Introduction
Lung cancer is one of the most common causes of
death, accounting for ~26 and 28% of all female and male
cancer-related deaths, respectively, in 2013. Non-small cell lung
cancer (NSCLC) accounts for >80% of all lung cancer cases, and
tops the list of the most commonly diagnosed type of cancer
(1). Although great progress has
been achieved in NSCLC therapy, the 5-year overall survival rate
has remained unchanged at <16% (2). Due to the poor prognosis, the
development of novel strategies for the diagnosis and therapy of
NSCLC is urgently required.
MicroRNAs (miRNAs/miRs) are single-stranded, small
non-coding RNAs of 18-22 nucleotides in length, which can modulate
gene expression. miRNAs bind to target mRNAs to regulate their
expression via blocking the transcription or promoting
transcriptional degradation (3). In
addition, they participate in several biological processes,
including cell proliferation, migration and apoptosis (4), and are involved in numerous types of
cancer, such as gastric (5), breast
(6) and lung cancer (7). miR-564 is a novel miRNA that has not
been widely studied. Previous studies have reported that miR-564 is
downregulated in prostate cancer, hepatocellular carcinoma,
osteosarcoma, gastric carcinoma and glioblastoma (8-12).
Furthermore, miR-564 acts as a tumor suppressor in the
aforementioned types of cancers via targeting various mRNAs. MiRs
may act as oncogenes or anti-cancer genes in different type of
cancers. So far, the function of miR-564 in the development of
NSCLC remains to be clarified. Moreover, the mechanism of miR-564
in regulating the cancer progression is limited. The present study
tried to elucidate the presice mechanism of miR-564 in the
development of NSCLC.
Plexin A4, a member of Plexin families, is the
receptor of Semaphorins (Semas), which function as axonal guidance
in nervous system (13). Plexin A4
modulates cell adhesion and migration via interacting with Sema6B,
which is a promoter of tumorigenesis, immune responses, and tissue
remodeling (14-16).
Moroever, overexpression of Plexin A4 contributes to the
inactivation of T cell (17).
Depletion of Plexin A4 in T cells facilitates proliferative ability
in vivo and in vitro. A previous study reveal that
Plexin-A4 enhances tumor progression and angiogenesis (18). However, the potential roles of
Plexin A4 in NSCLC has not been elucidated.
The current study demonstrated that miR-564 was
downregulated in NSCLC cell lines and tissues. The effects of
miR-564 on NSCLC cell proliferation and progression were also
investigated. Herein, the human NSCLC cell lines A594 and H460 were
utilized, whereas the effect of miR-564 on tumor growth in
vivo was investigated in a nude mice xenograft model. Finally,
bioinformatics analysis and luciferase reporter assays revealed
that plexin A4 was directly targeted by miR-564 in NSCLC cells.
Materials and methods
Clinical tissues collection
Human NSCLC tumor and adjacent normal tissues were
collected from 25 patients with NSCLC at the Huaian First People's
Hospital. The tissues were isolated and immediately stored in
liquid nitrogen. Patients signed informed consent in order for
their specimens to be used for scientific research. The current
study was approved by the Ethics Committee of the Huaian First
People's Hospital.
Cell culture
The human bronchial epithelial (HBE) cell line and
four NSCLC cell lines, namely A549, H1299, PC-9 and H460, were
purchased from the American Type Culture Collection. Cells were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with streptomycin (100 µg/ml), penicillin (100 U/ml)
and 10% FBS (Gibco; Thermo Fisher Scientific, Inc.), and incubated
at 37˚C in a humidified 5% CO2 incubator.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from NSCLC specimens and
cell lines using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions. A
total of 1 µg RNA was reverse transcribed into cDNA using the
PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd.).
Furthermore, qPCR was carried out using the SYBR Premix Ex Taq kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions, on a FAST7500 Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The PCR was
performed under the following thermocyling conditions: 95˚C for 30
sec, 40 cycles of 95˚C for 5 sec and 60˚C for 34 sec. U6 and GAPDH
served as loading control for miRNA and mRNA, respectively. The
results were calculated by 2-ΔΔCq method (19).
Cell transfection
miR-564 mimics, inhibitor and the corresponding
negative controls were obtained from Shanghai GenePharma Co., Ltd.
The construction of the plexin A4 overexpression plasmid
(pcDNA3.1/plexin A4) was also completed by Shanghai GenePharma Co.,
Ltd. The miR-564 antagomiR and negative control were purchased from
Guangzhou RiboBio Co., Ltd. Cells were transfected using
Lipofectamine 3000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol.
Cell viability assay
Subsequently, 1x103 A549 or H460 cells
were seeded into 96-well plates 24 h post-transfection. Following
12, 24, 48 and 72 h of incubation, the medium was replaced with
fresh medium supplemented with 10% Cell Counting Kit-8 (CCK-8)
reagent. Cells were then incubated for an additional 4 h. Finally,
the optical density value of each well was determined at 450 nm
using a microplate reader (Bio-Rad Laboratories, Inc.). Each
experiment was carried out ≥3 times.
Colony formation assay
Transfected NSCLC cells were seeded into 12-well
plates (500 cells/well) and incubated for 14 days. Subsequently,
cells were fixed with 4% paraformaldehyde for 15 min, stained with
0.1% crystal violet, and then photographed under a microscope
(Leica Microsystems GmbH). Only colonies with >50 cells were
counted.
