Introduction
In December 2019, a cluster of patients with
pneumonia caused by an unknown pathogen occurred in Wuhan (China)
and rapidly spread across the whole country (1,2).
Although China took an emergency response in the early days of the
outbreak, the infection has been spreading globally. From airway
epithelial cell samples, a novel coronavirus was determined by the
Chinese Center for Disease Control and Prevention using
next-generation sequencing technology and was named severe acute
respiratory syndrome coronavirus-2 (SARS-CoV-2) by the World Health
Organization. The sequence of SARS-CoV-2 is highly homologous to
severe acute respiratory syndrome coronaviruses (SARS-CoV)
(3).
Since the sequence of SARS-CoV-2 is highly
homologous to that of SARS-CoV, the clinical manifestations of the
patients with coronavirus disease 2019 (COVID-19) caused by
SARS-CoV-2 are similar to those of patients with SARS and Middle
East respiratory syndrome coronavirus (MERS-CoV) (4). The main manifestations are high fever
and cough, which may be accompanied by headache, myalgia, joint
pain, fatigue, diarrhea and chest tightness at onset. The patients
may have dyspnea, shortness of breath and then respiratory failure
at the later stage (5,6). SARS is a systemic organ damage
disease, the main target organs of which are the lung, immune
organs and systemic venules. In addition, virus-induced
orchitis-like pathological changes were detected during autopsy of
numerous patients who succumbed to SARS, even those with no related
symptoms during their lifetime (7).
Ma et al (8) reported that
the ratio of testosterone (T) to serum luteinizing hormone (LH) and
the ratio of follicle-stimulating hormone to LH were significantly
reduced in patients with COVID-19. Given that the sequences of
SARS-CoV-2 and SARS-CoV are highly homologous, SARS-CoV-2 may be
likely to injure testicular tissues.
In 2003, angiotensin-converting enzyme II (ACE2) was
identified as the functional receptor for SARS-CoV (9), which is able to bind to the
receptor-binding domain of viral Spike protein, causing endocytosis
and infection (10). It has been
reported that SARS-CoV-2 shares the same receptor with SARS-CoV,
i.e. ACE2 (11-13).
As the majority of ACE2-expressing cells in the respiratory tract
are type II alveolar epithelial cells (AT2), SARS-CoV-2 invasion
and the corresponding manifestations are mainly caused by
interaction with ACE2 receptors on the surface of AT2(14). A study has indicated that
ACE2-expressing epithelial cells of the small intestine were
susceptible to SARS-CoV-2 infection (15).
SARS-CoV-2 has been reported to damage multiple
organs and alter the ratio of male sex hormones in males with
COVID-19(8); however, its direct
effect on the testis has remained to be fully determined. In the
present study, the expression profiles of ACE2 were investigated by
analyzing single-cell RNA-sequencing (RNA-seq) datasets and
immunohistochemical (IHC) staining to explore the expression of
ACE2 and assess the effect of SARS-CoV-2 on the male productive
function.
Materials and methods
Data sources
Single-cell RNA-seq datasets (nos. GSE120506 and
GSE112013) were downloaded from the Gene Expression Omnibus (GEO)
database (https://www.ncbi.nlm.nih.gov/geo/). RNA-seq data of
normal human tissues from the GTEx database were downloaded using
the University of California Santa Cruz (UCSC) Xena browser
(https://xena.ucsc.edu/).
ACE2 expression patterns in different
tissues from normal human subjects from GTEx
The dataset gene expression RNAseq-TOIL RSEM
fragments per kilobase of exon model per million mapped fragments
and dataset phenotype-GTEx phenotype were downloaded from UCSC
Xena. Strawberry-perl 5.3.2.1 (https://strawberryperl.com/) was used to quantify the
mRNA expression of ACE2 in each organ. The R package ‘ggpubr’ was
used to draw boxplots of ACE2 mRNA expression in each organ.
Single-cell sequencing analysis
Single-cell RNA-seq datasets (nos. GSE120506 and
GSE112013) were downloaded from the GEO database. The Single cell
RNA sequencing (scRNA-seq) datasets of
GSE112013_Combined_UMI_table.txt and
GSE120506_infant_combined_UMI.txt were subsequently analyzed using
the R package Seurat (version 3.1.2). Strict parameters,
‘min.cells=3’ and ‘min. features=50’, were used to filter out
low-quality cells and low-quality genes in the function
CreateSeuratObject. Subsequently, the data were log-normalized
using the function NormalizeData with the default parameters. The
genes, with the largest standard deviation of expression in all
cells were determined by the function FindVariableGenes. After the
data were dealt with, the function ScaleData, principal component
analysis (PCA) dimensionality reduction was performed by using the
function RunPCA. By using the functions of JackStraw and
ScoreJackStraw, the PCA components with P<0.05 were selected for
further analysis using two-dimensional t-distributed stochastic
neighbor embedding. The setting of k.param was 20 in the function
FindNeighbors and the setting of the resolution was 0.5 in the
function FindClusters. The cell types were determined according to
the expression of marker genes. Gene expression of different cell
types was displayed using the functions DotPlot and VlnPlot.
IHC staining of human samples
Human normal testicular tissue sections (n=4)
included in normal tissue microarrays were obtained from Alenabio.
