Introduction
Renal fibrosis is caused by imbalance between
synthesis and degradation of the extracellular matrix (ECM)
constituents, including collagen I, III and IV, as a result of
various pathological factors, leading to glomerulosclerosis,
tubulointerstitial fibrosis, hyalinization and sclerosis of the
renal small vessels (1,2). These cellular and molecular events
ultimately result in parenchymal obstruction and renal failure
(3,4). Renal fibrosis can also result from a
variety of other factors that are associated with kidney injury,
including hypertension, inflammation, high sugar, high fat and drug
damage (5). However, previous
studies have shown that even if these factors are controlled
effectively, including blood pressure, blood glucose and drug
damage, the process of renal fibrosis remains difficult to prevent
(6). Therefore, investigating the
molecular mechanism underlying the occurrence and development of
fibrosis, identifying therapeutic agents and target genes that can
directly interfere with the fibrotic process has become an
important topic of study in recent years (7-9).
The formation and progression of renal fibrosis is a
complex and dynamic process that includes inflammatory cell
infiltration, fibroblast activation and proliferation, ECM
accumulation, tubular atrophy and microvascular degeneration
(10,11). A number of genes have been reported
to be involved in this process. Among them, transforming growth
factor (TGF)-β1 is considered to be the most important fibrogenic
factor, which initiates the occurrence and accelerates the progress
of fibrosis (12). In addition,
recent reports also found that a number of other factors, including
platelet-derived growth factor (PDGF), Smad3, connective tissue
growth factor and angiotensin II, enhanced the constituents of the
ECM (13,14). Although the mechanism behind the
formation and development of renal fibrosis has been extensively
studied, effective treatment strategies has yet to be found
(15,16). Therefore, it is of importance to
explore the mechanism of renal fibrosis further to find novel
potential therapeutic drugs.
Poria cocos Wolf (Polyporaceae) is a
traditional Chinese medicine that has been applied for >2,000
years in China that is also widely distributed and easy to obtain
because it exists in many plants (17,18).
Reported medicinal properties that have been associated with
Poria cocos include diuretic effects, invigorating the
spleen, tranquilizes the heart and edema elimination (19-22).
Poricoic acid A (PAA) is one of the main chemical constituents on
the surface layer of Poria cocos Wolf. PAA has been
documented to significantly alleviate oxidative stress and suppress
the increase in inflammatory factors during acute kidney injury
(23). Chen et al (24) reported that PAA exerted
renoprotective and antifibrotic effects by inhibiting TGF-β/Smad3
and Wnt/β-catenin signaling pathways, whilst other studies have
also demonstrated that PAA could suppress the renin-angiotensin
system (RAS) to prevent chronic kidney disease progression
(25,26). However, to the best of our
knowledge, the mechanistic effects of PAA on renal fibrosis remain
to be elucidated.
The present study aimed to investigate the effects
of PAA on TGF-β1-induced renal fibroblast proliferation and
fibrosis formation in addition to the potential mechanism
underlying the actions of PAA. Information from the present study
may provide a novel avenue for the treatment of fibrosis formation
during the development of renal failure.
Materials and methods
Cell culture and drug treatment
NRK-49F cells (cat. no. CRL-1570; American Type
Culture Collection), a rat renal interstitial fibroblast cell line
(27,28), were used to investigate the effects
of PAA on renal fibrosis. Cells were cultured in DMEM (cat. no.
11965-092; Gibco, Thermo Fisher Scientific, Inc.) supplemented with
10% FBS (cat. no. SH30071.02; HyClone; GE Healthcare Life Sciences)
in a 5% CO2 atmosphere at 37˚C.
To determine the optimal concentration of PAA
(Shanxi Medicine company), cells were exposed to serial
concentrations of PAA (1, 2, 5, 10, 15 and 20 µM) for 24 h at 37˚C
in a 5%CO2 incubator. The effects of PAA on renal
fibrosis were also determined using recombinant human TGF-β1 (5
ng/ml; cat. no. 240-B; R&D Systems, Inc.) in the presence or
absence of recombinant PDGF-C protein (50 ng/ml; cat. no. 1687-CC;
R&D Systems, Inc.) for 24 h at 37˚C in a 5% CO2
incubator.
Cell growth assay
NRK-49F cells were cultured in 96-well plates, 100
µl per well with 1x104 cells and exposed to 10 µmol PAA
for 24 h. Subsequently, 30 µl CCK-8 solution (Dojindo Molecular
Technologies, Inc.) was added to each well where the cells were
incubated for 4 h at 37˚C. The optical density (OD) value was
detected at a wavelength of 450 nm where a microplate reader
(Thermo Fisher Scientific, Inc.) was used to calculate cell
viability and inhibition rate. The detailed calculations are as
follows: Cell viability (%)=[(As-Ab)/(Ac-Ab)]x100; inhibition rate
(%)=[(Ac-As)/(Ac-Ab)] x100; As=OD value of the experimental wells;
Ab=OD value of blank wells; Ac=OD value of control wells.
Western blotting
The expression of PDGF-C/Smad3/MAPK or ECM related
proteins was analyzed by western blotting. Total protein was
isolated from NRK-49F cells with or without PAA treatments using
radioimmunoprecipitation assay (RIPA) buffer (cat. no. 89900;
Thermo Fisher Scientific, Inc.). Protein levels were quantified
using a BCA protein assay kit (Thermo Fisher Scientific, Inc.). A
total of 30 µg protein was loaded in each lane and subsequently run
on 12% SDS-PAGE gels. The separated proteins were subsequently
transferred to PVDF membranes. Then, the membranes were washed
three times with 0.1% Tween-20 TBS solution (TBST) and incubated
for 1 h at room temperature with 5% of non-fat milk (cat. no.
1706404; Bio-rad, Laboratories, Inc.). Subsequently, the following
primary antibodies were used at 1:1,000 dilution: E-cadherin (cat.
no. 14472S; Cell Signaling Technology, Inc.), collagen I (cat. no.
84336S; Cell Signaling Technology, Inc.), collagen IV (cat. no.
50273S; Cell Signaling Technology, Inc.), alpha smooth muscle actin
(α-SMA; cat. no. ab5694; Abcam), fibronectin (FN; cat. no. 26836S;
Cell Signaling Technology, Inc.), PDGF-C (cat. no. abc1392;
Sigma-Aldrich; Merck KGaA), phosphorylated (p)-Smad3 (cat. no.
