facilitated by vasodilation and the loosening of
endothelial cell contacts (14).
The pre-existing vessels will exhibit leakiness, which allows
extravascular plasma protein and extracellular matrix components to
migrate out from endothelial cells through the extracellular matrix
toward an angiogenic stimulus, such as VEGFA, that can promote
growing and survival of new blood vessels (14).
To the best of our knowledge, the effects of
gymnemic acid (GM) on VEGFA or Ang-1 have not yet been reported in
a rat model of diabetes-induced stroke.
Gymnema sylvestre (Asclepiadaceae; GS) is a
potent antidiabetic plant that is used in folk, ayurvedic and other
alternative medicines (15) to
treat of asthma, eye complaints such as cornea opacity,
inflammations, family planning by treating amenorrhea and
snakebites (15). GS has been
reported to exhibit antimicrobial, antihypercholesterolemic and
hepatoprotective activities (15).
GM is a mixture of oleanane-type triterpene saponins extracted from
the leaves of GS (16). GM is known
to suppress sweetness, whereby after chewing the leaves, sweetened
solutions with sucrose taste like water (17). A previous study demonstrated that GM
decreases blood glucose levels and increases plasma insulin levels
in diabetic rats (17). The
molecular structure of GM is similar to that of glucose; it can
bind to the glucose receptors on the taste buds of the tongue and
prevent its activation by sugar molecules from food and reducing
its sugary taste (18).
Additionally, the absorptive external layers of the intestine
exhibit receptors for GM molecules that prevent the absorption of
sugar molecules, lowering blood glucose levels (19). GM improves glucose and lipid
metabolism in type two DM model rats, enhances glucose uptake by
regulating the amelioration of endoplasmic reticulum stress and
insulin transduction in insulin resistant HepG2 cells (20), and inhibits GAPDH during glycerol
metabolism in rabbits (21).
The present study aimed to investigate the
restoration and improvement of GM on the blood vessels of the
anterior cerebral circulation that supply the majority of blood to
the brain, including arteries (ICA, MCA and ACA) and small vessels
(arterioles and capillary network) of the anterior cerebral
circulation, which provide crucial blood supply into the cerebral
circulation via the circle of Willis. The hypothalamus is an area
of the brain that is surrounded by branches of arteries that
originate near the stump of the circle of Willis (4). Therefore, this area of the brain was
selected for assessment in the present study. In addition, the
protein expression levels of VEGFA and Ang-1 in the hypothalamus
were analyzed to determine their association with affecting
angiogenesis or formation of new blood vessels in diabetic rat's
brain.
Materials and methods
Animals
All experimental protocols were reviewed and
approved by the Animal Ethics Committee of the Prince of Songkla
University (Songkhla, Thailand). A total of 80 male Wistar rats
(weight, 200-250 g; age, 8 weeks) were purchased from Nomura Siam
International Co., Ltd. Animals were housed under standard
laboratory conditions under a 12-h light/dark cycle, with good
lighting, moderate temperature (25+2˚C) with lights on at 7:00 a.m.
and humidity (50±10%) and adequate ventilation in a hygienic
environment. They were fed ad libitum with standard rat chow
containing protein, carbohydrate, fat, vitamins and minerals.
Induction and assessment of
diabetes
Experimental diabetic rats were intraperitoneally
injected with a single 60 mg/kg dose of streptozotocin (STZ;
Sigma-Aldrich; Merck KGaA) dissolved in 0.1 mol/l citrate buffer
(Sigma-Aldrich; Merck KGaA). Control rats were injected with 0.1
mol/l citrate buffer alone. Blood glucose levels were measured 3
days after STZ injection and persistent hyperglycemia was confirmed
1 week after STZ injection, using an Accu-Chek Active®
one-touch glucometer and test strips (Roche Diagnostics GmbH). Rats
with blood glucose levels >250 mg/dl were considered diabetic.
The rats were randomly divided into five groups (Fig. 1). (Normal control rats that received
a balanced standard diet (control group; n=10); 1B) gymnemic
control rats (CGM group; n=10) that received a balanced standard
diet supplemented with 400 mg/kg GM (Xi'an Guanyu Bio-Tech Co.)
(22) (purified >75% via
high-performance liquid chromatography analysis) in 0.5 ml 0.5%
Tween-80 solution (Sigma-Aldrich; Merck KGaA); 2) diabetic rats
that received a balanced standard diet and intraperitoneally
injected with 60 mg/kg STZ (STZ group; n=20 3) diabetic rats that
received a balanced standard diet supplemented with 400 mg/kg GM in
0.5 ml 0.5% Tween-80 solution (STZ + GM group; n=20); and 4)
diabetic rats that were treated with 4 mg/kg glibenclamide (GL)
(23) in 0.5 ml 0.5% Tween-80
solution (STZ + GL group; n=20). All animals were clinically
observed, such as weight loss, dehydration, cataracts, drowsiness,
and diabetic coma and weighed on a weekly basis. No deaths were
observed following supplementation with GM by being fed only one
time per day for a total of 8 weeks. Following animal sacrifice,
brain tissue was collected. Half of the rats in each group [n=5 (C
and CGM groups) n=10 (STZ, GM and GL groups)], tissue were used for
H&E staining and western blot analysis. The other half of rats
in each group were injected with resin for vascular corrosion
casting combined with scanning electron microscopy. The
experimental rats were euthanized by an excessive dose of sodium
pentobarbital (200 mg/kg; intraperitoneal injection). Death was
confirmed by observation of cardiac and respiratory arrest or fixed
and dilated pupils.
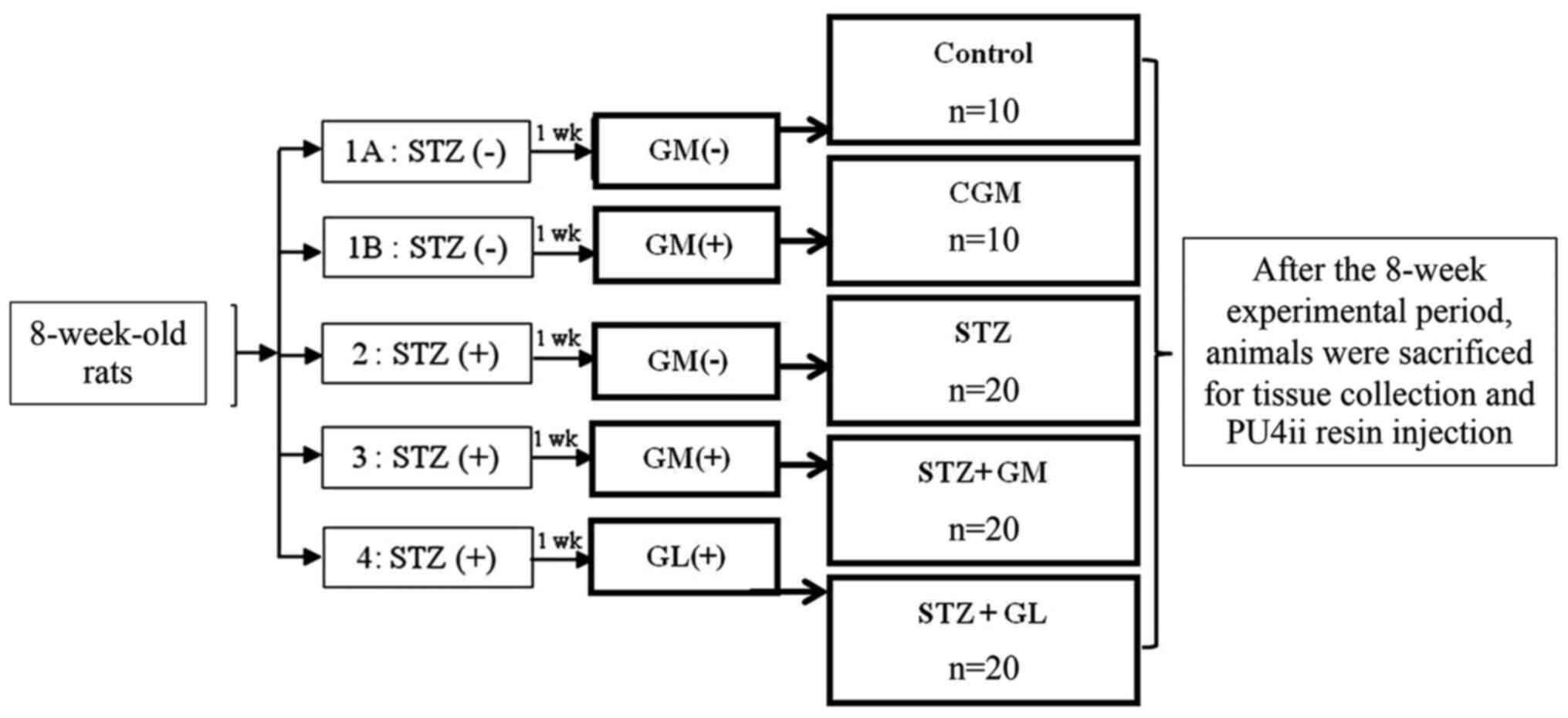 | Figure 1Experimental design. The rats were
randomly divided into five groups; 1A, Normal control rats that
received a balanced standard diet; 1B, gymnemic control rats (CGM)
that received a balanced standard diet supplemented with GM; 2,
diabetic rats that received a balanced standard diet and
intraperitoneally injected with 60 mg/kg STZ (STZ); 3, diabetic
rats that received a balanced standard diet supplemented with GM
(STZ + GM) and 4, diabetic rats that were treated with
glibenclamide (STZ + GL). After the 8-week experimental period,
animals were sacrificed for tissue collection and PU4ii resin
injection. Wk, week; GM, gymnemic acid; CGM, gymnemic control; STZ,
streptozotocin; GL, glibenclamide. |
Histological preparation for Masson's
trichrome staining
To analyze any histological changes and measure the
lumen diameter of the ICA, MCA and ACA, brain tissue from all
groups was dissected and immediately fixed in 10% formalin for 6 h
at room temperature. Tissue sections were dehydrated in a
descending ethanol series (70, 80, 90, 95 and 100%), with twice in
succession, each lasting 1 h. Prior to paraffin wax in filtration
and paraffin embedding, samples were washed three times with
clearing agent, xylene, at room temperature, with each wash lasting
30 min. Paraffin-embedded tissue samples were then cut into
5-µm-thick sections and stained at room temperature with Masson's
trichrome (Trichrome Stain (Masson) kit, HT15, Sigma-Aldrich; Merck
KGaA). This staining technique is used for the detection of
collagen fibers (24) in the
arterial wall of anterior circulation of brain. The brain tissue
slides were immersed in Bouin's solution at 56˚C for 15 min after
deparaffinization and rehydration. The brain sections were stained
for 5 min in Weigert's hematoxylin at room temperature and then
washed with tap water. The slides were stained in Biebrich
scarlet-acid fuchsin for 5 min, rinsed in distilled water,
incubated in phosphotungstic-phosphomolybdic acid for 5 min, dyed
with aniline blue for 5 min and fixed in 1% acetic acid for 2 min
at room temperature. After rinsing in distilled water, the slides
were dehydrated and mounted. The cross-sectional diameter of the
arterial lumen was measured from the outermost edge of the lumen
from left to right and top to bottom. The average value of these
measurements was considered the lumen diameter. All sections were
observed and captured under an Olympus BX50 light microscope using
an Olympus DP73 camera (Olympus Corporation; magnification, x600).
