Introduction
A hypertrophic scar (HS) is a severe fibrotic
cutaneous disorder characterized by morphological abnormality and
limited movement (1). It often
occurs after deep skin injuries such as extensive burns. The
incidence of HSs during cicatrix formation after a burn injury is
up to 70% (2). This not only
damages the physical and psychological health of patients, but also
places a heavy economic burden on their families and society.
Development of an effective therapy for HSs would be highly
beneficial.
The development of a curative strategy for
hypertrophic scarring relies on investigation into the underlying
pathophysiological mechanism (3).
Previous studies of HSs have revealed that their formation is
driven by an abnormal composition and excessive deposition of
extracellular matrix (ECM), and that overactive hypertrophic scar
fibroblasts (HSFs) are one of the main effector cells responsible
for these pathological changes (4).
Furthermore, the transforming growth factor β1 (TGF β1)/Smad3
pathway has been recognized as a principal cellular signaling
pathway in the promotion of fibrosis of HSFs (5). The phosphorylation of Smad3 is an
important step in this signaling cascade (6,7).
Regulation of this pathway in HSFs may have an essential role in
future HS therapies (3).
Application of mesenchymal stem cells (MSCs) in HS
therapy is a possible approach to treatment (8,9). MSCs
derived from different tissues have different biological
characteristics, but research has largely been focused on bone
marrow-derived MSCs (BM-MSCs) and adipose-derived MSCs (AD-MSCs)
(10-13).
There are few reports focusing on the therapeutic benefit of
umbilical cord-derived MSCs (UCMSCs), which possess a high
proliferative ability, weak immunogenicity and a specific
anti-fibrotic paracrine profile (14-16).
To the best of our knowledge, there has been limited research into
the exact mechanism of administration of UCMSCs for HS therapy.
In the present study the effect of UCMSCs on the
pro-fibrotic phenotype of HSFs in a co-culture system and the
potential molecular mechanisms underlying this regulation were
investigated. Pro-fibrotic phenotype was measured through
assessment of fibrosis-related cellular behaviours, including cell
proliferation, apoptosis, migration and the expression of
HS-associated genes and proteins.
Materials and methods
Isolation and culture of cells
UCMSCs were kindly provided by Stem Cell Bank,
Chinese Academy of Sciences and were cultured in DMEM containing
10% fetal bovine serum (FBS) and antibiotics (penicillin, 100 U/ml;
streptomycin, 0.1 mg/ml) at 37˚C with 5% CO2. These
UCMSCs possess the ability to differentiate into adipose tissue,
bone and cartilage and are CD29, CD44, CD73, CD90, CD105 and CD166
positive, while CD14, CD31, CD34, and CD45 negative.
Primary HSFs were cultured from the tissues of six
patients (3 male and 3 female; age, 17 months to 27 years) with HSs
who had received a cicatrectomy in the Department of Burn Surgery
of The First Hospital of Jilin University (Changchun, China) from
April 2016 to July 2017. Briefly, HS tissue mass was washed twice
with phosphate-buffered saline and cut into ~1-mm3
sections under sterile conditions. Washed sections were placed in a
culture plate with a distance of 1 cm between each other at 37˚C
for 30 min, and then incubated in DMEM containing 10% FBS and
antibiotics (penicillin, 100 U/ml; streptomycin, 0.1 mg/ml) at 37˚C
with 5% CO2. All cell culture reagents were supplied by
Gibco; Thermo Fisher Scientific, Inc. When UCMSCs and HSFs reached
90% confluence, they were trypsinized and prepared for subculture.
UCMSCs at passages 6-8 and HSFs at passages 2-4 were used in the
experiments described below.
Co-culture of cells
Transwell culture plates (Millicell®; EMD
Milllipore) were used to develop the co-culture system [a 96-well
culture plate was used for the Cell Counting Kit-8 (CCK-8) assay,
and six-well culture plates were used for the other experiments in
this study]. UCMSCs at passages 6-8 were harvested and then seeded
on the upper chamber of a transwell culture plate at a density of
1x104 cells/cm2. HSFs at passages 2-4 HSFs
were harvested and seeded on the lower chamber at the same density.
The co-culture system was cultured in DMEM containing 10% FBS and
antibiotics (penicillin, 100 U/ml; streptomycin, 0.1 mg/ml). The
cultured HSFs were divided into co-culture and control groups based
on the culture conditions. No UCMSCs were included in the upper
chamber of the control group. Cells were co-cultured for 12-72 h
for proliferation analysis, 48 h for apoptosis analysis, and for 72
h for migration analysis and detecting gene and protein expression
levels.
CCK-8 assay
CCK-8 (Beijing Solarbio Science & Technology
Co., Ltd.) was used to monitor cell proliferation. Briefly, HSFs
were seeded at 5x103 cells per well in a 96-well culture
plate with three replicates per sample. At 12, 24, 36, 48 and 72 h
after cell attachment, 10 µl of CCK-8 solution was added to each
well. After incubation for 1 h, the absorbance was measured at 450
nm using a microplate reader (Thermo Fisher Scientific, Inc.).
Immunofluorescence
The media of prepared HSFs were discarded, and
pre-cooled methanol was added to fix the cells for 30 min at room
temperature. Cells were incubated with PBS containing 0.2% Triton
X-100 for 15 min at room temperature for permeabilization, and with
TBST (0.5% Tween-20) containing 1% BSA for 30 min at room
temperature for blocking. For Ki67 staining, the cells were
incubated with a primary anti-Ki67 antibody (1:100; cat. no.
ab197234; Abcam) at 4˚C overnight followed by incubation with the
corresponding CY3-conjugated secondary antibody (1:2,000; cat. no.
ab6939; Abcam) at 37˚C for 2 h. For TUNEL staining, the cells were
incubated with a TUNEL reagent (Beyotime Institute of
Biotechnology) at 37˚C for 1 h. Finally, after the nuclei were
counterstained with DAPI (1:1,000; Beyotime Institute of
Biotechnology) for 10 min at room temperature, the slides were
mounted with Antifade Mounting Medium (Beyotime Institute of
Biotechnology). Images of each slide were captured at three random
fields of view using an inverted fluorescence microscope
(magnification, x100; IX73; Olympus Corporation). Total nuclei
(blue) and Ki67 or TUNEL-positive (red) cells were quantified using
ImageJ software (version 1.51w; National Institutes of Health).
