Introduction
Oral squamous cell carcinoma (OSCC), a type of head
and neck squamous cell carcinoma, is one of the most common
malignant tumors diagnosed worldwide with high morbidity and
mortality rates (1). According to
the 2018 global cancer statistics, OSCC accounted for ~3% of new
cases and 1.9% of deaths of all malignant cancers (2). Over the past two decades, although a
multidisciplinary approach, including chemotherapy, targeted
therapy and surgical therapy, has been applied for the treatment of
OSCC, the overall 5-year survival rate of patients with OSCC has
remained <50% (3,4). Hence, it is of great importance to
widen the current understanding of the mechanisms underlying OSCC
progression and discover effective biomarkers and therapeutic
strategies for improving OSCC treatment and prognosis.
Claudin-7 (CLDN7), one of the 24 members of the
Claudin family, is involved in regulating tight junction assembly,
intercellular exchange and cell polarity maintenance (5). An increasing number of studies have
confirmed that the abnormal expression of CLDN7 can disrupt the
integrity of tight junctions, thereby resulting in the loss of cell
polarity, and inducing abnormal proliferation, invasion and
metastasis (6,7). Accumulating evidence has shown that
CLDN7 is abnormally expressed in multiple types of cancer tissues,
such as ovarian carcinoma, salivary adenoid cystic carcinoma and
colorectal cancer (6,8,9),
indicating that alterations in CLDN7 expression may be closely
related to the occurrence of these tumors. Emerging evidence
supports the notion that low expression of CLDN7 serves as a
crucial feature of OSCC, and loss of CLDN7 is considered as a
negative prognostic factor for invasion and metastasis in OSCC
(10). However, the exact
regulatory mechanisms of CLDN7 in OSCC remains to be elucidated.
Interferon regulatory factor-2 (IRF2) was predicted as an upstream
regulator of CLDN7 expression, as determined using the PROMO
database, and the highest bound fraction between IRF2 and CLDN7 was
confirmed using the JASPAR database. The roles of IRF2 on
carcinogenesis are controversial. Initially, it was shown that
IRF-2 served as an oncogenic protein, and the expression levels of
IRF-2 were also found to be upregulated in esophageal and
pancreatic cancer (11,12). The downregulation of IRF2 is
responsible for the anti-tumorigenic effects in prostate cancer
cells overexpressing microRNA-221(13). In contrast, IRF2 served as a tumor
suppressor and its inactivation led to impaired P53 function
(14). IRF2 is reported to inhibit
the invasion and migration of gastric cancer cells by inactivating
MMP-1 expression (15). However,
the function of IRF2 in OSCC remains unknown, to the best of our
knowledge.
The present study aimed to investigate the effects
of CLDN7 on proliferation, invasion and migration of OSCC cells,
and identify whether IRF2 could transcriptionally activate CLDN7 to
affect the progression of OSCC.
Materials and methods
Cell culture
SCC-9, CAL-27, SCC-15 and SCC090 human OSCC cell
lines were purchased from the American Type Culture Collection. The
human oral keratinocyte HOK cell line was provided by Shanghai
Haoyuan Biotech Co., Ltd. These cell lines were maintained in DMEM
supplemented with 10% FBS (HyClone; Cytiva) at 37˚C in a humidified
incubator with 5% CO2.
Cell transfection
CAL-27 cells in the logarithmic growth phase were
collected and plated into 6-well plates at a density of
1x106 cells/well at 37˚C. When cells reached 85%
confluence, cells were transfected with 2 µg CLDN7 plasmid
(pc-CLDN7), 2 µg IRF2 plasmid (pc-IRF2) or 2 µg empty vector
plasmid (pcDNA3.1) using Lipofectamine® 3000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37˚C according to
the standard protocol (16). The
aforementioned plasmids were purchased from Shanghai GenePharma
Co., Ltd. Additionally, 2 µg short hairpin RNA (shRNA) targeting
CLDN7 (sh-CLDN7) was constructed in a U6/GFP/Neo plasmid
(GenePharma, Shanghai, China) and transfected into CAL-27 cells
using Lipofectamine® 2000 reagent at 37˚C. Western blot
analysis was employed to evaluate the transfection efficiency 48 h
post-transfection.
Cell viability assay
Cell Counting Kit-8 (CCK-8; Shanghai Yeasen
Biotechnology Co., Ltd.) assays were utilized to assess cell
viability, according to the manufacturer's instructions. CAL-27
cells were inoculated into 96-well plates and cultured at 37˚C
overnight. At the indicated time points (24, 48 or 72 h), 10 µl
CCK-8 solution was added to each well after 2 h of culture at 37˚C.
The absorbance was determined at 450 nm using a microplate reader
(Bio-Rad Laboratories, Inc.).
Colony formation assay
Cells in the logarithmic growth phase were seeded
into a 6-well culture plate (500 cells/well). Cells were cultured
at 37˚C for 2 weeks until colony formation became visible to the
naked eye. Then, cells were fixed with 4% paraformaldehyde for 30
min at room temperature, followed by staining with 0.5% crystal
violet solution (Sigma-Aldrich; Merck KGaA) for 5 min at room
temperature. The number of colony cells with a minimum of 50 cells
was counted under a light microscope (magnification, x200).
Transwell invasion assay
A Matrigel-based assay in 24-well 8-µm pore
Transwell chambers (Costar; Corning, Inc.) was utilized for the
evaluation of cell invasion. A total of 2.5x105 CAL-27
cells were inoculated in serum-free medium in the upper chamber.
DMEM supplemented with 10% FBS was added to the lower compartment
as a chemoattractant. After 24 h of incubation, cells remaining on
the top of the Matrigel were gently removed with a cotton swab.
Cells which had invaded to the lower surface of the membrane were
fixed in 4% paraformaldehyde for 20 min at 37˚C and stained with
0.1% crystal violet (Sigma-Aldrich; Merck KGaA) for 30 min at 37˚C.
Images were captured using an inverted fluorescence microscope
(magnification, x200; Olympus Corporation). The number of cells
which had invaded were counted using ImageJ version 1.52r (National
Institutes of Health).
Wound healing assay
CAL-27 cells were seeded into 6-well plates
(5x104 cells/well) and cultured to 80% confluence. The
cells were serum-starved overnight at 37˚C prior to the experiment.
Then, cells were scratched with a sterile plastic micropipette tip
to generate a wound in the monolayer. After 24 h of incubation, the
wound was imaged under an inverted microscope (Olympus Corporation)
and the separation distance was analyzed using ImageJ.