Lumen diameter of the arteries was measured using cellSens software
(v1.16, Olympus Corporation).
Western blot analysis of VEGFA and
Ang-1
The brain tissue lysates of control, CGM, STZ, STZ +
GM and STZ + GL rats at 8 weeks were prepared using ice-cold RIPA
buffer (Sigma-Aldrich; Merck KGaA) supplemented with 1X protease
inhibitor cocktail (MilliporeSigma). Homogenates were centrifuged
at 14,000 x g for 30 min at 4˚C to collect the supernatants. The
protein concentration of the supernatant was subsequently
determined using a BCA protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). Total protein (10 µg) was diluted (1:2) in
treatment buffer (0.125 M Tris-Cl, 4% SDS, 20% glycerol, 10%
2-mercaptoethanol and 0.2% bromophenol blue) and heated for 5 min
at 95˚C. Protein samples were subjected to 12% SDS-PAGE (100 V,
0.35 A and 300 W for 90 min) and subsequently transferred onto
nitrocellulose membranes (GE Healthcare). Membranes were then
blocked with 5% non-fat dry milk in 0.1% Tris-buffered saline with
Tween®-20 (TBS-T) for 1 h at room temperature. The
membranes were incubated with primary antibodies against VEGFA
(1:1,000; rabbit polyclonal antibody; cat. no. ab9570; Abcam) Ang-1
(1:1,000; rabbit polyclonal antibody; cat. no. PA1-32150; Thermo
Fisher Scientific, Inc.) and β-actin (1:3,000; rabbit polyclonal
antibody; cat. no. ab8227; Abcam) at 4˚C for 24 h. Membranes were
washed three times with TBS-T, followed by incubation for 2 h at
room temperature with goat anti-rabbit horseradish
peroxidase-conjugated immunoglobulin G secondary antibodies
(1:5,000; ab6721; Abcam). Protein bands were visualized using an
enhanced chemiluminescence (ECL) (Pierce™ ECL Western Blotting
Substrate; Thermo Fisher Scientific, Inc.) detection system with
Amersham Hyperfilm™ ECL (GE Healthcare). Resulting bands were
subjected to densitometric analysis using Scion Image 4.0 software
(Scion Corporation).
Vascular corrosion casting
technique
Vascular corrosion casting was performed via an
intravascular injection of 0.5 ml heparin (5,000 IU/ml; Leo Pharma
UK/IE), which was immediately injected into the left ventricle to
prevent blood clotting. The right atrium functioned as the outlet
for blood and injected fluid. A second injection of 400-500 ml 0.9%
normal saline solution (General Hospital Products Public Co., Ltd.)
was administered to remove the blood from the vascular bed.
Subsequently, PU4ii polyurethane-based casting resin (vasQtec) was
immediately injected into the cannula through the ascending aorta
until reflux from the venous vessels became evident. Each rat was
left at room temperature to allow the casting medium to settle for
2 h. The brain was subsequently excised and immersed in water at
80˚C to complete the hardening process. Thereafter, the brain
underwent a corrosive process by adding 10% KOH solution (Vidhyasom
Co., Ltd.) at room temperature for 1 month. Tissues were removed
and washed three times with distilled water. The brain vascular
cast was then dissected and were assessed using a stereomicroscope
(OLYMPUS SZ2 series) to obtain small specimens and air dried for 2
weeks. The vascular cast of the brain was placed on a metal stub
with double side adhesive tape, sprayed with carbon paint and
coated with gold on a sputtering apparatus prior to preparation for
scanning electron microscopy observation (JEOL JSM-5400; JEOL,
Ltd.) at 10-15 KV. The diameter of brain blood vessels was measured
using SemAfore 5.2 software (JEOL, Ltd.).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8.0 (GraphPad Software, Inc.). One-way ANOVA followed by
Bonferroni post-hoc analysis was used. Each experiment was repeated