Scratch wound closure assay
Cell migration was tested using a scratch wound
closure assay. In brief, HSFs of co-culture and control groups were
cultured in a six-well culture plate at a density of
1x104 cells/cm2 with DMEM containing 10% FBS
until the cell confluence reached 100%. A 1-ml pipette tip was used
to make a scratch wound in the middle of each well. After washing
three times with PBS, the medium was changed to serum-free DMEM for
72 h. Images were acquired using an inverted microscope
(magnification, x40; Olympus Corporation) after 24, 48 and 72 h.
Wound area was measured using ImageJ software (version 1.51;
National Institutes of Health). Results are shown as the percentage
of area closed, which was calculated through normalizing the wound
space of each time point to that of 0 h. Images of each well were
captured at three random fields of view.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated, and RT was performed as
previously described (17).
Briefly, total RNA was isolated from HSFs from the control group
and the group co-cultured with UCMSCs for 3 days using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). cDNA was synthesized using the TransScript All-in-One
First-Strand cDNA Synthesis SuperMix (Beijing Transgen Biotech Co.,
Ltd.). FastStart Universal SYBR Green Master (ROX) (Roche
Diagnostics GmbH) was used for qPCR using a Stratagene Mx3005P
instrument (Agilent Technologies, GmbH). The reaction conditions
were: Initial denaturation at 95˚C for 10 min; 40 cycles of
denaturation at 95˚C for 30 sec and annealing at 60˚C for 1 min;
dissociation at 95˚C for 1 min, annealing at 55˚C for 30 sec, and
final extension at 95˚C for 30 sec. The primers used in this study
for gene amplification are listed in Table I. Expression levels of target genes
were normalized to that of GAPDH and the 2-∆∆Cq
method was used to calculate the relative expression levels of
genes (18). Each sample was run in
triplicate wells.
 | Table ISequences of primers used for
PCR. |
Table I
Sequences of primers used for
PCR.
| | Primer sequence
(5'→3') |
|---|
| Gene | Forward | Reverse |
|---|
| COL1A2 |
GAGGGCAACAGCAGGTTCACTTA |
TCAGCACCACCGATGTCCA |
| COL3A1 |
CCACGGAAACACTGGTGGAC |
GCCAGCTGCACATCAAGGAC |
| ACTA2 |
GACAATGGCTCTGGGCTCTGTAA |
TGTGCTTCGTCACCCACGTA |
| TGFB1 |
AACTCCGGTGACATCAAAAGATAA |
TGCTGAGGCTCAAGTTAAAAGT |
| CTGF |
CTGGAAGGACTCTCCGCTGCGG |
GCGACCCGCACAAGGGCCTAT |
| TIMP1 |
ACCCACAGACGGCCTTCTGCAATT |
AACGCTGGTATAAGGTGGTCTGGTT |
| POSTN |
CTCAGAGCAGATGCCAAGCCTAATTG |
GTGTGATCCATTTGATTGATCAGGTCCT |
| GAPDH |
GCACCGTCAAGCTGAGAAC |
TGGTGAAGACGCCAGTGGA |
Western blotting
HSFs of co-culture and control groups were harvested
and washed with PBS. The cells were processed with RIPA lysis
buffer (CST Biological Reagents Co., Ltd.) supplemented with
phenylmethylsulphonyl fluoride (Thermo Fisher Scientific, Inc.),
protease inhibitor cocktail (TransGen Biotech Co., Ltd.) and
phosphatase inhibitor cocktail (TransGen Biotech Co., Ltd.).
Protein concentration was determined using a bicinchoninic acid
(BCA) protein assay kit (Beyotime Institute of Biotechnology).
Western blotting was performed as previously described (17). In brief, 30 µg of protein from each
sample was separated in 8% (for collagen I and III), 10% [for TGF
β1, α-smooth muscle actin (α-SMA), Smad3 and phosphorylated
(p)-Smad3] or 12% (for caspase-3) SDS-PAGE gels, and then
electro-transferred to PVDF membranes (Immobilon-P, EMD Millpore)
for immunoblotting analysis. After blocking with 5% bovine serum
albumin (Sigma-Aldrich; Merck KGaA) in TBST (0.5% Tween-20) for 1 h
at room temperature, the primary antibodies anti-collagen I
(1:1,000; cat. no. 14695-1-AP; ProteinTech Group, Inc.),
anti-collagen III (1:500; cat. no. 22734-1-AP; ProteinTech Group,
Inc.), anti-α-SMA (1:1,000; cat. no. ab32575; Abcam), anti-β-actin
(1:2,000; cat. no. 60008-1-Ig; ProteinTech, Group Inc.), anti-TGF
β1 (1:1,000; cat. no. 21898-1-AP; ProteinTech Group, Inc.),
anti-Smad3 (1:1,000; cat. no. ab40854; Abcam), anti-p-Smad3 (1:500;
cat. no. ab52903; Abcam) and anti-caspase-3 (1:500; cat. no.
ab13847; Abcam) were incubated with the PVDF membranes at 4˚C
overnight. Following incubation with the appropriate horseradish
peroxidase-conjugated secondary antibodies (1:5,000; goat
anti-mouse cat. no. SA00001-1; goat anti-rabbit cat. no. SA00001-2;
ProteinTech Group, Inc.) for 1 h at room temperature, proteins were
detected by chemiluminescence using the EasySee Western Blot kit
(TransGen Biotech Co., Ltd) in a myECL™ imager (Thermo
Fisher Scientific, Inc.), and the band intensities were quantified
using ImageJ software.