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to extract the total RNA from
CAL-27 cells, according to the manufacturer's instructions. Then,
RNA was reverse transcribed into cDNA using a RT kit according to
the manufacturer's protocol (Beijing Transgen Biotech Co., Ltd.).
qPCR was performed using SYBR Premix Ex Taq (Takara Biotechnology
Co., Ltd.) on an ABI 7500 Real-Time PCR Detection system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The RT-qPCR conditions
were as follows: Pre-denaturation at 95˚C for 2 min; followed by 35
cycles of 95˚C for 35 sec, 58˚C for 45 sec and 72˚C for 30 sec; and
72˚C for 5 min. The sequences of the primers were: CLDN7 forward,
5'-AAAGTGAAGAAGGCCCGTATA-3' and reverse,
5'-TAATGTTGGTAGGGATCAAAGG-3'; IRF2 forward,
5'-TCACTAGTGTTATTACATCCTTGTGGCAC-3' and reverse,
5'-GAACTAGTGAAGTCATGCAAAACGCTCA-3'; GAPDH forward,
5'-ACAACTTTGGTATCGTGGAAGG-3' and reverse,
5'-GCCATCACGCCACAGTTTC-3'. Analysis of gene expression was
performed using the 2-IICq method (17). GAPDH was used as an internal
reference gene.
Chromatin immunoprecipitation (ChIP)
assay
The binding of IRF2 to the CLDN7 promoter was
examined using a ChIP assay with EZ-ChIPTM Chromatin
Immunoprecipitation Kit (Millipore), according to the
manufacturer's protocol. After fixing with formaldehyde (1%) for 10
min at room temperature to produce cross-linked protein and DNA,
cultured CAL-27 cells were incubated with 1X Buffer B for 10 min at
room temperature and incubated with ice-cold 1X Buffer C for a
further 10 min. Cells were subsequently incubated with the 1X
Buffer D/PI mix and chromatin fragments were obtained using
sonication (5 times for 10 sec each) on ice for 10 min. IRF2
antibody (cat. no. sc-374327; Santa Cruz Biotechnology, Inc.) was
utilized to generate immunoprecipitants, whereas an IgG antibody
(cat. no. sc-69786; Santa Cruz Biotechnology, CA, USA) was used as
the blank control group to exclude the influence of other factors.
The recuperated DNA fragments were evaluated via qPCR. The relative
levels of CLDN7 promoter was normalized according to the average
level of the IgG group.
Luciferase reporter assay
Luciferase reporter plasmids (Promega Corporation)
were constructed using wild-type and mutant 3' untranslated regions
of the CLDN7 promoter. Firefly luciferase activity represented the
primary reporter used to monitor the binding of proteins to cloned
target sequences. Renilla luciferase was regarded as a
control reporter for normalization. The luciferase reporter
plasmids and regulating factors were co-transfected into CAL-27
cells using Lipofectamine® 3000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). After 48 h, the luciferase
activities were quantified using the Dual-Luciferase Reporter assay