at least twice and data are presented as the mean ± SEM. P<0.05
was considered to indicate a statistically significant
difference.
Results
Effect of GM on blood glucose
levels
The effect of GM on blood glucose levels in diabetic
rats is presented in Table I. The
blood glucose levels in the control, CGM, STZ, STZ + GM and STZ +
GL groups were compared at 8 weeks. The results demonstrated that
blood glucose levels were significantly elevated in the STZ, STZ +
GM and STZ + GL groups compared with the control rats (P<0.001).
Furthermore, blood glucose levels of STZ + GM rats and STZ + GL
rats significantly decreased from week 3 to week 8 compared with
STZ rats (P<0.01 and P<0.001).
 | Table IComparison of blood glucose levels in
different groups. |
Table I
Comparison of blood glucose levels in
different groups.
| Week | Control
(mg/dl) | CGM (mg/dl) | STZ (mg/dl) | STZ+GM (mg/dl) | STZ+GL (mg/dl) |
|---|
| 1 | 91.8±3.9 | 104.3±4.0 |
255.7±4.8a |
368.3±14.2a,c |
207.3±19.0a |
| 2 | 99.7±4.9 | 106.8±10.5 |
348.3±31.9a |
395.5±24.8a,b |
199.3±35.9b |
| 3 | 104.2±2.6 | 109.7±2.8 |
541.2±22.6a |
318.8±39.4a,b |
166.5±29.6b |
| 4 | 96.2±2.6 | 106.0±2.2 |
383.3±53.3a |
247.0±40.3a,b |
254.0±19.3c |
| 5 | 103.0±2.1 | 96.3±2.5 |
457.2±27.0a |
230.3±44.3a,b |
190.7±14.3b |
| 6 | 102.5±2.3 | 105.2±1.8 |
418.5±13.1a |
203.3±44.9a,b |
177.7±12.2b |
| 7 | 96.7±2.3 | 110.3±2.6 |
439.0±16.8a |
273.2±44.5a,b |
121.2±10.4b |
| 8 | 89.2±3.7 | 92.7±2.4 |
413.2±11.5a |
211.0±53.0a,b |
106.2±9.7b |
Effect of GM on body weight
The effect of GM on the body weight of diabetic rats
was evaluated in Table II. The
results demonstrated that body weight was significantly lower in
the STZ group compared with the control group (P<0.001). After
supplementation of GM, body weight was significantly increased
every week in the STZ + GM group compared with the STZ group
(P<0.01 and P<0.001).
| Table IIComparison of body weight in
different groups. |
Table II
Comparison of body weight in
different groups.
| Week | Control (g) | CGM (g) | STZ (g) | STZ+GM (g) | STZ+GR (g) |
|---|
| 1 | 310.3±8.4 | 322.2±6.7 |
213.3±15.5a |
284.4±14.3c |
229.7±10.1a |
| 2 | 327.2±9.2 | 336.7±5.7 |
212.2±19.5a |
294.4±35.6c |
233.3±10.9a |
| 3 | 340.7±8.4 | 356.7±6.3 |
227.6±20.2a |
311.3±16.9c |
329.4±23.3c |
| 4 | 351.4±8.5 | 369.2±7.3 |
230.0±20.1a |
317.5±18.7c |
267.8±12.8a |
| 5 | 364.1±8.1 | 386.3±7.5 |
239.6±20.9a |
318.0±21.1d |
299.4±14.3c |
| 6 | 382.9±9.2 | 352.5±6.9 |
245.6±21.9b |
346.9±19.2d |
286.1±14.6b |
| 7 | 402.1±9.2 | 418.3±3.8 |
286.1±28.7b |
361.3±19.49c |
299.4±16.7b |
| 8 | 409.3±9.6 | 432.2±7.7 |
255.4±24.6a |
367.2±18.7d |
303.3±17.2b |
Masson's trichome histological
observation of arteries in the anterior cerebral circulation
Rat brains were histologically analyzed for ICA, MCA
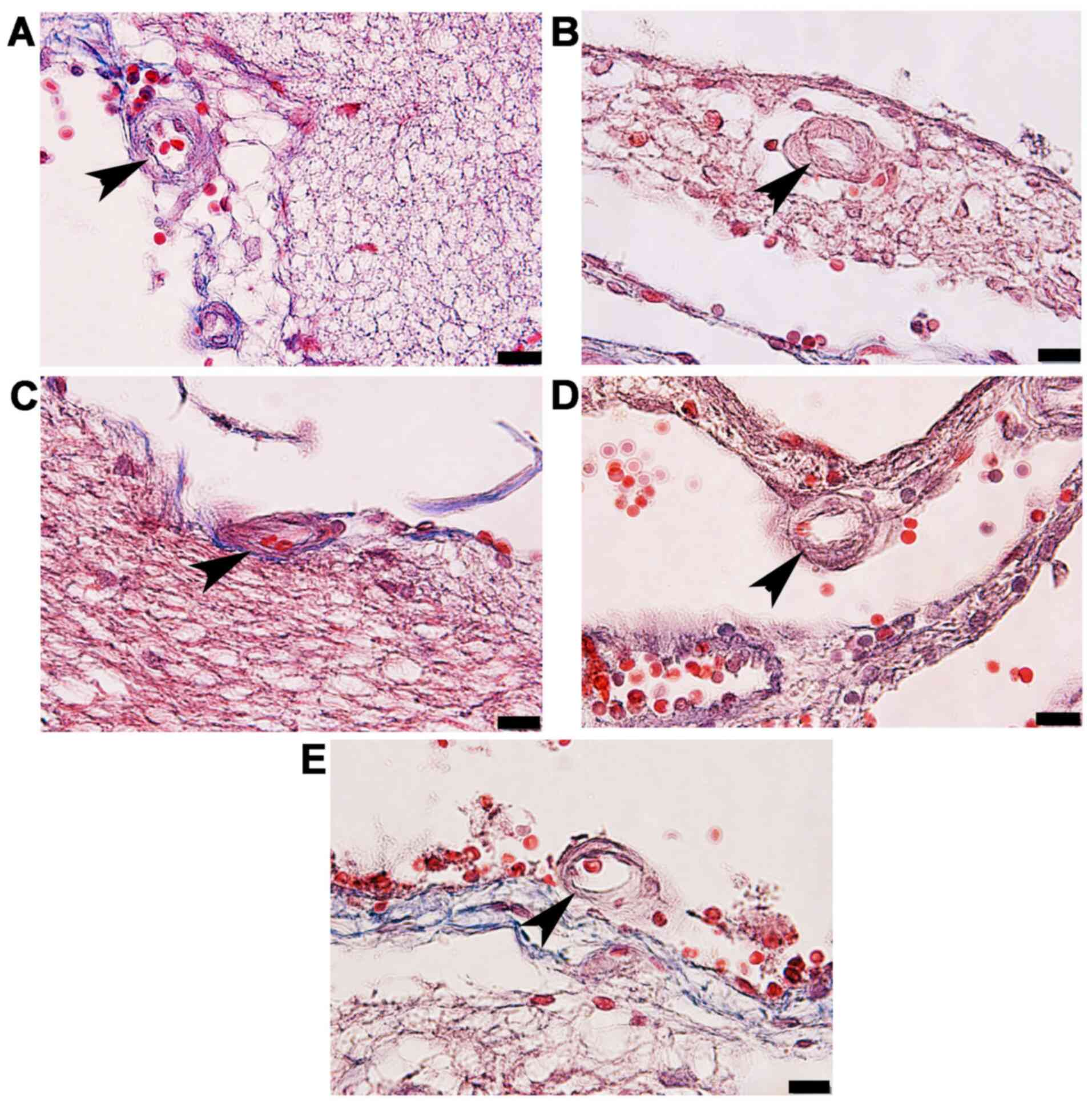
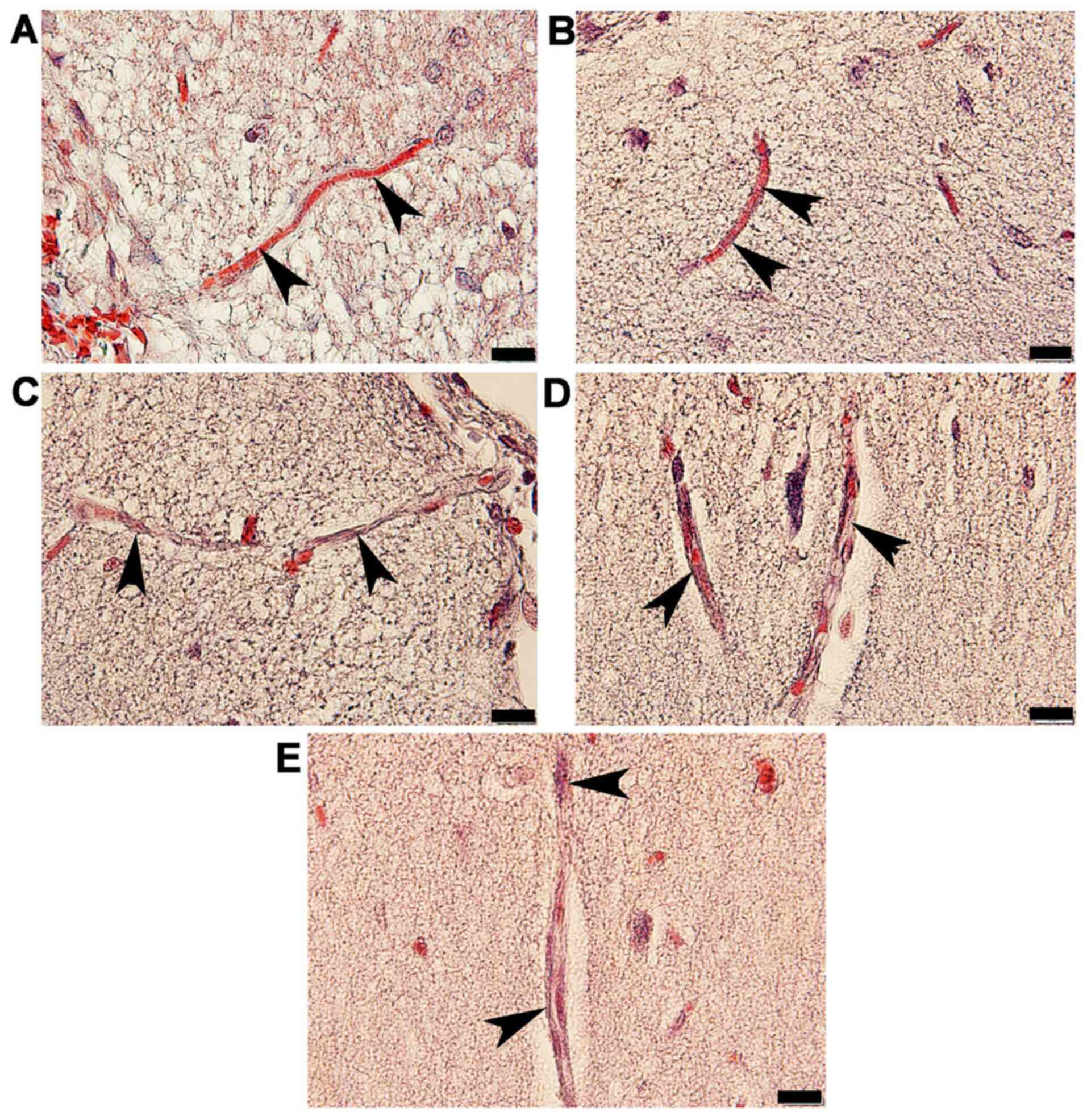
and ACA wall thickness, as well as arterial stenosis (Fig. 2). The results demonstrated that in
the STZ group, collagen accumulation (stained in blue) was present
in the in tunica media and tunica adventitia of the arterial walls,
and a markedly smaller lumen diameter was also demonstrated in the
three types of arteries. The arterioles (Fig. 3C) and capillaries (Fig. 4C) exhibited increased collagen
thickness in the vessel wall with indications of collagen fibers
(blue staining). Decreased collagen fiber accumulation in STZ + GM
(Figs. 3D and 4D) and STZ + GL rats (Figs. 3E and 4E was also revealed. The wall thickness of
ICA, MCA and ACA arteries were measured and presented in Table III. The wall thickness of each type
of artery in the STZ group was significantly increased (P<0.001)
