Introduction
Oral squamous cell carcinoma (OSCC) is the most
frequent type of head and neck cancer in China (1). The current treatment modalities for
OSCC include surgery, which can be functionally debilitating or
disfiguring, radiotherapy and chemotherapy. For patients with
terminal or advanced cancer, the survival times may be decreased to
a few months, which highlights the urgent need for novel
therapeutic strategies (2,3).
Curcumin [Cur; 1,7-bis(4-hydroxy-3-methoxyphenyl)-1,
6-heptadiene-3,5-dione] is a natural phenolic compound commonly
known as the dietary spice turmeric (Curcuma longa), which
is derived from the rhizome of an East Indian plant (4). Cur has been considered as
pharmacologically safe for dietary consumption over several
centuries (5). Extensive research
has revealed that Cur is an effective molecular agent with a broad
spectrum of biological activities against aging, inflammation and
cancer (6,7). Previous studies have reported that Cur
inhibits proliferation, metastasis and initiation of several
malignances, including lung, breast, hepatocellular, pancreatic and
gastric cancers (8-12).
In addition, epidemiological study has suggested that a diet rich
in Cur may decrease the incidence of colon cancer (13).
Cur significantly inhibits the activity of the
nuclear factor κB (NF-κB) pathway in various human carcinomas, such
as liver and thyroid carcinoma (14,15).
Cur may therefore, be considered as an inhibitor of NF-κB by
regulating the proteins associated with the NF-κB pathway. It was
demonstrated that Cur can decrease the expression of p65, which
specifically targets cancer stem cell populations and kills liver
cancer cells (16). Under hypoxic
tumor microenvironment, Cur can effectively inhibit
hypoxia-inducible factor 1α (HIF-1α) and nuclear p65 expression in
breast and lung cancer cells, which leads to tumor growth
inhibition (10). In addition, Cur
can inhibit NF-κB activity in an AKT-dependent manner in head and
neck squamous cell carcinoma (17).
Specificity protein 1 (Sp1) is a member of the
Sp-family, which is involved in multiple biological processes,
including cell apoptosis, differentiation, cell cycle progression
and proliferation (18-20).
The expression and activation of Sp1 have been reported to be
associated with the development of human cancers and prognosis
(21). Sp1 is overexpressed in
several cancers, including pancreatic and gastric cancers, and
appears to be associated with poor prognosis, suggesting that Sp1
could be a potential therapeutic strategy for cancer treatment
(22,23). Sp1 is also associated with
proliferation, apoptosis and metastasis in oral cancer. Suppression
of the tissue inhibitor of metalloproteinase-3 by promoter
methylation of Sp1 contributes to oral cancer metastasis (24). Interfering Sp1 with small
interfering (si)RNA or mithramycin A increasingly trigger apoptosis
in oral cancer cells (25). A
series of Sp1 special target inhibitors were reported to be
effective for OSCC treatment, including honokiol, β-lapachone and
esculetin (26-29).
In addition, Sp1 can regulate NF-κB pathway activity by increasing
the transcriptional level of p65 and p50 in pancreatic cancer and
OSCC (30,31).
The present study aimed to investigate the effects
and underlying mechanisms of Cur in OSCC. The results indicated
that Cur inhibited the proliferation and NF-κB activity of OSCC
cells. Importantly, Cur decreased the expression of Sp1, p65 and
HSF1 in OSCC cells. Finally, Sp1 knockdown contributed to the
effect of Cur on p65 and HSF1 in OSCC cells, resulting in a
decreased NF-κB activity and cell viability.
Materials and methods
Cell lines and reagents
HSC3 and CAL33 cell lines were used in the present
study. The HSC3 cell line was kindly provided by Dr Xin Zhang
(University of Wuhan). The CAL33 cell line was a gift from Dr Juhua
Zhou (Hubei University of Medicine). All cells were cultured in
DMEM (HyClone; Cytiva) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) and 1% antibiotic
(penicillin/streptomycin; Sigma-Aldrich; Merck KGaA) and placed at
37˚C in a humidified incubator containing 5% CO2. Cur
(Sigma-Aldrich; Merck KGaA) was dissolved in DMSO (Sigma-Aldrich;
Merck KGaA) as a stock solution of 100 mM.
Cell viability assay
Previous studies have used 5-20 µM Cur as a work
dose in experiments on OSCC and other malignances (32,33).
In the present study, HSC3 and CAL33 cells were seeded in 96-well
plates (1x104 cells/well) and treated with 0.01% DMSO
(control) or increasing concentrations of Cur (5-20 µM) at 37˚C.
After 24 or 48 h treatment, cells were incubated with 10 µl Cell
Counting Kit-8 reagent (Dojindo Molecular Technologies, Inc.) for 1
h at 37˚C in the dark. The optical density was measured using an
ELx800 microimmunoanalyser at 450 nm (BioTek Instruments, Inc.).
Data were normalized to the control sample.
Colony formation assay
HSC3 and CAL33 (500 cells) were seeded in 35-mm
culture dishes and grown in standard medium with DMSO (0.01%) or
Cur (0.5-2 µM). After 14 days of treatment, cells were incubated
with 0.5% crystal violet-glutaraldehyde solution (0.6%, v/v) for 1
min. After washing with PBS, cell colonies were photographed
(34,35).
Western blotting
HSC3 and CAL33 cells were treated with DMSO (0.01%)
or Cur (5, 10 and 20 µM) for 48 h. Cells were washed, harvested and
lysed in RIPA lysis buffer (Beyotime Institute of Biotechnology)
containing 0.5% cocktail protease inhibitor (Roche Diagnostics) on
ice for 10 min, collected and sonicated for 15 sec. Protein
concentration was measured by BCA assay (Bio-Rad Laboratories,
Inc.). Proteins were mixed with 5X loading buffer (Chunfeng Lv,
China) and boiled for 5 min. Proteins (20 µg) were separated by 10%
SDS-PAGE and transferred onto PVDF membranes. After blocking with
5% skimmed milk in PBS supplemented with 0.01% Tween-20 for 1 h at
room temperature, membranes were incubated with primary antibodies
against β-actin (cat. no. A-1978; 1:2,000; Sigma-Aldrich; Merck
KGaA); Sp1 (cat. no. ab124804; 1:1,000; Abcam); p65 (cat. no.
ab16502; 1:1,500; Abcam) and HSF1 (cat. no. 12972; 1:1,000; Cell
Signaling Technology, Inc.) overnight at 4˚C. After washing three
times with PBS-0.01% Tween-20, membranes were incubated with
HRP-conjugated goat anti-rabbit (cat. no. A9169; 1:2,000) or goat
anti-mouse (cat. no. A9309; 1:2,000; both from Sigma-Aldrich; Merck
KGaA) secondary antibodies. An ECL kit (cat. no. 1705060; Bio-Rad
Laboratories, Inc.) was used to detect the signal. Western blot
gray values were determined using ImageJ software (version 1.8.0;
National Institutes of Health).