9520; Cell Signaling Technology, Inc.), Smad3 (cat. no. 9523; Cell
Signaling Technology, Inc.), p-ERK1/2 (cat. no. 4370; Cell
Signaling Technology, Inc.), ERK1/2 (cat. no. 4695; Cell Signaling
Technology, Inc.), p-p38 (cat. no. 4511; Cell Signaling Technology,
Inc.), p38 (cat. no. 8690; Cell Signaling Technology, Inc.) and
GAPDH (cat. no. sc-137179; Santa Cruz Biotechnology, Inc.).
Following incubation with the respective primary antibodies
overnight at 4˚C, the membranes were washed three times with TBST
solution. Subsequently, the membranes were exposed to horseradish
peroxidase-conjugated goat anti-rabbit (1:5,000; cat. no. ab6721;
Abcam), goat anti-mouse (1:5,000; cat. no. ab05719; Abcam), and
rabbit anti-goat (1:5,000; cat. no. ab5755; Abcam) secondary
antibodies for 1 h at room temperature prior to treatment with ECL
reagent (cat. no. RPN2232; GE Healthcare), using autoluminography
to visualize protein bands. Protein expression was then quantified
using ImageJ (version 1.48V; National Institutes of Health) with
GAPDH as the loading control.
Cell proliferation assay
To assess kidney cell proliferation,
2x105 NRK-49F cells/per well in 1ml culture medium were
plated onto coverslips in a 6-well plate and allowed to grow
overnight at 37˚C in a 5% CO2 incubator. Cells were
incubated with Alexa Fluor 555 labeled with
5-ethynyl-2'-deoxyuridine (EdU; cat. no. A10044; Invitrogen; Thermo
Fisher Scientific, Inc.) at a final concentration of 10 µM for 24 h
at 37˚C in 5% CO2 before being harvested and subjected
to treatments in accordance with the Click-iT®EdU Alexa
Fluor® 555 Imaging kit (cat. no. C10353; Invitrogen;
Thermo Fisher Scientific, Inc., cat. no. C10353). All procedures
were performed according to the manufacturer's instructions. Nuclei
were stained with DAPI at final a concentration of 3 µM.
EdU-positive cells with DAPI-labeled nuclei were counted to
evaluate cell proliferation.
Statistical analysis
Data are presented as the mean ± SD. Statistical
analysis was performed using GraphPad Prism 6.0 (GraphPad Software,
Inc.). One-way ANOVA followed by Tukey's post hoc test was used for
comparisons between multiple groups. P<0.05 was considered to
indicate a statistically significant difference. All data presented
are from at least three independent experiments.
Results
In vitro screening for the optimal
concentration of PAA
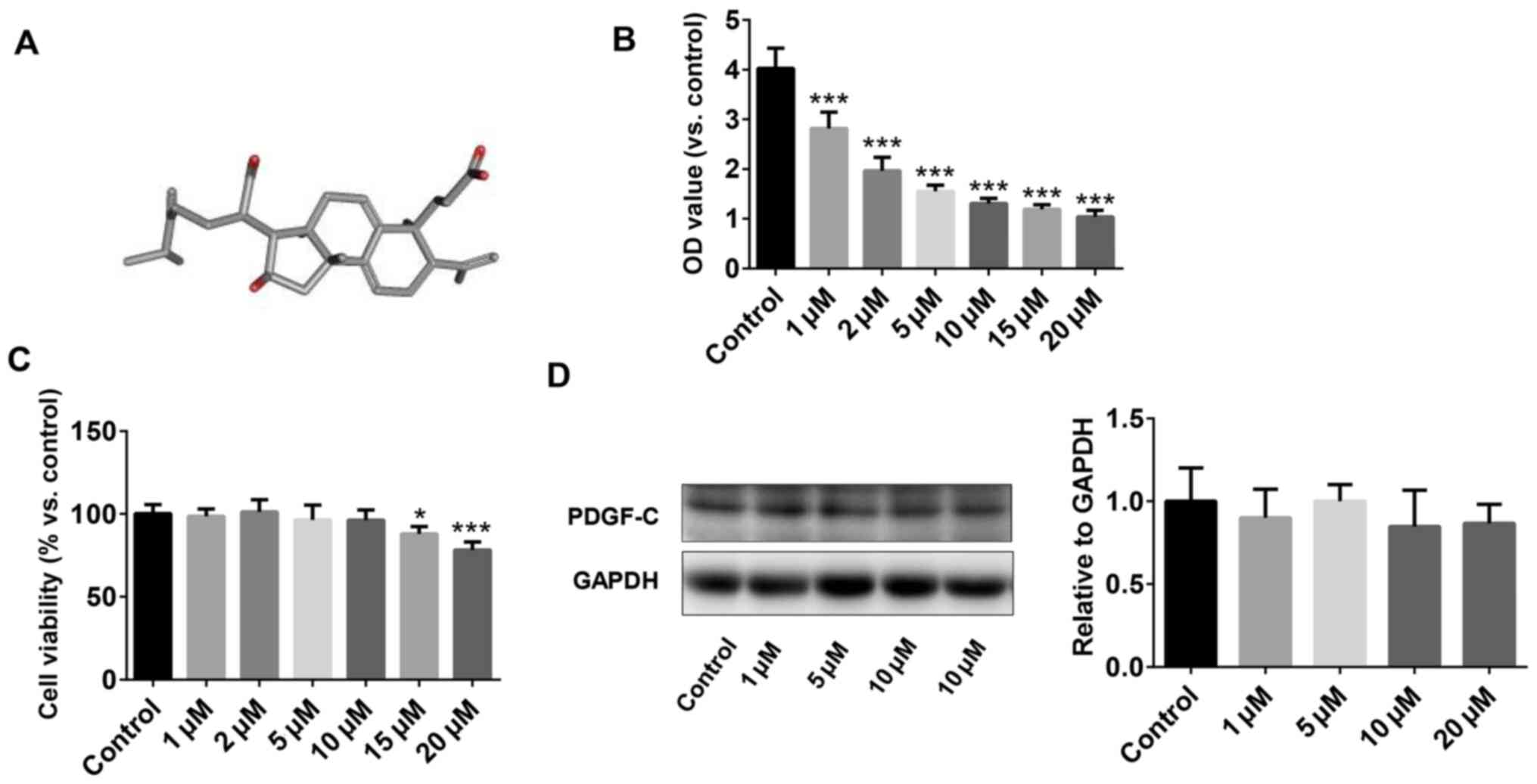
The chemical structure of PAA extracted from the
epidermis of Poria cocos is shown in Fig. 1A. The effect of PAA on cell
viability was measured using CCK-8 assay. To test the optimal
concentration of PAA on cell growth, serial concentrations of PAA,
1-20 µM final concentration were used. The results revealed that
concentrations of PAA >1 µM reduced NRK-49F cell viability in a
dose-dependent manner. However, the inhibitory effect did not
increase further when the concentration of PAA applied was >10
µM (Fig. 1B). The CCK-8 assay
results demonstrated that PAA exerted no cytotoxic effects on
NRK-49F cells at concentrations <10 µM, but cell viability was
significantly reduced following treatment with 15 and 20 µM PAA
(Fig. 1C). Therefore, 10 µM PAA was
applied for subsequent experiments. The effects of different
concentrations of PAA on PDGF-C protein expression were explored
further by western blotting (Fig.