when compared with control and CGM rats. By contrast, they were
significantly reduced in STZ + GM and STZ + GL rats (P<0.001 and
P<0.01, respectively) compared with the STZ group. The lumen
diameter of the ICA, MCA and ACA was significantly decreased in the
STZ group compared with the control and CGM groups (Fig. 5). Conversely, the lumen diameters
were increased in STZ + GM and STZ + GL rats compared with the STZ
group (Fig. 5).
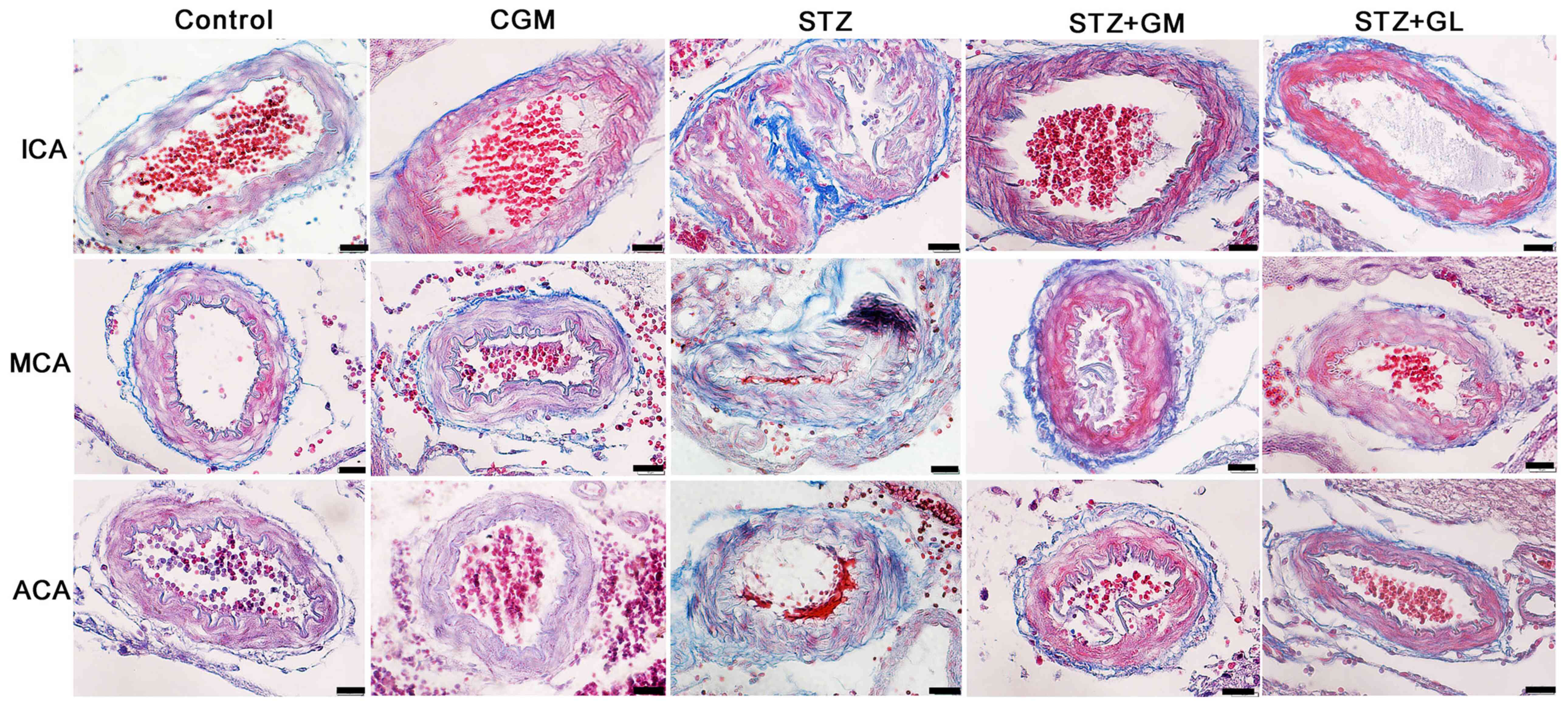 | Figure 2Photomicrographs demonstrating the
histological structure of the anterior arteries included in the
circle of Willis. Samples obtained from rats in the C, CGM, STZ,
STZ + GM and STZ + GL groups were subjected to Masson's trichrome
staining (magnification, x600). ACA, anterior cerebral artery; C,
control; CGM, gymnemic control; GL, glibenclamide; GM, gymnemic
acid; ICA, internal carotid artery; MCA, middle cerebral artery;
STZ, streptozotocin. |
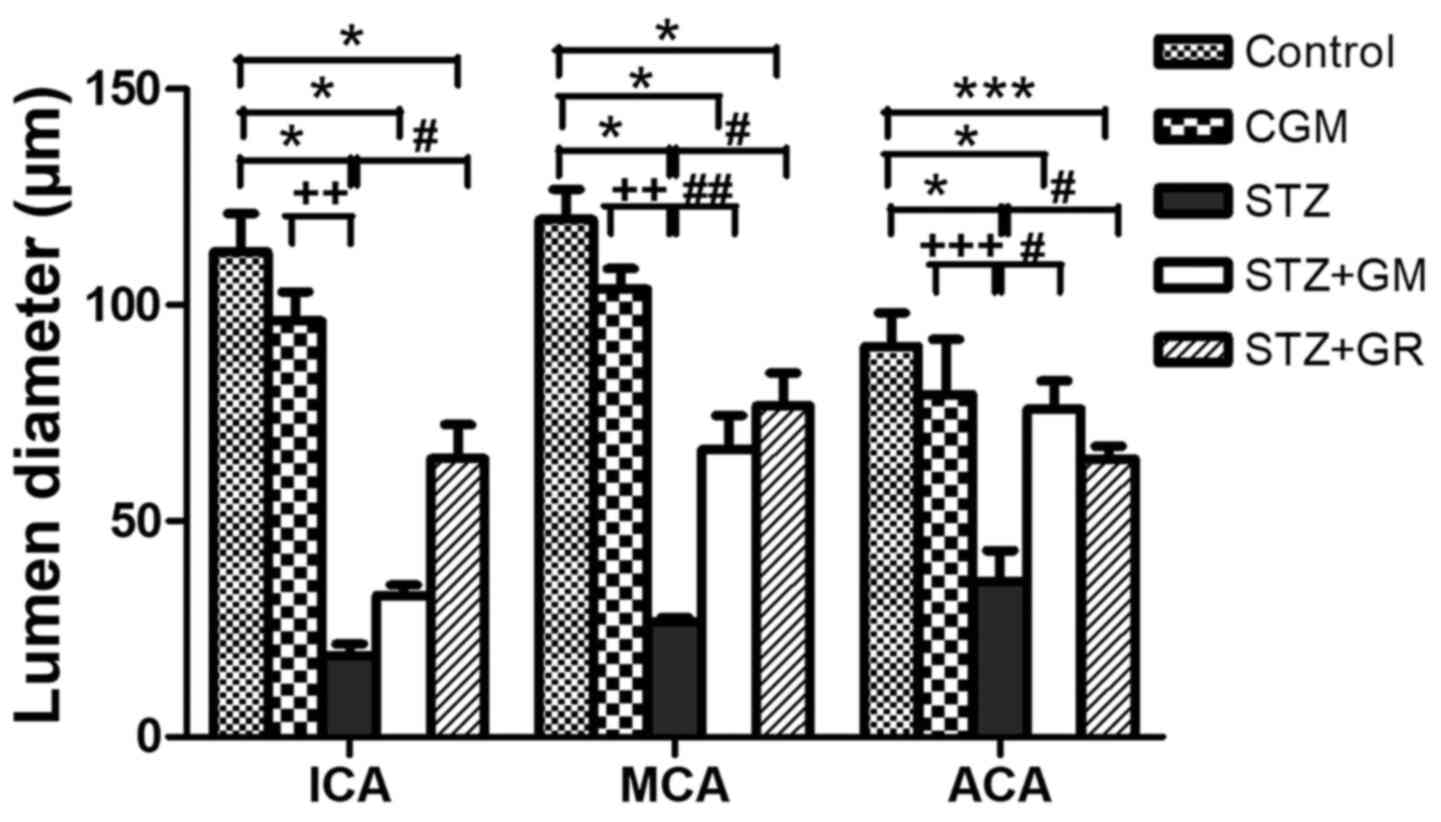 | Figure 5Average lumen diameters of the
anterior blood vessels included in the circle of Willis. Tissues
collected from the C, CGM, STZ, STZ + GM and STZ + GL groups were
subjected to Masson's trichrome staining following 8 weeks of
treatment. *P<0.001 and ***P<0.01 vs.
the control group, #P<0.001 and
##P<0.01 vs. STZ; ++P<0.01 and
+++P<0.05 vs. CGM. ACA, anterior cerebral artery; C,
control; CGM, gymnemic control; GL, glibenclamide; GM, gymnemic
acid; ICA, internal carotid artery; MCA, middle cerebral artery;
STZ, streptozotocin. |
Vascular corrosion cast
examination
Vascular corrosion casts of the anterior circulation
of the circle of Willis in control, CGM, STZ, STZ + GM and STZ + GL
rats were assessed using a stereomicroscope at low magnification
(Fig. 6A). The results demonstrated
that the size of the brain vascular cast in the STZ group is
smaller and exhibited a greater dropout of vessel density (Fig. 6A) compared with the other groups.
The cerebral arterial circle of Willis in the control group
exhibited anastomosing branches at the ventral view of the brain in
the low magnification SEM scan (Fig.
6B). It formed a polygon surrounding the optic chiasm, optic
tracts, pituitary stalk and basal hypothalamus by two ICAs, and the
rostral branches (referred to as the posterior cerebral of basilar
artery). The anterior and posterior circulation of the polygon was
completed by the anastomotic branches (the anterior and posterior
communicating arteries) (4).
According to the anterior circulation of circle of Willis, the
ACAs, which are branches of the two internal carotid arteries, run
medially and rostrally downward in front of the optic chiasm. The
anterior part of the hypothalamus is supplied by the branches of
the anterior cerebral and anterior communicating arteries (25). The MCAs are extensive branches of
ICAs. They continue into the lateral sulcus where it then branches
and projects to several sections of the cerebral hemisphere. The
relative diameter of arteries from the anterior circulation of the
circle of Willis among the control, CGM, STZ, STZ + GM and STZ + GL
groups at 8 weeks were demonstrated ventrally within the brain.
Marked shrinkage of the ICA, ACA and MCA was exhibited in the STZ
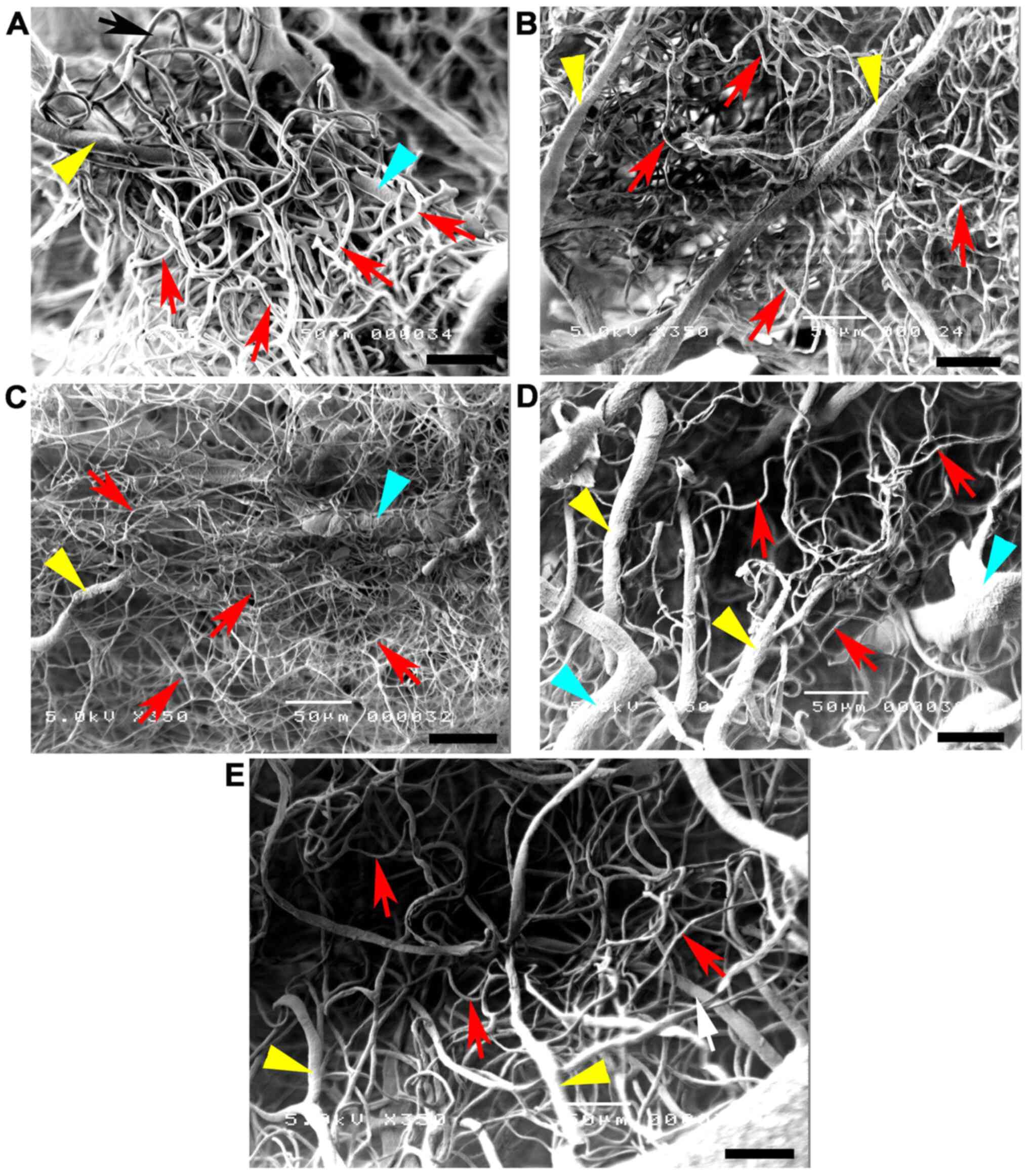
group compared with the other groups (Fig. 7C). At high magnification, the
vascular cast of the small blood vessels in the control and CGM
groups exhibited the typical pattern of arterioles and capillaries
in the hypothalamus and were surrounded by branches of the circle
of Willis (Fig. 8A and B). Conversely, the injured blood vessels
presented with smaller diameters, stenosis and shrinkage of the
arterioles and capillaries in the STZ group (Fig. 8C).