Statistical analysis
The quantified data are presented as the mean ± SD
for the CCK-8 assay, immunofluorescence (percentage of Ki67- or
TUNEL-positive cells), RT-qPCR and scratch wound closure assay, and
as the mean ± SEM for western blotting. The differences between the
two groups were analysed by Student's t-test using GraphPad Prism
(version 7.00; Graph Pad Software Inc.) statistical package. A
value of P<0.05 was considered to indicate a statistically
significant difference. All experiments were repeated at least
three times.
Results
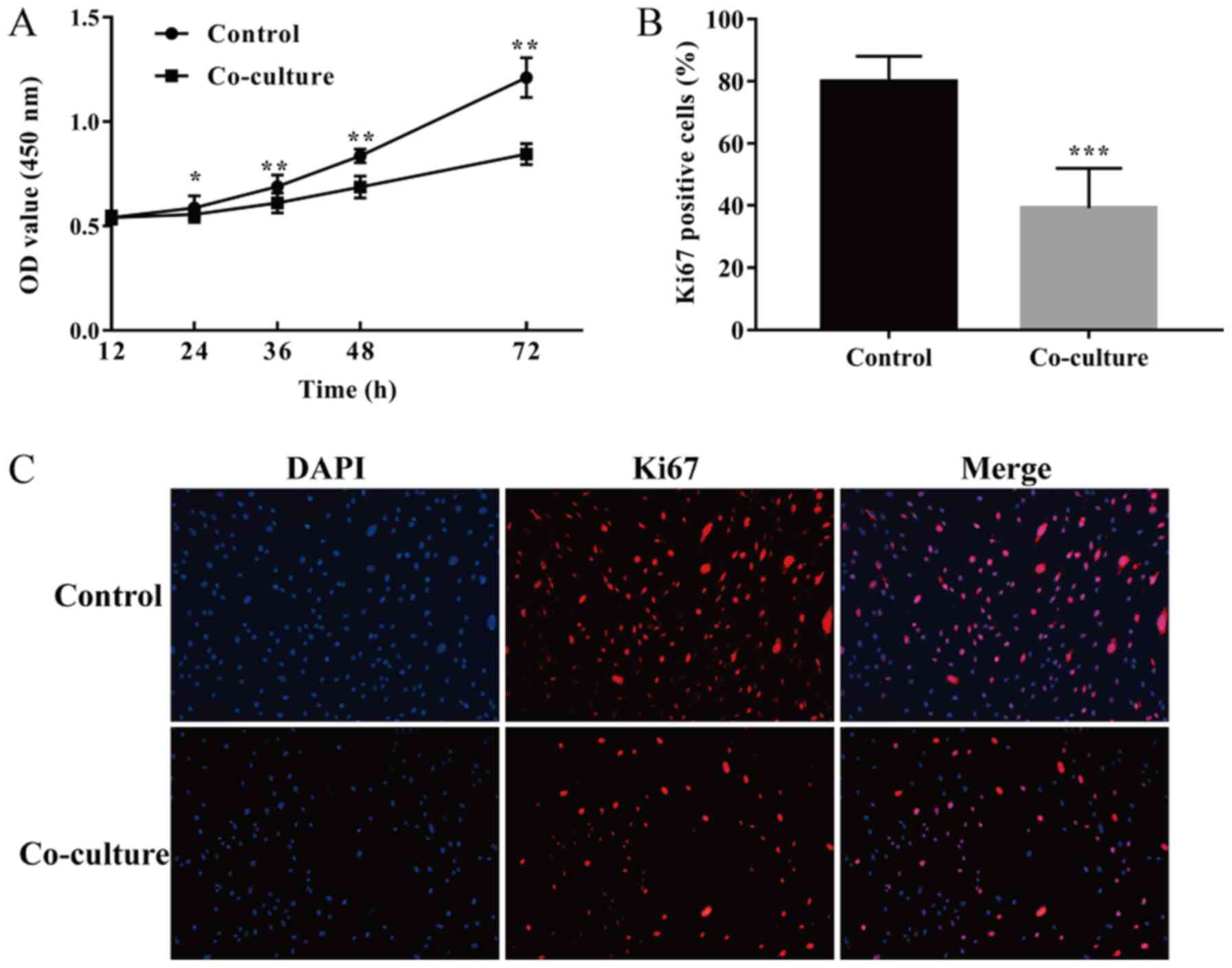
UCMSCs suppress the proliferation of
HSFs in co-culture
In order to determine whether UCMSCs affect the
proliferation of HSFs in a co-culture system, the cell viability of
HSFs cultured with or without UCMSCs at different time points was
assessed using a CCK-8 assay and the percentage of Ki67-positive
fibroblasts determined through immunofluorescence. After co-culture
with UCMSCs for 24, 36, 48, and 72 h, the cell viability of HSFs
was significantly lower compared with their respective control
groups (24 h, P<0.05; 36, 48 and 72 h, P<0.01; Fig. 1A). Additionally, the percentages of
Ki67-positive fibroblasts in the co-culture group were
significantly lower than that in the control group after culturing
for 48 h (P<0.001, Fig. 1B and
C). These differences suggested
that UCMSCs could suppress the proliferative ability of HSFs in
co-culture.