1D). PAA did not exert significant effects on PDGF-C protein
expression at all concentrations tested compared with that of the
control group.
PAA inhibits TGF-β1-induced ECM
accumulation, fibrosis formation and proliferation by NRK-49F
cells
To explore the effects of PAA on TGF-β1-induced
NRK-49F cell ECM accumulation and fibrosis formation, western
blotting was performed to examine the expression of E-cadherin,
collagen I, α-SMA, FN and collagen IV, proteins associated with ECM
and fibrosis. The results showed that PAA treatment alone had no
significant effect on the expression levels of these proteins
compared with those of the control group. In contrast, TGF-β1 alone
decreased the expression of E-cadherin and increased the expression
of collagen I, α-SMA, FN and collagen IV. However, in the presence
of TGF-β1, PAA significantly increased E-cadherin expression and
significantly reducing those of collagen I, α-SMA, FN and collagen
IV in NRK-49F cells compared with their expression in cells
following TGF-b1 treatment alone (Fig.
2A). In addition, EdU staining results confirmed that the
TGF-β1-induced NRK-49F cell proliferation was significantly
attenuated by PAA (Fig. 2B).
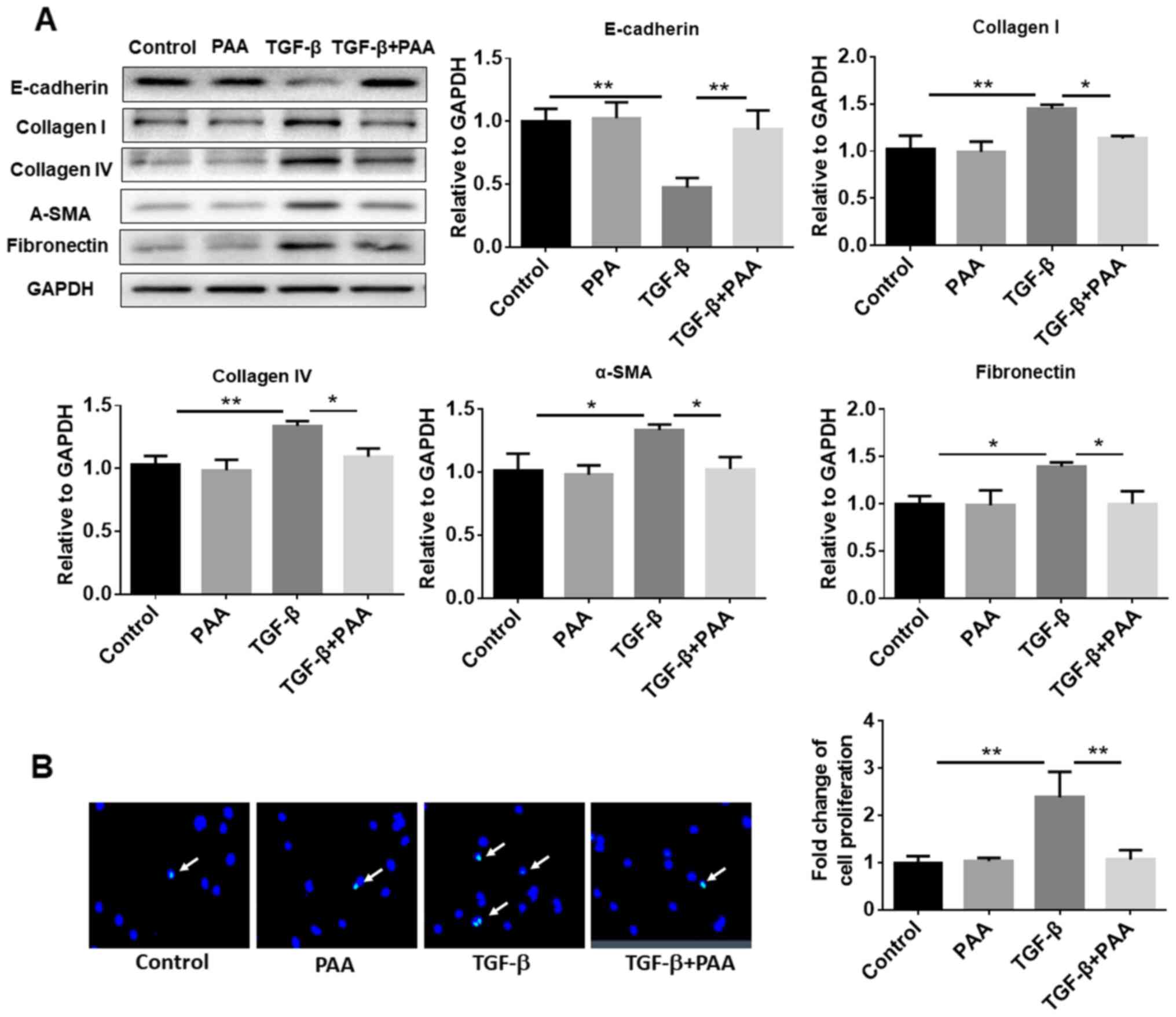 | Figure 2Effect of PAA on the expression of
ECM proteins and TGF-β1-induced cell proliferation. (A) Expression
of E-cadherin, collagen I, collagen IV, α-SMA and fibronectin,
proteins associated with ECM, were measured by western blotting
following treatment with PAA, TGF-β1 or a combination of both PAA
and TGF-β1. (B) Cell proliferation was measured after TGF-β1 and/or
PAA treatment using 5-ethynyl-2'-deoxyuridine staining. n=3.