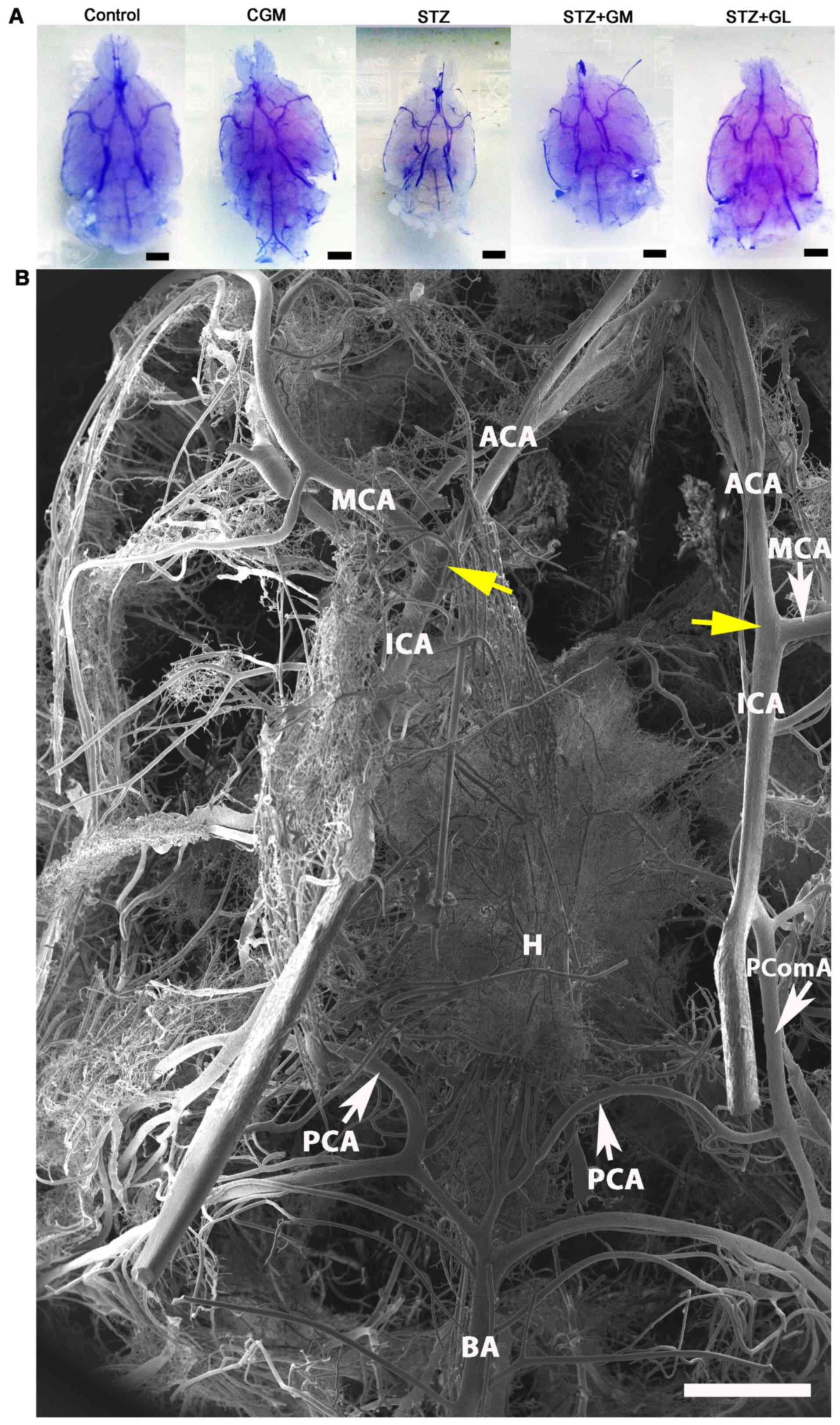 | Figure 6Vascular corrosion casts of the brain
of Wistar rats. (A) Stereomicrographs of the ventral views of the
brain vascular corrosion cast in control, CGM, STZ, STZ + GM and
STZ + GL rats showing the branches from circle of Willis (scale
bar, 1 mm). (B) Low magnification scanning electron micrograph of
the circle of Willis in a representative control group rat. The
image presents the ventral view of the brain. The circle of Willis
is a circulatory anastomosis (yellow arrows) that supplies blood to
the brain. A ring is formed surrounding the basal hypothalamus.
ACA, anterior cerebral artery; BA, basilar artery; CGM, gymnemic
control; GL, glibenclamide; GM, gymnemic acid; H, hypothalamus;
ICA, internal carotid artery; MCA, middle cerebral artery; PCA,
posterior cerebral artery; PComA, posterior communicating artery;
STZ, streptozotocin (scale bar, 1 mm). |
 | Figure 7High magnifications of vascular
corrosion casts of the anterior circulatory anastomosis of the
brain. Scanning electron micrographs of the ventral views of the
ICA, ACA and MCA from anterior cerebral circulation of circle of
Willis in (A) control, (B) gymnemic control, (C) STZ, (D) STZ + GM
and (E) STZ + GL rats; scale bars, 200 µm. The STZ group
demonstrated a marked shrinkage of the ICA, ACA and MCA. Signs of
vessel restoration and improvement were exhibited by the
increasingly healthy and nourished blood vessels of the STZ + GM
and STZ + GL groups. ACA, anterior cerebral artery; GL,
glibenclamide; GM, gymnemic acid; ICA, internal carotid artery;
MCA, middle cerebral artery; STZ, streptozotocin. |
Signs of vessel restoration were demonstrated by
increased healthy and nourished ICA, ACA and MCA in the STZ + GM
and STZ + GL groups (Fig. 7D and
E, respectively). ACA and MCA
diameters were significantly decreased in the STZ group compared
with the control (P<0.001) and CGM (P<0.01) groups; however,
no significant differences were observed between the STZ + GM and
STZ + GL groups (Fig. 9).
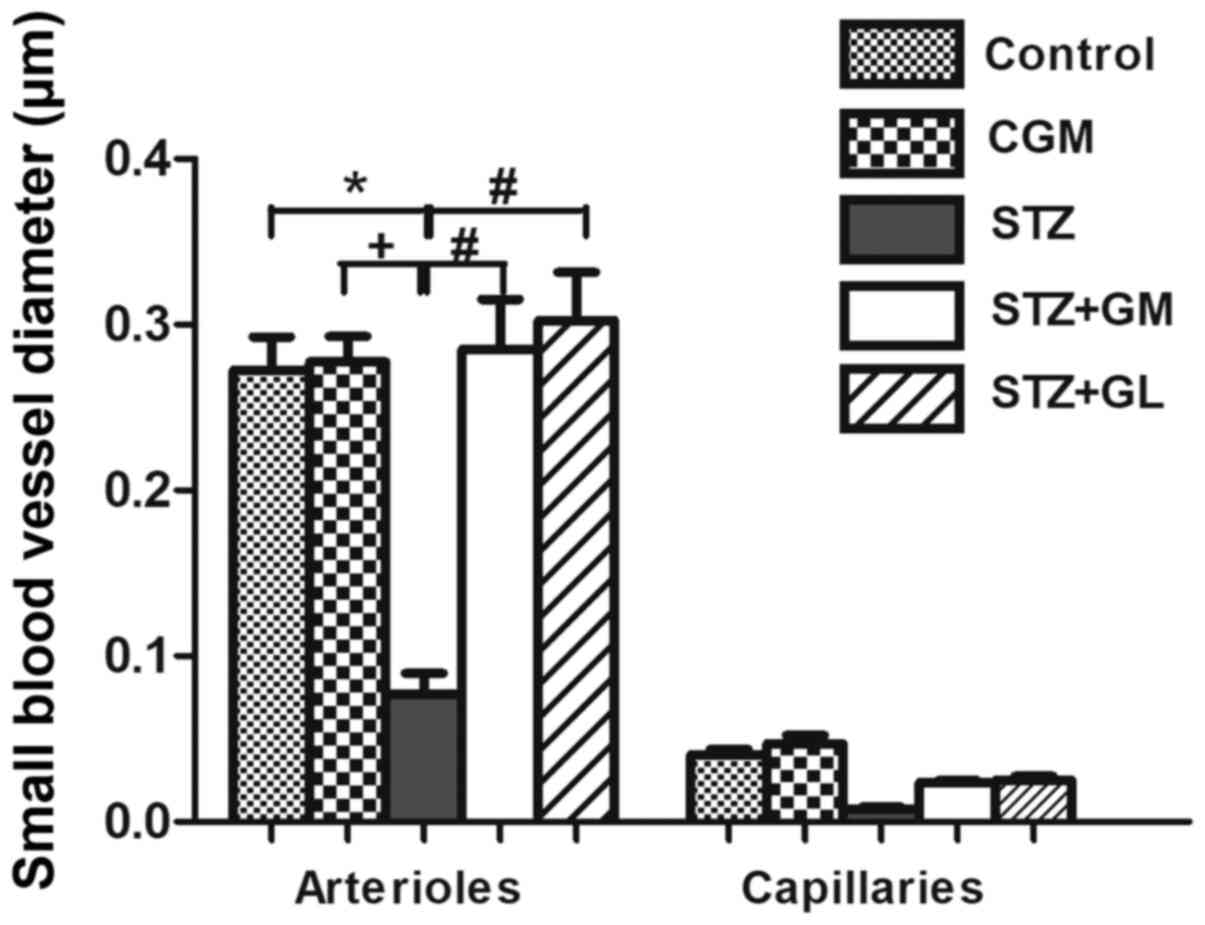
Furthermore, the STZ arterioles from the circle of Willis in the
hypothalamus exhibited decreased diameter sizes compared with all
other experimental groups (Fig.
10). The same effect was demonstrated in capillaries, although
it was not significant. The diameter of arterioles in the STZ group
were significantly decreased (P<0.001) when compared with the
control and CGM groups (Fig. 10).
Following supplementation with GM and GL, the well-organized
architecture of the arteries was restored in the STZ + GM (Fig. 8D) and STZ + GL group (Fig. 8E). Signs of vessel restoration and
improvement were exhibited by increased diameters, as well as
healthy and nourished arterioles and capillaries in the
hypothalamus of the STZ + GM and STZ + GL groups. Additionally, the
diameter of arterioles was significantly increased in STZ + GM and
STZ + GM groups compared with STZ rats (P<0.001).
