Introduction
Polycystic ovary syndrome (PCOS) is the most common
endocrine and metabolic disorder in females of reproductive age
worldwide, with a prevalence of 6-20%, depending on the diagnostic
criteria applied (1). The key
features of PCOS include hyperandrogenism, polycystic ovarian
morphology, ovulatory dysfunction and irregular menstrual cycles
(2). PCOS is a primary cause of
infertility, resulting in abnormal follicle development and a high
rate of follicular atresia in females with PCOS (3). Increasing evidence indicates that
granulosa cell (GC) apoptosis serves an important role in
follicular atresia and PCOS development (4,5). GC
apoptosis is increased in patients with PCOS and in PCOS model
animals, compared with control patients with normal ovarian
function and healthy animals respectively (6,7), and
inhibition of GC apoptosis improved the ovary functions of PCOS
model rats (8). The hyperandrogenic
condition is a key pathogenic factor that mediates GC apoptosis in
PCOS (9). It was reported that
androgens induce GC apoptosis via activation of the intrinsic and
extrinsic apoptotic signaling pathways (7,9).
However, the mechanisms underlying androgen-mediated induction of
GC apoptosis are not completely understood.
MicroRNAs (miRs/miRNAs) are short non-coding RNA
molecules that are widely present in different tissues (10). miRNAs regulate gene expression by
destabilizing target mRNAs or inhibiting translation (10). miR-874-3p, a newly identified miRNA,
is associated with several pathological processes, including
cancer, myocardial infarction, bone formation and erectile
dysfunction (11-15).
However, the role of miR-874-3p in PCOS and GC apoptosis is yet to
be elucidated. Several targets of miR-874-3p have been identified,
one of which is histone deacetylase (HDAC)1(14). Although the effects of HDAC1 on
apoptosis have been widely studied (16,17),
to the best of our knowledge, the role of HDAC1 in GC apoptosis,
especially in hyperandrogenic conditions, has not been previously
reported.
The present study assessed the miR-874-3p expression
profile in PCOS and testosterone-induced GC apoptosis.
Additionally, the present study investigated the role of miR-874-3p
in testosterone-induced GC apoptosis, as well as the underlying
mechanisms.
Materials and methods
Subjects and collection of human
GCs
The present study was approved by the Medical Ethics
Committee of Zaozhuang Maternal and Child6 Health Care Hospital.
All patients were female and hospitalized in Zaozhuang Maternal and
Child Health Care Hospital (Zaozhuang, China) from February 2016 to
May 2018. Written informed consent was obtained from all patients.
A total of 16 patients diagnosed with PCOS according to the
Rotterdam criteria (18) were
recruited into the PCOS group. The control group consisted of 11
patients with normal ovarian function and regular periods, where
infertility was ascribed to tubal factors or male infertility.
Individuals with ovarian tumors, congenital adrenal hyperplasia,
androgen secreting tumors, Cushing's syndrome and endometriosis
were excluded. All patients received controlled ovarian
hyperstimulation as previously reported (6). Briefly, follicular growth was promoted
with recombinant follicle stimulating hormone (FSH; LiShenBao,
Lizhu Pharmaceutical; https://en.livzon.com.cn/product/14.html) on the third
day of the menstrual cycle. The dosages were adjusted according to
the follicular size and serum hormonal level of each patient. When
at least three of the follicles displayed diameters of 18 mm,
10,000 IU human chorionic gonadotropin (Lizhu Pharmaceutical;
https://en.livzon.com.cn/product/9.html) was
administered to induce ovulation. At 36 h post-administration,
oocyte retrieval was performed via ultrasound-guided puncture.
Follicular fluids (diameter, >18 mm) were carefully collected
and immediately transported to the laboratory on ice.
Human GCs were isolated using a previously described
strainer methodology (19). First,
the follicular fluids were filtered through a 40-µm cell strainer.
The strainer was rinsed with PBS and then backwashed with PBS to
collect GCs. Following incubation for 5 min at 4˚C, the cell
suspension was repeatedly aspirated using Pasteur pipettes to
mechanically break up aggregates. The resulting cell suspension was
filtered through a 70-µm cell strainer to remove unwanted and
undispersed material. Finally, the filtrate was centrifuged at 600
x g for 5 min at 4˚C. GCs in the cell pellet were used for
subsequent experiments. Human GCs were cultured under the same
conditions as mouse GCs, which was in DMEM/F12 (HyClone; Cytiva)
supplemented with 10% FBS (HyClone; Cytiva) and 1%
penicillin/streptomycin (HyClone; Cytiva) at 37˚C with 5%
CO2
Isolation, culture, and treatment of
mouse GCs
All animal procedures were approved by the Animal
Ethics Committee of Zaozhuang Maternal and Child Health Care
Hospital. In total, 84 C57BL/6 mice (weight, 10.3±0.9 g; age, 21
days; total number, 84; immature females) were from Shanghai SLAC
Laboratory Animal Co., Ltd. The mice were housed in a temperature
(22-24˚C) and humidity (50-60%) controlled environment with a 12-h
light/dark cycle and given ad libitum access to food and
water. The mice were intraperitoneally injected with 8 IU pregnant
mare serum gonadotrophin (Ningbo Sansheng Pharmaceutical Industry
Co., Ltd.). At 47 h post-injection, mice were sacrificed by
cervical dislocation, the ovaries were collected and the
surrounding tissues removed. GCs were isolated from the ovaries by
puncturing the follicles with 26-gauge needles, followed by
centrifugation at 300 x g for 10 min at 4˚C. GCs were cultured in
DMEM/F12 (HyClone; Cytiva) supplemented with 10% FBS (HyClone;
Cytiva) and 1% penicillin/streptomycin (HyClone; Cytiva) at 37˚C
with 5% CO2. At 1 day post-transduction, cells were
incubated with 10 µM testosterone (Sigma-Aldrich; Merck KGaA) or
vehicle (0.1% DMSO) for 24 h at 37˚C to induce apoptosis. In
addition, cells were treatment with 10 µM testosterone together
with 1 µM flutamide (Sigma-Aldrich; Merck KGaA) at 37˚C for 24
h.