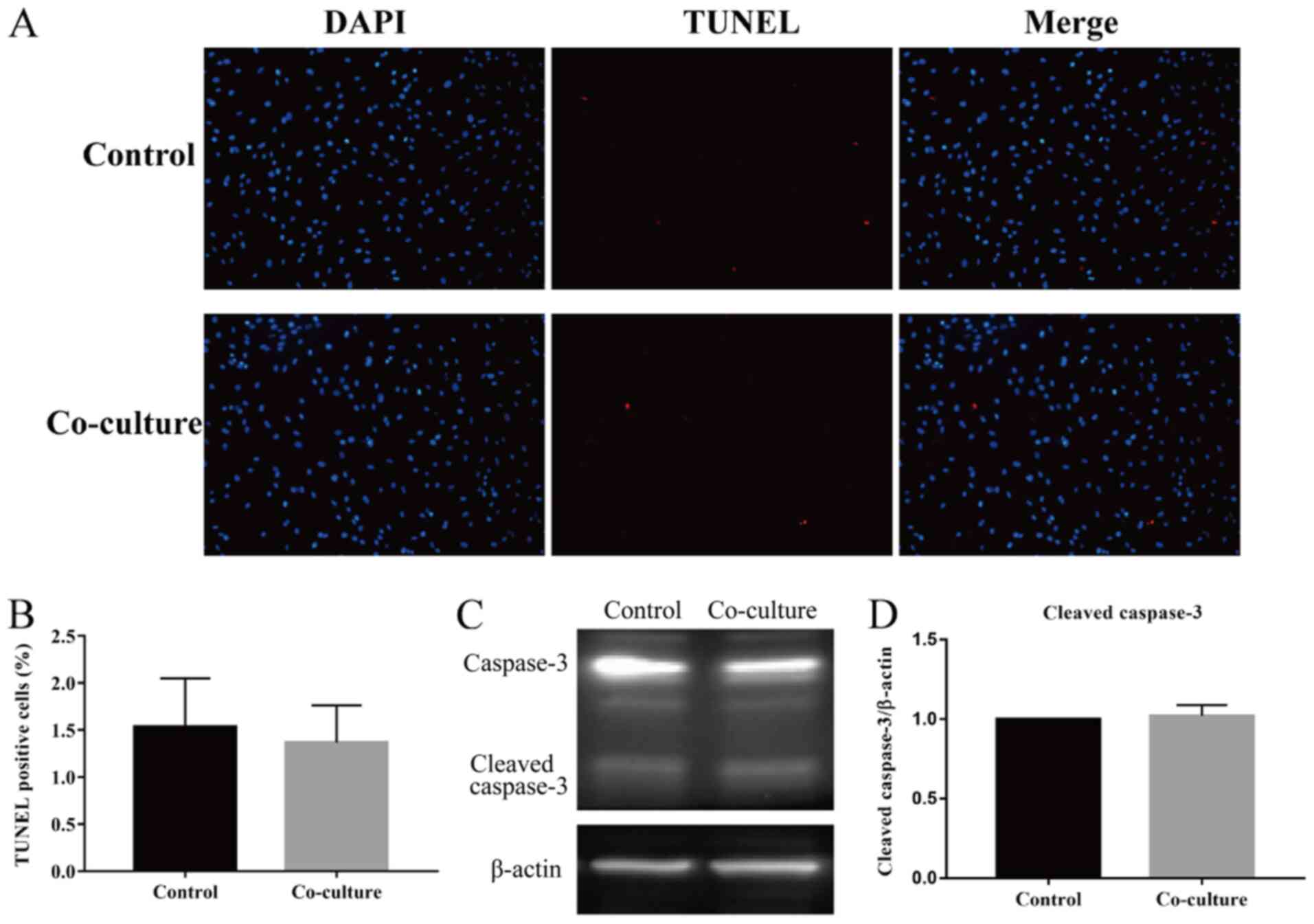
UCMSCs do not influence the apoptosis
of HSFs in co-culture
The apoptosis of HSFs was measured via a TUNEL assay
and by assessing the expression of caspase-3, a key
apoptosis-related protein, through western blotting after 48 h of
co-culture. As shown in Fig. 2A and
B, the TUNEL assay revealed that
there was no significant difference between the percentage of
apoptosis-positive cells from the two groups. Moreover, the protein
levels of cleaved caspase-3, which indicated the apoptosis level,
revealed that there was no significant difference between levels in
the HSFs of co-cultured groups compared with those in the controls
(Fig. 2C and D). Taken together, the results
demonstrated that UCMSCs had no influence on the rate of apoptosis
of HSFs in a co-culture system.
UCMSCs inhibit the migration of
co-cultured HSFs
To study the effect of UCMSCs on the migration of
HSFs in co-culture, a scratch wound healing assay was performed.
Representative images of this assay at the time points 24, 48 and
72 h after scratching showed that the migration ability of
co-cultured HSFs into the scratched space was inhibited compared
with that of controls (Fig. 3A).
The scratch wound of the controls cultured without UCMSCs was
almost closed after 72 h; however, the corresponding co-cultured
subset was not. The percentage of wound closure area at different
time points compared with the 0 h controls was quantified. As shown
in Fig. 3B, after 24, 48 and 72 h
of culture, respectively, 40.23±1.73, 70.4±1.8 and 86.52±1.1% of
the scratched space was filled by the migrated HSFs in the control
groups; by contrast, 28.69±1.85, 36.64±0.9 and 47.19±1.36% of space
was filled by the migrated HSFs in the co-culture groups,
indicating a significant difference between the two groups in terms
of HSF migration (24, 48 and 72 h, P<0.01; Fig. 3B). These data demonstrated that
UCMSCs could significantly inhibit the migration ability of HSFs in
co-culture.