system (Promega Corporation).
Western blot analysis
For immunoblotting, total cellular extracts were
prepared using RIPA lysis buffer containing protease inhibitor
cocktail (Beyotime Institute of Biotechnology). Protein
concentration was determined using a bicinchoninic acid kit
(Beyotime Institute of Biotechnology). From each sample, 40 µg
protein was added and separated via SDS-PAGE on a 10% gel, and
separated proteins were subsequently transferred to a
nitrocellulose blotting membrane (Cytiva). Non-specific binding was
blocked by 5% skimmed milk for 1.5 h at room temperature and then
the membranes were incubated overnight at 4˚C with specific primary
antibodies. The following day, the membranes were incubated with
HRP-conjugated goat anti-rabbit secondary antibody (cat. no. 7074S;
Cell Signaling Technology, Inc.) for 1 h at room temperature.
Signals were visualized using the Odyssey Western Blot Analysis
system (LI-COR Biosciences). Anti-CLDN7 (cat. no. ab265583)
antibody was provided by Abcam, anti-IRF2 (cat. no. sc-374327) and
anti-GAPDH (cat. no. sc-47724) antibodies were purchased from Santa
Cruz Biotechnology, Inc. The relative intensity of each band was
semi-quantified using ImageJ. GAPDH was used as the loading
control.
Statistical analysis
All experiments were repeated independently three
times. Data are presented as the mean ± standard deviation.
Comparisons between two groups were performed using a Student's
t-test. A one-way ANOVA followed by a Tukey's post-hoc test was
used to assess differences between multiple groups. All analyses
were performed using GraphPad Prism version 6.0 (GraphPad Software,
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Bioinformatics
PROMO (alggen.lsi.upc.es/; version, 3.0.2) was utilized to
predict potential transcription factors of CLDN7, and JASPAR
(jaspar.genereg.net/; version, 2020) was
used to predict the potential binding sites between IRF2 and the
CLDN7 promoter.
Results
CLDN7 expression is significantly
downregulated in OSCC cell lines
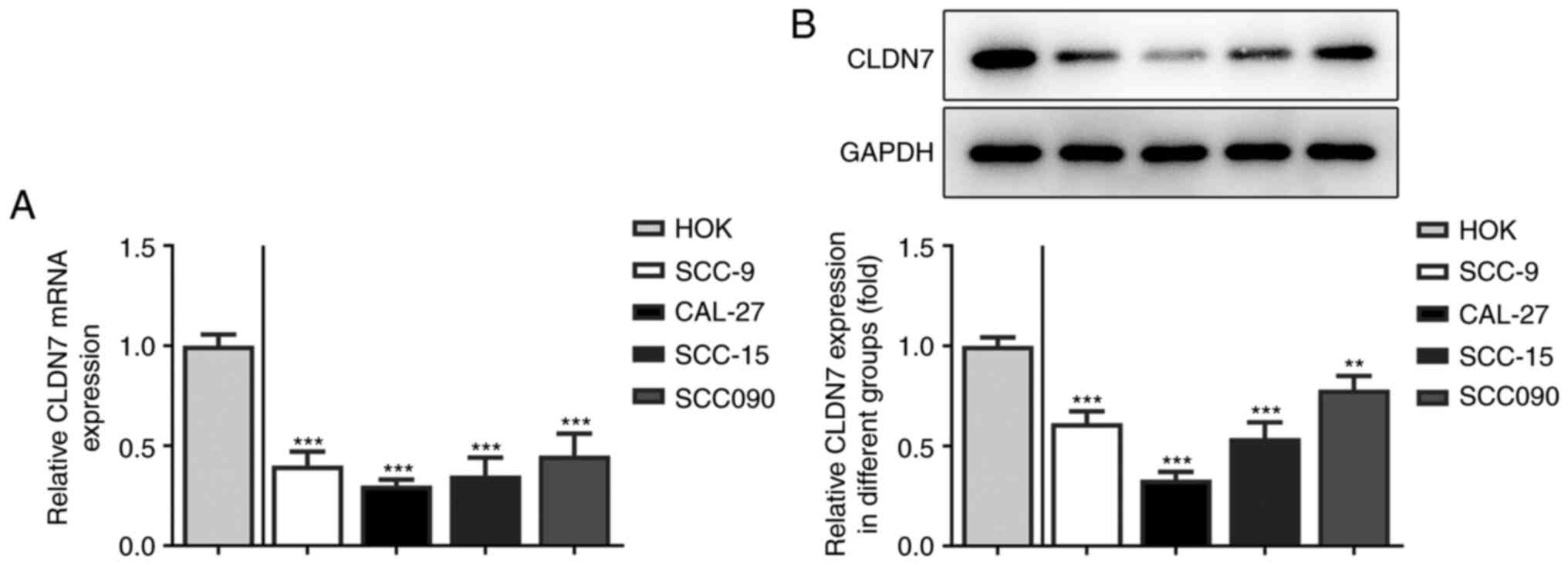
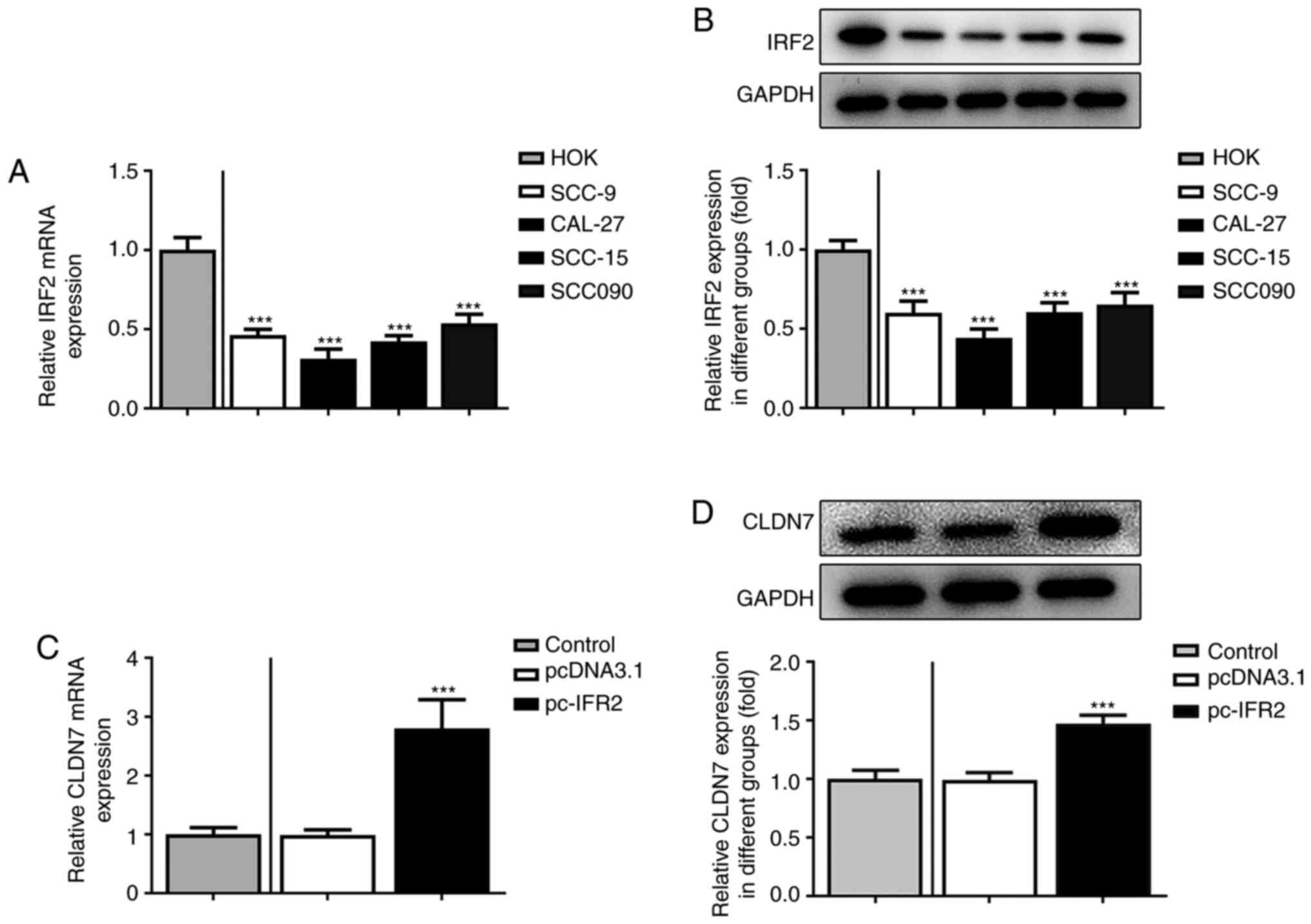
Firstly, the expression of CLDN7 in several OSCC
cell lines (SCC-9, CAL-27, SCC-15 and SCC090) was detected. As
shown in Fig. 1A and B, the expression levels of CLDN7 at the
mRNA and protein level were notably reduced in the OSCC cell lines
compared with the human oral keratinocyte HOK cell line. Lowest
CLDN7 expression was observed in the CAL-27 cells, therefore, this
cell line was used in subsequent experiments.
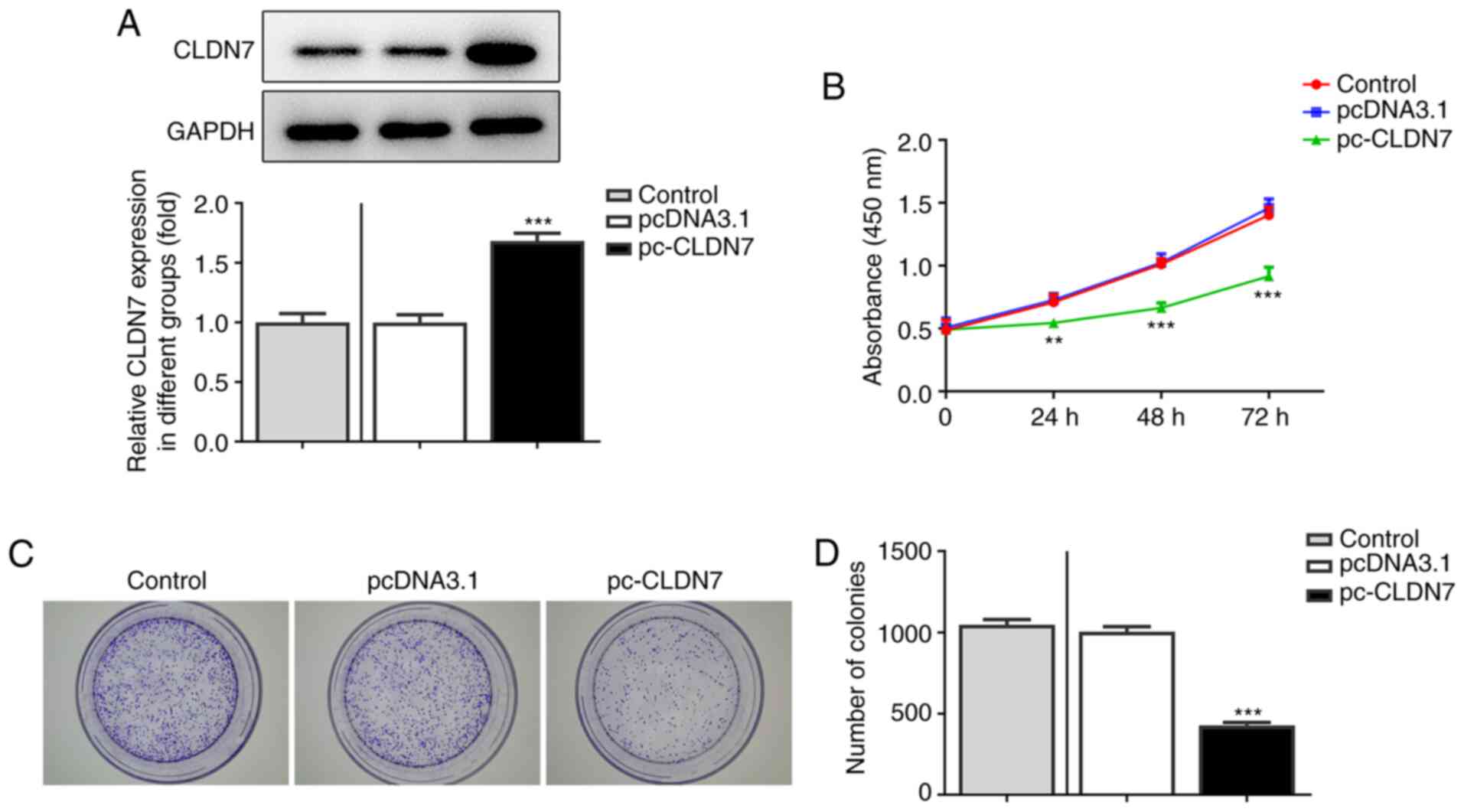
CLDN7 overexpression inhibits the
proliferation, invasion and migration of OSCC cells
To study the effects of CLDN7 on the progression of
OSCC, CLDN7 was overexpressed by transfection with a CLDN7 plasmid.