In addition, the human testicular tissues from 11 patients with
prostate cancer were obtained from the Third Affiliated Hospital of
Guangzhou Medical University (Guangzhou, China). The specimen
collection started on 1 September 2020 and was completed by 31
October 2020. For IHC staining, 3-µm sections were prepared. IHC
staining was performed as described in a previous study by our
group (16). In brief, the sections
were dewaxed, hydrated and incubated with anti-ACE2 antibody (1:400
dilution; cat. no. ab15348; Abcam) overnight at 4˚C. Subsequently,
HRP-conjugated secondary antibody (1:100 dilution; cat. no. BM3894;
Boster Biotechnology, Inc.) was added to the sections and incubated
for 50 min at 37˚C. All of the sections were stained with
diaminobenzidine and counterstained with hematoxylin. Images were
captured using an upright microscope and the expression levels of
ACE2 were evaluated by two experimenters. The data were
independently collected and evaluated by two experimenters (GM and
RZ) in a double-blind manner.
Statistical analysis
The Graphpad Prism 5 software package (GraphPad
Software, Inc.) was used to analyze the data and draw the
statistical charts. The two-tailed Student's t-test was used to
statistically analyze the results. P<0.05 was considered to
indicate a statistically significant difference.
Results
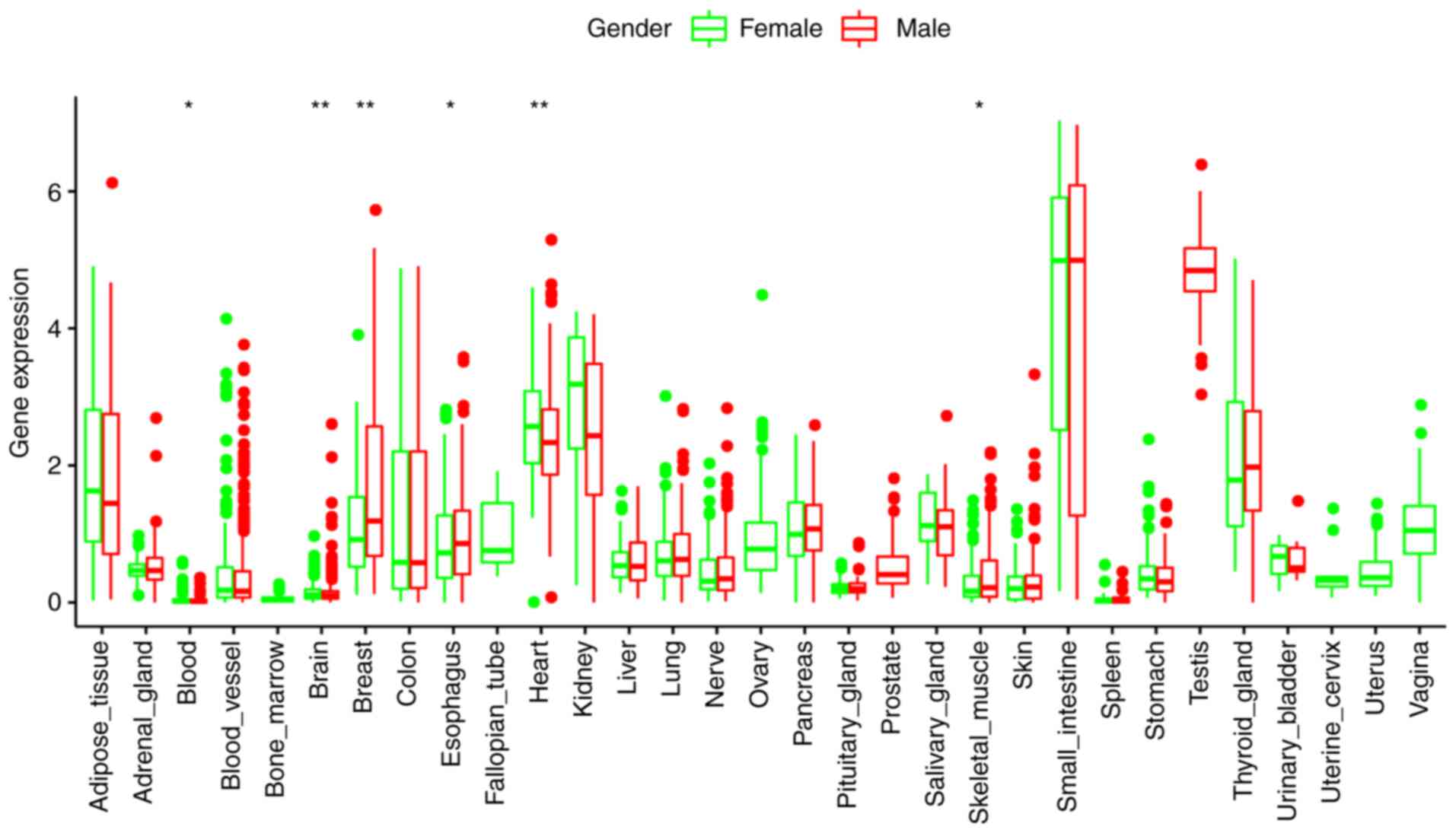
ACE2 mRNA expression in different
organs of normal human subjects
ACE2 mRNA expression was detected in different
organs of normal human subjects by analyzing RNA-seq data from the
GTEx database. ACE2 mRNA was indicated to be highly expressed both
in the small intestine and testis (Fig.