Reverse transcription-quantitative
(RT-q)PCR
HSC3 and CAL33 cells (1x105 cells/well)
were seeded in 24-well plates overnight. Cells were washed with
PBS, treated with DMSO (0.01%) or Cur (10 or 20 µM) and incubated
at 37˚C for 24 h. Cells were washed with PBS and total RNA was
extracted using 0.5 ml TRIzol reagent (Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. Subsequently,
RNA was then reverse transcribed into cDNA using RT kit (Takara
Bio, Inc.) according to the manufacturer's instructions. cDNA
levels were quantified using the SYBR Premix Ex Taq kit (Takara
Bio, Inc.) with the CFX96 Real-Time PCR Detection System. The
thermocycling conditions were as follows: Denaturation at 95˚C for
15 sec, annealing at 60˚C for 15 sec and extension at 72˚C for 15
sec. The replication cycles were repeated 30 times. The sequences
of the primers were as follows: p65, forward, 5'-CGGGATGGCTTCTATGA
GG-3' and reverse, 5'-CTCCAGGTCCCGCTTCTT-3'; HSF1, forward,
5'-ACCTTCATCGGAAACTCCAAAG-3' and reverse,
5'-CTGTTAGGCTGGGAAAAGTTAGG-3'; Sp1, forward,
5'-GGAGAGCAAAACCAGCAGAC-3' and reverse, 5'-AAGGTGATTGTTTGGGCTTG-3';
and GAPDH, forward, 5'-AGGTCGGTGTGAACGGATTTG-3' and reverse,
5'-TGTAGACCATGTAGTTGAGGTCA-3'. The relative expression levels were
normalized to endogenous control and were expressed as
2-ΔΔCq (36).
Cell transfection
HSC3 cells were transfected with Sp1-specific short
hairpin RNA (shRNA; 5'-GCATATTTGCCACATCCAAGG-3', Sp1-Homo-1828;
GenePharma, Co., Ltd.) or a non-specific control (NC;
5'-TTCTCCGAACGTGTCACGT-3'; Shanghai GenePharma Co., Ltd.) using
X-treme GENE HP DNA Transfection Reagent (Roche Diagnostics; 40
pmol for each shRNA) according to the manufacturer's instructions.
Furthermore, PCDNA3.1 control vector and pCDNA3.1-Sp1 plasmid were
donated by Dr Zilong Gao (Taihe Hospital, Shiyan). HSC3 cells were
transfected with pCDNA3.1 and pCDNA3.1-Sp1 (2 µg) by using X-treme
GENE HP DNA Transfection Reagent. At 4 h following transfection,
Cur or DMSO control was added. Cells were incubated and harvested
for the next experiments as follows: Western blotting (48 h),
Dual-Luciferase reporter assay (12 h) and Cell Counting Kit-8 assay
(24 h).
Dual-Luciferase reporter assay
The NF-κB activity plasmid and PRL-TK plasmid were
kindly provided by Professor H. Shu (University of Wuhan). HSC3
cells (1x105 cells/well) were seeded into 24-well plates
and incubated at 37˚C overnight. Plasmids were co-transfected using
X-treme GENE HP DNA Transfection Reagent into the cells for 24 h.
Cells were treated with Cur (10 or 20 µM) or DMSO (0.01%) for 12 or
24 h, then collected and analyzed using the Dual-Luciferase
reporter assay system according to the manufacturer's instructions
(Promega Corporation). The intensity of NF-κB activity was
normalized to the Renilla luciferase activity from the
PRL-TK plasmid.
Statistical analysis
Experimental data were presented as the mean ±
standard deviation of three independent experiments. Student's
t-test was used for comparing significance between two groups.
One-way ANOVA followed by Newman-Keuls or Bonferroni post hoc tests
was used to compare data between >2 groups. Statistical analysis
was performed using SPSS version 12.0 (SPSS, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
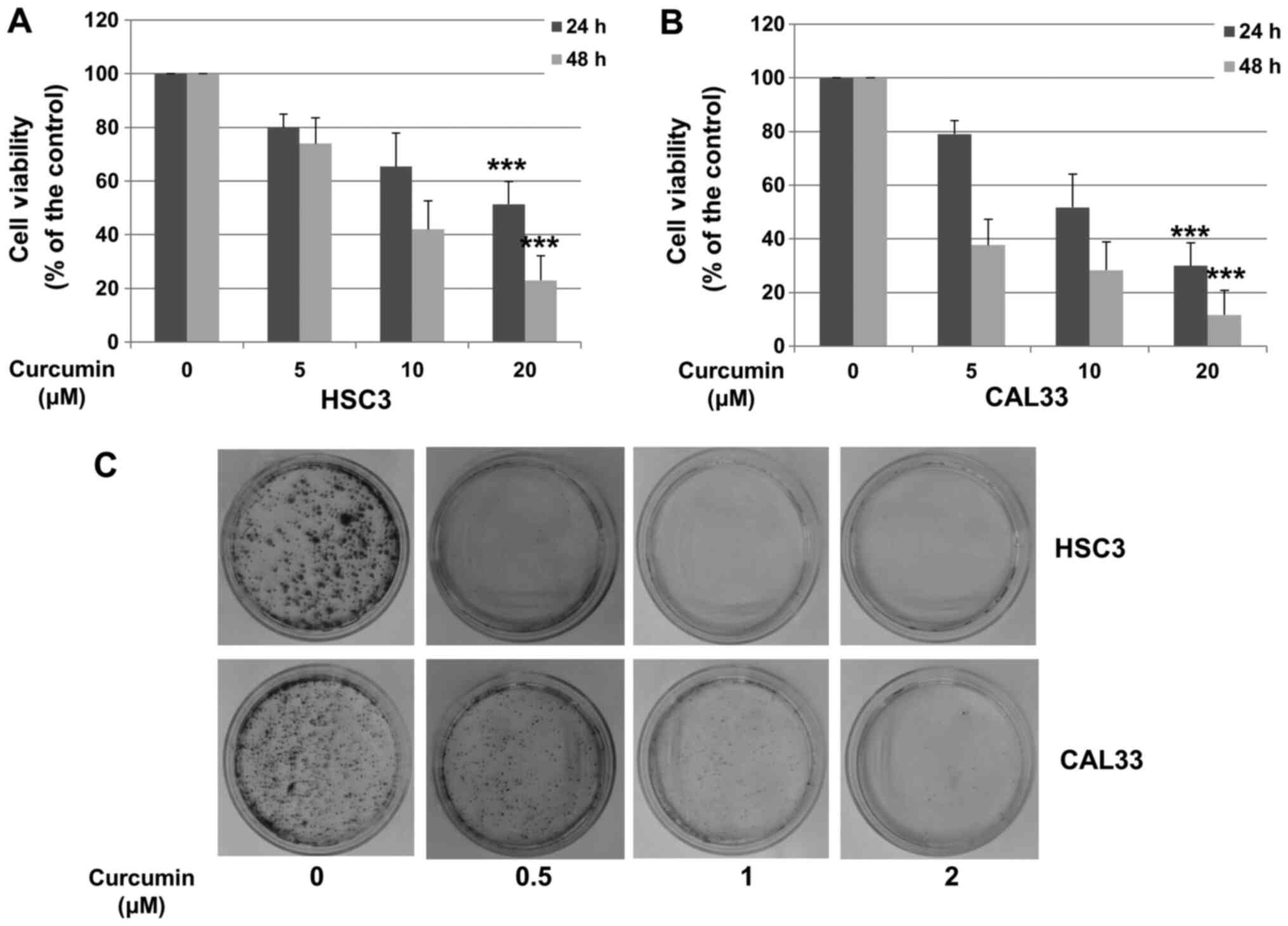
Cur inhibits OSCC cell
proliferation
To determine the effect of Cur on the viability of
OSCC cells, HSC3 and CAL33 cells were treated with various doses of
Cur for 24 or 48 h. Cell growth inhibition was compared with DMSO
control alone. As presented in Fig.