As shown in Fig. 2A, CLDN7
expression was notably upregulated after transfection compared with
the empty vector group. Next, the proliferative ability of CAL-27
cells was evaluated using CCK-8 and colony formation assays. It was
found that CLDN7 overexpression markedly suppressed cell viability
compared with the pcDNA3.1 group (Fig.
2B). Consistently, a significant decrease in the number of
colonies was observed after CLDN7 overexpression compared with the
vector control group (Fig. 2C and
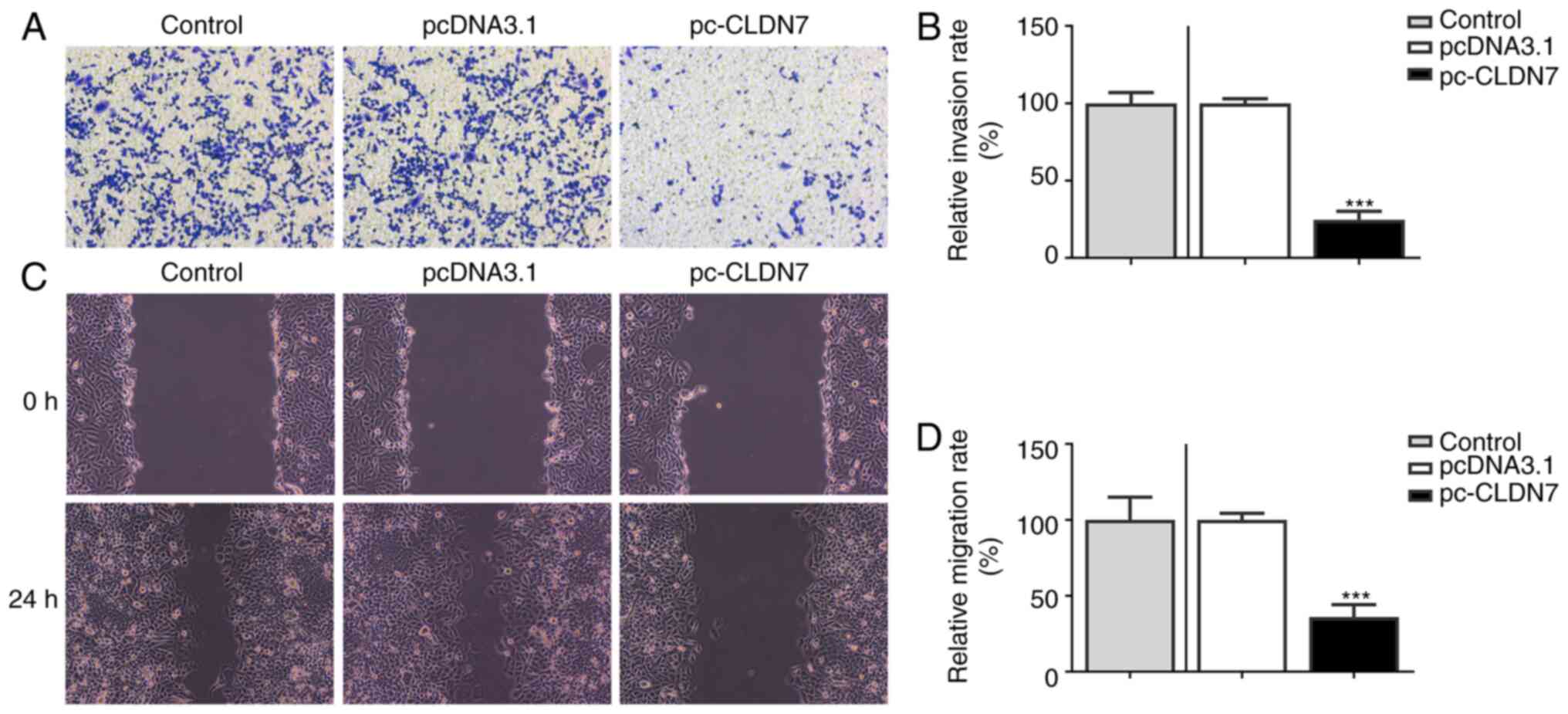
D). Subsequently, Transwell and
wound healing assays were used to evaluate cell invasion and
migration. As shown in Fig. 3A-D,
CLDN7 overexpression notably inhibited the invasion and migration
of CAL-27 cells. These data showed that overexpression of CLDN7
suppressed the proliferation, invasion and migration of OSCC
cells.