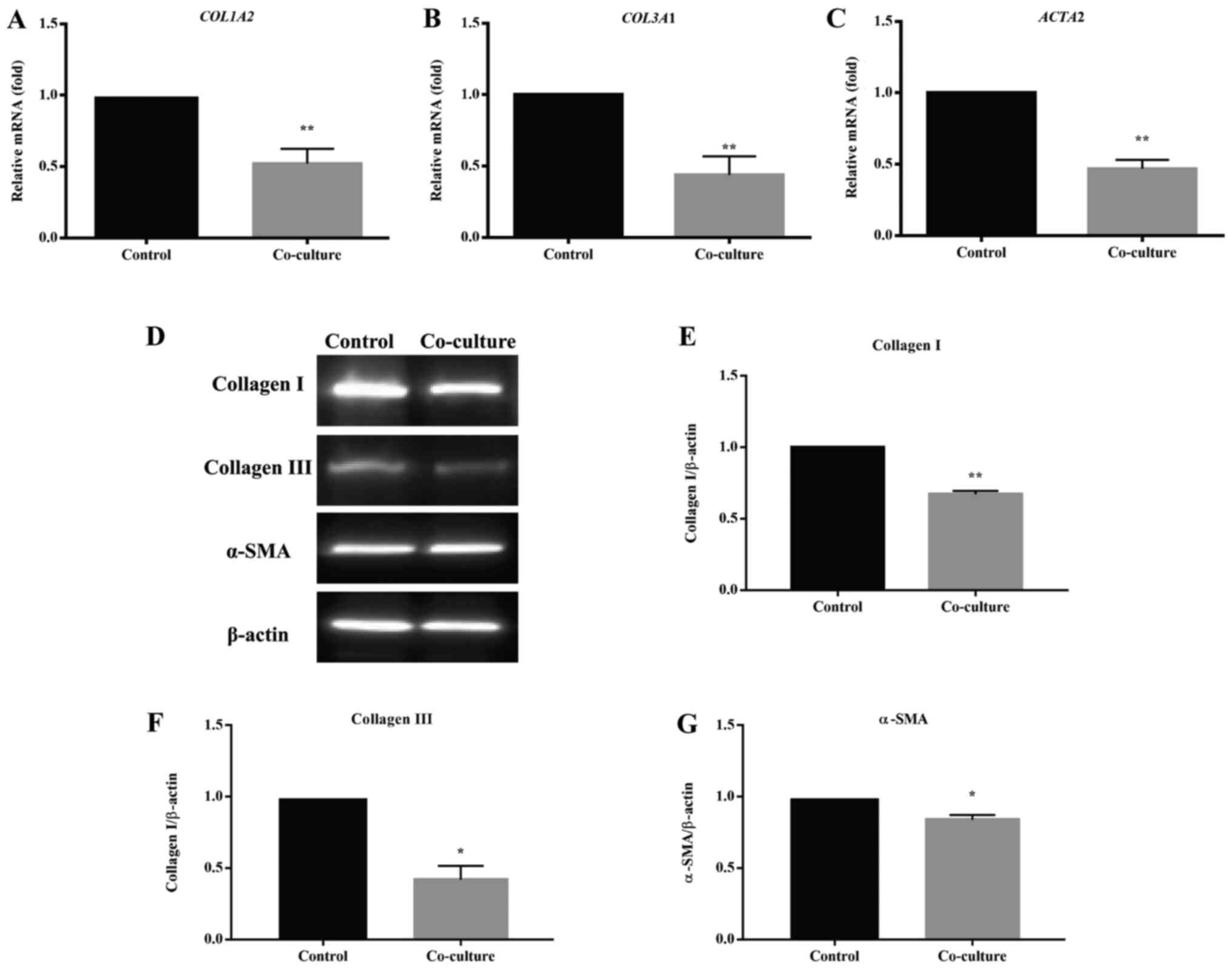
UCMSCs reduce HS-associated gene and
protein expression in HSFs in co-culture
The expression of HS-associated genes and proteins
was measured to examine the effect of UCMSCs on the pro-fibrotic
phenotype of HSFs in a co-culture system. The mRNA levels of
collagen type I α 2 chain (COL1A2), collagen type III α 1
chain (COL3A1) and actin α 2 smooth muscle (ACTA2),
and the protein levels of collagen I, collagen III and α-SMA, which
play essential roles in HS formation, were assessed by RT-qPCR and
western blotting, respectively (Fig.
4). RT-qPCR showed that the mRNA levels of COL1A2,
COL3A1 and ACTA2 of HSFs from the co-culture groups
were significantly lower than those from their respective control
groups (all, P<0.01; Fig. 4A-C).
Consistent with the changes in the mRNA levels, the protein levels
of collagen I, collagen III and α-SMA in the co-cultured HSFs
decreased compared with the controls, as shown through western
blotting (Fig. 4D) and further
quantitative and statistical analysis (collagen I, P<0.05,
Fig. 4E; collagen III and α-SMA,
P<0.01, Fig. 4F and G). All the above results suggested that
UCMSCs could inhibit the pro-fibrotic phenotype of HSFs in
co-culture.
TGF β1/Smad3 signaling pathway was
inhibited in HSFs co-cultured with UCMSCs
To further investigate the potential mechanism
underlying the anti-fibrotic effect of UCMSCs, levels of the
related key protein molecules of the TGF β1/Smad3 pathway in HSFs
were assessed. As shown in Fig. 5A,
the levels of TGF β1 and p-Smad3 in HSFs co-cultured with UCMSCs
were significantly reduced (TGF β1, P<0.05, Fig. 5B; p-Smad3, P<0.01, Fig. 5D), whereas no significant difference
between the levels of total Smad3 (t-Smad3) protein was observed
(Fig. 5C). However, the ratio of
p-Smad3 to t-Smad3 was significantly decreased in co-culture group
compared with the control (P<0.01, Fig. 5E). The mRNA levels of TGFB1
and other important genes targeted by this pathway [cellular
communication network factor 2 (CTGF), metalloproteinase
inhibitor 1 (TIMP1) and periostin (POSTN)] were then
examined. Statistical analysis indicated that the transcription of
these genes in HSFs from the co-cultured group was inhibited
compared with the control (TGFB1, P<0.01, Fig. 5F; CTGF, P<0.01, Fig. 5G; TIMP1, P<0.05, Fig. 5H; POSTN, P<0.0001,
Fig. 5I). This suggested that
UCMSCs may inhibit the fibrosis of HSFs by inhibiting the TGF
β1/Smad3 pathway, and that the reduction of TGF β1 and inhibited
phosphorylation of Smad3 played key roles in this regulation.