1A and B, Cur significantly
decreased the viability of HSC3 and CAL33 cells in a
concentration-dependent manner (20 µM; P<0.001). A colony
formation assay was performed to further confirm these results.
HSC3 and CAL33 cells were treated with DMSO or Cur for 14 days and
subjected to crystal violet staining. As presented in Fig. 1C, Cur significantly inhibited the
colony formation of both HSC3 and CAL33 cells. These results
suggested that Cur inhibited the proliferation of HSC3 and CAL33
cell lines.
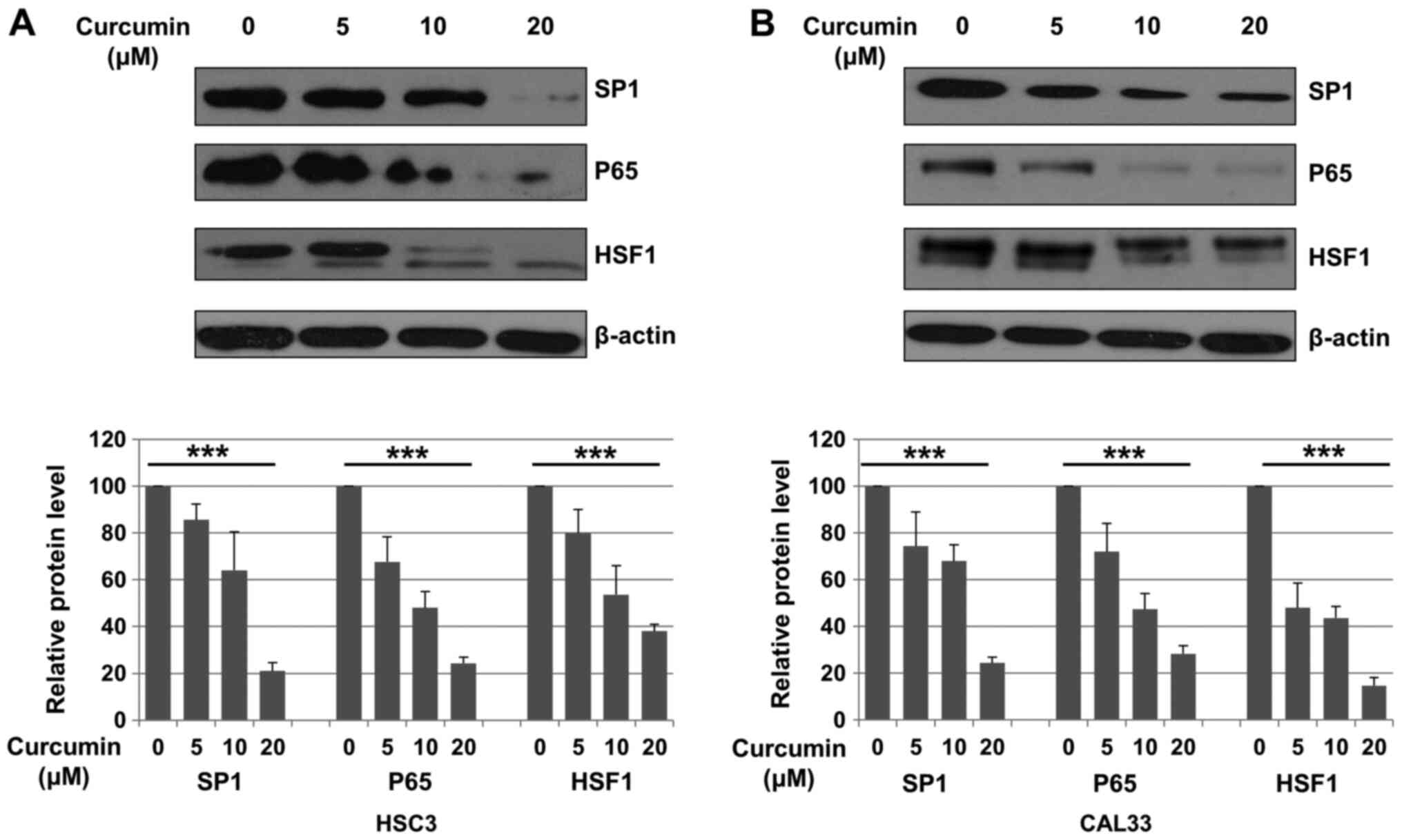
Cur downregulates Sp1, p65 and HSF1
expression in OSCC cells
The expression of Sp1, p65 and HSF1 following
treatment with DMSO (control) or Cur (5, 10 and 20 µM) for 48 h was
evaluated in HSC3 and CAL33 cells. Compared with control, Sp1
expression was significantly decreased after Cur treatment in both
HSC3 and CAL33 cells (P<0.001; Fig.
2A and B). It has been reported
that Sp1 is a transcription factor of p65 and HSF1(31). Subsequently, the expression of p65
and HSF1 was also evaluated. As presented in Fig. 2A and B, Cur treatment significantly decreased
the expression of p65 (P<0.001) and HSF1 (P<0.001) in OSCC
cells compared with the control.