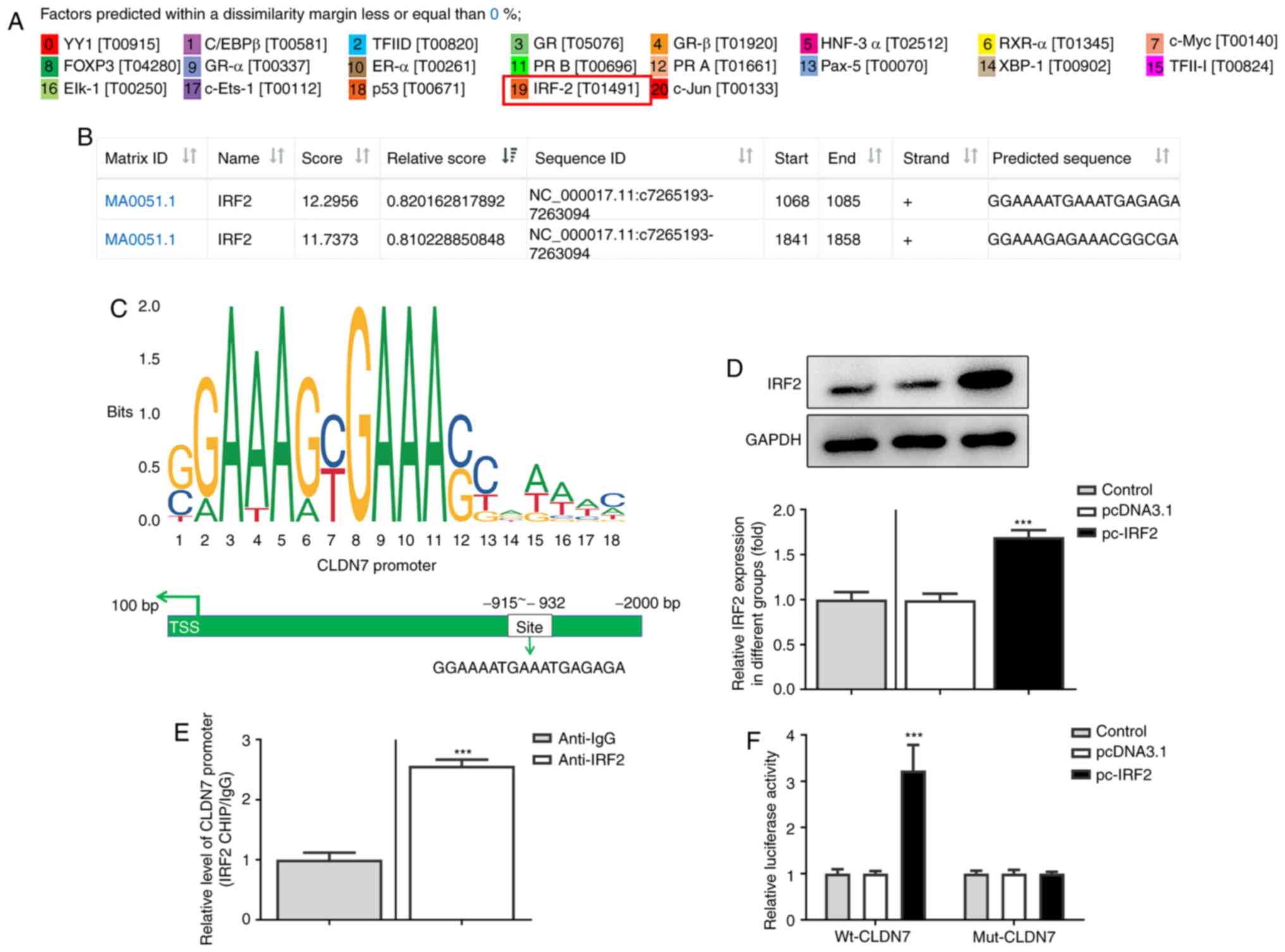
IRF2 regulates CLDN7 expression by
directly binding to the CLDN7 promoter
To further explore the molecular mechanism by which
CLDN7 regulated proliferation, invasion and migration of OSCC
cells, the potential transcription factor binding sites in the
promoter region of CLDN7 were predicted using PROMO and JASPAR
databases. IRF2 was predicted as a potential transcription factor
upstream of CLDN7, and the highest bound fraction between the IRF2
and CLDN7 promoter was confirmed using the JASPAR database
(Fig. 4A and B). As presented in Fig. 4C, one possible binding site of IRF2
was identified in the CLDN7 promoter (site-932 to -915). Next, IRF2
was overexpressed by transfection (Fig.
4D). In subsequent ChIP assays performed with extracts from
CAL-27 cells, notable enrichment of the CLDN7 promoter sequence was
obtained through immunoprecipitation with an anti-IRF2 antibody,
but not with the control IgG antibody (Fig. 4E). Moreover, the luciferase activity
of Wt-CLDN7 was activated by IRF2 overexpression (Fig. 4F). These observations showed that
IRF2 promoted CLDN7 transcription through directly binding to the
CLDN7 promoter region.
IRF2 overexpression promotes the
expression of CLDN7 in OSCC cells
Subsequently, the expression of IRF2 in several OSCC
cell lines was assessed using RT-qPCR and western blotting. As
shown in Fig. 5A and B, IRF2 mRNA and protein expression levels
were significantly downregulated in OSCC cell lines compared with
the HOK cells. Moreover, IRF2 was overexpressed by transfection
with the IRF2 plasmid in CAL-27 cells, and the increase in IRF2
expression is shown in Fig. 5C.
Subsequently, CLDN7 expression was markedly enhanced following IRF2
overexpression (Fig. 5D). Overall,
these data suggested that IRF2 overexpression promoted the
expression of CLDN7 in CAL-27 cells.
CLDN7 knockdown reverses the
inhibitory effects of IRF2 overexpression on the proliferation,
invasion and migration of CAL-27 cells
To further confirm the regulation of CLDN7 and IRF2,
function-related rescue experiments were performed. Firstly, CLDN7
expression was silenced by transfection with shCLDN7, which is
presented in Fig. 6A. As shown in
Fig. 6B-D, the proliferative
ability of CAL-27 cells was significantly inhibited following IRF2
overexpression, whereas this inhibitory effect was alleviated
following the knockdown of CLDN7 expression. Consistently, notably
suppressed invasion (Fig. 7A and
B) and migration (Fig. 7C and D) of CAL-27 cells was observed in the
pc-IRF2 group compared with the pcDNA3.1 group, which was restored
after co-transfection with pc-IRF2 and shCLDN7 into CAL-27
cells.