Transwell assay
A total of 1x105 cells were harvested,
resuspended in 100 µl serum-free medium and plated into the upper
chambers of 24-well Transwell plates (8.0-µm pore size; Corning,
Inc.). The lower chamber was supplemented with 600 µl medium
containing 10% FBS. Following incubation for 24 h, the medium was
removed, and the chamber was washed twice with PBS. Cells on the
upper chamber were removed using cotton swabs, while those on the
lower surface of the chamber were fixed with 4% paraformaldehyde
for 15 min, stained with 0.1% crystal violet for 5 min, and then
observed under a light microscope (Leica Microsystems GmbH).
Luciferase assay
TargetScan (http://www.targetscan.org/) and MIRDB (http://mirdb.org/) databases were used to predict
whether miR-564 could directly target plexin A4. The
3'-untranslated region (3'UTR) of plexin A4 was synthesized by
Shanghai GenePharma Co., Ltd. and subcloned into a pmirGLO vector
(Promega Corporation). Subsequently, A549 and H460 cells were
co-transfected with miR-564 mimics or mimics control and plasmids
encompassing the wild-type (wt) or mutant (mut) plexin A4-3'UTR
using Lipofectamine 3000 (Thermo Fisher Scientific, Inc.).
Western blot analysis
Cells were lysed with RIPA lysis buffer (Beyotime
Institute of Biotechnology), and the protein concentration was
determined with the NanoDrop 2000 system (Thermo Fisher Scientific,
Inc.). A total of 40 µg protein was separated by 10% SDS-PAGE and
then electrotransferred onto polyvinylidene difluoride (PVDF)
membranes. Following blocking with 5% milk for 60 min at room
temperature, the membranes were incubated with specific primary
antibodies against plexin A4 and GAPDH at 4˚C overnight.
Subsequently, membranes were incubated with the corresponding
secondary antibodies at room temperature for ~2 h. Finally, the
blots were visualized using an ECL kit (Beyotime Institute of
Biotechnology).
Nude mice xenograft model
Male BALB/c nude mice (6-weeks-old) were obtained
from the Vital River Laboratory Animal Technology Company (Beijing,