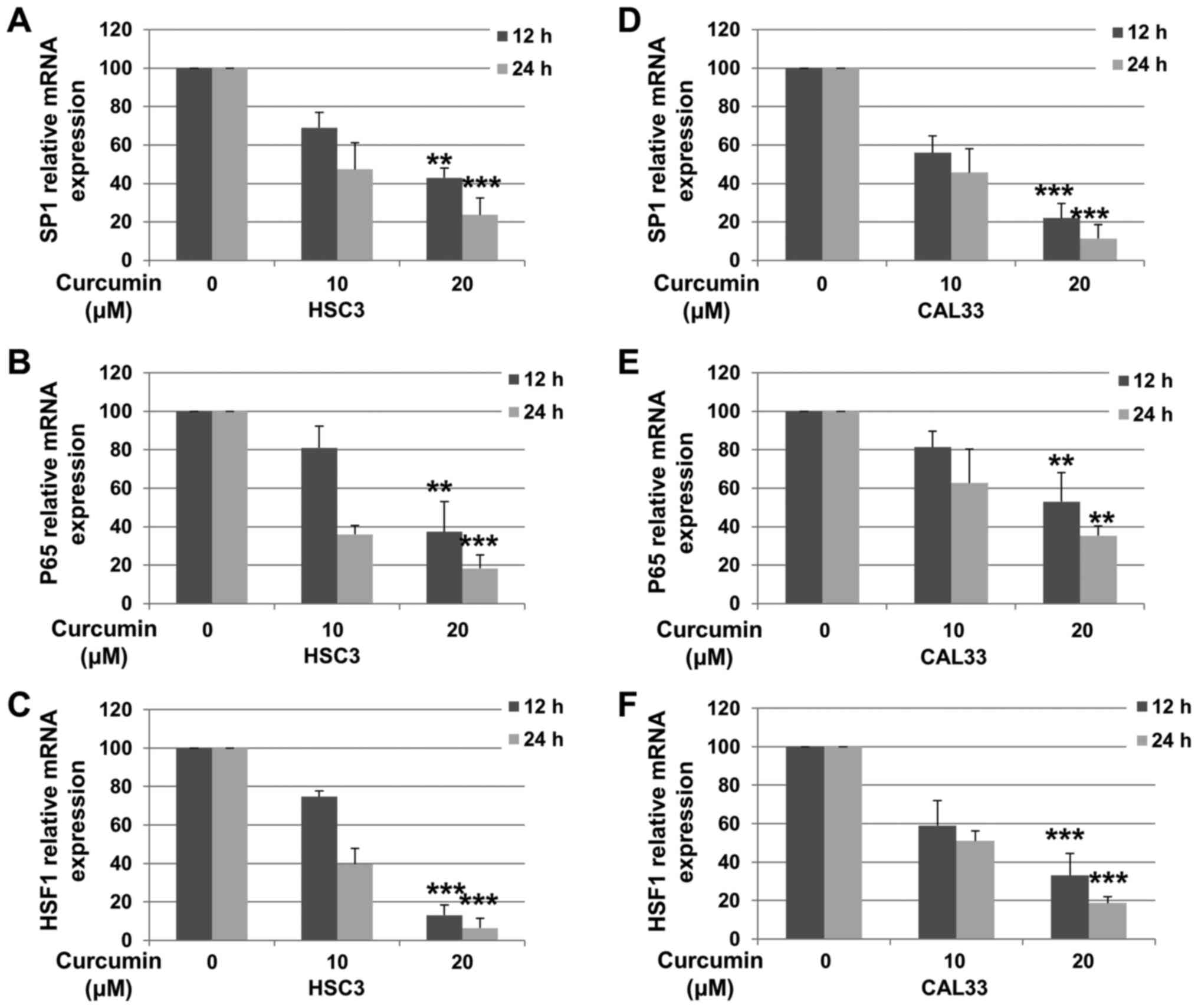
Cur decreases the expression level of
Sp1, p65 and HSF1 in OSCC cells
In order to determine whether the decreased protein
expression in OSCC cells was associated with decreased mRNA levels,
RT-qPCR was performed. HSC3 and CAL33 cells were incubated with or
without Cur for 12 or 24 h. As presented in Fig. 3A and D, the mRNA expression of Sp1 in HSC3 [12 h
(P=0.0036) and 24 h (P<0.001)] and CAL33 [12 h (P<0.001) and
24 h (P<0.001)] cells was decreased following treatment with 20
µM Cur. Furthermore, Cur treatment significantly downregulated the
expression levels of p65 [Fig. 3B
(12 h, P=0.0052; 24 h, P<0.001) and Fig. 3E (12 h, P=0.0083; 24 h, P=0.0021)]
and HSF1 [Fig. 3C (12 h,
P<0.001; 24 h, P<0.001) and Fig.
3E (12 h, P<0.001; 24 h, P<0.001)] in both OSCC cell
lines. These results suggested that Cur decreased the expression of
Sp1, p65 and HSF1 in OSCC cells by downregulating their
transcriptional levels.
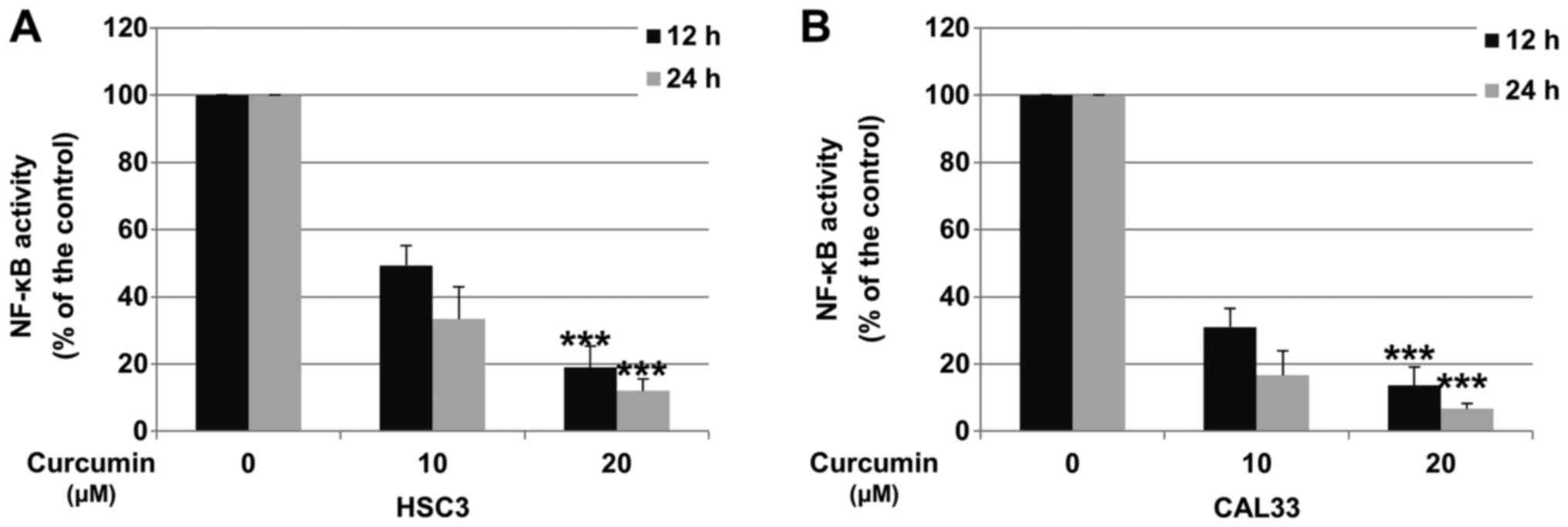
Cur decreases NF-κB activity in OSCC
cells
Downregulation of p65 was reported to decrease the
activity of the NF-κB pathway (31). To determine whether Cur could
inhibit NF-κB activity in OSCC cells, HSC3 and CAL33 cells were
incubated with DMSO (control) or Cur, and the activity of the NF-κB
pathway was determined via Dual-Luciferase reporter assay. As
presented in Fig. 4A and B, after 24 h treatment, 20 µM Cur
significantly decreased the activity of NF-κB by 88.2 and 95.4% in
HSC3 and CAL33 cell lines, respectively.