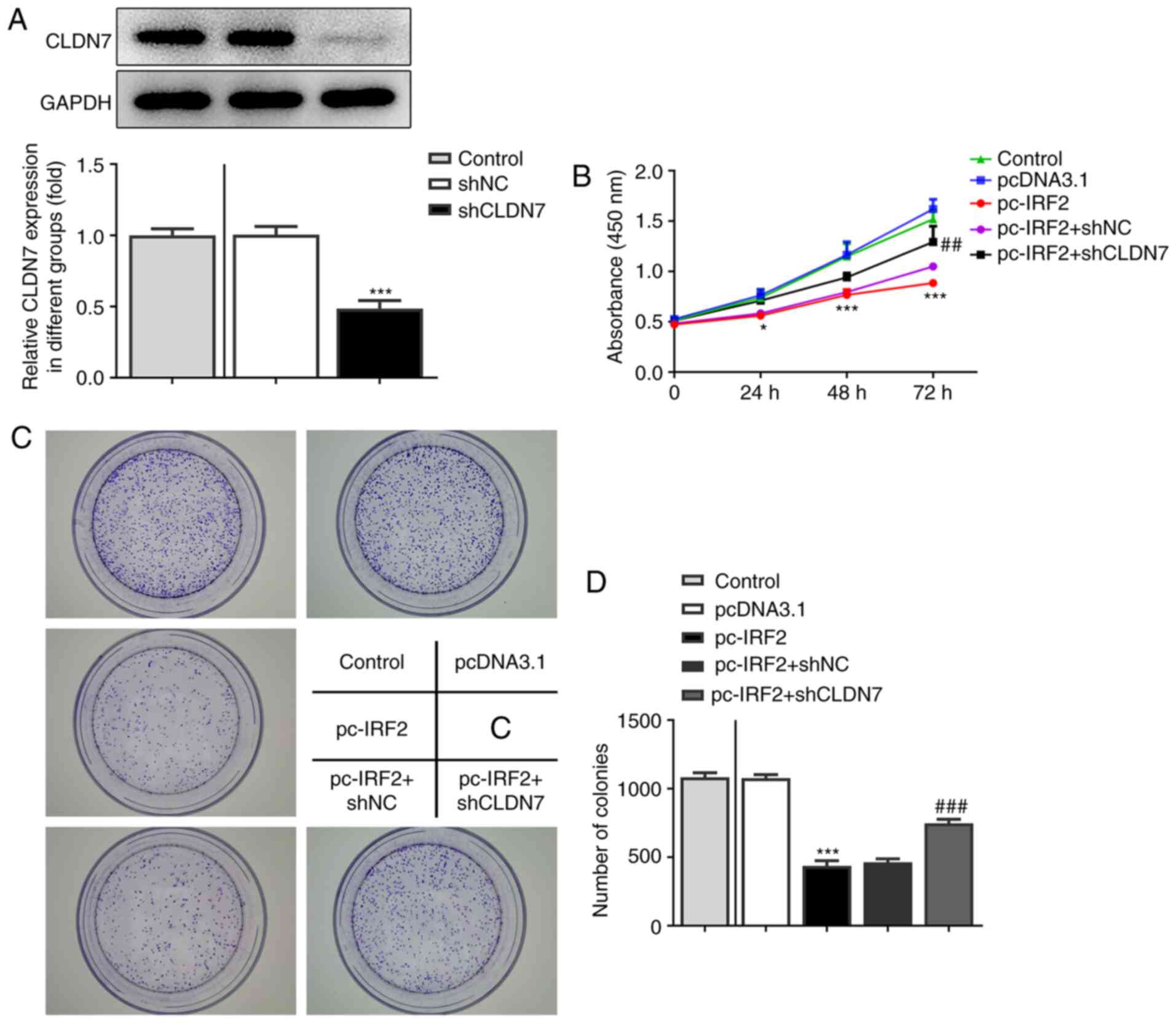 | Figure 6CLDN7 knockdown reverses the
inhibitory effects of IRF2 overexpression on cell proliferation in
OSCC cells. (A) Knockdown efficiency of CLDN7 was demonstrated
using western blotting. (B) A Cell Counting Kit-8 assay was used to
evaluate cell viability in the Control group, pcDNA3.1 group,
pc-IRF2 group, pc-IRF2 + shNC group and pc-IRF2 + shCLDN7 group. (C
and D) Colony formation assays were used to assess the number of
cell colonies in the Control group, pcDNA3.1 group, pc-IRF2 group,
pc-IRF2 + shNC group and pc-IRF2 + shCLDN7 group.
*P<0.05, ***P<0.001 vs. pcDNA3.1;
##P<0.01, ###P<0.001 vs. pc-IRF2 +
shNC. CLDN7, Claudin-7; IRF2, interferon regulatory factor-2; OSCC,
oral squamous cell carcinoma; sh, short hairpin RNA; NC, negative
control. |
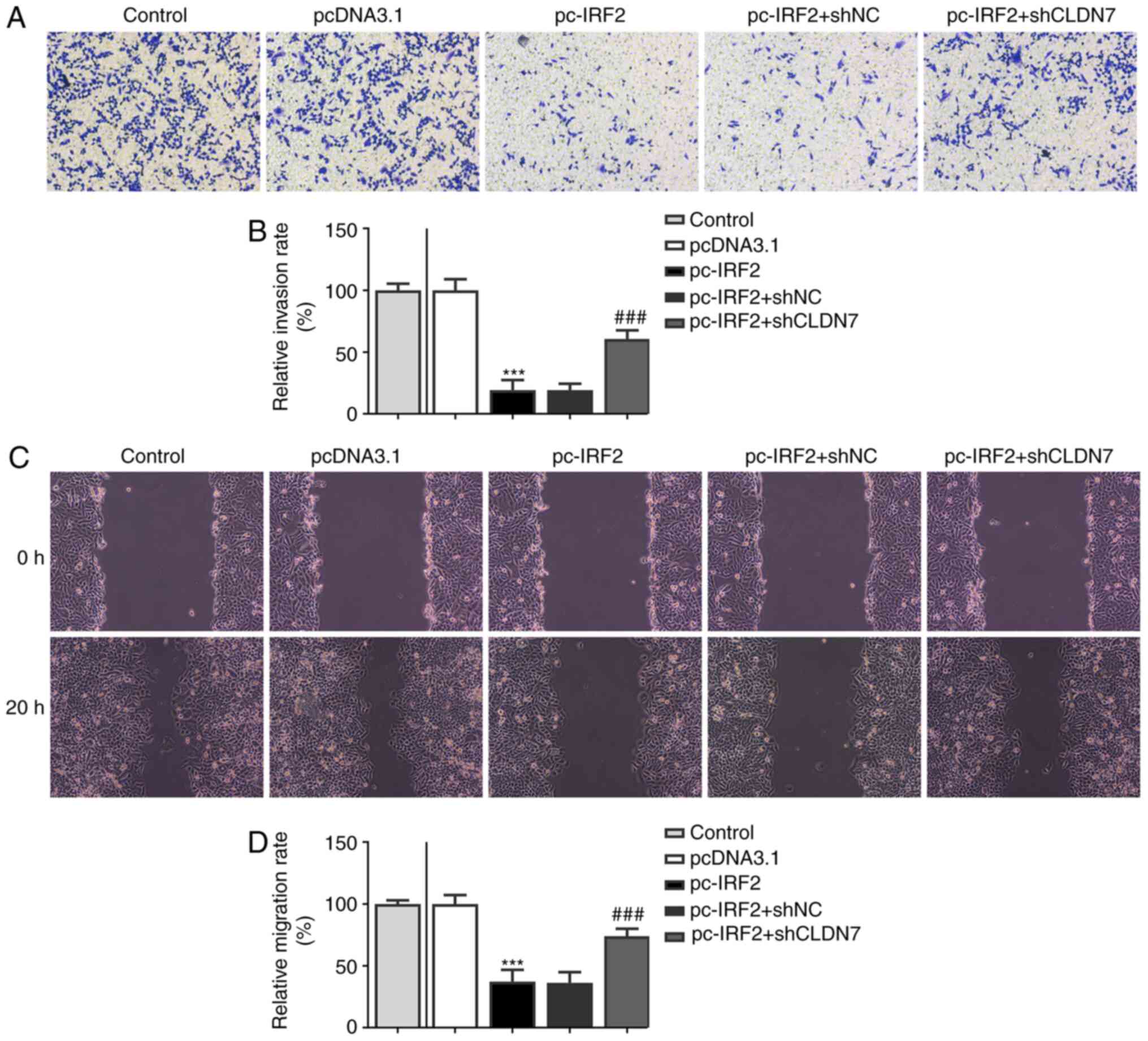 | Figure 7CLDN7 knockdown reverses the
inhibitory effects of IRF2 overexpression on cell migration and
invasion in OSCC cells. (A and B) Invasive ability of CAL-27 cells
in the Control group, pcDNA3.1 group, pc-IRF2 group, pc-IRF2 + shNC
group and pc-IRF2 + shCLDN7 group was measured using a Transwell
assay. (C and D) Migration rates of CAL-27 cells in the Control
group, pcDNA3.1 group, pc-IRF2 group, pc-IRF2 + shNC group and
pc-IRF2 + shCLDN7 group was detected using a wound healing assay.