1). These results indicated that testis may be susceptible to
SARS-CoV-2 because SARS-CoV-2 invades host cells via ACE2
receptors.
Identification of cell types in the
testes of adults and infants
Given that SARS-CoV-2 invades host cells via ACE2
receptors and SARS-CoV is able to cause orchitis (7), ACE2-expression by cell type was
detected in the present study by analyzing single-cell RNA-seq
data. The R package Seurat was used to re-analyze the scRNA-seq
datasets GSE112013 and GSE120506. In order to normalize the data,
the data of the datasets were filtered and normalized. After the
data were normalized, 1,500 genes from GSE112013 (Fig. S1) and 5,000 genes from GSE120506
(Fig. S2), with larger
coefficients of variation (min cell=3, min features=50), were
selected for subsequent analysis and the top 10 genes with the
largest standard deviation were displayed. In order to reduce the
linear dimensionality, a PCA algorithm was used to analyze the
dataset. It was indicated that the PCs (P<0.05) of the dataset
from the adult testes (Fig. S3)
and the infant testes (Fig. S4)
were suitable for further analysis.
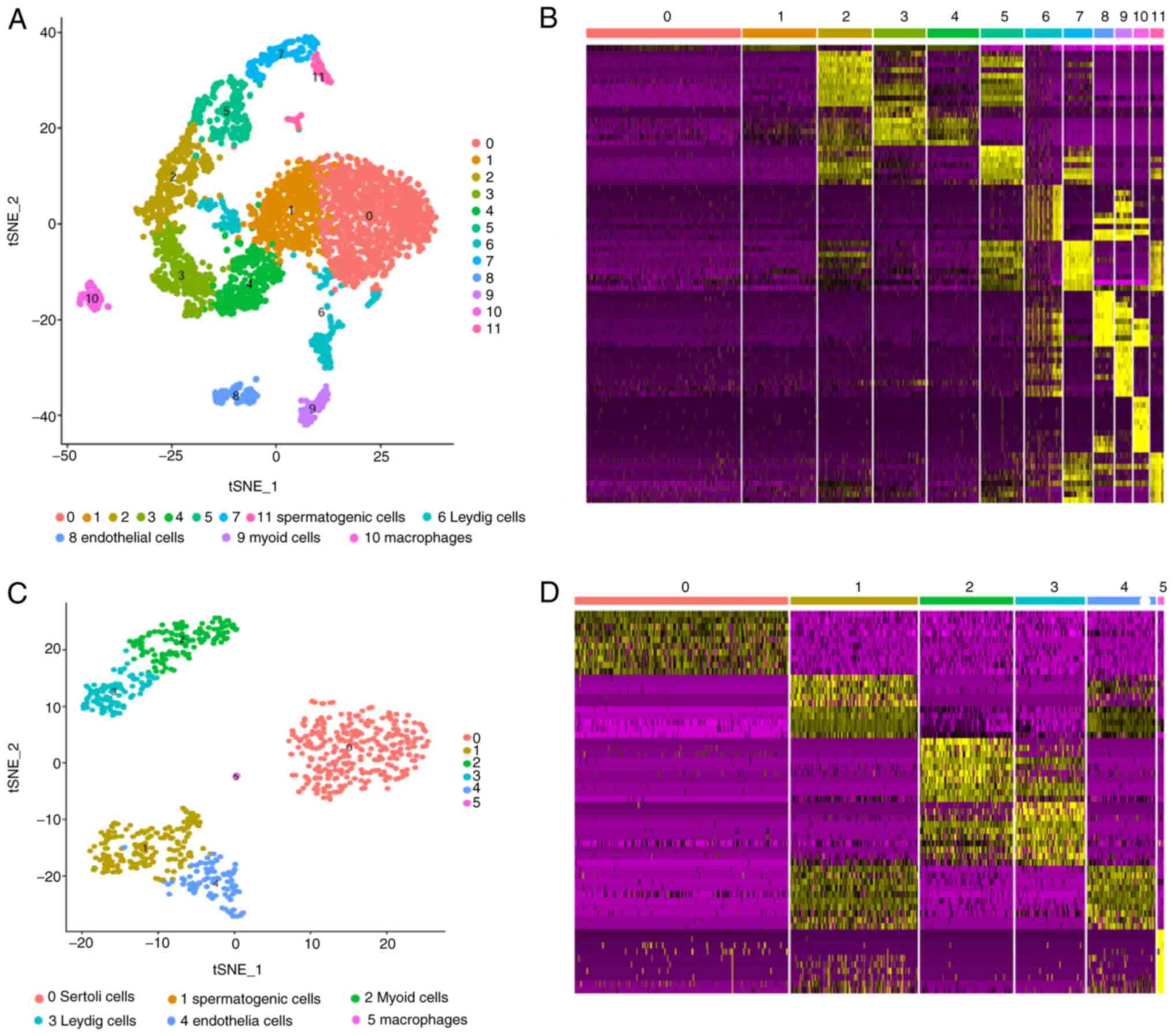
A total of 3,652 individual cells from the testes of
three healthy males of peak reproductive age and 813 individual
cells from the testes of two infants (age, 13 months) were
analyzed. Using unsupervised graph-based clustering, it was
indicated that at least 12 distinct cell clusters were present in
the testicular tissues of the adult males (Fig. 2A and B) and six cell clusters were identified in
the infant testicular tissues (Fig.