Cur inhibits cell proliferation and
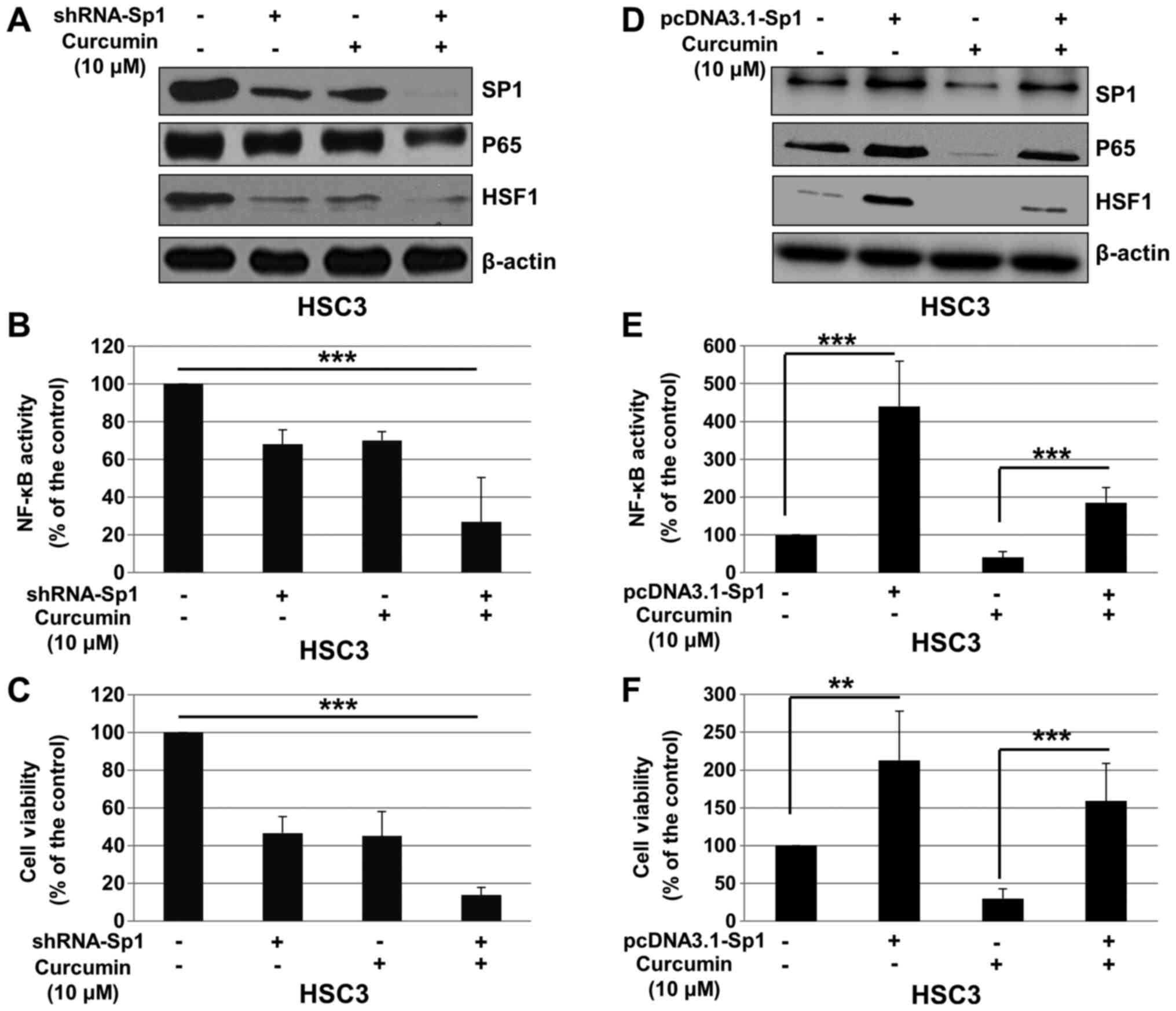
NF-κB activity in a Sp1-dependent manner
To determine whether the effects of Cur on cell
viability and NF-κB pathway are dependent of Sp1, HSC3 cells were
transfected with control shRNA or shRNA-Sp1, and treated with Cur
or DMSO (control). As presented in Fig.
5A, shRNA-Sp1 significantly downregulated the expression of
Sp1, p65 and HSF1, and enhanced the inhibitory effect of Cur on the
expression of these proteins. In addition, Sp1 knockdown
significantly enhanced the effect of Cur on NF-κB activity
(Fig. 5B) and proliferation of HSC3
cells (Fig. 5C). To further confirm
these results, pcDNA3.1 or pcDNA3.1-Sp1 was transfected into HSC3
cells, which were subsequently treated with Cur or DMSO (control).
As presented in Fig. 5D, pcDNA3.1
significantly upregulated Sp1, p65 and HSF1 expression, and
attenuated the effect of Cur on the expression of these proteins.
In addition, overexpression of Sp1 significantly reversed the
effect of Cur on NF-κB activity (Fig.
5E) and proliferation of HSC3 cells (Fig. 5F). These results suggested that Cur
may inhibit OSCC cell proliferation and NF-κB activity via Sp1
regulation.
Discussion
Chemotherapy is a treatment of choice for OSCC. Most
anticancer drugs (~80%) are derived from natural products or
analogues based on natural products, such as triptolide and
baicalein (37). Numerous studies
have reported the potent antitumor activities of Cur in various
types of cancer in vivo and in vitro, including
pancreatic, lung, breast, colorectal, ovarian, gastric and head and
neck cancers (17,38-42).
In phase I clinical trials, Cur is not associated with significant
side effects in animals or patients with breast, bladder or
pancreatic cancers (43-45).
In the present study, Cur significantly inhibited the proliferation
of OSCC cells, and significantly decreased the protein and mRNA
levels of Sp1, p65 and HSF1. In addition, Cur inhibited NF-κB
activity in OSCC cells. Furthermore, the downregulation of Sp1
significantly decreased the expression of p65 and HSF1 and enhanced
the inhibitory effect of Cur on cell proliferation and NF-κB
activity.