China). The mice were housed in specific pathogen free (SPF)
facilities under a 12 h light/dark cycle, and temperature- and
humidity-controlled conditions. Nude mice were subcutaneously
injected with 5x106 A549 cells. Following 7 days, the
tumors were injected with 3 nmol of antagomiR-564 or agomir-564
(RiboBio Co., Ltd.) as well as their negative controls
respectively. The injections were carried out every 2 days for a
period of 20 days. The tumor size (width and length) was measured
every 2 days, and tumor volume was calculated using the following
formula: Tumor volume=1/2x (width)2 x length. The animal
experiments were approved by the Ethics Committee of the Huaian
First People's Hospital. At the end, the mouse were euthanized by
intraperitoneal injection of 4% pentobarbital (180 mg/kg). Then,
the tumors were removed.
Immunohistochemistry (IHC)
Paraffin-embedded tumor tissues were cut into 4-µm
thick sections. Then, the sections were deparaffinized in xylene
and rehydrated. Antigen retrieval was performed in a pressure
cooker for 30 min with 10 mM citrate buffer and the activity of
endogenous peroxidase was blocked using 0.5% hydrogen peroxide.
Subsequently, the sections were incubated with primary antibodies
against Ki67 or plexin A4 overnight at 4˚C. Immunostaining was
carried out using a diaminobenzidine (DAB) staining kit according
to the manufacturer's instructions. The stained slides were
visualized under a microscope.
Statistical analysis
All statistical analyses were performed with SPSS
20.0 software (IBM Corp.). Data are expressed as the mean ±
standard deviation (SD). Paired and unpaired Student's t-tests were
used to evaluate the differences in gene expression between two
groups. One-way analysis of variance (ANOVA) with Tukey's test were
used to analyze differences between two and multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-564 expression is downregulated in
NSCLC tissues and cell lines
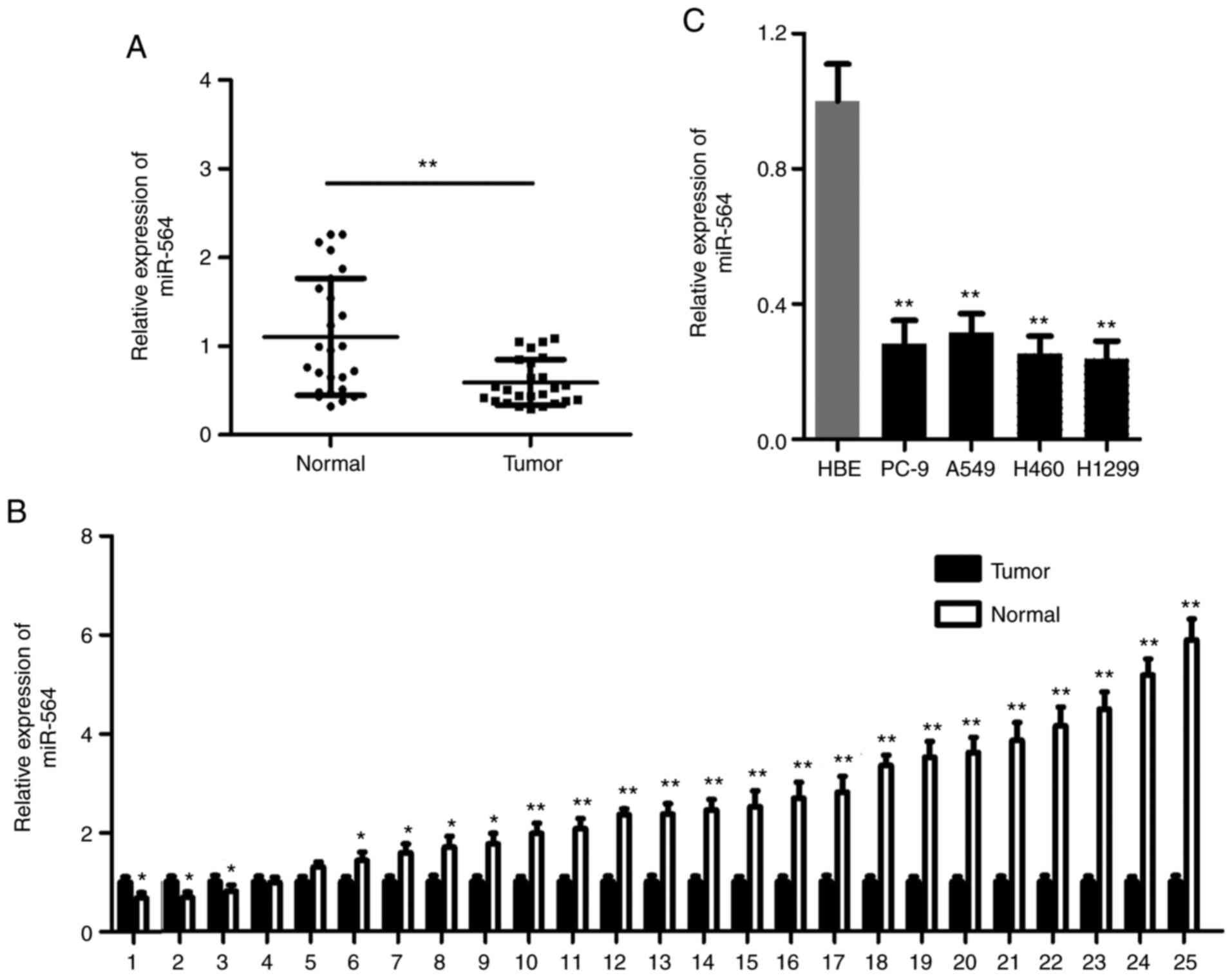
RT-qPCR analysis was used to detect the expression