2C and D) according to their
corresponding marker gene expression profiles.
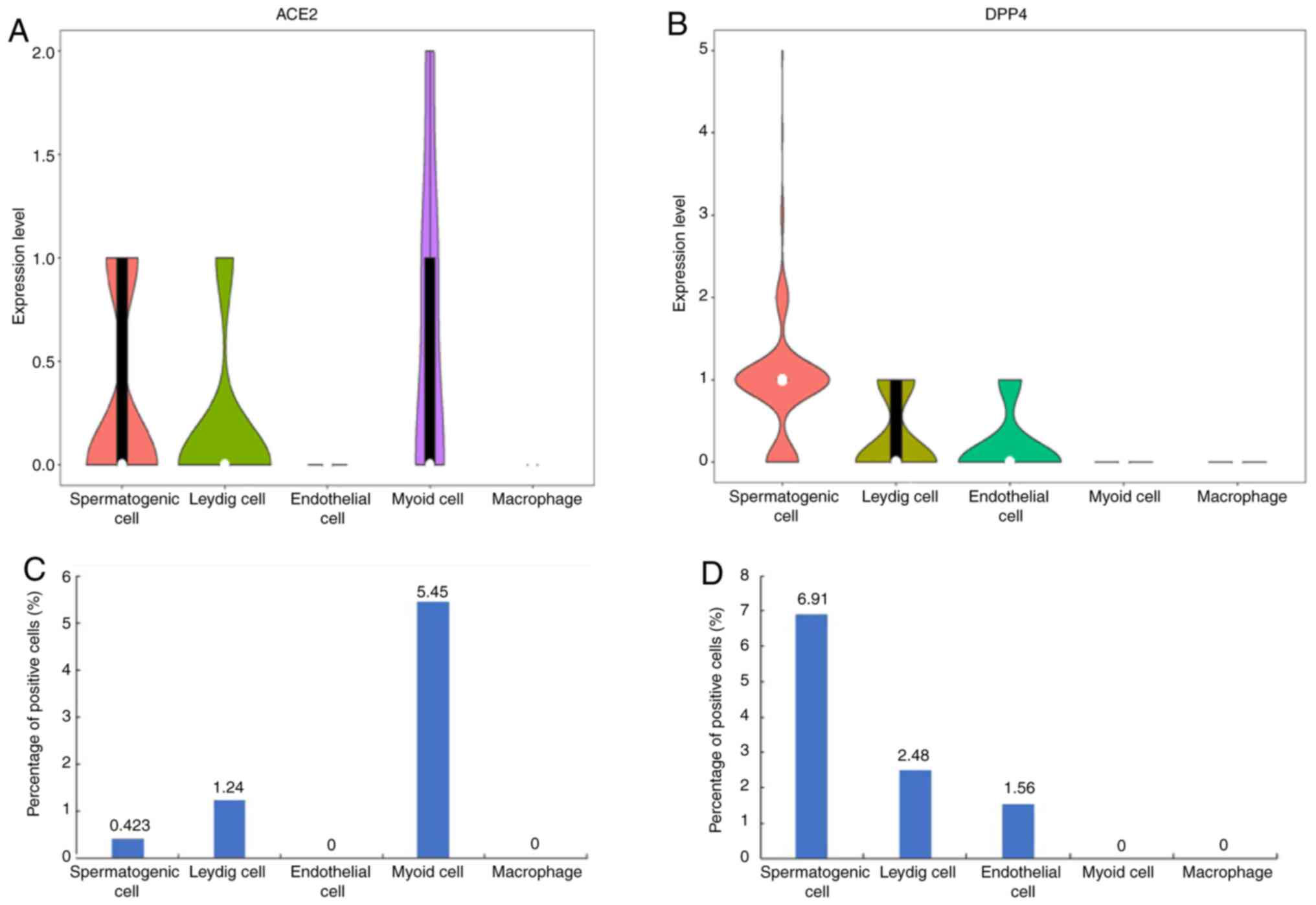
Identification of ACE2-expressing
cells in adult testes
To determine the cell types expressing ACE2, the
mRNA expression of ACE2 was analyzed from scRNA-seq datasets. The
violin plots indicated that ACE2 was highly expressed in
spermatogenic cells, Leydig cells and myoid cells of the adult male
testicular tissues, but hardly expressed in endothelial cells and
macrophages (Fig. 3A). Of note,
investigation of the expression profiles of another virus receptor,
dipeptidyl peptidase 4 (DPP4) receptor for MERS-CoV virus,
indicated that DPP4 was highly expressed in spermatogenic cells,
Leydig cells and endothelial cells of the adult male testicular
tissues, while its expression was low in myoid cells and
macrophages. Furthermore, the expression of DPP4 in spermatogenic
cells was higher compared with that of ACE2 (Fig. 3B). ACE2-expressing myoid cells,
Leydig cells and spermatogenic cells were determined to account for
5.45, 1.24 and 0.423% of adult testicular cells, respectively
(Fig. 3C). Furthermore,
DPP4-expressing spermatogenic cells, Leydig cells and endothelial
cells accounted for 6.91, 2.48 and 1.56% of adult testicular cells,
respectively (Fig. 3D). These
results indicated that SARS-CoV-2 and MERS-CoV may damage the adult
testis, as these two viral receptors have significantly overlapped
and increased expression profiles.