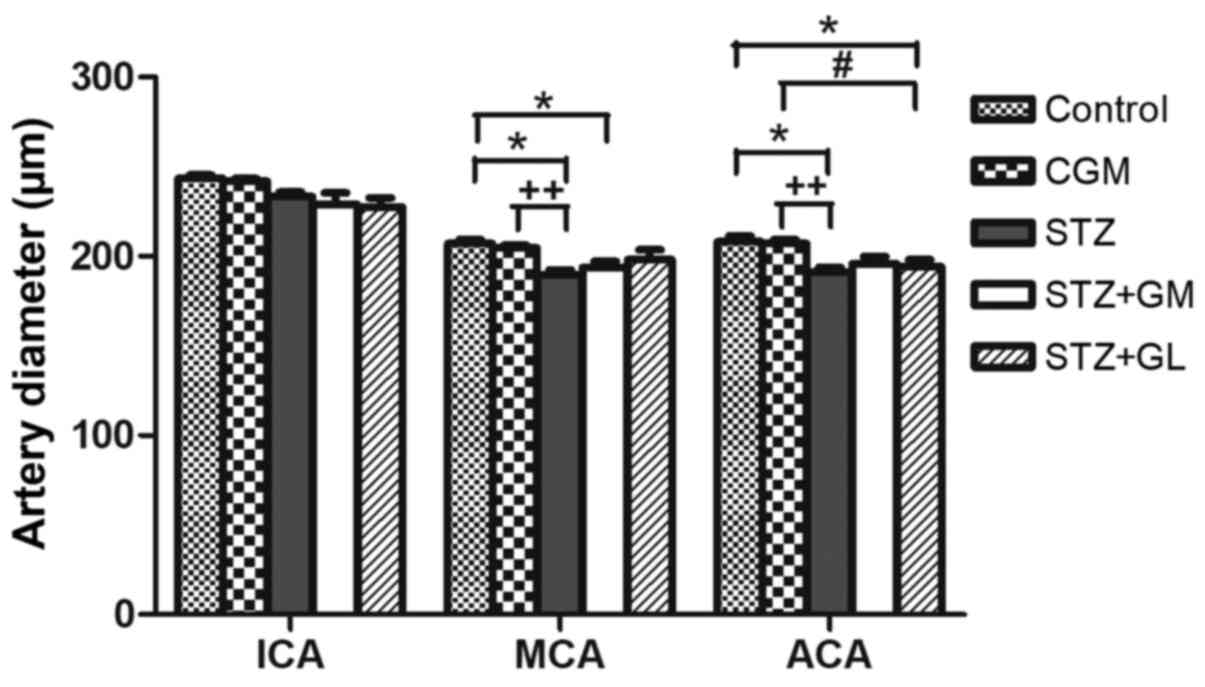 | Figure 9Average diameter of arteries from the
anterior circulation of the circle of Willis in control, CGM, STZ,
STZ + GM and STZ + GL rats at 8 weeks. Data are presented as the
mean ± SEM. *P<0.001 vs. control;
#P<0.001 vs. STZ; ++P<0.01 vs. CGM.
ACA, anterior cerebral artery; CGM, gymnemic control; GL,
glibenclamide; GM, gymnemic acid; ICA, internal carotid artery;
MCA, middle cerebral artery; STZ, streptozotocin. |
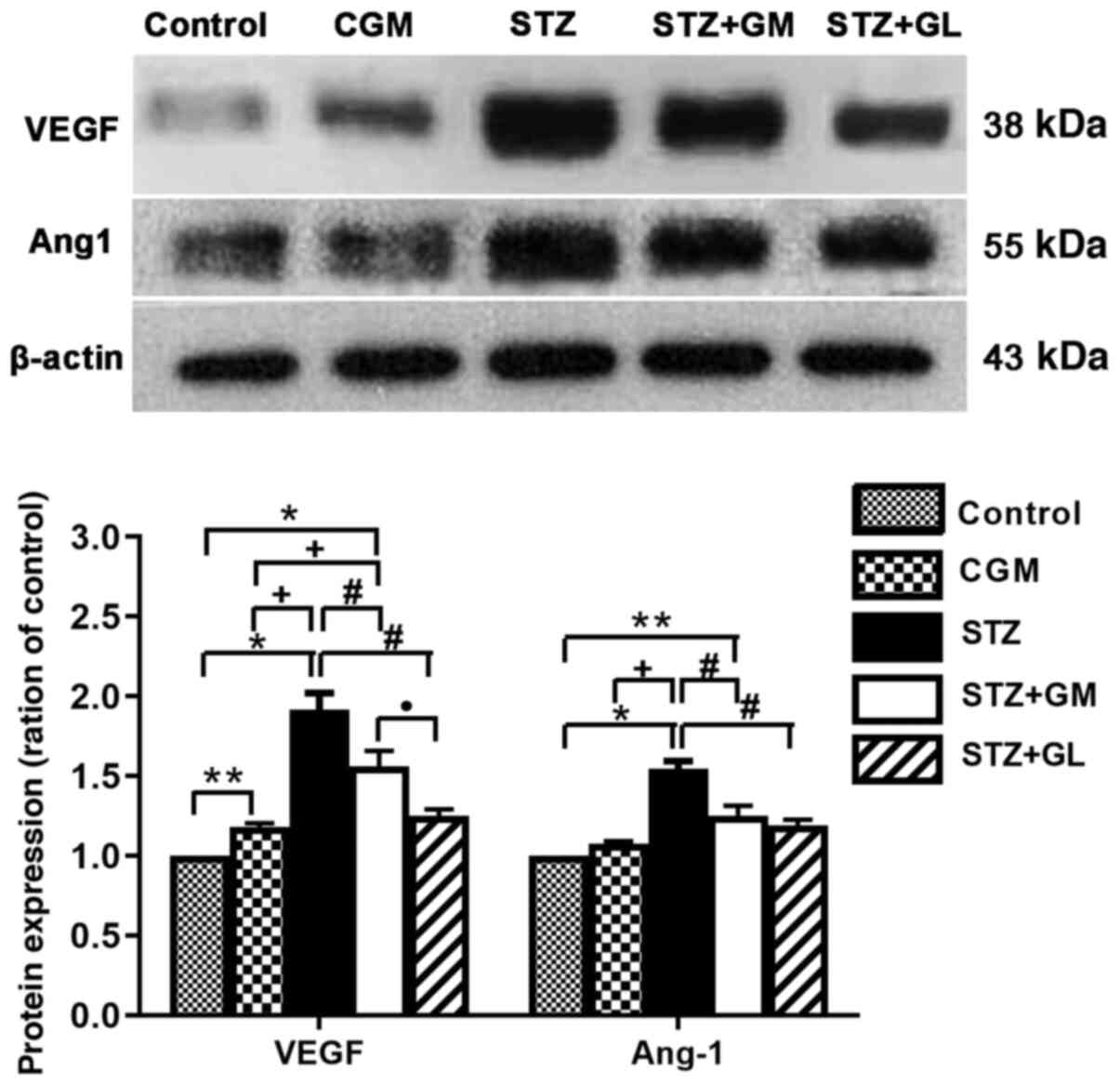
Western blot analysis
The western blotting results demonstrated that the
protein expression levels of VEGFA and Ang-1 (Fig. 11A) were significantly increased in
STZ rats (P<0.001) compared with the control and CGM rats,
whereas VEGFA and Ang-1 expression levels were decreased in STZ +
GM and STZ + GL groups compared with STZ rats. Furthermore, VEGFA
and Ang-1 protein expression levels were significantly increased in
STZ + GM rats compared with the control and CGM (P<0.0001 and
P<0.001). However, the protein expression levels of VEGFA and
Ang-1 were significantly decreased in STZ + GM rats compared with
STZ rats (P<0.001; Fig.
11).
Discussion
In the present study, the ability of GM to attenuate
the diabetes-induced morphological and histopathological
alterations of blood vessels in the anterior cerebral circulation
of rats was examined. The results demonstrated significantly
decreased blood glucose levels in the STZ rats treated with 400
mg/kg GM for a total period of 8 weeks compared with untreated
diabetic groups. This result is consistent with previous studies,
which have indicated that GS extracts may possess antidiabetic
properties (16,23). The antidiabetic and antioxidative
properties may be due to a number of bioactive compounds present in
GS extracts, including oleanane-type triterpenoid saponins (known
as GM), alkaloids, acidic glycosides and anthraquinones and their
derivatives (26).
In the present study, histological analysis by
Masson's trichrome staining of ICA, MCA and ACA from the brain
tissues of STZ rats demonstrated wall thickening and increased
collagen fiber deposition in the tunica media and tunica adventitia
of arteries. Furthermore, increased collagen thickness was
demonstrated in the walls of arterioles and capillaries. Moreover,
STZ rats exhibited lumen narrowing in each type of artery. Collagen
deposition can lead to the stenosis of arteries, whereas immoderate
collagen breakdown combined with insufficient synthesis weakens
plaques, thereby making them rupture (27). Newly synthesized collagen may work
as a substrate leading to luminal narrowing (27). Arterial stenosis is the narrowing of
large arteries that transfer blood to the body, including the head,
face and brain (28). The
destruction of the vascular wall results in vascular complications
in diabetes (28). Early structural
impairment of the common carotid artery is often associated with
type I diabetes and is considered an early sign of cerebral
atherosclerosis, which is also reflected by the increased wall
thickness of the tunica intima and tunica media (29). These histopathological changes
result in blood vessels becoming inflexible, thus becoming unable
to react to exogenous or endogenous stimuli to effectively regulate
blood flow (2). Atherosclerosis is
a regressive blood vessel disorder that generally affects medium to
large-sized arteries. In the brain, the vessels of the circle of
Willis are often affected byy atherosclerosis (30).
Vascular corrosion casting demonstrated significant
destruction of arteries (ICA, MCA and ACA) and small vessels
(arterioles and capillaries) in the brains of the STZ group. Vessel
diameter is a key determinant of stroke extension; under normal
physiological conditions, vessel diameter is regulated by several
pathways, such as intracellular signaling, cytokine and nervous
control that ensure adequate blood flow (30). Following cerebral artery occlusion,
the consecutive drop in local CBF is offset by recruitment of the
collateral artery and vasodilation, thus limiting the extension of
the lesion; this effect is mediated by nitric oxide (NO) (31). VEGF mediates the endothelial nitric
oxide synthase (eNOS) expression of NO following cerebral ischemia
(31). Physical activity improves
long-term stroke outcome through eNOS-dependent mechanisms that
increase angiogenesis and CBF (32). In the vascular system, VEGF and
Ang-1 (with its receptor Tie2), are essential angiogenic factors
that control angiogenesis between small and large blood vessels
(33). VEGF serves a crucial role
in angiogenesis and neuronal regeneration following ischemic stroke
(34). VEGF is a master regulator
of angiogenesis that has been shown to directly regulate lumen
size. Changes in lumen diameter occur in response to blood pressure
and blood flow, and subsequently control oxygen delivery and
immunological surveillance. VEGF and Ang-1 are highly effective at
inducing endothelial proliferation and promoting hyperplasia,
tortuosity and decreased lumen diameter. Therefore, increased
expression of these two proteins results in the narrowing of the
lumen diameter (35).