levels of miR-564 in 25 paired NSCLC tissues and cell lines. The
expression levels of miR-564 were notably reduced in NSCLC tissues
compared with those in the matched normal tissues (Fig. 1A and B). Consistent with the previous finding,
miR-564 was downregulated in all four NSCLC cell lines (A549, H460,
PC-9, and H1200), to differing degrees, compared with the HBE group
(Fig. 1C). These results indicated
that miR-564 could promote NSCLC development.
miR-564 inhibits A549 and H460 cell
proliferation and migration
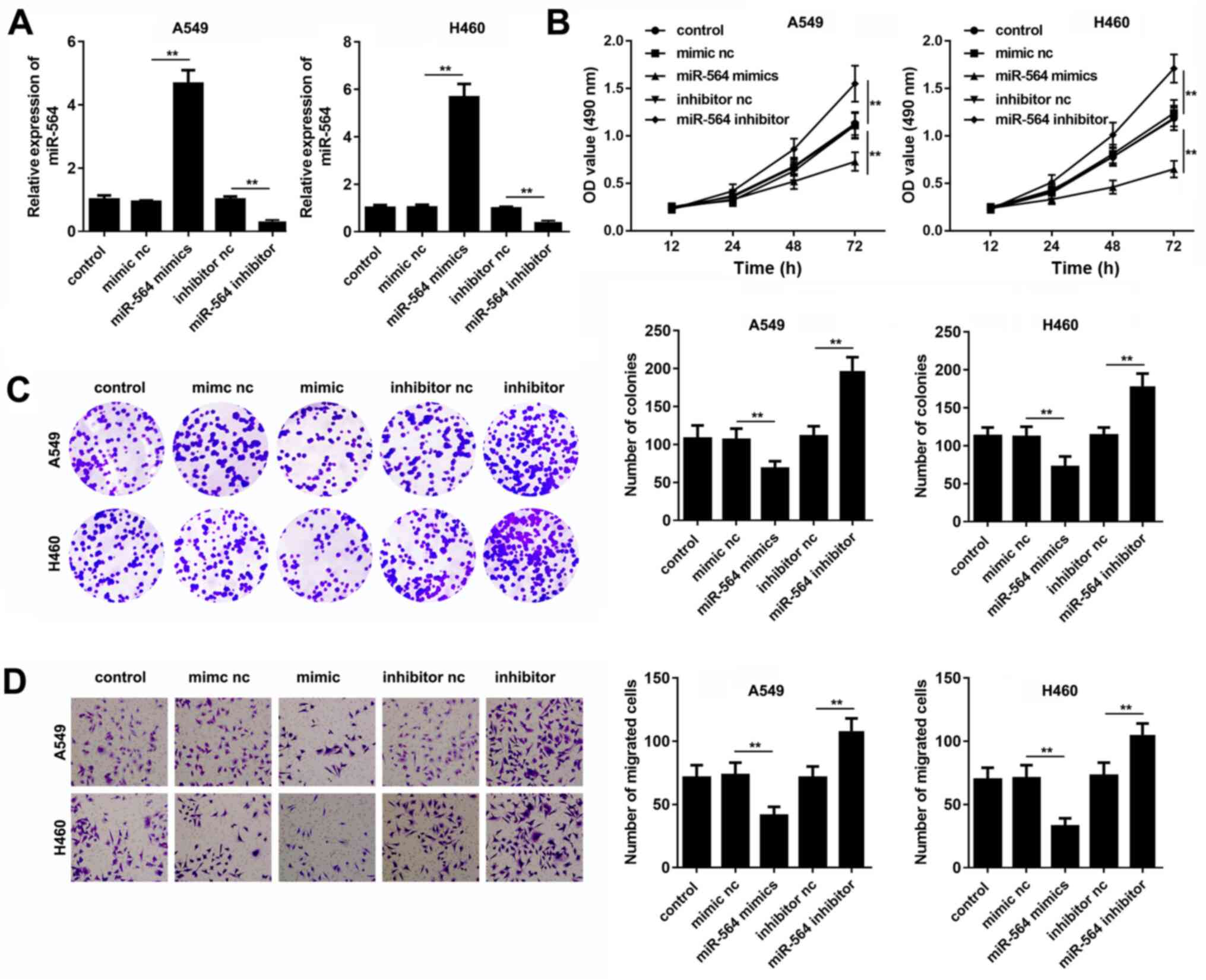
A549 and H460 cells were transfected with miR-564
mimics, inhibitor or their corresponding negative controls and
RT-qPCR analysis was carried out. The results showed that the cell
transfection with miR-564 mimics or inhibitor notably increased and
decreased the miR-564 expression levels, respectively (Fig. 2A). Furthermore, the CCK-8 and colony
formation assays demonstrated that miR-564 mimics inhibited, while
the miR-564 inhibitor promoted, NSCLC cell viability (Fig. 2B and C). Similarly, the Transwell assays
revealed that the cell migration ability was attenuated in A549 and
H460 cells following the transfection with miR-564 mimics. By
contrast, the cell transfection with miR-564 inhibitor induced the
A549 and H460 cell migratory ability (Fig. 2D). Overall, these findings suggested
that miR-564 could attenuate NSCLC cell proliferation and
migration.
miR-564 directly targets plexin A4 in
NSCLC cells
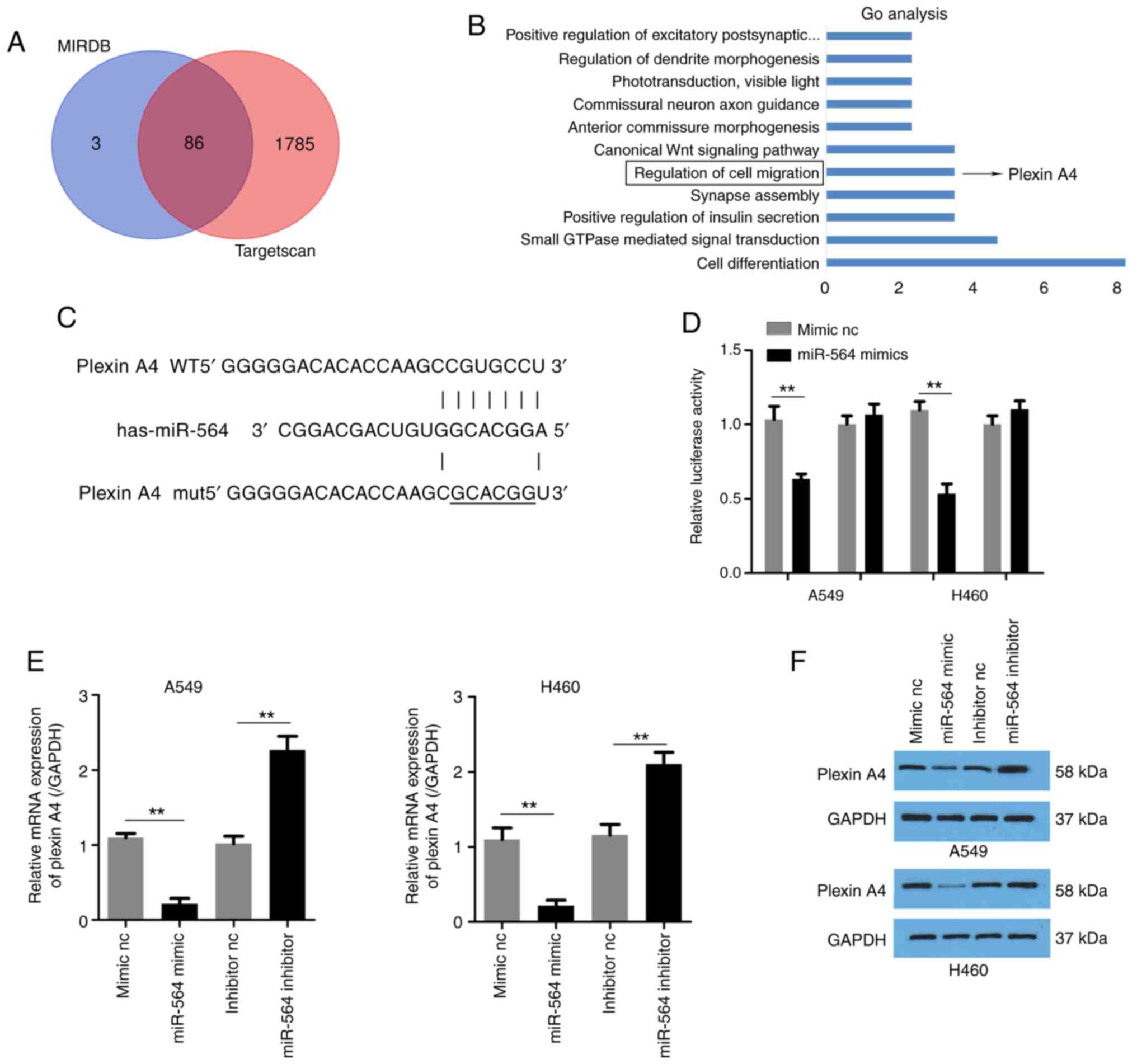
To determine the mechanisms underlying the effects
of miR-564 in NSCLC, the mRNA targets of miR-564 were identified
using TargetScan and MiRDB databases. A total of 86 genes were
predicted as potential targets of miR-564 using both softwares
(Fig. 3A). The results of the Gene
Ontology (GO) enrichment analysis are shown in Fig. 3B. Among the 86 potential target
genes, plexin A4 was selected for subsequent experiments, since it
was predicted to be involved in the regulation of cell migration.