NF-κB highly contributes to tumor cell survival,
proliferation and metastasis and exerts anti-apoptotic effects on
various types of cancer, including OSCC (46). Tumor necrosis factor-α is known to
enhance the invasive and metastatic abilities of OSCC cells by
increasing the expression of p65 and IKKβ (47). NF-κB has been reported to increase
the expression of matrix metalloprotease-9, which is associated
with distant lymph node metastasis and poor survival of patients
with OSCC (48). It has been
demonstrated that upregulated expression of activator protein 1
(AP-1) and NF-κB in OSCC tissues is involved in Bcl-2 gene
regulation, which promotes cancer progression and resistance to
chemoradiotherapy (49). In
previous clinical studies, NF-κB overexpression has been associated
with negative prognosis of patients with OSCC, pancreatic cancer
and laryngeal squamous cell carcinoma (50-52).
p65 may therefore serve a critical role in the development and
progression of OSCC and may be considered as a potent therapeutic
target.
Cur is commonly known as a special inhibitor
targeting p65 and suppressing NF-κB activity in multiple human
cancer cells, including lung, breast and liver cancer cells
(10,14). In addition to targeting p65, Cur has
been reported to sensitize cancer cells to radiation by decreasing
the expression of inhibitor of NF-κB α (IκBα) and inhibiting NF-κB
activity (53). Furthermore, Cur
can promote paclitaxel-induced apoptosis of human
papillomavirus-positive cervical cancer cells via the
NF-κB-p53-caspase-3 pathway (54).
Previous studies have demonstrated that Cur inhibits OSCC growth by
inhibiting NF-κB pathway through various mechanisms (32,33,55).
By inhibiting NF-κB activity, Cur has been indicated to enhance
OSCC radiosensitivity in vivo and in vitro (32). Cur treatment has been revealed to
inhibit the release of epithelial-to-mesenchymal transition (EMT)
mediators in carcinoma-associated fibroblasts and induce the
reversal of EMT in tumor cells, which was indicated to decrease the
invasion efficiency of OSCC cells (33). To the best of our knowledge, the
present study was the first to demonstrate that Cur could
downregulate p65, which may account for the decreased NF-κB
activity and viability of OSCC cells.
It has been reported that Sp1 is overexpressed in
OSCC tissues compared with adjacent normal oral mucosal tissues,
suggesting that Sp1 may be considered as a potential target for
treating OSCC (56). Numerous drugs
targeting Sp1, including mithramycin A, have exhibited strong
inhibitory effects against OSCC cell proliferation (56-58).
It was previously reported that baicalein decreases the expression
of the NF-κB subunits p50 and p65, in a Sp1-dependent manner in
OSCC cells, indicating that baicalein may inhibit NF-κB pathway and
OSCC cell proliferation (30).
Since Cur has been reported to decrease Sp1 expression in various
cancer cells, including osteosarcoma and non-small cell lung cancer
cells (59,60), the expression of Sp1 was detected in
OSCC cells following Cur treatment. The results demonstrated that
Sp1 expression was significantly decreased in OSCC cells following
Cur treatment. In addition, Sp1 silencing significantly
downregulated the expression levels of p65 and HSF1, which were
detected to evaluate the effect of Cur on Sp1 expression.
Furthermore, shRNA-Sp1 significantly contributed to the effect of
Cur on cell viability and NF-κB pathway activity, which suggested
that Cur may inhibit OSCC cell proliferation in a Sp1-dependent
manner. One limitation of the present study was that transfection
experiments were only performed on HSC3 cell line.
In conclusion, the present study demonstrated that
Cur inhibited the proliferation and NF-κB activity of OSCC cells.
In addition, Cur was shown to decrease the expression of Sp1, p65
and HSF1 in OSCC cells. Sp1 knockdown contributed to the effect of
Cur on p65 and HSF1 in OSCC cells, resulting in the decreased
activity of NF-κB and cell viability. These findings suggested that
Cur may inhibit OSCC cell proliferation throuh a
Sp1/NF-κB-dependent pathway.
Acknowledgements
The authors would like to thank Dr Xiao Zhang
(University of Wuhan, China), Dr Jie Zhou (Hubei University of
Medicine, Shiyan, China), Dr Zilong Gao (Taihe Hospital, Shiyan)
and Dr Hongbin Shu (University of Wuhan, Wuhan) for supplying the
cell lines and plasmids.
Funding
The present study was supported by the Nature
Science Foundation of Hubei province for Young Scholars (grant no.
H3561204), without commercial or not-for-profit sectors.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HL designed the study. TLo performed the
experiments. TLi prepared the new reagents/analytic tools. TLo
analyzed the data. TLo and TLi wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gansler T, Ganz PA, Grant M, Greene FL,
Johnstone P, Mahoney M, Newman LA, Oh WK, Thomas CR Jr, Thun MJ, et
al: Sixty years of CA: A cancer journal for clinicians. CA Cancer J
Clin. 60:345–350. 2010.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Sinevici N and O'Sullivan J: Oral cancer:
Deregulated molecular events and their use as biomarkers. Oral
Oncol. 61:12–18. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Forastiere A, Koch W, Trotti A and
Sidransky D: Head and neck cancer. N Engl J Med. 345:1890–1900.
2001.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Hatcher H, Planalp R, Cho J, Torti FM and
Torti SV: Curcumin: From ancient medicine to current clinical
trials. Cell Mol Life Sci. 65:1631–1652. 2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ammon HP and Wahl MA: Pharmacology of
curcuma longa. Planta Med. 57:1–7. 1991.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Aggarwal BB: Targeting
inflammation-induced obesity and metabolic diseases by curcumin and
other nutraceuticals. Annu Rev Nutr. 30:173–199. 2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Sri Ramya PV, Guntuku L, Angapelly S,
Karri S, Digwal CS, Babu BN, Naidu VGM and Kamal A: Curcumin
inspired 2-chloro/phenoxy quinoline analogues: Synthesis and
biological evaluation as potential anticancer agents. Bioorg Med
Chem Lett. 28:892–898. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Lelli D, Pedone C, Majeed M and Sahebkar
A: Curcumin and lung cancer: The role of microRNAs. Curr Pharm Des.
23:3440–3444. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Basha R, Connelly SF, Sankpal UT, Nagaraju
GP, Patel H, Vishwanatha JK, Shelake S, Tabor-Simecka L, Shoji M,
Simecka JW and El-Rayes B: Small molecule tolfenamic acid and
dietary spice curcumin treatment enhances antiproliferative effect
in pancreatic cancer cells via suppressing Sp1, disrupting NF-kB
translocation to nucleus and cell cycle phase distribution. J Nutr
Biochem. 31:77–87. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chatterjee B, Ghosh K, Suresh L and Kanade
SR: Curcumin ameliorates PRMT5-MEP50 arginine methyltransferase
expression by decreasing the Sp1 and NF-YA transcription factors in
the A549 and MCF-7 cells. Mol Cell Biochem. 455:73–90.
2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hassanalilou T, Ghavamzadeh S and Khalili
L: Curcumin and gastric cancer: A review on mechanisms of action. J
Gastrointest Cancer. 50:185–192. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang HH, Zhang Y, Cheng YN, Gong FL, Cao
ZQ, Yu LG and Guo XL: Metformin incombination with curcumin
inhibits the growth, metastasis, and angiogenesis of hepatocellular
carcinoma in vitro and in vivo. Mol Carcinog. 57:44–56.