***P<0.001 vs. pcDNA3.1; ###P<0.001 vs.
pc-IRF2 + shNC. CLDN7, Claudin-7; IRF2, interferon regulatory
factor-2; OSCC, oral squamous cell carcinoma; sh, short hairpin
RNA; NC, negative control. |
Discussion
OSCC is one of the most aggressive neoplasms amongst
head and neck malignant tumors, and is associated with a poor
prognosis (18). Improving our
understanding of the underlying mechanisms of the development and
progression of OSCC is critical for identifying effective targets
with therapeutic potential to improve the survival rates of
patients with OSCC. The primary aim of the present study was to
investigate the role of CLDN7 in the progression of OSCC. It The
results showed that CLDN7 expression was significantly decreased in
OSCC cells, and CLDN7 overexpression notably inhibited the
proliferation, invasion and migration of OSCC cells, which was
directly induced by the IRF2 transcription factor.
Proliferation and metastasis are both hallmarks of
the malignant biological behavior of OSCC, and the inhibition of
these processes is a crucial factor to improve biomedical treatment
worldwide (19). CLDN7, a member of
the Claudin family, is reported to be a tumor-related gene. At
present, it is known that CLDN7 plays either tumor promoting or
tumor suppressing roles based on the specific type of tumor. A
growing body of literature has shown that the loss of CLDN7 is
closely associated with a poor prognosis in nasopharyngeal cancer,
ovarian carcinoma and esophageal squamous cell carcinoma (8,20,21).
CLDN7 silencing induces invasion and metastasis in colorectal
cancer via the promotion of epithelial-mesenchymal transition
(6). Dysregulation of CLDN7 results
in loss of E-Cadherin expression and the enhanced invasion of
esophageal squamous cell carcinoma cells (22). Conversely, CLDN7 is notably elevated
in cervical and gastric cancer (23,24).
CLDN7 suppresses proliferation and metastasis in salivary adenoid
cystic carcinoma (9). Of note, loss
of CLDN7 expression may be associated with the pathogenesis of OSCC
and this loss is associated with a poor prognosis (25). Loss of CLDN7 is considered a
negative prognostic factor for invasion and metastasis in OSCC
(10). However, the exact
regulatory mechanisms by which CLDN7 is regulated in OSCC remains
unknown. The current study revealed that CLDN7 was downregulated in
OSCC cell lines, and CLDN7 overexpression markedly inhibited the
proliferation, invasion and migration of OSCC cells, suggesting the
potential tumor suppressor role of CLDN7 in OSCC.
According to PROMO and JASPAR database analyses,
IRF2 could potentially bind to the promoter region of CLDN7. IRF2,
one of the members of the IRF family of transcription factors, can
transcriptionally induce multiple direct target genes, which has
been identified to participate in the regulation of immune
responses and immune cell development (26-28).
An increasing number of studies have confirmed that IRF2 is closely
related to the physiological and pathological processes of various
types of cancer (13,29). Existing reports have shown that
IRF-2 inhibits the invasion and migration of gastric cancer through
downregulation of MMP-1 expression (15). MicroRNA-664 suppresses the
progression of cutaneous squamous cell carcinoma via inhibiting
IRF2 expression (30). IRF2 serves
an inhibitory effect on the invasion and migration of osteosarcoma,
where it is regulated by microRNA-18a-5p (31). Of note, IRF2 has been reported to
bind to inositol polyphosphate 4-phosphatase type II (INPP4B)
promoter and increase INPP4B expression, thereby participating in
the development of acute myeloid leukemia (32). In the present study, significantly
downregulated levels of IRF2 expression were observed in OSCC
cells, and overexpression of IRF2 significantly suppressed the
proliferation, invasion and migration of OSCC cells. Furthermore,
the binding effect of IRF2 to the promoter of CLDN7 was confirmed
via ChIP and luciferase reporter assays. The rescue experiments
demonstrated that CLDN7 silencing blocked the inhibitory effects of
IRF2 overexpression on the proliferation, invasion and migration of
OSCC cells.
In summary, the present study found that both CLDN7
and IRF2 expression levels were downregulated in OSCC cells. IRF2
was determined to have a direct regulatory effect on the
transcription of CLDN7, and the biological functions of CLDN7 in
OSCC cells were closely regulated by the transcriptional factor
IRF2. These findings provided evidence for a novel regulatory
mechanism involving CLDN7 and IRF2 in OSCC, thus potentially
identifying novel targets for therapy. However, the use of only one
OSCC cell line to clarify the effects of CLDN7 and IRF2 in OSCC is
a potential limitation of the present study. Subsequent experiments
will incorporate more typical OSCC cell lines to support this
conclusion.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and WY designed the experiments. XL analyzed the
experimental data and wrote the manuscript. WY helped to correct
the manuscript. XL carried out the experiments. XL and WY confirmed
the authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sasahira T and Kirita T: Hallmarks of
cancer-related newly prognostic factors of oral squamous cell
carcinoma. Int J Mol Sci. 19(2413)2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Sequeira I, Neves JF, Carrero D, Peng Q,
Palasz N, Liakath-Ali K, Lord GM, Morgan PR, Lombardi G and Watt
FM: Immunomodulatory role of Keratin 76 in oral and gastric cancer.
Nat Commun. 9(3437)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Li YY, Tao YW, Gao S, Li P, Zheng JM,
Zhang SE, Liang J and Zhang Y: Cancer-associated fibroblasts
contribute to oral cancer cells proliferation and metastasis via
exosome-mediated paracrine miR-34a-5p. EBioMedicine. 36:209–220.
2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Gao D, Xu T, Qi X, Ning W, Ren S, Ru Z, Ji
K, Ma Y, Yu T, Li Y, et al: CLAUDIN7 modulates trophectoderm
barrier function to maintain blastocyst development in pigs.
Theriogenology. 158:346–357. 2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Wang K, Li T, Xu C, Ding Y, Li W and Ding
L: Claudin-7 downregulation induces metastasis and invasion in
colorectal cancer via the promotion of epithelial-mesenchymal
transition. Biochem Biophys Res Commun. 508:797–804.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Poon CE, Madawala RJ, Day ML and Murphy
CR: Claudin 7 is reduced in uterine epithelial cells during early
pregnancy in the rat. Histochem Cell Biol. 139:583–593.