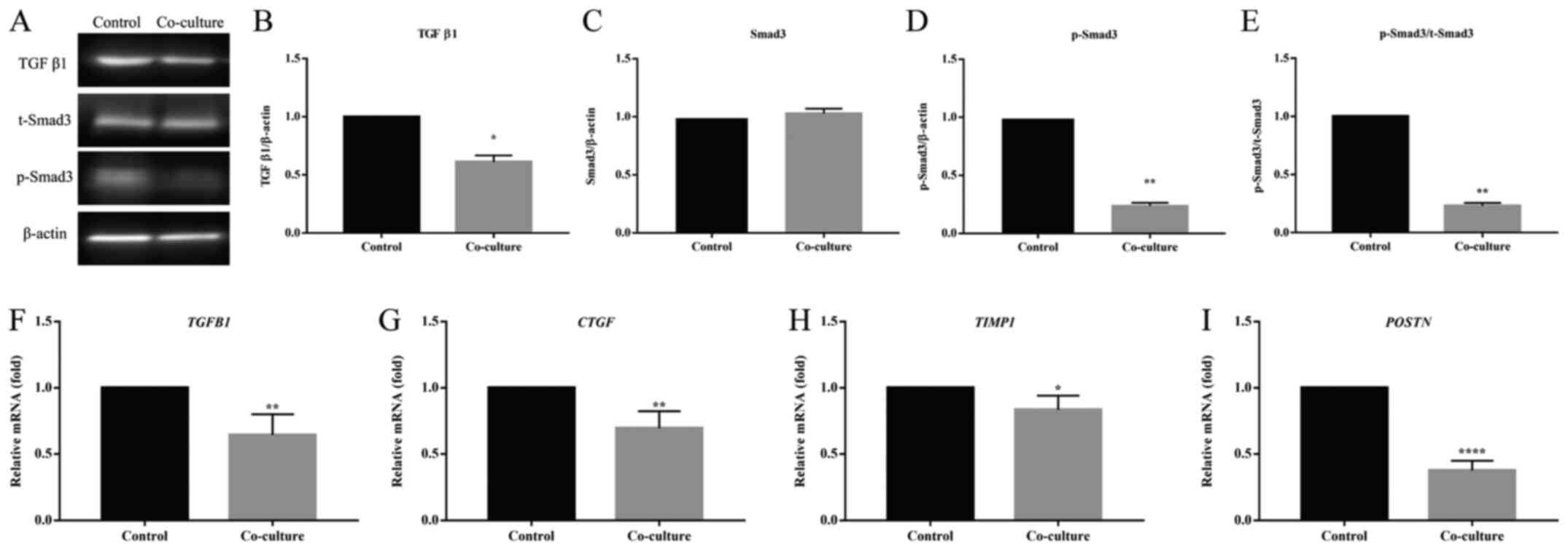 | Figure 5Effect of UCMSCs on the activation of
the TGF β1/Smad3 pathway of HSFs in a co-culture system. (A)
Western blotting was used to determine the protein levels of (B)
TGF β1, (C) t-Smad3 and (D) p-Smad3, and (E) the ratio of p-Smad3
to t-Smad3. Data are presented as the mean ± SEM. The relative mRNA
levels of (F) TGFB1, (G) CTGF, (H) TIMP1 and
(I) POSTN were detected by reverse
transcription-quantitative PCR. Data are presented as the mean ±
SD. *P<0.05, **P<0.01 and
****P<0.0001 vs. the control group. CTGF, cellular
communication network factor 2; HSF, hypertrophic scar fibroblasts;
p, phosphorylated; t, total; TGFB1, transforming growth factor β1;
TIMP1, metalloproteinase inhibitor 1; POSTN, periostin; UCMSC,
umbilical cord-derived mesenchymal stem cells. |
Discussion
Due to improvements in acute burn care, in recent
decades mortality due to extensive deep burns has significantly
decreased (19). In recent years,
HSs, which can cause severe physical and psychological problems in
patients who survive massive burns, have become the greatest unmet
challenge in burn care (1). In
order to address this problem a variety of treatments has been
developed, including surgical approaches, compressive dressing,
laser therapy and local drug injection (20). Existing studies and trials have
shown that though these methods are effective in addressing HSs,
they have limitations and can be accompanied by severe side-effects
(3,20). The development of a comprehensive
treatment for this disease remains a long and arduous task.
MSC therapy in wound healing has been widely studied
(21). Previous studies have
demonstrated that MSCs attenuate scarring through their involvement
in all overlapping scar formation phases: inflammation,
proliferation and remodeling (22).
Liu et al (23) reported
that BM-MSCs repressed HS formation through inflammatory regulation
in a rabbit model. Domergue et al (24) proposed that, in a nude mouse model,
injection of AD-MSCs produced an anti-fibrotic effect during the
remodeling phase of HS formation. However, Ding et al
(25) found that the fibrosis of
deep dermal fibroblasts was reinforced by BM-MSCs in a co-culture
model. These studies illustrate the regulatory effect of MSCs on
scar formation, and indicate that MSCs could activate normal dermal
fibroblasts and inhibit HS fibroblasts (3). Among the literature on MSCs as a
therapy for HSs, there are few reports focusing on UCMSCs. However,
there is increasing evidence to suggest that UCMSCs may inhibit
fibrosis in other fibrotic diseases (26-28).