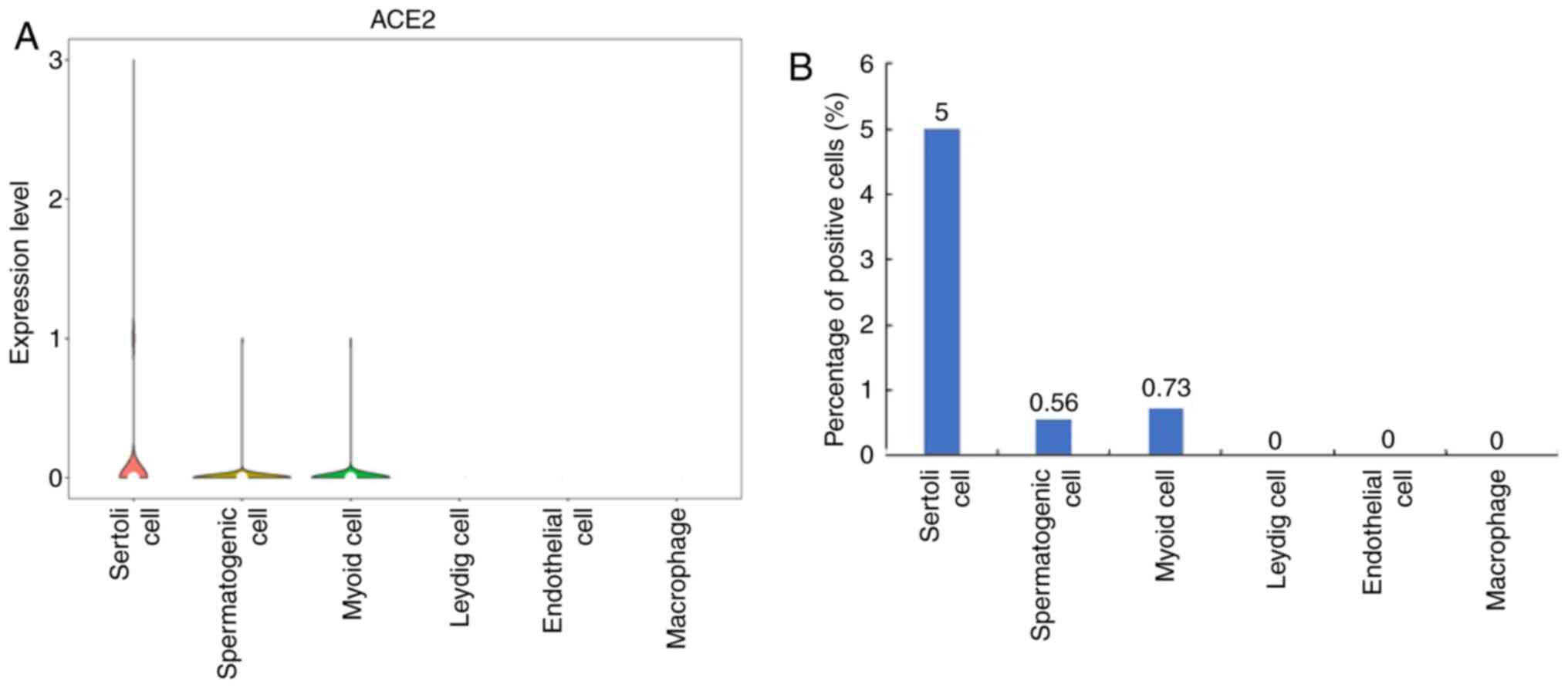
Identification of ACE2-expressing
cells in infant testes
To predict the effect of SARS-CoV-2 on infant testes
and determine the cell types expressing ACE2 in infant testes, the
mRNA expression of ACE2 in infant testicular tissues was also
analyzed. It was determined that ACE2 was highly expressed in
Sertoli cells of the infant testicular tissues, but hardly
expressed in spermatogenic cells and myoid cells (Fig. 4A). ACE2-expressing Sertoli cells,
spermatogenic cells and myoid cells accounted for 5.00, 0.56 and
0.73% of infant testicular cells, respectively (Fig. 4B). These results suggested that
SARS-CoV-2 may also damage the testes of infants.