The predicted binding site between miR-564 and plexin A4 is
presented in Fig. 3C. Subsequently,
a luciferase reporter assay was performed. The results demonstrated
that miR-564 overexpression notably reduced the activity of the
reporter plasmid carrying plexin A4-3'-UTR-wt but not that of
plexin A4-3'UTR-mut (Fig. 3D). In
addition, miR-564 expression negatively regulated the mRNA and
protein expression levels of plexin A4 in A549 and H460 cells
(Fig. 3E and F). The aforementioned findings suggested
that plexin A4 was a direct target of miR-564 in A549 and H460
NSCLC cells.
Plexin A4 overexpression reverses the
effect of miR-564 on NSCLC cells
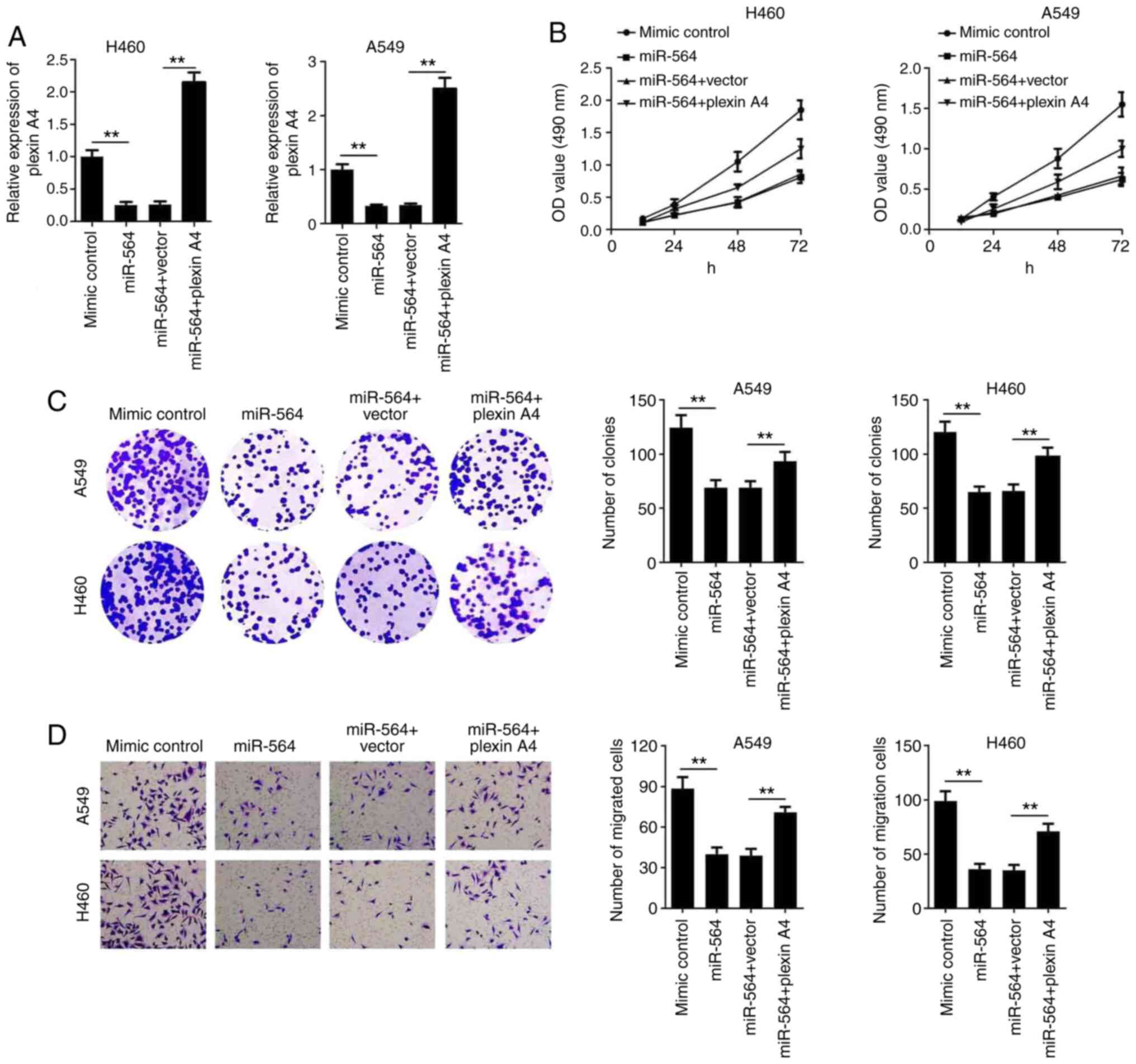
To verify whether plexin A4 could affect the
function of miR-564 on NSCLC cell proliferation and migration, A549
and H460 cells were co-transfected with a plexin A4 overexpression
plasmid and miR-564 mimics. Then, the expression of plexin A4 was
determined by RT-qPCR. The results showed that transfection with a
plexin A4 overexpression plasmid significantly elevated the
expression level of plexin A4 (Fig.