*P<0.05 and **P<0.01. Magnification
x40. PAA, poricoic acid A; TGF-β1, transforming growth factor-β1;
α-SMA, α-smooth muscle actin; ECM, extracellular matrix. |
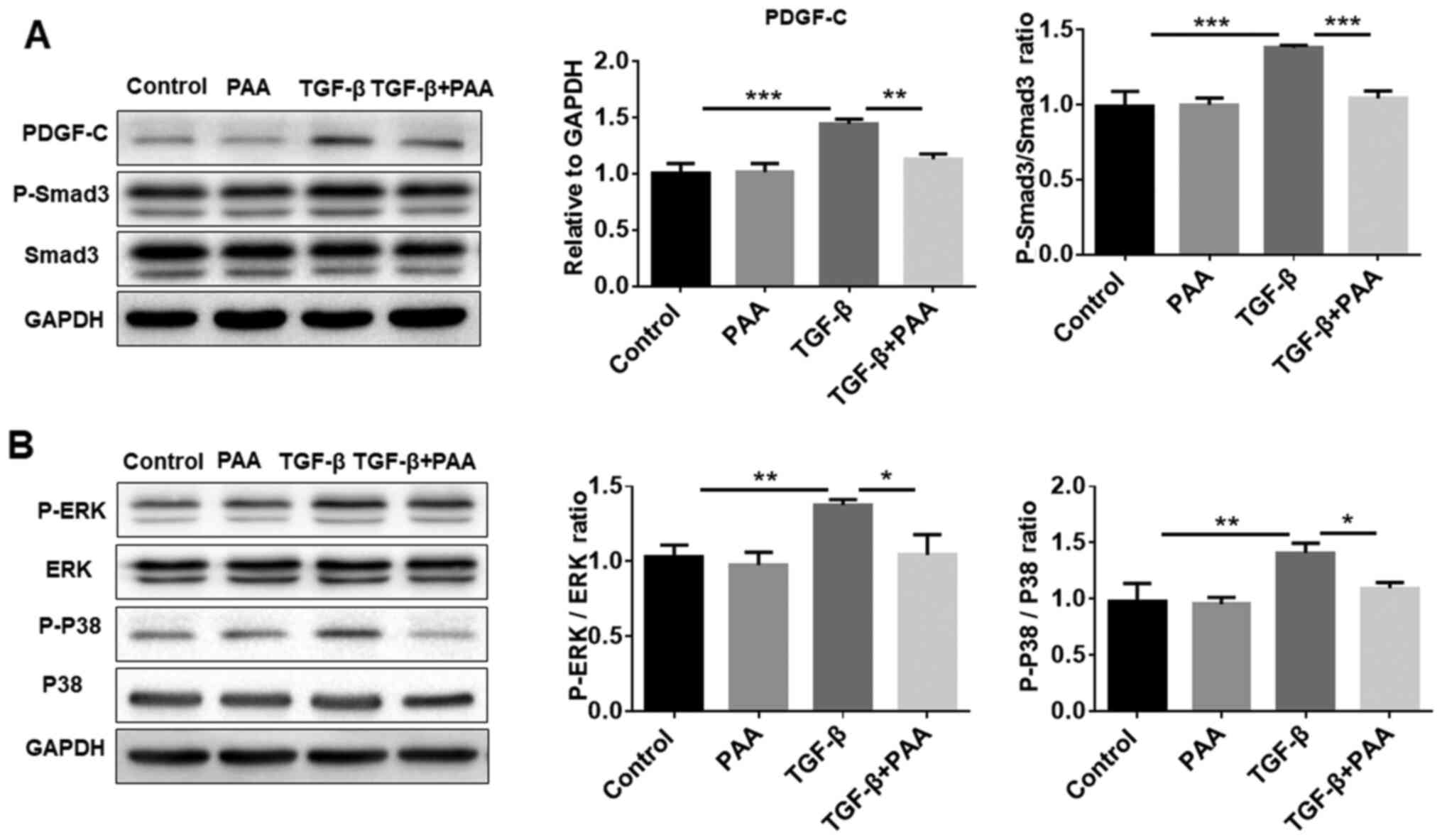
PDGF-C and Smad3/MAPK pathways are
suppressed by PAA treatment in NRK-49F cells
Previous studies have shown that PDGF-C serves an
important role in renal fibrosis as a mediator of renal
interstitial fibrosis, where the Smad3 and MAPK pathways are
reported to be involved (29,30).
To explore further whether PAA can regulate the PDGF-C, Smad3 and
MAPK signaling pathways in NRK-49F cells, western blotting was
performed to examine Smad3 and MAPK phosphorylation. The results
showed that PAA alone did not significantly alter PDGF-C and
p-Smad3 levels compared with those in the control group; however,
PAA treatment significantly reversed TGF-β1-induced PDGF-C
expression, Smad3 (Fig. 3A), ERK1/2
and P38 MAPK phosphorylation (Fig.
3B). These data suggest that PAA can inhibit TGF-β1-induced
activation of the Smad and MAPK signaling pathways.
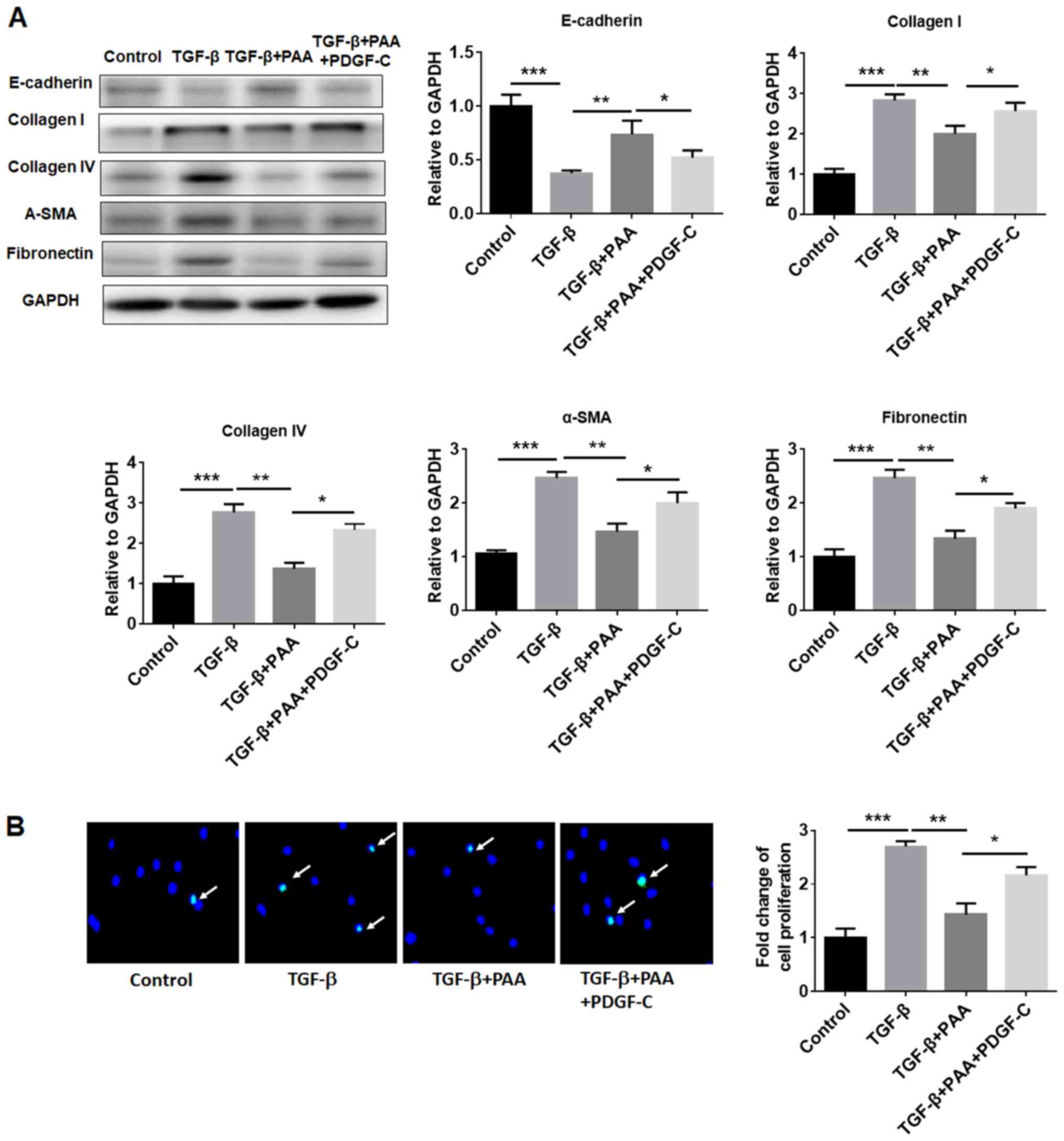
PDGF-C reverses the inhibitory effects
of PAA on TGF-β1-stimulated NRK-49F cells
To evaluate the effects of PDGF-C on the inhibitory
effects of PAA on NRK-49F cell ECM accumulation, fibrosis formation