In chronic and weak cerebral hypoxia demonstrated in
patients with large cerebral artery stenosis, VEGF expression may
increase prior to the onset of a stroke and may function as an
angiogenic molecule (36). However,
VEGF has been reported to also cause leakage of the blood-brain
barrier, inflammation and brain edema (37). Ang-1 modulates the maturation and
stabilization of newly formed vessels, including adherence and
maintaining the survival of vascular epithelial cells (38). Ang-1 decreases permeability of the
endothelial cell and increases vascular stabilization by recruiting
pericytes and smooth muscle cells to the growing blood vessels.
Together, VEGF and Ang-1 enhance angiogenesis and are more
effective in decreasing blood-brain barrier leakage compared with
VEGF alone (39).
In the present study, western blotting demonstrated
that VEGF and Ang-1 protein expression levels were increased in the
STZ group compared with the control group. This angiogenic factors
are increased in cerebral microvessels of diabetic animals is
consistent with previous results on other diabetic animal models,
such as high-fat-diet, Goto-Kakizaki rats and db/db mice (40). These changes were associated with
increased VEGF signaling (41). An
increased understanding of the regulation of cerebral
neovascularization in diabetes is required to identify preventive
and therapeutic strategies against cerebral complications (42). Hypoperfusion of the brain results in
hypoxia-induced angiogenesis through the upregulated expression of
hypoxia-inducible genes, such as VEGF and Ang-1(43). Arterial stenosis or atherosclerotic
changes of the vessels caused by hypoxia are considered to serve an
important role in the pathogenesis of brain ischemia (44). During inflammation, there is a
pathological increase in vascular leakage mediated by VEGF
(45); Ang-1 has been reported to
counteract the VEGF-induced inflammation and vascular leakage in
endothelial cells as well as exhibiting an additive effect on
vessel formation (4).
GM is the active compound of GS and is found in all
parts of the plant (46). GS
possesses glucose lowering effects, which are associated with
reduced ROS in rats and improved antioxidant parameters, such as
like glutathione, glutathione peroxidase, catalase and
malondialdehyde (47). GAPDH is a
key enzyme in the glycolysis pathway (18); it has been reported to interact with
GAPDH and has been indicated to serve an important role in
hypoglycemic activity (19).
Previous studies have demonstrated that GM inhibits Na+
dependent active glucose transport in the small intestine (47) and inhibits glucose-stimulated
gastric inhibitory peptide (48).
GM molecules prevent glucose absorption in the intestine of rats by
filling the receptor location in the absorptive external layers of
the intestinal wall (19).
Furthermore, GM can inhibit glucose uptake in the intestine of
guinea pigs, which occurs from to the suppressive effect of GM on
high K+-induced contraction in guinea pig ileal
longitudinal muscles (49). Thus,
the use of GM can be beneficial for diabetic animals owing to its
anti-inflammatory properties along with anti-oxidant potential.
Hyperglycemia, a high blood glucose can affect
endothelial cells dysfunction of brain blood vessels in STZ
induced-diabetic rats, resulting in brain hypoxia-induced
angiogenesis by upregulating expression of the hypoxia-inducible
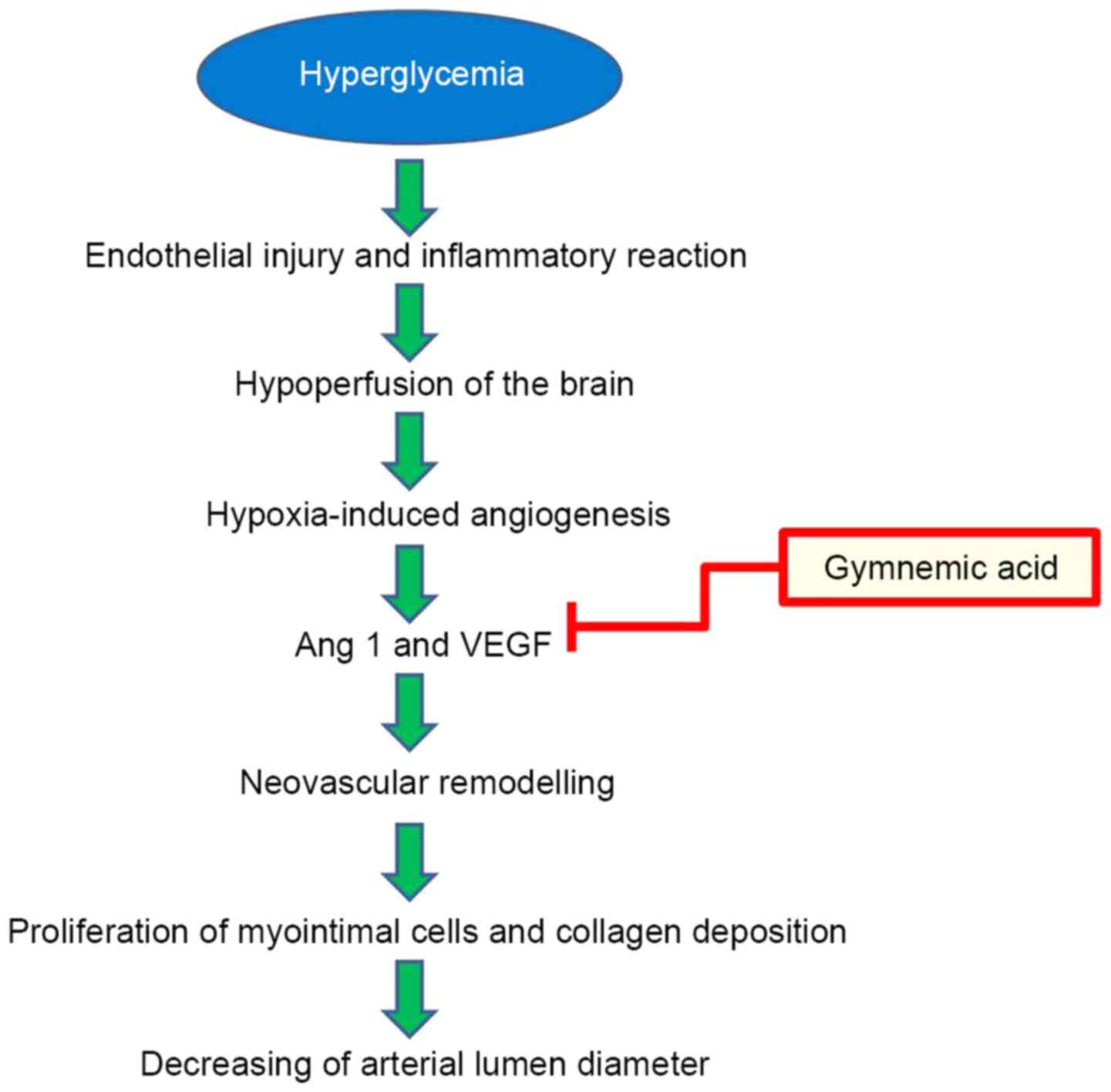
gene (50). The beneficial effects
of GM treatment (400 mg/kg) improved the pathological changes in
brain blood vessels by reducing neovascular remodeling via the
inhibition of VEGF and Ang-1 (Fig.
12). VEGF protein and receptors increase lumen formation and
decrease lumen diameter by proliferation of myointimal cells and
collagen deposition (51), and
Ang-1 serves important roles in angiogenesis and vascular
development. Ang-1 has powerful vascular protective effects, such
as suppressing plasma leakage, inhibiting vascular inflammation and
preventing endothelial death (52).
In the present study, vascular corrosion casting and
microscopy demonstrated that treatment with GM in the diabetic rat
group recovered the healthy architecture of vessels. GM
administration resulted in the improvement and restoration of
injured brain blood vessels induced by hyperglycemia. The lumen
diameter, wall thickness of arteries and the diameter of arteries,
arterioles and capillaries were all improved.
The present study is not without limitations. For
example, the study only assessed the effects of GM substances for 8
weeks to determine whether they could restore the small blood
vessels of the brain. A longer period of time is required to
determine the effective regeneration of large blood vessels.
Taken together, the results of the present study
demonstrated that the restoration and improvement of brain vascular
characteristics in the hypothalamus, through decreased VEGF
expression and Ang-1 secretion, may be beneficial for investigating
the role of GM as a therapeutic target for stroke following
treatment for diabetes.
Acknowledgements
The authors would like to thank Mrs. Anna Chatthong
from International Relations Officer, Faculty of Science, Prince of
Songkla University for their assistance with revising the
manuscript.
Funding
The present study was supported by the Prince of Songkla
University Research Fund (grant no. SCI610515S).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WK designed and conducted the research. NS and PB
prepared animal tissue and performed staining. WK, NS and RJ
performed vascular corrosion casting. NS and MK performed the
western blot analysis. WK wrote the manuscript and performed
statistical analysis. The authenticity of all raw data was assessed
by WK and NS. All authors have read and approved the final version
of the manuscript.
Ethics approval and consent to
participate
The experimental protocol used was approved by the
Animal Ethics Committee of the Prince of Songkla University (Hat
Yai, Songkhla, Thailand).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
American Diabetes Association. Diagnosis
and classification of diabetes mellitus. Diabetes Care. 32 (Suppl
1):S62–S67. 2009.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Enache AL, Slujitoru AS, Pintea IL,
Stocheci CM, Mateescu GO and Gheorghişor I: Histological and
immunohistochemical aspects of cerebral vessels of the elderly. Rom
J Morphol Embryol. 53:1043–1050. 2012.PubMed/NCBI
|
|
3
|
Naderi A, Zahed R, Aghajanpour L, Amoli FA
and Lashay A: Long term features of diabetic retinopathy in
streptozotocin-induced diabetic Wistar rats. Exp Eye Res.