S1). In addition, miR-564 overexpression notably reduced the
expression levels of plexin A4, however, its expression levels were
restored following transfection with a plexin A4 overexpression
plasmid (Fig. 4A). Furthermore, the
CCK-8 and colony formation assays revealed that cell
co-transfection with miR-564 mimics and pcDNA3.1-plexin A4 reversed
the effect of miR-564 on NSCLC cell proliferation (Fig. 4B and C). Additionally, the overexpression of
plexin A4 also reversed the effect of miR-564 on NSCLC cell
migration (Fig. 4D). These findings
indicated that miR-564 could inhibit the proliferative and
migratory ability of NSCLC cells via targeting plexin A4.
miR-564 attenuates tumor growth in
mice
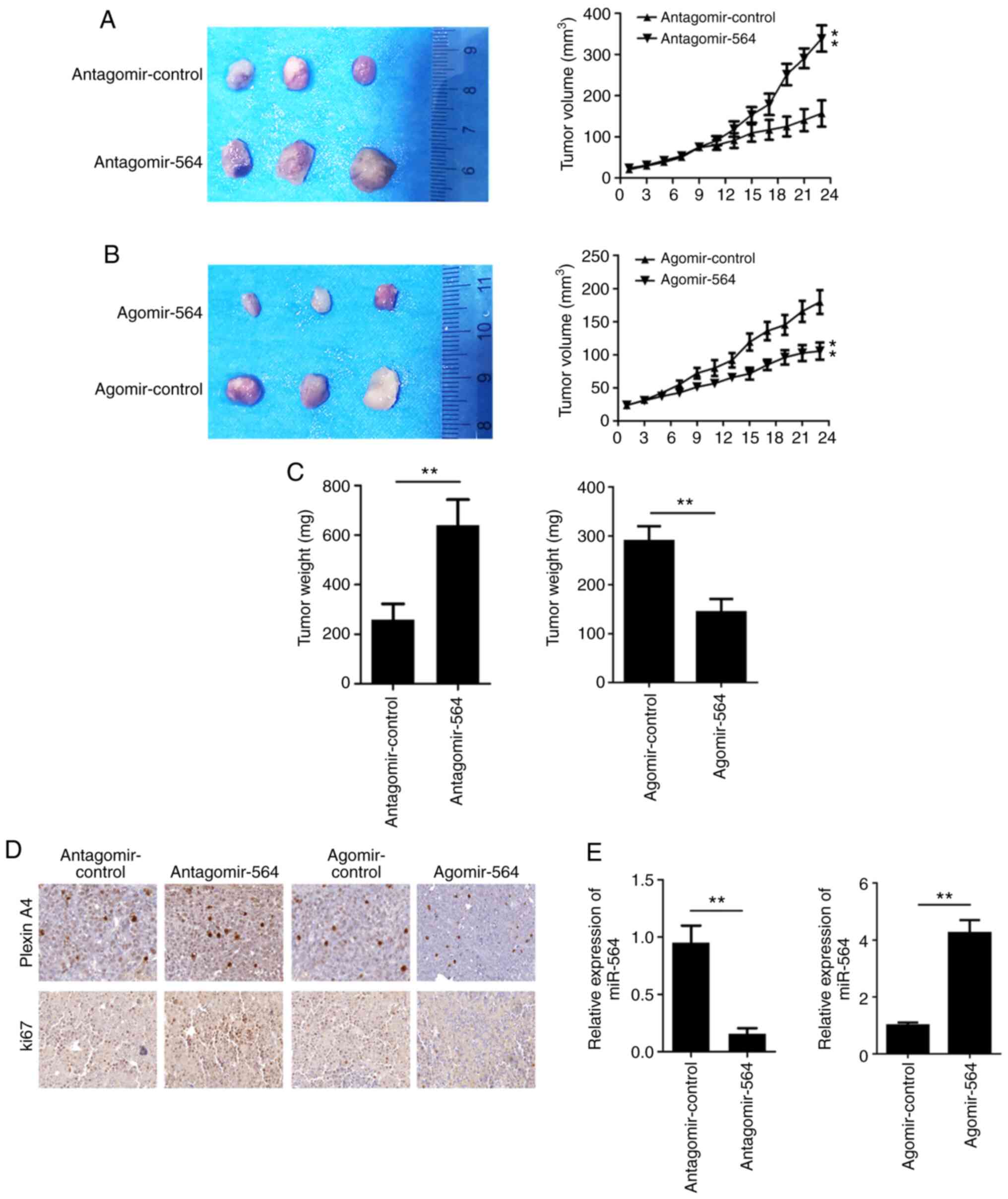
The antagomir-564 and agomir-564 were used to
inhibit or overexpress the expression of miR-564 in the tumor
tissues, respectively. MiR-564 overexpression notably inhibited
while miR-564 knockdown promoted the growth of NSCLC cells in
vivo (Fig. 5A and B). Furthermore, the tumor weight in the
miR-564 silenced group was notably increased compared with the
control group while miR-564 overexpression reduced the tumor weight
(Fig. 5C). The largest tumor
diameter is 12.8 mm. In addition, the IHC results revealed that the
expression of Ki67 and plexin A4 was upregulated after miR-564
silencing while downregulated by miR-564 overexpression (Fig. 5D). To further verify whether the
antogomir-564 could inhibit miR-564, the expression of miR-564 was
determined in tumor tissues. The results indicated that
antogomir-564 could notably attenuate the expression levels of
miR-564 while agomir-564 promoted that (Fig. 5E).