and proliferation further, western blotting and EdU
immunofluorescence staining were performed. The western blotting
results revealed that the inhibitory effects of PAA on
TGF-β1-induced renal fibroblast ECM accumulation and fibrosis
formation was significantly reversed by PDGF-C treatment compared
with those observed in the TGF-β1+PAA group (Fig. 4A). PDGF-C treatment also reversed
the inhibitory effects of PAA on TGF-β1-induced NRK-49F cell
proliferation (Fig. 4B).
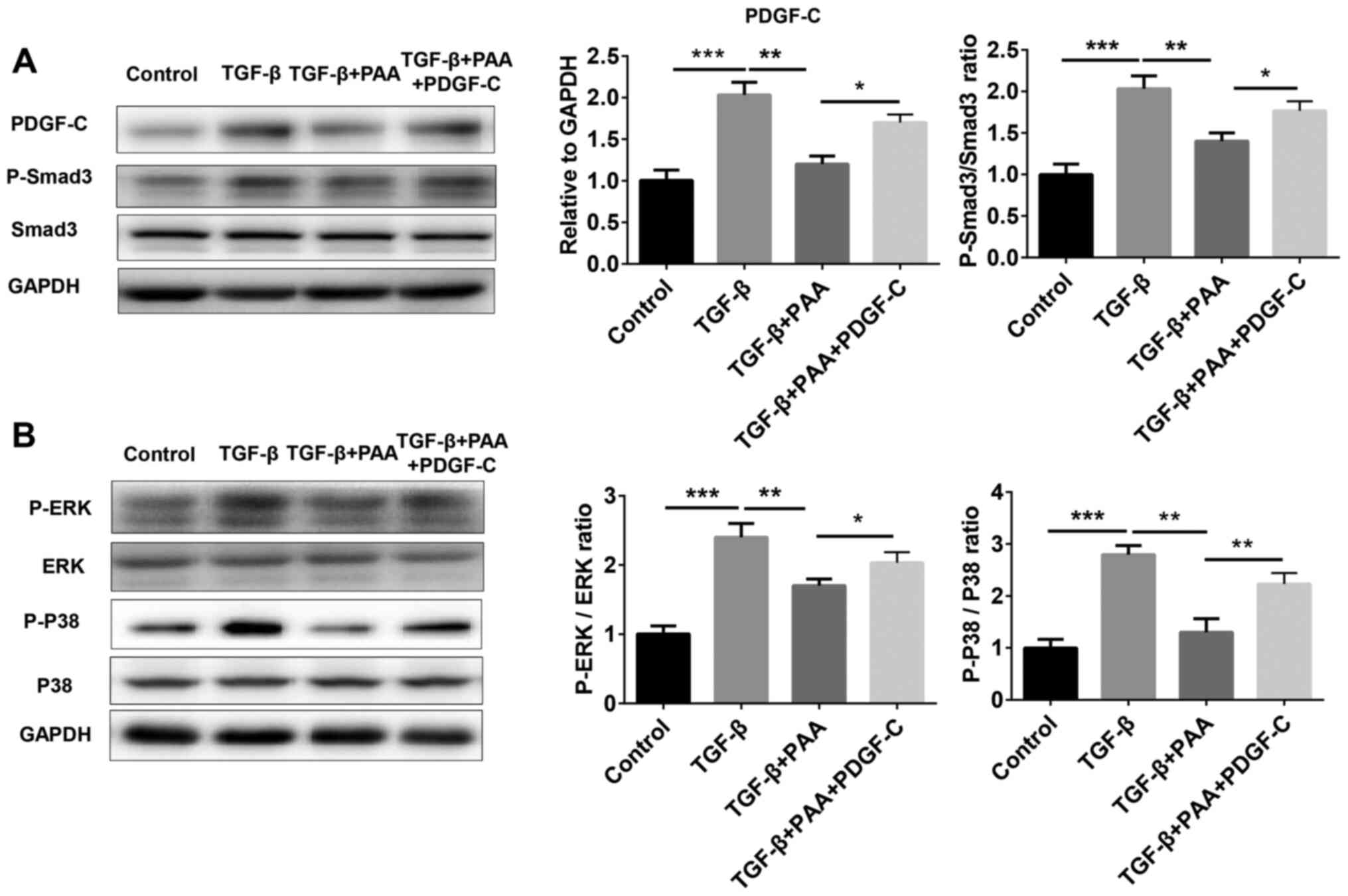
PDGF-C treatment eliminates the
inhibition of TGF-β1-stimulatedSmad3 and MAPK signaling by PAA in
NRK-49F cells
To further examine whether PDGF-C is involved in the
inhibitory role of PAA on the Smad3 and MAPK signaling activation
induced by TGF-β1, the expression of proteins associated with the
Smad3 and MAPK pathways were measured by western blotting in cells
treated with or without TGF-β1, PAA and PDGF-C. The results showed
that the increased expression of proteins associated with the Smad3
and MAPK pathway induced by TGF-β1 was significantly reduced after
PAA treatment (Fig. 5A). By
contrast, additive PDGF-C treatment significantly increased the
expression of these proteins compared with that in the TGF-β1+PAA
groups (Fig. 5).