ACE2 protein is highly expressed in
normal testicular tissues
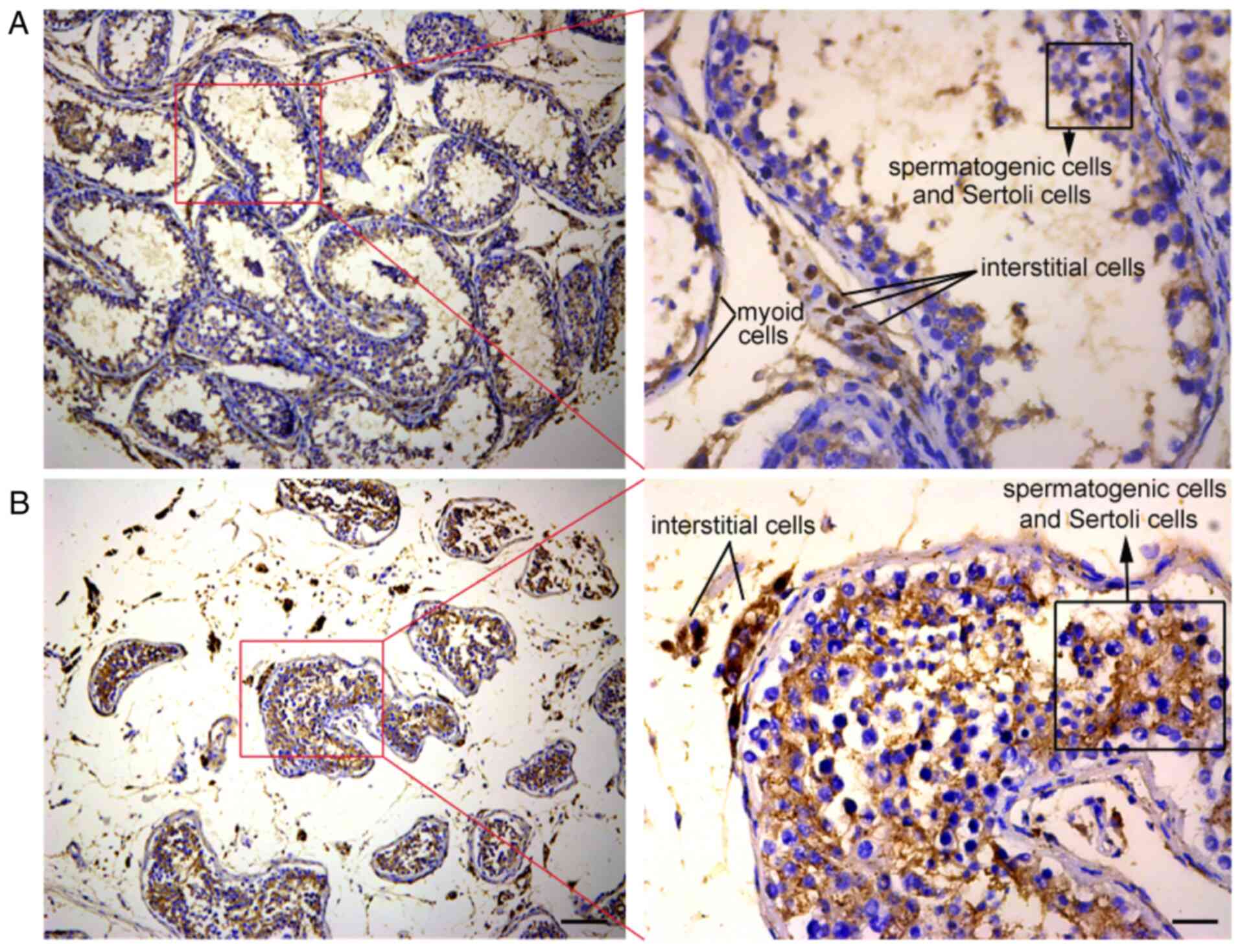
To determine the expression of ACE2 protein in
normal testicular tissues, the testicular tissues from 15 patients
with prostate cancer were sectioned and subjected to IHC analysis.
IHC staining for ACE2 was performed on the testicular tissue
sections. The results indicated that ACE2 protein was highly
expressed in the normal testicular tissues, including spermatogenic
cells, Sertoli cells, interstitial cells and myoid cells (Fig. 5A and B). These results further demonstrated that
the testis may be target organs of SARS-CoV-2 and imply that
SARS-CoV-2 may cause testicular damage and impair reproductive
function, and even lead to male infertility.
Discussion
Recently, COVID-19 has caused a great health threat
throughout the world and led to the declaration of a public health
emergency of international concern by the World Health Organization
(6,13). The pathogen is SARS-CoV-2, a
single-stranded RNA virus, which has a stronger transmission
ability than SARS. Apart from lung injury as the most common
manifestation, COVID-19 is able to cause systemic/multiple-organ
damage, including the heart, kidney and liver. However, whether
SARS-CoV-2 infects the testis and causes damage to male fertility
function has remained elusive.
As mentioned above, as ACE2 is the cell entry
receptor for SARS-CoV-2, it is worthwhile to detect ACE2 expression
in testicular tissue in order to explore whether male testes are a
potential target of virus invasion. In the present study,
single-cell RNA-seq expression profiles were analyzed to reveal
that ACE2 was highly expressed in spermatogenic cells, Leydig cells
and myoid cells of adult testes and Sertoli cells of infant
testicular tissues, which demonstrated that not only lung and heart
but also testes are potential target organs. These results suggest
that SARS-CoV-2 may cause testicular damage and impair reproductive
function, even leading to male infertility.