Discussion
In the current study, the expression of miR-564 was
significantly decreased in both NSCLC tissues and cell lines. To
elucidate the function of miR-564, A549 and H460 cells were
transfected with miR-564 mimics, to increase its expression, or
with miR-564 inhibitor to downregulate its endogenous expression.
The functional studies revealed the inhibitory effect of miR-564 on
NSCLC cell proliferation and migration. Mechanistically, miR-564
directly targeted plexin A4 to inhibit its expression.
It has been reported that miR-564 exerts an
antitumor effect in several types of cancer, including NSCLC. More
specifically, miR-564 is downregulated in NSCLC (20). Herein, the A549 and H460 NSCLC cell
lines were used for the in vitro experiments, which was
inconsistent with a previous study, where A549 and H1299 cell lines
were utilized. In addition, the study assessed the effect of
ectopic miR-564 expression on tumor growth in vivo. In the
present study, the role of miR-564 knockdown on NSCLC growth was
investigated in vivo via injecting the miR-564 antagomiR
into mice. Antagomirs and agomirs have been widely used in
vitro and in vivo to knockdown target genes (21,22).
The results demonstrated that the miR-564 antagomiR significantly
downregulated miR-564 in tumor tissues. Furthermore, miR-564
knockdown notably enhanced NSCLC cell growth. Therefore, the
current findings further confirmed the antitumor effect of miR-564
in NSCLC. However, in the present study, the tumor size reached
about 30 mm3 when the treatment was performed. In the future study,
we will perform the administration of antagomir-564 or agomir-564
when the tumors reach a higher volume to check if miR-564 can still
change the growth of NSCLC cells.
miRNAs can modulate the expression of their target
genes via binding to specific regions (23,24).
It has been reported that miR-564 directly targets
astrocyte-elevated gene-1 (AEG1), transforming growth factor
(TGF)-β1 and E2F transcription factor 3 (E2F3) (12,25,26).
Herein, the possible targets of miR-564 were accurately predicted
using TargetScan and MirDB software. Among the possible target
genes, plexin A4 was selected for further experiments, since it was
predicted to be involved in cell migration.
Except from class 3 semaphorins (Semas) such as
Sema3a, which binds to both neuropilin (Nrp)-1 and Nrp-2, plexins
are receptors of Semas. Sema3a signaling is mediated through plexin
A4, as well as other plexin A proteins (27,28).
In addition, the Sema3a/Nrp-1/plexinA4 signaling pathway is
involved in the inflammatory immune response and tumor cell growth
(29). In the present study, the
luciferase reporter assay and rescue experiments predicted and
verified that miR-564 could directly target plexin A4 in NSCLC
cells.
It has been reported that plexin A4 modulates the
phosphorylation status of phosphatase and tensin homolog (PTEN)
(29). Therefore, the effect of
miR-564 on PTEN phosphorylation will be further investigated in
future studies.
In conclusion, the current study showed that miR-564
was downregulated in NSCLC. Furthermore, miR-564 overexpression
attenuated NSCLC proliferation and migration via upregulating
plexin A4 in vitro and in vivo. Therefore, miR-564
and plexin A4 could be considered as potential therapeutic targets
for NSCLC.
Supplementary Material
Plexin A4 overexpression plasmid
significantly promotes the expression of plexin A4. Reverse
transcription-quantitative PCR was used to evaluate the expression
of plexin A4 after transfection. **P<0.01.
Acknowledgements
Not applicable.
Funding
Funding: Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HD performed the in vitro study. BG performed
the in vivo study and the histological examination. LL
analyzed and interpreted the patient data. YN contributed to the
statistical analysis. SC designed the study and wrote the
manuscript and agreed to be accountable for all aspects of the work
in ensuring that questions related to the accuracy or integrity of
any part of the work are appropriately investigated and resolved.
HD, BG and SC confirmed the authenticity of the raw data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The current study was approved by the Ethics
Committee of the Huaian First People's Hospital and was performed
in accordance with the Declaration of Helsinki. Patients signed
informed consent in order for their specimens to be used for
scientific research.
Patient consent for publication
Not Applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang X, Xiong Y, Xia Q, Wu F, Liu L, Zhou
Y, Zeng L, Zhou C, Xia C, Jiang W, et al: Efficacy and safety of
apatinib plus vinorelbine in patients with wild-type advanced
non-small cell lung cancer after second-line treatment failure: A
nonrandomized clinical trial. JAMA Netw Open.
3(e201226)2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Innos K, Oselin K, Laisaar T and Aareleid
T: Patterns of survival and surgical treatment in lung cancer
patients in Estonia by histologic type and stage, 1996-2016. Acta
Oncol. 58:1549–1556. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zare A, Ganji M, Omrani MD, Alipoor B and
Ghaedi H: Gastric cancer MicroRNAs Meta-signature. Int J Mol Cell
Med. 8:94–102. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Jafari N and Abediankenari S: MicroRNA-34
dysregulation in gastric cancer and gastric cancer stem cell.
Tumour Biol. 39(1010428317701652)2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kanchan RK, Siddiqui JA, Mahapatra S,
Batra SK and Nasser MW: microRNAs orchestrate pathophysiology of
breast cancer brain metastasis: Advances in therapy. Mol Cancer.
19(29)2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Gon Y, Shimizu T, Mizumura K, Maruoka S
and Hikichi M: Molecular techniques for respiratory diseases:
MicroRNA and extracellular vesicles. Respirology. 25:149–160.