184:213–220. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Javed K and Das MJ: Neuroanatomy,
Posterior Cerebral Arteries. StatPearls Publishing, Treasure
Island, FL, 2020.
|
|
5
|
Mukherjee D, Jani ND, Narvid J and Shadden
SC: The role of circle of Willis anatomy variations in
cardio-embolic stroke: A patient-specific simulation based study.
Ann Biomed Eng. 46:1128–1145. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yu R and Lui F: Neuroanatomy, Brain
Arteries. StatPearls Publishing, Treasure Island, FL, 2019.
|
|
7
|
Shi Y and Vanhoutte PM: Macro- and
microvascular endothelial dysfunction in diabetes. J Diabetes.
9:434–449. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Conway EM, Collen D and Carmeliet P:
Molecular mechanisms of blood vessel growth. Cardiovasc Res.
49:507–521. 2001.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Rodella LF and Favero G: Atherosclerosis
and current anti-oxidant strategies for atheroprotection. Available
from: https://app.dimensions.ai/details/publication/pub.1024158465.
|
|
10
|
Shibuya M: Vascular endothelial growth
factor (VEGF) and its receptor (VEGFR) signaling in angiogenesis: A
crucial target for anti- and pro-angiogenic therapies. Genes
Cancer. 2:1097–1105. 2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Aguilera KY and Brekken RA: Recruitment
and retention: Factors that affect pericyte migration. Cell Mol
Life Sci. 71:299–309. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wirostko B, Wong TY and Simó R: Vascular
endothelial growth factor and diabetic complications. Prog Retin
Eye Res. 27:608–621. 2008.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Akwii RG, Sajib MS, Zahra FT and Mikelis
CM: Role of Angiopoietin-2 in vascular physiology and
pathophysiology. Cells. 8(471)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Melgar-Lesmes P and Edelman ER:
Monocyte-endothelial cell interactions in the regulation of
vascular sprouting and liver regeneration in mouse. J Hepatol.
63:917–925. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Thakur G, Sharma R, Sanodiya BS, Pandey M
and Bisen P: Gymnema sylvestre: An alternative therapeutic
agent for management of diabetes. Pharm Sci Technol Today. 2:1–6.
2012.
|
|
16
|
Ahamad J, Amin S and Mir SR: Simultaneous
quantification of gymnemic acid as gymnemagenin and charantin as
β-sitosterol using validated HPTLC densitometric method. J
Chromatogr Sci. 53:1203–1209. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
El Shafey AAM, El-Ezabi MM, Seliem MME,
Ouda HHM and Ibrahim DS: Effect of Gymnema sylvestre R. Br.
leaves extract on certain physiological parameters of diabetic
rats. J King Saud Univ Sci. 25:135–141. 2013.
|
|
18
|
Tiwari P, Mishra BN and Sangwan NS:
Phytochemical and pharmacological properties of Gymnema
sylvestre: An important medicinal plant. BioMed Res Int.
2014(830285)2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Sahu NP, Mahato SB, Sarkar SK and Poddar
G: Triterpenoid saponins from Gymnema sylvestre.
Phytochemistry. 41:1181–1185. 1996.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Li Y, Sun M, Liu Y, Liang J, Wang T and
Zhang Z: Gymnemic acid alleviates type 2 diabetes mellitus and
suppresses endoplasmic reticulum stress in vivo and in vitro. J
Agric Food Chem. 67:3662–3669. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Nazaruk J and Borzym-Kluczyk M: The role
of triterpenes in the management of diabetes mellitus and its
complications. Phytochem Rev. 14:675–690. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Prabhu S and Vijayakumar S: Antidiabetic,
hypolipidemic and histopathological analysis of Gymnema
sylvestre (R. Br) leaves extract on streptozotocin induced
diabetic rats. Biomed Prev Nutr Internet. 4:425–430. 2014.
|
|
23
|
Wu F, Jin Z and Jin J: Hypoglycemic
effects of glabridin, a polyphenolic flavonoid from licorice, in an
animal model of diabetes mellitus. Mol Med Rep. 7:1278–1282.
2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Komolkriengkrai M, Nopparat J,
Vongvatcharanon U, Anupunpisit V and Khimmaktong W: Effect of
glabridin on collagen deposition in liver and amelioration of
hepatocyte destruction in diabetes rats. Exp Ther Med.
18:1164–1174. 2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cui Y, Xu T, Chen J, Tian H and Cao H:
Anatomic variations in the anterior circulation of the circle of
Willis in cadaveric human brains. Int J Clin Exp Med.
8:15005–15010. 2015.PubMed/NCBI
|
|
26
|
Fatani AJ, Al-Rejaie SS, Abuohashish HM,
Al-Assaf A, Parmar MY, Ola MS and Ahmed MM: Neuroprotective effects
of Gymnema sylvestre on streptozotocin-induced diabetic
neuropathy in rats. Exp Ther Med. 9:1670–1678. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Rekhter MD: Collagen synthesis in
atherosclerosis: Too much and not enough. Cardiovasc Res.
41:376–384. 1999.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Grinberg LT and Thal DR: Vascular
pathology in the aged human brain. Acta Neuropathol. 119:277–290.
2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chen R, Ovbiagele B and Feng W: Diabetes
and stroke: Epidemiology, pathophysiology, pharmaceuticals and
outcomes. Am J Med Sci. 351:380–386. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Castilla-Guerra L and Fernandez-Moreno MC:
Stroke in diabetic patients: Is it really a macrovascular
complication? Stroke. 38:e106. 2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Poittevin M, Bonnin P, Pimpie C, Rivière
L, Sebrié C, Dohan A, Pocard M, Charriaut-Marlangue C and Kubis N:
Diabetic microangiopathy: Impact of impaired cerebral
vasoreactivity and delayed angiogenesis after permanent middle
cerebral artery occlusion on stroke damage and cerebral repair in
mice. Diabetes. 64:999–1010. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Yin K-J, Hamblin M and Chen YE:
Angiogenesis-regulating microRNAs and ischemic stroke. Curr Vasc
Pharmacol. 13:352–365. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Marti HJ, Bernaudin M, Bellail A, Schoch
H, Euler M, Petit E and Risau W: Hypoxia-induced vascular
endothelial growth factor expression precedes neovascularization
after cerebral ischemia. Am J Pathol. 156:965–976. 2000.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Zhang Z and Chopp M: Vascular endothelial
growth factor and angiopoietins in focal cerebral ischemia. Trends
Cardiovasc Med. 12:62–66. 2002.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Niu G and Chen X: Vascular endothelial
growth factor as an anti-angiogenic target for cancer therapy. Curr
Drug Targets. 11:1000–1017. 2010.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Matsuo R, Ago T, Kamouchi M, Kuroda J,
Kuwashiro T, Hata J, Sugimori H, Fukuda K, Gotoh S, Makihara N, et
al: Clinical significance of plasma VEGF value in ischemic stroke -
research for biomarkers in ischemic stroke (REBIOS) study. BMC
Neurol. 13(32)2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Fagiani E, Lorentz P, Kopfstein L and
Christofori G: Angiopoietin-1 and -2 exert antagonistic functions
in tumor angiogenesis, yet both induce lymphangiogenesis. Cancer
Res. 71:5717–5727. 2011.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Akwii RG, Sajib MS, Zahra FT and Mikelis
CM: Mikelis Role of Angiopoietin-2 in vascular physiology and
pathophysiology. Cells. 8(471)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Peplow PV: Growth factor- and
cytokine-stimulated endothelial progenitor cells in post-ischemic
cerebral neovascularization. Neural Regen Res. 9:1425–1429.
2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Prakash R, Somanath PR, El-Remessy AB,
Kelly-Cobbs A, Stern JE, Dore-Duffy P, Johnson M, Fagan SC and
Ergul A: Enhanced cerebral but not peripheral angiogenesis in the
Goto-Kakizaki model of type 2 diabetes involves VEGF and
peroxynitrite signaling. Diabetes. 61:1533–1542. 2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Abdelsaid M, Coucha M, Hafez S, Yasir A,
Johnson MH and Ergul A: Enhanced VEGF signalling mediates cerebral
neovascularisation via downregulation of guidance protein ROBO4 in
a rat model of diabetes. Diabetologia. 60:740–750. 2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Pugh CW and Ratcliffe PJ: Regulation of
angiogenesis by hypoxia: Role of the HIF system. Nat Med.
9:677–684. 2003.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Pogue AI and Lukiw WJ: Angiogenic
signaling in Alzheimer's disease. Neuroreport. 15:1507–1510.
2004.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Gamble JR, Drew J, Trezise L, Underwood A,
Parsons M, Kasminkas L, Rudge J, Yancopoulos G and Vadas MA:
Angiopoietin-1 is an antipermeability and anti-inflammatory agent
in vitro and targets cell junctions. Circ Res. 87:603–607.
2000.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Thurston G, Suri C, Smith K, McClain J,
Sato TN, Yancopoulos GD and McDonald DM: Leakage-resistant blood
vessels in mice transgenically overexpressing angiopoietin-1.
Science. 286:2511–2514. 1999.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Ibrahim A, Onyike E, Nok AJ and Umar IA:
Combined effect on antioxidant properties of Gymnema
sylvestre and combretum micranthum leaf extracts and the
relationship to hypoglycemia. Eur Sci J: Dec 31, 2017 (Epub ahead
of print). doi: org/10.19044/esj.2017.v13n36p266.
|
|
47
|
Yoshioka S: Inhibitory effects of gymnemic
acid and an extract from the leaves of Zizyphus jujuba on
glucose absorption in the rat small intestine. J Yonago Med Assoc.
37:142–154. 1986.
|
|
48
|
Fushiki T, Kojima A, Imoto T, Inoue K and
Sugimoto E: An extract of Gymnema sylvestre leaves and
purified gymnemic acid inhibits glucose-stimulated gastric
inhibitory peptide secretion in rats. J Nutr. 122:2367–2373.
1992.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Shimizu K, Iino A, Nakajima J, Tanaka K,
Nakajyo S, Urakawa N, Atsuchi M, Wada T and Yamashita C:
Suppression of glucose absorption by some fractions extracted from
Gymnema sylvestre leaves. J Vet Med Sci. 59:245–251.
1997.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Krock BL, Skuli N and Simon MC:
Hypoxia-induced angiogenesis: Good and evil. Genes Cancer.
2:1117–1133. 2011.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Xu J and Shi GP: Vascular wall
extracellular matrix proteins and vascular diseases. Biochim
Biophys Acta. 1842:2106–2119. 2014.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Brindle NPJ, Saharinen P and Alitalo K:
Signaling and functions of angiopoietin-1 in vascular protection.
Circ Res. 98:1014–1023. 2006.PubMed/NCBI View Article : Google Scholar
|