2018.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Mohandas KM and Desai DC: Epidemiology of
digestive tract cancers in India V. Large and small bowel. Indian J
Gastroenterol. 18:118–121. 1999.PubMed/NCBI
|
|
14
|
Marquardt JU, Gomez-Quiroz L, Arreguin
Camacho LO, Pinna F, Lee YH, Kitade M, Domínguez MP, Castven D,
Breuhahn K, Conner EA, et al: Curcumin effectively inhibits
oncogenic NF-κB signaling and restrains stemness features in liver
cancer. J Hepatol. 63:661–669. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Schwertheim S, Wein F, Lennartz K, Worm K,
Schmid KW and Sheu-Grabellus SY: Curcumin induces G2/M arrest,
apoptosis, NF-κB inhibition, and expression of differentiation
genes in thyroid carcinoma cells. J Cancer Res Clin Oncol.
143:1143–1154. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Huang Y, Hu L, Huang S, Xu W, Wan J, Wang
D, Zheng G and Xia Z: Curcumin-loaded galactosylated BSA
nanoparticles as targeted drug delivery carriers inhibit
hepatocellular carcinoma cell proliferation and migration. Int J
Nanomedicine. 13:8309–8323. 2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wang D, Veena MS, Stevenson K, Tang C, Ho
B, Suh JD, Duarte VM, Faull KF, Mehta K, Srivatsan ES and Wang MB:
Liposome-encapsulated curcumin suppresses growth of head and neck
squamous cell carcinoma in vitro and in xenografts through the
inhibition of nuclear factor kappaB by an AKT-independent pathway.
Clin Cancer Res. 14:6228–6236. 2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Shi Q, Le X, Abbruzzese JL, Peng Z, Qian
CN, Tang H, Xiong Q, Wang B, Li XC and Xie K: Constitutive Sp1
activity is essential for differential constitutive expression of
vascular endothelial growth factor in human pancreatic
adenocarcinoma. Cancer Res. 61:4143–4154. 2001.PubMed/NCBI
|
|
19
|
Abdelrahim M, Smith R III, Burghardt R and
Safe S: Role of Sp proteins in regulation of vascular endothelial
growth factor expression and proliferation of pancreatic cancer
cells. Cancer Res. 64:6740–6749. 2004.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Black AR, Black JD and Azizkhan-Clifford
J: Sp1 and kruppel-like factor family of transcription factors in
cell growth regulation and cancer. J Cell Physiol. 188:143–160.
2001.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Safe S and Abdelrahim M: Sp transcription
factor family and its role in cancer. Eur J Cancer. 41:2438–2448.
2005.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Hu H, Wu LL, Han T, Zhuo M, Lei W, Cui JJ,
Jiao F and Wang LW: Correlated high expression of FXR and Sp1 in
cancer cells confers a poor prognosis for pancreatic cancer: A
study based on TCGA and tissue microarray. Oncotarget.
8:33265–33275. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jiang W, Jin Z, Zhou F, Cui J and Wang L
and Wang L: High co-expression of Sp1 and HER-2 is correlated with
poor prognosis of gastric cancer patients. Surg Oncol. 24:220–225.
2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Su CW, Chang YC, Chien MH, Hsieh YH, Chen
MK, Lin CW and Yang SF: Loss of TIMP3 by promoter methylation of
Sp1 binding site promotes oral cancer metastasis. Cell Death Dis.
10(793)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Yu HJ, Shin JA, Nam JS, Kang BS and Cho
SD: Apoptotic effect of dibenzylideneacetone on oral cancer cells
via modulation of specificity protein 1 and Bax. Oral Dis.
19:767–774. 2013.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Cho JH, Shin JC, Cho JJ, Choi YH, Shim JH
and Chae JI: Esculetin (6,7-dihydroxycoumarin): A potential cancer
chemopreventive agent through suppression of Sp1 in oral squamous
cancer cells. Int J Oncol. 46:265–271. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jeon YJ, Bang W, Choi YH, Shim JH and Chae
JI: Beta-lapachone suppresses non-small cell lung cancer
proliferation through the regulation of specificity protein 1. Biol
Pharm Bull. 38:1302–1308. 2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Jeon YJ, Bang W, Shin JC, Park SM, Cho JJ,
Choi YH, Seo KS, Choi NJ, Shim JH and Chae JI: Downregulation of
Sp1 is involved in β-lapachone-induced cell cycle arrest and
apoptosis in oral squamous cell carcinoma. Int J Oncol.
46:2606–2612. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Kim DW, Ko SM, Jeon YJ, Noh YW, Choi NJ,
Cho SD, Moon HS, Cho YS, Shin JC, Park SM, et al:
Anti-proliferative effect of honokiol in oral squamous cancer
through the regulation of specificity protein 1. Int J Oncol.
43:1103–1110. 2013.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Gao Z, Zhang Y, Zhou H and Lv J: Baicalein
inhibits the growth of oral squamous cell carcinoma cells by
downregulating the expression of transcription factor Sp1. Int J
Oncol. 56:273–282. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Banerjee S, Sangwan V, McGinn O, Chugh R,
Dudeja V, Vickers SM and Saluja AK: Triptolide-induced cell death
in pancreatic cancer is mediated by O-GlcNAc modification of
transcription factor Sp1. J Biol Chem. 288:33927–33938.
2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Chiang IT, Liu YC, Hsu FT, Chien YC, Kao
CH, Lin WJ, Chung JG and Hwang JJ: Curcumin synergistically
enhances the radiosensitivity of human oral squamous cell carcinoma
via suppression of radiation-induced NF-κB activity. Oncol Rep.
31:1729–1737. 2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Dudás J, Fullar A, Romani A, Pritz C,
Kovalszky I, Hans Schartinger V, Mathias Sprinzl G and Riechelmann
H: Curcumin targets fibroblast-tumor cell interactions in oral
squamous cell carcinoma. Exp Cell Res. 319:800–809. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Jiang L and Xiao J:
2-phenylethynesulfonamide inhibits growth of oral squamous cell
carcinoma cells by blocking the function of heat shock protein 70.
Biosci Reps. 40(BSR20200079)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhang Y, Wang H, Liu Y, Wang C, Wang J,
Long C, Guo W and Sun X: Baicalein inhibits growth of Epstein-Barr
virus-positive nasopharyngeal carcinoma by repressing the activity
of EBNA1 Q-promoter. Biomed Pharmacothe. 102:1003–1014.
2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Newman DJ, Cragg GM and Snader KM: Natural
products as sources of new drugs over the period 1981-2002. J Nat
Prod. 66:1022–1037. 2003.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Zhang Q, Qiao H, Wu D, Lu H, Liu L, Sang
X, Li D and Zhou Y: Curcumin potentiates the galbanic acid-induced
anti-tumor effect in non-small cell lung cancer cells through
inhibiting Akt/mTOR signaling pathway. Life Sci.