2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Liang C, Xu Y, Ge H, Xing B, Li G, Li G
and Wu J: miR-564 inhibits hepatocellular carcinoma cell
proliferation and invasion by targeting the GRB2-ERK1/2-AKT axis.
Oncotarget. 8:107543–107557. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ru N, Zhang F, Liang J, Du Y, Wu W, Wang F
and Liu X: MiR-564 is down-regulated in osteosarcoma and inhibits
the proliferation of osteosarcoma cells via targeting Akt. Gene.
645:163–169. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Meng FJ, Meng FM, Wu HX and Cao XF:
miR-564 inhibited metastasis and proliferation of prostate cancer
by targeting MLLT3. Eur Rev Med Pharmacol Sci. 21:4828–4834.
2017.PubMed/NCBI
|
|
11
|
Guo Y, Qi Y, Guo A, Du C, Zhang R and Chu
X: miR-564 is downregulated in gastric carcinoma and targets E2F3.
Oncol Lett. 13:4155–4160. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Jiang C, Shen F, Du J, Hu Z, Li X, Su J,
Wang X and Huang X: MicroRNA-564 is downregulated in glioblastoma
and inhibited proliferation and invasion of glioblastoma cells by
targeting TGF-β1. Oncotarget. 7:56200–56208. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tran TS, Kolodkin AL and Bharadwaj R:
Semaphorin regulation of cellular morphology. Annu Rev Cell Dev
Biol. 23:263–292. 2007.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Matsuoka RL, Nguyen-Ba-Charvet KT, Parray
A, Badea TC, Chedotal A and Kolodkin AL: Transmembrane semaphorin
signalling controls laminar stratification in the mammalian retina.
Nature. 470:259–263. 2011.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Mauti O, Domaniskaya E, Andermatt I, Sadhu
R and Stoeckli ET: Semaphorin6A acts as a gate keeper between the
central and the peripheral nervous system. Neural Dev.
2(28)2007.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Catalano A, Lazzarini R, Di Nuzzo S,
Orciari S and Procopio A: The plexin-A1 receptor activates vascular
endothelial growth factor-receptor 2 and nuclear factor-kappaB to
mediate survival and anchorage-independent growth of malignant
mesothelioma cells. Cancer Res. 69:1485–1493. 2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yamamoto M, Suzuki K, Okuno T, Ogata T,
Takegahara N, Takamatsu H, Mizui M, Taniguchi M, Chédotal A, Suto
F, et al: Plexin-A4 negatively regulates T lymphocyte responses.
Int Immunol. 20:413–420. 2008.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kigel B, Rabinowicz N, Varshavsky A,
Kessler O and Neufeld G: Plexin-A4 promotes tumor progression and
tumor angiogenesis by enhancement of VEGF and bFGF signaling.
Blood. 118:4285–4296. 2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Yang B, Jia L, Guo Q, Ren H, Hu D, Zhou X,
Ren Q, Hu Y and Xie T: MiR-564 functions as a tumor suppressor in
human lung cancer by targeting ZIC3. Biochem Biophys Res Commun.
467:690–966. 2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang H, Chao K, Ng SC, Bai AH, Yu Q, Yu J,
Li M, Cui Y, Chen M, Hu JF and Zhang S: Pro-inflammatory miR-223
mediates the cross-talk between the IL23 pathway and the intestinal
barrier in inflammatory bowel disease. Genome Biol.
17(58)2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wu Y, Heinrichs J, Bastian D, Fu J, Nguyen
H, Schutt S, Liu Y, Jin J, Liu C, Li QJ, et al: MicroRNA-17-92
controls T-cell responses in graft-versus-host disease and leukemia
relapse in mice. Blood. 126:1314–1323. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Whipple AJ, Breton-Provencher V, Jacobs
HN, Chitta UK, Sur M and Sharp PA: Imprinted maternally expressed
microRNAs antagonize paternally driven gene programs in neurons.
Mol Cell. 78:85–95.e8. 2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Basak A, Munschauer M, Lareau CA,
Montbleau KE, Ulirsch JC, Hartigan CR, Schenone M, Lian J, Wang Y,
Huang Y, et al: Control of human hemoglobin switching by
LIN28B-mediated regulation of BCL11A translation. Nat Genet.
52:138–145. 2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Xiao L, Tang T, Huang Y and Guo J: MiR-564
promotes hypertrophic scar formation through TGF-β1 upregulation. G
Ital Dermatol Venereol. 154:186–191. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Song Z, Yang H, Wu X, Kong C and Xu C:
microRNA-564 inhibits the aggressive phenotypes of papillary
thyroid cancer by directly targeting astrocyte-elevated gene-1.
Onco Targets Ther. 12:4869–4881. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Casazza A, Laoui D, Wenes M, Rizzolio S,
Bassani N, Mambretti M, Deschoemaeker S, Van Ginderachter JA,
Tamagnone L and Mazzone M: Impeding macrophage entry into hypoxic
tumor areas by Sema3A/Nrp1 signaling blockade inhibits angiogenesis
and restores antitumor immunity. Cancer Cell. 24:695–709.
2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
He Z and Tessier-Lavigne M: Neuropilin is
a receptor for the axonal chemorepellent Semaphorin III. Cell.
90:739–751. 1997.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Podojil JR, Chiang MY, Ifergan I, Copeland
R, Liu LN, Maloveste S, Langermann S, Liebenson D, Balabanov R, Chi
H, et al: B7-H4 modulates regulatory CD4(+) T Cell induction and
function via ligation of a semaphorin 3a/Plexin A4/Neuropilin-1
complex. J Immunol. 201:897–907. 2018.PubMed/NCBI View Article : Google Scholar
|