Discussion
Since renal interstitial fibrosis serves a role in
the development of renal failure, the degree of fibrotic
development is closely associated with the prognosis of chronic
kidney disease and renal failure (31). Therefore, research on the prevention
and treatment of renal interstitial fibrosis has garnered attention
over recent years (32).
Traditional Chinese medicine has also made significant progress in
the study of renal fibrosis treatment (33). In the present study, PAA
significantly inhibited TGF-β1-induced renal fibroblast
proliferation, ECM production and fibrosis formation. The present
results suggested that PAA serves a role in the progression of
renal fibrosis.
Among the factors associated with renal interstitial
fibrosis, TGF-β1 has been most extensively studied (34). A number of studies have shown that a
variety of cytokines and growth factors participate in the
occurrence and development of renal interstitial fibrosis, of which
TGF-β1 appeared to serve the most significant role (35,36).
The biological functions of TGF-β1 in kidney development include
promotion of ECM accumulation and synthesis of matrix components
(37). TGF-β1 has also been
implicated in epithelial-mesenchymal transition, whose markers
include FN, E-cadherin, vimentin and α-SMA (38). Although several clinical trials
targeting TGF-β1 using anti-TGFβ1 antibodies in kidney disease as
well as animal model have been performed, no significant
improvement in the symptoms have been observed in either patients
or mouse models (39-41).
Therefore, the present study used a rat renal fibroblast cell line
treated with TGF-β1 as a cell model. TGF-β1 significantly promoted
fibroblast proliferation and the expression of fibrosis-related
proteins. PAA alone did not alter the expression of these proteins,
including collagen I, collagen IV, α-SMA, FN, PDGF-C, as well as
the phosphorylation of Smad3, ERK and p38, but it could
significantly suppress the expressions of these proteins induced by
TGF-β1. Other previous studies suggested that PAA could suppress
renal fibrosis via inhibiting the functions of TGF-β1 via the
RAS/TGF-β/Smad axis (26) or the
Wnt/β-catenin pathway (25). The
current results showed that PAA extenuated renal cell proliferation
through suppressing the Smad3/MAPK pathway. These results can be
compared with those obtained in animal models, such as unilateral
ureteral obstruction (42) or 5/6
nephrectomy mice (43), where
alleviation of renal fibrosis was mediated through the inhibition
of the TGF-β/Smad and Wnt/β-catenin signaling pathways in the
kidneys or RAS. In addition, other proteins associated with renal
fibrosis, including angiotensinogen, renin, angiotensin-converting
enzymes and tubular angiotensin II type 1 receptors (25) can be measured in clinical patients
in future studies. The results of the present study showed that the
inhibitory effects of PAA on TGF-β1-induced renal fibroblast
proliferation were mediated by acting the Smad3/MAPK pathway.
PDGF-C, originally identified in platelets, has a
variety of documented biological functions (44). Previous studies demonstrated that
PDGF-C expression in renal interstitial fibrosis was significantly
increased during progression to renal fibrosis (45,46).
Inhibition of PDGF-C activity can significantly inhibit the
development of renal fibrosis (46,47).
In the present study, although PAA alone did not affect PDGF-C
expression in NRK-49F cells, PDGF-C significantly reversed the
inhibitory effects of PAA in the presence of TGF-β1. By contrast,
treatment with PDGF-C can partially reverse the effects of PAA in
TGF-β1-induced renal fibroblasts. The present study reported that
PDGF-C stimulated mesenchymal cells proliferation through binding
to the PDGF-α receptor (44). The
interaction between proximal tubule cells and adjacent fibroblasts
is critical for renal fibrosis (48). NRK-49F cells were used in the
present study since it exhibits a characteristic fibroblast
spindle-like morphology that is easy to monitor in response to PDGF
(27,28). These results suggested that PAA
treatment inhibited the proliferation, ECM aggregation and fibrosis
of renal fibroblasts by inhibiting the functions of TGF-β1.
However, the inhibitory effects could be reversed by PDGF-C.
Previous studies have revealed that Smad3 and MAPK
signaling pathways are involved in the development of renal
fibrosis (49,50). The expression of Smad3 and MAPK is
significantly activated during renal fibrosis (50,51)
and the inhibition of Smad and MAPK pathways has been reported to
significantly reduce renal ECM aggregation and fibrosis formation
(52). The present study showed
that PAA could suppress the increased upregulation of Smad and MAPK
signaling induced by TGF-β1, which were reversed by the presence of
PDGF-C.
In conclusion, data from the present study suggest
that PAA treatment suppressed TGF-β1-induced renal fibroblast ECM
accumulation, fibrosis formation and proliferation by inhibiting
the PDGF-C, Smad3 and MAPK signaling pathways. PDGF-C can block the
effect of PAA on TGF-β1-induced fibroblasts and promoted the
activation of the Smad and MAPK pathways induced by TGF-β1. These
findings revealed the potential of PAA application in protecting
against the fibrosis of renal fibroblasts. The present study also
provided a new insight into the prevention of fibrosis formation
through regulation by PDGF-C. However, further in vivo
studies are necessary to fully clarify the role of PAA in renal
fibrosis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
QL and GW conceived and designed the experiment. QL,
YM and HJ performed experiments and collected and analyzed the
data. QL and GW wrote the manuscript. All authors revised and
approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shiber S, Eliakim-Raz N and Yair M:
Retroperitoneal fibrosis: Case series of five patients and review
of the literature. Rev Bras Reumatol Engl Ed. 56:101–104.
2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Zhou D and Liu Y: Renal fibrosis in 2015:
Understanding the mechanisms of kidney fibrosis. Nat Rev Nephrol.
12:68–70. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hu C, Sun L, Xiao L, Han Y, Fu X, Xiong X,
Xu X, Liu Y, Yang S, Liu F and Kanwar YS: Insights into the
mechanisms involved in the expression and regulation of
extracellular matrix proteins in diabetic nephropathy. Curr Med
Chem. 22:2858–2870. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wang J, Yang Q, Nie Y, Guo H, Zhang F,
Zhou X and Yin X: Tetrahydrobiopterin contributes to the
proliferation of mesangial cells and accumulation of extracellular
matrix in early-stage diabetic nephropathy. J Pharm Pharmacol.