By investigating the expression of ACE2 in normal
human testicles, it was revealed that ACE2 protein was highly
expressed in spermatogenic cells, Leydig cells and myoid cells of
the adult testes. Ding et al (17) reported that SARS-CoV was detected in
numerous organs and tissues, including the lung, trachea, bronchus,
stomach and small intestine, by using IHC and in situ
hybridization, but not in the testes. However, Zhao et al
(18) detected SARS-CoV in the
seminiferous epithelial and mesenchymal cells of the testes using
electron microscopic examination of post-mortem materials. In the
present study, the expression of ACE-2 in tissues from patients
with COVID-19 was not examined, as it was not possible to obtain
tissues from post-mortem patients. Since ACE2 is highly expressed
in testicular cells, it may be speculated that the testis are also
a potential target of SARS-CoV-2 infection.
SARS-CoV is able to cause testicular damage,
manifested as a thickened basement membrane of seminiferous tubules
and fibrosis of seminiferous tubules. The seminiferous epithelial
cells were necrotic and fell off to different degrees, resulting in
a significant decrease in the number of spermatogenic cells.
Extensive germ cell destruction with few or no spermatozoa in the
lumen of seminiferous tubules were observed. Vascular congestions,
vasodilation and leukocyte infiltration were present in the
testicular stroma (7). Furthermore,
Ma et al (8) indicated that
serum LH and PRL levels were significantly elevated and the ratio
of T:LH was significantly decreased in patients with COVID-19,
implying dysfunction of the testes. Xu et al (1) also reported that RNA and viral
particles of SARS-CoV-2 were detected in testes, and testes were
subjected to various degrees of spermatogenic cell reduction and
injury. The present results also suggested that ACE2 mRNA and
protein were highly expressed in the testis. SARS-CoV-2 invade into
the cells via the ACE2 receptor, the same as SARS-CoV. Therefore,
SARS-CoV-2 may cause severe pathological damage in the testes of
infants and adults.
It has been reported that the virus not only causes
orchitis but also causes infertility and even increases the
incidence of testicular cancer. 59.2% of patients with COVID-19
were reported to be male, of whom 56% were 0-49 years old.
Therefore, close attention should be paid for the reproductive
function of male inpatients with COVID-19, particularly those with
severe disease. If clinically cured patients were to be further
followed up and were subjected to periodic comprehensive testing of
the sperm count, morphology and vitality, it would further the
understanding of COVID-19, and is directly related to the recovery
of patients with COVID-19 and their quality of life.
Supplementary Material
Raw data of the dataset GSE112013 from
single-cell RNA sequencing of the adult testis were filtered and
normalized. (A) The low-quality data of the dataset were filtered
as follows: Genes expressed in <3 cells and samples with <50
cells were detected. (B) No correlation was present between the
percentage of mitochondrial genes and the sequencing depth. A
positive correlation was detected between the gene number and
sequencing depth (R=0.86; P<0.05). (C) The data were
log-normalized and genes with larger coefficients of variation were
selected. 1,500 differential genes with the largest standard
deviation of expression in all cells were selected for subsequent
analysis and the top 10 differential genes were displayed. Mt,
mitochondrial genes.
Raw data of the dataset GSE120506 from
single-cell RNA sequencing of infant testis were filtered and
normalized. (A) The filtered data of the dataset. (B) No
correlation was present between the percentage of mitochondrial
genes and the sequencing depth. A positive correlation was detected
between the gene number and sequencing depth (R=0.97, P<0.05).
(C) The data were log-normalized and genes with a larger
coefficient of variation were selected. A total of 5,000
differential genes with the largest standard deviation of
expression in all cells were selected for subsequent analysis and
the top 10 differential genes were displayed. Mt, mitochondrial
genes.
PCA of the dataset GSE112013 from
single-cell RNA sequencing of adult testis. (A) A PCA algorithm was
used to analyze the dataset. (B) The top 20 genes in the top 4 PCs
and the correlation between genes and each component. (C) Heat map
displaying the marker genes in the top 4 PCs. (D) P-value and mean
distribution of the 20 PCs. PCA, principal component analysis.
PCA of the dataset GSE120506 from
single-cell RNA sequencing of infant testis. (A) A PCA algorithm
was used to analyze the dataset. (B) The top 20 genes in the top 4
PC and the correlation between genes and each component. (C) Heat
map displaying the marker genes in the top 4 PC. (D) P-value and
mean distribution of the 20 PCs. PCA, principal component
analysis.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National Science
Foundation of China (grant no. 81773095 to CQ and 31771578 to LW)
and the Youth Research Fund of the Third Affiliated Hospital of
Guangzhou Medical University (grant no. 2017Q12 to TC).
Availability of data and materials
The single scRNA-seq datasets generated and/or
analyzed during the current study are available in the GEO
repository (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE120506
and https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE112013).