2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Romani C, Zizioli V, Silvestri M,
Ardighieri L, Bugatti M, Corsini M, Todeschini P, Marchini S,
D'Incalci M, Zanotti L, et al: Low expression of Claudin-7 as
potential predictor of distant metastases in high-grade serous
ovarian carcinoma patients. Front Oncol. 10(1287)2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ji H, Ding X, Zhang W, Zheng Y, Du H,
Zheng Y, Song H, Li M, Jiang Y, Xie J, et al: Claudin-7 inhibits
proliferation and metastasis in salivary adenoid cystic carcinoma
through Wnt/β-catenin signaling. Cell Transplant.
29(963689720943583)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yoshizawa K, Nozaki S, Kato A, Hirai M,
Yanase M, Yoshimoto T, Kimura I, Sugiura S, Okamune A, Kitahara H,
et al: Loss of claudin-7 is a negative prognostic factor for
invasion and metastasis in oral squamous cell carcinoma. Oncol Rep.
29:445–450. 2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wang Y, Liu DP, Chen PP, Koeffler HP, Tong
XJ and Xie D: Involvement of IFN regulatory factor (IRF)-1 and
IRF-2 in the formation and progression of human esophageal cancers.
Cancer Res. 67:2535–2543. 2007.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Cui L, Deng Y, Rong Y, Lou W, Mao Z, Feng
Y, Xie D and Jin D: IRF-2 is over-expressed in pancreatic cancer
and promotes the growth of pancreatic cancer cells. Tumour Biol.
33:247–255. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kneitz B, Krebs M, Kalogirou C, Schubert
M, Joniau S, van Poppel H, Lerut E, Kneitz S, Scholz CJ, Ströbel P,
et al: Survival in patients with high-risk prostate cancer is
predicted by miR-221, which regulates proliferation, apoptosis, and
invasion of prostate cancer cells by inhibiting IRF2 and SOCS3.
Cancer Res. 74:2591–2603. 2014.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Pettersson S, Kelleher M, Pion E, Wallace
M and Ball KL: Role of Mdm2 acid domain interactions in recognition
and ubiquitination of the transcription factor IRF-2. Biochem J.
418:575–585. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chen YJ, Liang L, Li J, Wu H, Dong L, Liu
TT and Shen XZ: IRF-2 inhibits gastric cancer invasion and
migration by down-regulating MMP-1. Dig Dis Sci. 65:168–177.
2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Tian JB, Cao L and Dong GL: Long noncoding
RNA DDX11-AS1 induced by YY1 accelerates colorectal cancer
progression through targeting miR-873/CLDN7 axis. Eur Rev Med
Pharmacol Sci. 23:5714–5729. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Binmadi NO and Basile JR: Perineural
invasion in oral squamous cell carcinoma: A discussion of
significance and review of the literature. Oral Oncol.
47:1005–1010. 2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Takahara T, Kasamatsu A, Yamatoji M, Iyoda
M, Kasama H, Saito T, Takeuchi S, Endo-Sakamoto Y, Shiiba M,
Tanzawa H and Uzawa K: SIPA1 promotes invasion and migration in
human oral squamous cell carcinoma by ITGB1 and MMP7. Exp Cell Res.
352:357–363. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Suren D, Yildirim M, Kaya V, Elal R,
Selcuk OT, Osma U, Yildiz M, Gunduz S and Sezer C: Expression
patterns of claudins 1, 4, and 7 and their prognostic significance
in nasopharyngeal carcinoma. J BUON. 20:212–217. 2015.PubMed/NCBI
|
|
21
|
Usami Y, Chiba H, Nakayama F, Ueda J,
Matsuda Y, Sawada N, Komori T, Ito A and Yokozaki H: Reduced
expression of claudin-7 correlates with invasion and metastasis in
squamous cell carcinoma of the esophagus. Hum Pathol. 37:569–577.
2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lioni M, Brafford P, Andl C, Rustgi A,
El-Deiry W, Herlyn M and Smalley KS: Dysregulation of claudin-7
leads to loss of E-cadherin expression and the increased invasion
of esophageal squamous cell carcinoma cells. Am J Pathol.
170:709–721. 2007.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhang B, Lin Y, Bao QF, Zheng YT and Lan
L: MiR-1193 inhibits the malignancy of cervical cancer cells by
targeting claudin 7 (CLDN7). OncoTargets Ther. 13:4349–4358.
2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wu Z, Shi J, Song Y, Zhao J, Sun J, Chen
X, Gao P and Wang Z: Claudin-7 (CLDN7) is overexpressed in gastric
cancer and promotes gastric cancer cell proliferation, invasion and
maintains mesenchymal state. Neoplasma. 65:349–359. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Lourenço SV, Coutinho-Camillo CM, Buim ME,
de Carvalho AC, Lessa RC, Pereira CM, Vettore AL, Carvalho AL,
Fregnani JH, Kowalski LP and Soares FA: Claudin-7 down-regulation
is an important feature in oral squamous cell carcinoma.
Histopathology. 57:689–698. 2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Tamura T, Yanai H, Savitsky D and
Taniguchi T: The IRF family transcription factors in immunity and
oncogenesis. Annu Rev Immunol. 26:535–584. 2008.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Sun L, Jiang Z, Acosta-Rodriguez VA,
Berger M, Du X, Choi JH, Wang J, Wang KW, Kilaru GK, Mohawk JA, et
al: HCFC2 is needed for IRF1- and IRF2-dependent Tlr3 transcription
and for survival during viral infections. J Exp Med. 214:3263–3277.
2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhao GN, Jiang DS and Li H: Interferon
regulatory factors: At the crossroads of immunity, metabolism, and
disease. Biochim Biophys Acta. 1852:365–378. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Sakai T, Mashima H, Yamada Y, Goto T, Sato
W, Dohmen T, Kamada K, Yoshioka M, Uchinami H, Yamamoto Y and
Ohnishi H: The roles of interferon regulatory factors 1 and 2 in
the progression of human pancreatic cancer. Pancreas. 43:909–916.
2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Li X, Zhou C, Zhang C, Xie X, Zhou Z, Zhou
M, Chen L and Ding Z: MicroRNA-664 functions as an oncogene in
cutaneous squamous cell carcinomas (cSCC) via suppressing
interferon regulatory factor 2. J Dermatol Sci. 94:330–338.
2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Lu C, Peng K, Guo H, Ren X, Hu S, Cai Y,
Han Y, Ma L and Xu P: miR-18a-5p promotes cell invasion and
migration of osteosarcoma by directly targeting IRF2. Oncol Lett.
16:3150–3156. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhang F, Zhu J, Li J, Zhu F and Zhang P:
IRF2-INPP4B axis participates in the development of acute myeloid
leukemia by regulating cell growth and survival. Gene. 627:9–14.
2017.PubMed/NCBI View Article : Google Scholar
|