239(117044)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li W, Wang Z, Xiao X, Han L, Wu Z, Ma Q
and Cao L: Curcumin attenuates hyperglycemia-driven EGF-induced
invasive and migratory abilities of pancreatic cancer via
suppression of the ERK and AKT pathways. Oncol Rep. 41:650–658.
2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Pereira MC, Mohammed R, Van Otterlo WAL,
De Koning CB and Davids H: In vitro analysis of the combinatory
effects of novel aminonaphthoquinone derivatives and curcumin on
breast cancer progression. Anticancer Res. 40:229–238.
2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Tian M, Tian D, Qiao X, Li J and Zhang L:
Modulation of Myb-induced NF-kB-STAT3 signaling and resulting
cisplatin resistance in ovarian cancer by dietary factors. J Cell
Physiol. 234:21126–21134. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Boven L, Holmes SP, Latimer B, McMartin K,
Ma X, Moore-Medlin T, Khandelwal AR, McLarty J and Nathan CO:
Curcumin gum formulation for prevention of oral cavity head and
neck squamous cell carcinoma. Laryngoscope. 129:1597–1603.
2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Cheng AL, Hsu CH, Lin JK, Hsu MM, Ho YF,
Shen TS, Ko JY, Lin JT, Lin BR, Ming-Shiang W, et al: Phase I
clinical trial of curcumin, a chemopreventive agent, in patients
with high-risk or pre-malignant lesions. Anticancer Res.
21:2895–2900. 2001.PubMed/NCBI
|
|
44
|
Kanai M, Yoshimura K, Asada M, Imaizumi A,
Suzuki C, Matsumoto S, Nishimura T, Mori Y, Masui T, Kawaguchi Y,
et al: A phase I/II study of gemcitabine-based chemotherapy plus
curcumin for patients with gemcitabine-resistant pancreatic cancer.
Cancer Chemother Pharmacol. 68:157–164. 2011.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Bayet-Robert M, Kwiatkowski F, Leheurteur
M, Gachon F, Planchat E, Abrial C, Mouret-Reynier MA, Durando X,
Barthomeuf C and Chollet P: Phase I dose escalation trial of
docetaxel plus curcumin in patients with advanced and metastatic
breast cancer. Cancer Biol Ther. 9:8–14. 2010.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Monisha J, Roy NK, Bordoloi D, Kumar A,
Golla R, Kotoky J, Padmavathi G and Kunnumakkara AB: Nuclear factor
Kappa B: A potential target to persecute head and neck cancer. Curr
Drug Targets. 18:232–253. 2017.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Tang D, Tao D, Fang Y, Deng C, Xu Q and
Zhou J: TNF-alpha promotes invasion and metastasis via NF-Kappa B
pathway in oral squamous cell carcinoma. Med Sci Monit Basic Res.
23:141–149. 2017.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Bond M, Fabunmi RP, Baker AH and Newby AC:
Synergistic upregulation of metalloproteinase-9 by growth factors
and inflammatory cytokines: An absolute requirement for
transcription factor NF-kappa B. FEBS Lett. 435:29–34.
1998.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Alam M, Kashyap T, Pramanik KK, Singh AK,
Nagini S and Mishra R: The elevated activation of NFκB and AP-1 is
correlated with differential regulation of Bcl-2 and associated
with oral squamous cell carcinoma progression and resistance. Clin
Oral Investig. 21:2721–2731. 2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Yoshida K, Sasaki R, Nishimura H, Okamoto
Y, Suzuki Y, Kawabe T, Saito M, Otsuki N, Hayashi Y, Soejima T, et
al: Nuclear factor-kappaB expression as a novel marker of
radioresistance in Early-stage laryngeal cancer. Head Neck.
32:646–655. 2010.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Yang SH, Hsu CH, Lee JC, Tien YW, Kuo SH
and Cheng AL: Nuclear expression of glioma-associated oncogene
homolog 1 and nuclear factor-κB is associated with a poor prognosis
of pancreatic cancer. Oncology. 85:86–94. 2013.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Jiang LZ, Wang P, Deng B, Huang C, Tang
WX, Lu HY and Chen HY: Overexpression of forkhead box M1
transcription factor and nuclear factor-κB in laryngeal squamous
cell carcinoma: A potential indicator for poor prognosis. Hum
Pathol. 42:1185–1193. 2011.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Qiao Q, Jiang Y and Li G: Curcumin
improves the antitumor effect of X-ray irradiation by blocking the
NF-κB pathway: An in-vitro study of lymphoma. Anticancer Drugs.
23:597–605. 2012.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Teymouri M, Pirro M, Johnston TP and
Sahebkar A: Curcumin as a multifaceted compound against human
papilloma virus infection and cervical cancers: A review of
chemistry, cellular, molecular, and preclinical features.
Biofactors. 43:331–346. 2017.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Shin HK, Kim J, Lee EJ and Kim SH:
Inhibitory effect of curcumin on motility of human oral squamous
carcinoma YD-10B cells via suppression of ERK and NF-kappaB
activations. Phytother Res. 24:577–582. 2010.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Shin JA, Kim JJ, Choi ES, Shim JH, Ryu MH,
Kwon KH, Park HM, Seo JY, Lee SY, Lim DW, et al: In vitro apoptotic
effects of methanol extracts of dianthus chinensis and Acalypha
australis L targeting specificity protein 1 in human oral cancer
cells. Head Neck. 35:992–998. 2013.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Sachita K, Yu HJ, Yun JW, Lee JS and Cho
SD: YM155 induces apoptosis through downregulation of specificity
protein 1 and myeloid cell leukemia-1 in human oral cancer cell
lines. J Oral Pathol Med. 44:785–791. 2015.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Hsieh MJ, Chen JC, Yang WE, Chien SY, Chen
MK, Lo YS, His YT, Chuang YC, Lin CC and Yang SF:
Dehydroandrographolide inhibits oral cancer cell migration and
invasion through NF-κB-, AP-1-, and SP-1-modulated matrix
metalloproteinase-2 inhibition. Biochem Pharmacol. 130:10–20.
2017.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Chen P, Huang HP, Wang Y, Jin J, Long WG,
Chen K, Zhao XH, Chen CG and Li J: Curcumin overcome primary
gefitinib resistance in non-small-cell lung cancer cells through
inducing autophagy-related cell death. J Exp Clin Cancer Res.
38(254)2019.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Lima FT, Seba V, Silva G, Torrezan GS,
Polaquini CR, Pinhanelli VC, Baek SJ, Fachin AL, Regasini LO and
Marins M: The curcumin analog CH-5 exerts anticancer effects in
human osteosarcoma cells via modulation of transcription factors
p53/Sp1. Int J Mol Sci. 19(1909)2018.PubMed/NCBI View Article : Google Scholar
|