69:182–190. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Klinkhammer BM, Goldschmeding R, Floege J
and Boor P: Treatment of renal fibrosis-turning challenges into
opportunities. Adv Chronic Kidney Dis. 24:117–129. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sandner P and Stasch JP: Anti-fibrotic
effects of soluble guanylate cyclase stimulators and activators: A
review of the preclinical evidence. Respir Med. 122 (Suppl
1):S1–S9. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Wang Y, Cai J, Tang C and Dong Z:
Mitophagy in acute kidney injury and kidney repair. Cells. 9:
pii(E338)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Tang J, Goldschmeding R, Samarakoon R and
Higgins PJ: Protein phosphatase Mg(2+)/Mn(2+) dependent-1A and PTEN
deregulation in renal fibrosis: Novel mechanisms and co-dependency
of expression. FASEB J. 34:2641–2656. 2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Marko L, Park JK, Henke N, Rong S, Balogh
A, Klamer S, Bartolomaeus H, Wilck N, Ruland J, Forslund SK, et al:
B-cell lymphoma/leukemia 10 (Bcl10) and angiotensin II-induced
kidney injury. Cardiovasc Res. 116:1059–1070. 2019.
|
|
10
|
Li O, Ma Q, Li F, Cai GY, Chen XM and Hong
Q: Progress of small ubiquitin-related modifiers in kidney
diseases. Chin Med J (Engl). 132:466–473. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Genovese F, Manresa AA, Leeming DJ,
Karsdal MA and Boor P: The extracellular matrix in the kidney: A
source of novel non-invasive biomarkers of kidney fibrosis?
Fibrogenesis Tissue Repair. 7(4)2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Castellone MD and Laukkanen MO: TGF-beta1,
WNT, and SHH signaling in tumor progression and in fibrotic
diseases. Front Biosci (Schol Ed). 9:31–45. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
van Roeyen CRC, Martin IV, Drescher A,
Schuett KA, Hermert D, Raffetseder U, Otten S, Buhl EM, Braun GS,
Kuppe C, et al: Identification of platelet-derived growth factor C
as a mediator of both renal fibrosis and hypertension. Kidney Int.
95:1103–1119. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ghayur A, Padwal MK, Liu L, Zhang J and
Margetts PJ: SMAD3-dependent and-independent pathways in glomerular
injury associated with experimental glomerulonephritis. Am J
Physiol Renal Physiol. 317:F152–F62. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Breyer MD and Susztak K: The next
generation of therapeutics for chronic kidney disease. Nat Rev Drug
Discov. 15:568–588. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Mencke R, Olauson H and Hillebrands JL:
Effects of Klotho on fibrosis and cancer: A renal focus on
mechanisms and therapeutic strategies. Adv Drug Deliv Rev.
121:85–100. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Li S, Wang Z, Gu R, Zhao Y, Huang W, Wang
Z and Xiao W: A new epidioxy-tetracyclic triterpenoid from Poria
cocos Wolf. Nat Prod Res. 30:1712–1717. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Lee SR, Lee S, Moon E, Park HJ, Park HB
and Kim KH: Bioactivity-guided isolation of anti-inflammatory
triterpenoids from the sclerotia of Poria cocos using
LPS-stimulated Raw264.7 cells. Bioorg Chem. 70:94–99.
2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Li S, Zhang J, Li S, Liu C, Liu S and Liu
Z: Extraction and separation of lactate dehydrogenase inhibitors
from Poria cocos (Schw.) Wolf based on a hyphenated
technique and in vitro methods. J Sep Sci. 40:1773–1783.
2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhao YY, Feng YL, Du X, Xi ZH, Cheng XL
and Wei F: Diuretic activity of the ethanol and aqueous extracts of
the surface layer of Poria cocos in rat. J Ethnopharmacol.
144:775–778. 2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Feng YL, Lei P, Tian T, Yin L, Chen DQ,
Chen H, Mei Q, Zhao YY and Lin RC: Diuretic activity of some
fractions of the epidermis of Poria cocos. J Ethnopharmacol.
150:1114–1118. 2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhao YY, Feng YL, Bai X, Tan XJ, Lin RC
and Mei Q: Ultra performance liquid chromatography-based
metabonomic study of therapeutic effect of the surface layer of
Poria cocos on adenine-induced chronic kidney disease
provides new insight into anti-fibrosis mechanism. PLoS One.
8(e59617)2013.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Chen DQ, Feng YL, Chen L, Liu JR, Wang M,
Vaziri ND and Zhao YY: Poricoic acid A enhances melatonin
inhibition of AKI-to-CKD transition by regulating
Gas6/AxlNFkappaB/Nrf2 axis. Free Radic Biol Med. 134:484–497.
2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen DQ, Cao G, Zhao H, Chen L, Yang T,
Wang M, Vaziri ND, Guo Y and Zhao YY: Combined melatonin and
poricoic acid A inhibits renal fibrosis through modulating the
interaction of Smad3 and beta-catenin pathway in AKI-to-CKD
continuum. Ther Adv Chronic Dis.
10(2040622319869116)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wang M, Chen DQ, Chen L, Cao G, Zhao H,
Liu D, Vaziri ND, Guo Y and Zhao YY: Novel inhibitors of the
cellular renin-angiotensin system components, poricoic acids,
target Smad3 phosphorylation and Wnt/β-catenin pathway against
renal fibrosis. Br J Pharmacol. 175:2689–2708. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang M, Chen DQ, Chen L, Liu D, Zhao H,
Zhang ZH, Vaziri ND, Guo Y, Zhao YY and Cao G: Novel RAS Inhibitors
Poricoic Acid ZG and Poricoic Acid ZH attenuate renal fibrosis via
a Wnt/β-catenin pathway and targeted phosphorylation of smad3
Signaling. J Agric Food Chem. 66:1828–1842. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tan RJ, Zhou D and Liu Y: Signaling
crosstalk between tubular epithelial cells and interstitial
fibroblasts after kidney injury. Kidney Dis (Basel). 2:136–144.
2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Tang N, Cunningham K and Enger MD: TGF
beta elicits opposite responses in clonal subpopulations of NRK-49F
cells. Exp Cell Res. 196:13–19. 1991.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Floege J, Eitner F and Alpers CE: A new
look at platelet-derived growth factor in renal disease. J Am Soc
Nephrol. 19:12–23. 2008.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Huang C, Day ML, Poronnik P, Pollock CA
and Chen XM: Inhibition of KCa3.1 suppresses TGF-β1 induced MCP-1
expression in human proximal tubular cells through Smad3, p38 and
ERK1/2 signaling pathways. Int J Biochem Cell Biol. 47:1–10.