Authors' contributions
CQ and LW conceived and designed the study. TC and
MF acquired and analyzed the single-cell RNA-seq data. RZ and GM
performed the IHC staining. TC and CQ performed the statistical
analysis. XY evaluated the immunohistochemical results, reviewed
the manuscript as a clinical laboratory specialist and provided
critical guidance in the process of manuscript revision. CQ wrote
and modified the manuscript. CQ and TC confirmed the authenticity
of the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All patients provided written informed consent for
their tissues to be used for medical research. Ethical approval for
this study was obtained from the Ethics committee of the Third
Affiliated Hospital of Guangzhou Medical University (Guangzhou,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xu B, Yao XH, Ping YF, Yu SC, Shi Y, Luo
T, et al: Autopsy of COVID-19 victims in China. National Science
Review 2020. https://academic.oup.com/nsr/advance-article/doi/10.1093/nsr/nwaa123/5854209?searchresult=1.
|
|
2
|
Chen N, Zhou M, Dong X, Qu J, Gong F, Han
Y, Qiu Y, Wang J, Liu Y, Wei Y, et al: Epidemiological and clinical
characteristics of 99 cases of 2019 novel coronavirus pneumonia in
Wuhan, China: A descriptive study. Lancet. 395:507–513.
2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hayashi T, Abiko K, Mandai M, Yaegashi N
and Konishi I: Highly conserved binding region of ACE2 as a
receptor for SARS-CoV-2 between humans and mammals. Vet Q.
40:243–249. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Zhou J, Li C, Zhao G, Chu H, Wang D, Yan
HHN, Poon VKM, Wen L, Wong BHY, Zhao X, et al: Human intestinal
tract serves as an alternative infection route for Middle East
respiratory syndrome coronavirus. Sci Adv.
3(eaao4966)2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Guan WJ, Ni ZY, Hu Y, Liang WH, Ou CQ, He
JX, Liu L, Shan H, Lei CL, Hui DSC, et al: Clinical characteristics
of coronavirus disease 2019 in China. N Engl J Med. 382:1708–1720.
2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Huang C, Wang Y, Li X, Ren L, Zhao J, Hu
Y, Zhang L, Fan G, Xu J, Gu X, et al: Clinical features of patients
infected with 2019 novel coronavirus in Wuhan, China. Lancet.
395:497–506. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xu J, Qi L, Chi X, Yang J, Wei X, Gong E,
Peh S and Gu J: Orchitis: A complication of severe acute
respiratory syndrome (SARS). Biol Reprod. 74:410–416.
2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Ma L, Xie W, Li DY, Shi L, Mao Y, Xiong Y,
Zhang Y and Zhang M: Effect of SARS-CoV-2 infection upon male
gonadal function: A single center-based study. medRxiv: Mar 30,
2020 (Epub ahead of print). doi:
org/10.1101/2020.03.21.20037267.
|
|
9
|
Li W, Moore MJ, Vasilieva N, Sui J, Wong
SK, Berne MA, Somasundaran M, Sullivan JL, Luzuriaga K, Greenough
TC, et al: Angiotensin-converting enzyme 2 is a functional receptor
for the SARS coronavirus. Nature. 426:450–454. 2003.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kuiken T, Fouchier RA, Schutten M,
Rimmelzwaan GF, van Amerongen G, van Riel D, Laman JD, de Jong T,
van Doornum G, Lim W, et al: Newly discovered coronavirus as the
primary cause of severe acute respiratory syndrome. Lancet.
362:263–270. 2003.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wan Y, Shang J, Graham R, Baric RS and Li
F: Receptor recognition by the novel coronavirus from Wuhan: An
analysis based on decade-long structural studies of SARS
Coronavirus. J Virol. 94(e00127)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Letko M, Marzi A and Munster V: Functional
assessment of cell entry and receptor usage for SARS-CoV-2 and
other lineage B betacoronaviruses. Nat Microbiol. 5:562–569.
2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhou P, Yang XL, Wang XG, Hu B, Zhang L,
Zhang W, Si HR, Zhu Y, Li B, Huang CL, et al: A pneumonia outbreak
associated with a new coronavirus of probable bat origin. Nature.
579:270–273. 2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhang H, Kang Z, Gong H, Xu D, Wang J, Li
Z, Li Z, Cui X, Xiao J, Zhan J, et al: Digestive system is a
potential route of COVID-19: An analysis of single-cell
coexpression pattern of key proteins in viral entry process. Gut.
69:1010–1018. 2020.
|
|
15
|
Liang W, Feng Z, Rao S, Xiao C, Xue X, Lin
Z, Zhang Q and Qi W: Diarrhoea may be underestimated: A missing
link in 2019 novel coronavirus. Gut. 69:1141–1143. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Qi C, Wei B, Zhou W, Yang Y, Li B, Guo S,
Li J, Ye J, Li J, Zhang Q, et al: P-selectin-mediated platelet
adhesion promotes tumor growth. Oncotarget. 6:6584–6596.
2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Ding Y, He L, Zhang Q, Huang Z, Che X, Hou
J, Wang H, Shen H, Qiu L, Li Z, et al: Organ distribution of severe
acute respiratory syndrome (SARS) associated coronavirus (SARS-CoV)
in SARS patients: Implications for pathogenesis and virus
transmission pathways. J Pathol. 203:622–630. 2004.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhao JM, Zhou GD, Sun YL, Wang SS, Yang
JF, Meng EH, Pan D, Li WS, Zhou XS, Wang YD, et al: Clinical
pathology and pathogenesis of severe acute respiratory syndrome.
Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi. 17:217–221.
2003.PubMed/NCBI(In Chinese).
|