UCMSCs are characterized by their high proliferation rate, weak
immunogenicity, non-invasive acquisition and special paracrine
factors (14-16).
When compared with other tissue-sourced MSCs, the paracrine profile
of UCMSCs contains a higher level of hepatocyte growth factor (HGF)
(29), which is defined as an
anti-fibrotic factor, and lower levels of vascular endothelial
growth factor and epidermal growth factor (16), which have been demonstrated to
promote the progression of fibrosis. These findings support the
hypothesis that UCMSCs are a promising target for treatment against
HS formation. Consistent with this hypothesis, the present study
demonstrated that UCMSCs could inhibit proliferation and migration,
as well as the expression of HS-related genes in HSFs in a
co-culture system. UCMSCs had no effect on the apoptosis of HSFs.
These results suggest that UCMSCs may be beneficial as a clinical
treatment against HS formation. Further research is required to
confirm this and will be focused on the in vivo experiments
and the clinical application of UCMSCs.
Investigation into the mechanism underlying the
anti-fibrotic function of UCMSCs suggested that UCMSCs could
regulate the TGF β1/Smad3 pathway of HSFs in vitro, through
inhibition of TGF β1 expression and the phosphorylation of Smad3.
TGF β1 plays a fundamental role in HS formation, and Smad3 acts as
a convergent node in the pathway downstream of TGF β1 receptors.
Smad3 forms a complex with Smad2 and Smad4 and functions as a
transcription factor that induces aberrant ECM deposition and
hyperactivity of fibroblasts (5-7).
Additionally, Smad2 has been reported to be involved in the
regulation of the differentiation of myofibroblasts mediated by
UCMSC-derived exosomal microRNAs (30). The mechanisms underlying the
anti-fibrotic effect of UCMSCs on HSFs must be further elucidated
in order to develop treatment against HSs.
In previous studies, MSC conditioned medium and the
isolated exosomes of MSCs have been used to stimulate target cells
in order to test the effects of MSCs in vitro (31,32).
The present study adopted a transwell co-culture system to
investigate the interaction between UCMSCs and HSFs. In
vivo, target cells are not only unilaterally subject to the
effect of MSCs, but they can also simultaneously influence the
paracrine release of MSCs via interaction in the unique
microenvironment (33). A
co-culture system provided a better in vitro simulation for
cell-cell interaction compared with the administration of
conditioned medium or isolated exomes (34,35).
However, there are still numerous problems worth studying to
explore this complex interaction, such as the mechanism of direct
cell-cell interaction and interaction in a 3D co-culture system
(36).
In conclusion, the present study indicates that
UCMSCs may play a valuable role in HS therapy by exerting an
anti-fibrotic action on HSFs, inhibiting their proliferation and
migration, and reducing the expression of HS-associated genes and
proteins. Suppression of the TGF β1/Smad3 pathway appears to be a
part of the molecular mechanism underlying this regulation.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Natural
Science Foundation of Jilin Provincial Science and Technology
Department Project (grant no. 20160101141JC).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
The study was conceived and designed by JY. XM, XG
and XC conducted the experiments and acquired the data. XM
interpreted the results and wrote the first draft of the
manuscript. JY made revisions to the final manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All protocols of the present study were subject to
approval by the Ethics Committee of The First Hospital of Jilin
University. Written consents were obtained from all participants
before surgery.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Finnerty CC, Jeschke MG, Branski LK,
Barret JP, Dziewulski P and Herndon DN: Hypertrophic scarring: The
greatest unmet challenge after burn injury. Lancet. 388:1427–1436.
2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bombaro KM, Engrav LH, Carrougher GJ,
Wiechman SA, Faucher L, Costa BA, Heimbach DM, Rivara FP and Honari
S: What is the prevalence of hypertrophic scarring following burns?
Burns. 29:299–302. 2003.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Amini-Nik S, Yousuf Y and Jeschke MG: Scar
management in burn injuries using drug delivery and molecular
signaling: Current treatments and future directions. Adv Drug Deliv
Rev. 123:135–154. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wang H, Pieper J, Peters F, van
Blitterswijk CA and Lamme EN: Synthetic scaffold morphology
controls human dermal connective tissue formation. J Biomed Mater
Res A. 74:523–532. 2005.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Cutroneo KR: TGF-beta-induced fibrosis and
SMAD signaling: Oligo decoys as natural therapeutics for inhibition
of tissue fibrosis and scarring. Wound Repair Regen. 15 (Suppl
1):S54–S60. 2007.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ashcroft GS and Roberts AB: Loss of Smad3
modulates wound healing. Cytokine Growth Factor Rev. 11:125–131.
2000.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Roberts AB, Russo A, Felici A and Flanders
KC: Smad3: A key player in pathogenetic mechanisms dependent on
TGF-beta. Ann N Y Acad Sci. 995:1–10. 2003.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Brunt KR, Weisel RD and Li RK: Stem cells
and regenerative medicine-future perspectives. Can J Physiol
Pharmacol. 90:327–335. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Li Q, Zhang C and Fu X: Will stem cells
bring hope to pathological skin scar treatment? Cytotherapy.
18:943–956. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kim KH, Blasco-Morente G, Cuende N and
Arias-Santiago S: Mesenchymal stromal cells: Properties and role in
management of cutaneous diseases. J Eur Acad Dermatol Venereol.
31:414–423. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yun IS, Jeon YR, Lee WJ, Lee JW, Rah DK,
Tark KC and Lew DH: Effect of human adipose derived stem cells on
scar formation and remodeling in a pig model: A pilot study.
Dermatol Surg. 38:1678–1688. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Lam MT, Nauta A, Meyer NP, Wu JC and
Longaker MT: Effective delivery of stem cells using an
extracellular matrix patch results in increased cell survival and
proliferation and reduced scarring in skin wound healing. Tissue
Eng Part A. 19:738–747. 2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hu L, Wang J, Zhou X, Xiong Z, Zhao J, Yu
R, Huang F, Zhang H and Chen L: Exosomes derived from human adipose
mensenchymal stem cells accelerates cutaneous wound healing via
optimizing the characteristics of fibroblasts. Sci Rep.