2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu M, Ning X, Li R, Yang Z, Yang X, Sun S
and Qian Q: Signalling pathways involved in hypoxia-induced renal
fibrosis. J Cell Mol Med. 21:1248–1259. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Waasdorp M, de Rooij DM, Florquin S,
Duitman J and Spek CA: Protease-activated receptor-1 contributes to
renal injury and interstitial fibrosis during chronic obstructive
nephropathy. J Cell Mol Med. 23:1268–1279. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Liu W, Lin S, Cai Q, Zhang L, Shen A, Chen
Y, Chu J and Peng J: Qingxuan jiangya decoction mitigates renal
interstitial fibrosis in spontaneously hypertensive rats by
regulating transforming growth factor-β1/Smad signaling pathway.
Evid Based Complement Alternat Med. 2017(1576328)2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lawson JS, Liu HH, Syme HM, Purcell R,
Wheeler-Jones CPD and Elliott J: The cat as a naturally occurring
model of renal interstitial fibrosis: Characterisation of primary
feline proximal tubular epithelial cells and comparative
pro-fibrotic effects of TGF-β1. PLoS One.
13(e0202577)2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Xu J, Yu TT, Zhang K, Li M, Shi HJ, Meng
XJ, Zhu LS and Zhu LK: HGF alleviates renal interstitial fibrosis
via inhibiting the TGF-β1/SMAD pathway. Eur Rev Med Pharmacol Sci.
22:7621–7627. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Loeffler I: MKP2 suppresses TGF-β1-induced
epithelial-to-mesenchymal transition through JNK inhibition. Clin
Sci (Lond). 133:545–550. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Okuda S, Languino LR, Ruoslahti E and
Border WA: Elevated expression of transforming growth factor-beta
and proteoglycan production in experimental glomerulonephritis.
Possible role in expansion of the mesangial extracellular matrix. J
Clin Invest. 86:453–462. 1990.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Choi HI, Kim DH, Park JS, Kim IJ, Kim CS,
Bae EH, Ma SK, Lee TH and Kim SW: Peroxiredoxin V (PrdxV)
negatively regulates EGFR/Stat3-mediated fibrogenesis via a
Cys48-dependent interaction between PrdxV and Stat3. Sci Rep.
9(8751)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Vincenti F, Fervenza FC, Campbell KN, Diaz
M, Gesualdo L, Nelson P, Praga M, Radhakrishnan J, Sellin L, Singh
A, et al: A phase 2, double-blind, placebo-controlled, randomized
study of fresolimumab in patients with steroid-resistant primary
focal segmental glomerulosclerosis. Kidney Int Rep. 2:800–810.
2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Trachtman H, Fervenza FC, Gipson DS,
Heering P, Jayne DR, Peters H, Rota S, Remuzzi G, Rump LC, Sellin
LK, et al: A phase 1, single-dose study of fresolimumab, an
anti-TGF-β antibody, in treatment-resistant primary focal segmental
glomerulosclerosis. Kidney Int. 79:1236–1243. 2011.PubMed/NCBI View Article : Google Scholar
|
|
41
|
McGaraughty S, Davis-Taber RA, Zhu CZ,
Cole TB, Nikkel AL, Chhaya M, Doyle KJ, Olson LM, Preston GM,
Grinnell CM, et al: Targeting Anti-TGF-β therapy to fibrotic
kidneys with a dual specificity antibody approach. J Am Soc
Nephrol. 28:3616–3626. 2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Zhang ZH, He JQ, Zhao YY, Chen HC and Tan
NH: Asiatic acid prevents renal fibrosis in UUO rats via promoting
the production of 15d-PGJ2, an endogenous ligand of PPAR-γ. Acta
Pharmacol Sin. 41:373–382. 2020.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Bian X, Bai Y, Su X, Zhao G, Sun G and Li
D: Knockdown of periostin attenuates 5/6 nephrectomy-induced
intrarenal renin-angiotensin system activation, fibrosis, and
inflammation in rats. J Cell Physiol. 234:22857–22873.
2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Li X, Ponten A, Aase K, Karlsson L,
Abramsson A, Uutela M, Bäckström G, Hellström M, Boström H, Li H,
et al: PDGF-C is a new protease-activated ligand for the PDGF
alpha-receptor. Nat Cell Biol. 2:302–309. 2000.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Eitner F, Bucher E, van Roeyen C, Kunter
U, Rong S, Seikrit C, Villa L, Boor P, Fredriksson L, Bäckström G,
et al: PDGF-C is a proinflammatory cytokine that mediates renal
interstitial fibrosis. J Am Soc Nephrol. 19:281–289.
2008.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Boor P, Babickova J, Steegh F, Hautvast P,
Martin IV, Djudjaj S, Nakagawa T, Ehling J, Gremse F, Bücher E, et
al: Role of platelet-derived growth factor-CC in capillary
rarefaction in renal fibrosis. Am J Pathol. 185:2132–2142.
2015.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Wang Y, Abu-Asab MS, Yu CR, Tang Z, Shen
D, Tuo J, Li X and Chan CC: Platelet-derived growth factor (PDGF)-C
inhibits neuroretinal apoptosis in a murine model of focal retinal
degeneration. Lab Invest. 94:674–682. 2014.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Chen CL, Chou KJ, Lee PT, Chen YS, Chang
TY, Hsu CY, Huang WC, Chung HM and Fang HC: Erythropoietin
suppresses epithelial to mesenchymal transition and intercepts Smad
signal transduction through a MEK-dependent mechanism in pig kidney
(LLC-PK1) cell lines. Exp Cell Res. 316:1109–1118. 2010.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Grynberg K, Ma FY and Nikolic-Paterson DJ:
The JNK signaling pathway in renal fibrosis. Front Physiol.
8(829)2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Xu ZJ, Shu S, Li ZJ, Liu YM, Zhang RY and
Zhang Y: Liuwei Dihuang pill treats diabetic nephropathy in rats by
inhibiting of TGF-β/SMADS, MAPK, and NF-kB and upregulating
expression of cytoglobin in renal tissues. Medicine (Baltimore).
96(e5879)2017.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Nakagawa T, Lan HY, Glushakova O, Zhu HJ,
Kang DH, Schreiner GF, Böttinger EP, Johnson RJ and Sautin YY: Role
of ERK1/2 and p38 mitogen-activated protein kinases in the
regulation of thrombospondin-1 by TGF-beta1 in rat proximal tubular
cells and mouse fibroblasts. J Am Soc Nephrol. 16:899–904.
2005.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Okano K, Hibi A, Miyaoka T, Inoue T,
Sugimoto H, Tsuchiya K, Akiba T and Nitta K: Inhibitory effects of
the transcription factor Ets-1 on the expression of type I collagen
in TGF-β1-stimulated renal epithelial cells. Mol Cell Biochem.
369:247–254. 2012.PubMed/NCBI View Article : Google Scholar
|