6(32993)2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ding DC, Chang YH, Shyu WC and Lin SZ:
Human umbilical cord mesenchymal stem cells: A new era for stem
cell therapy. Cell Transplant. 24:339–347. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
El Omar R, Beroud J, Stoltz JF, Menu P,
Velot E and Decot V: Umbilical cord mesenchymal stem cells: The new
gold standard for mesenchymal stem cell-based therapies? Tissue Eng
Part B Rev. 20:523–544. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Dabrowski FA, Burdzinska A, Kulesza A,
Sladowska A, Zolocinska A, Gala K, Paczek L and Wielgos M:
Comparison of the paracrine activity of mesenchymal stem cells
derived from human umbilical cord, amniotic membrane and adipose
tissue. J Obstet Gynaecol Res. 43:1758–1768. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Xie C, Shi K, Zhang X, Zhao J and Yu J:
MiR-1908 promotes scar formation post-burn wound healing by
suppressing Ski-mediated inflammation and fibroblast proliferation.
Cell Tissue Res. 366:371–380. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Brusselaers N, Monstrey S, Vogelaers D,
Hoste E and Blot S: Severe burn injury in Europe: A systematic
review of the incidence, etiology, morbidity, and mortality. Crit
Care. 14(R188)2010.PubMed/NCBI View
Article : Google Scholar
|
|
20
|
Friedstat JS and Hultman CS: Hypertrophic
burn scar management: What does the evidence show? A systematic
review of randomized controlled trials. Ann Plast Surg.
72:S198–S201. 2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Lee DE, Ayoub N and Agrawal DK:
Mesenchymal stem cells and cutaneous wound healing: Novel methods
to increase cell delivery and therapeutic efficacy. Stem Cell Res
Ther. 7(37)2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Jackson WM, Nesti LJ and Tuan RS:
Mesenchymal stem cell therapy for attenuation of scar formation
during wound healing. Stem Cell Res Ther. 3(20)2012.PubMed/NCBI View
Article : Google Scholar
|
|
23
|
Liu S, Jiang L, Li H, Shi H, Luo H, Zhang
Y, Yu C and Jin Y: Mesenchymal stem cells prevent hypertrophic scar
formation via inflammatory regulation when undergoing apoptosis. J
Invest Dermatol. 134:2648–2657. 2014.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Domergue S, Bony C, Maumus M, Toupet K,
Frouin E, Rigau V, Vozenin MC, Magalon G, Jorgensen C and Noël D:
Comparison between stromal vascular fraction and adipose
mesenchymal stem cells in remodeling hypertrophic scars. PLoS One.
11(e0156161)2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ding J, Ma Z, Shankowsky HA, Medina A and
Tredget EE: Deep dermal fibroblast profibrotic characteristics are
enhanced by bone marrow-derived mesenchymal stem cells. Wound
Repair Regen. 21:448–455. 2013.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Weiss DJ: Stem cells and cell therapies
for cystic fibrosis and other lung diseases. Pulm Pharmacol Ther.
21:588–594. 2008.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tsai PC, Fu TW, Chen YM, Ko TL, Chen TH,
Shih YH, Hung SC and Fu YS: The therapeutic potential of human
umbilical mesenchymal stem cells from Wharton's jelly in the
treatment of rat liver fibrosis. Liver Transpl. 15:484–495.
2009.PubMed/NCBI View
Article : Google Scholar
|
|
28
|
Alatab S, Najafi I, Atlasi R, Pourmand G,
Tabatabaei-Malazy O and Ahmadbeigi N: A systematic review of
preclinical studies on therapeutic potential of stem cells or stem
cells products in peritoneal fibrosis. Minerva Urol Nefrol.
70:162–178. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Balasubramanian S, Venugopal P, Sundarraj
S, Zakaria Z, Majumdar AS and Ta M: Comparison of chemokine and
receptor gene expression between Wharton's jelly and bone
marrow-derived mesenchymal stromal cells. Cytotherapy. 14:26–33.
2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Fang S, Xu C, Zhang Y, Xue C, Yang C, Bi
H, Qian X, Wu M, Ji K, Zhao Y, et al: Umbilical cord-derived
mesenchymal stem cell-derived exosomal MicroRNAs suppress
myofibroblast differentiation by inhibiting the transforming growth
factor-β/SMAD2 pathway during wound healing. Stem Cells Transl Med.
5:1425–1439. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Skalnikova H, Motlik J, Gadher SJ and
Kovarova H: Mapping of the secretome of primary isolates of
mammalian cells, stem cells and derived cell lines. Proteomics.
11:691–708. 2011.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Yu B, Zhang X and Li X: Exosomes derived
from mesenchymal stem cells. Int J Mol Sci. 15:4142–4157.
2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kusuma GD, Carthew J, Lim R and Frith JE:
Effect of the microenvironment on mesenchymal stem cell paracrine
signaling: Opportunities to engineer the therapeutic effect. Stem
Cells Dev. 26:617–631. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Paschos NK, Brown WE, Eswaramoorthy R, Hu
JC and Athanasiou KA: Advances in tissue engineering through stem
cell-based co-culture. J Tissue Eng Regen Med. 9:488–503.
2015.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Goers L, Freemont P and Polizzi KM:
Co-culture systems and technologies: Taking synthetic biology to
the next level. J R Soc Interface. 11: pii(20140065)2014.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Wrzesinski K and Fey SJ: From 2D to 3D-a
new dimension for modelling the effect of natural products on human
tissue. Curr Pharm Des. 21:5605–5616. 2015.PubMed/NCBI View Article : Google Scholar
|