Lentiviral vectors
miR-874-3p mimic non-targeting negative control (NC;
5'-ACUACUGAGUGACAGUAGA-3') or miR-874-3p mimic
(5'-CUGCCCUGGCCCGAGGGACCGA-3') were inserted into the GV309 vector
(Shanghai GeneChem Co., Ltd.); miR-874-3p inhibitor non-targeting
NC (5'-CAGUACUUUUGUGUAGUACAA-3') or miR-874-3p inhibitor
(5'-UCGGUCCCUCGGGCCAGGGCAG-3') were inserted into the GV280 vector
(Shanghai GeneChem Co., Ltd.). For HDAC1 overexpression, the coding
sequence of HDAC1 was inserted into the GV358 vector (Shanghai
GeneChem Co., Ltd.). The empty vector was used as the NC for HDAC1
overexpression. 293T cells (American Type Culture Collection) were
transfected with 20 µg the GV vector (GV309, GV280 or GV358)
together with 15 µg pHelper 1.0 and 10 µg pHelper 2.0 vectors (both
from Shanghai GeneChem Co., Ltd.) and then cultured at 37˚C in a
humidified incubator in an atmosphere of 5% CO2 for 48 h
to produce lentiviral particles. Transduction was performed when
cells were growing exponentially and were 70-80% confluent.
Polybrene (8 µg/ml, Sigma-Aldrich; Merck KGaA) and appropriate
lentiviral particles (108 TU/ml, 5 µl) were added to
cells for 16 h at 37˚C in a humidified incubator in an atmosphere
of 5% CO2. The medium containing lentiviral particles
were removed from wells and fresh medium were added. Subsequent
experiments were performed 24 h later.
Flow cytometry analysis
GC apoptosis (early and late) was quantified by flow
cytometry using an Annexin V-FITC Apoptosis Detection kit (cat. no.
556547; BD Bioscience) according to the manufacturer's
instructions. Briefly, cells were harvested, washed twice with cold
PBS and re-suspended in the binding buffer. The cell suspension
(100,000 cells/ml) was incubated with 5 µl Annexin V-FITC and 10 µl
PI for 15 min at room temperature in the dark. Subsequently, flow
cytometry was performed using a flow cytometer (CytoFlex S; Beckman
Coulter, Inc.). The data were analyzed with FlowJo software (v10;
FlowJo, LLC.).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated using an RNeasy Plus Mini kit
(Qiagen GmbH) according to the manufacturer's instructions. Total
RNA was reverse transcribed into cDNA using the PrimeScript™ RT
reagent kit (Takara Biotechnology Co., Ltd.). The reverse
transcription reactions were performed at 37˚C for 15 min, followed
by inactivation of the reverse transcriptase at 85˚C for 5 sec.
Subsequently, qPCR was performed using SYBR Premix Ex Taq (Takara
Biotechnology Co., Ltd.). The following thermocycling conditions
were used for qPCR: 95˚C for 10 min; followed by 40 cycles of 95˚C
for 15 sec and 60˚C for 1 min. The following primers were used for
qPCR: miR-874-3p forward, GAACTCCACTGTAGCAGAGATGGT and reverse,
CATTTTTTCCACTCCTCTTCTCTC; HDAC1 (human) forward,
GAATCCGCATGACTCATAAT and reverse, GCTGTGGTACTTGGTCATCT; HDAC1
(mouse) forward, CTGAATACAGCAAGCAGATGCAGAG and reverse,
TCCCGTGGACAACTGACAGAAC; U6 (human and mouse) forward,
CTCGCTTCGGCAGCACA and reverse, AACGCTTCACGAATTTGCGT; GAPDH (human)
forward, GCACCGTCAAGGCTGAGAAC and reverse, TGGTGAAGACGCCAGTGGA; and
GAPDH (mouse) forward, AACGGGAAGCTCACTGGCAT and reverse,
GCTTCACCACCTTCTTGATG. miRNA and mRNA expression levels were
quantified using the 2-ΔΔCq method (20) and normalized to the internal
reference genes U6 and GAPDH, respectively.
Western blotting
Proteins were extacted using a Western Cell lysis
Buffer kit (Beyotime Institute of Biotechnology) and the
concentrations were determined by a bicinchninic acid (BCA) Protein
Assay Kit (Beyotime Institute of Biotechnology). In total, 30 µg
total proteins were loaded per lane. Proteins were separated via
10% SDS-PAGE and transferred to PVDF membranes (EMD Millipore).
Following blocking with 5% non-fat milk at room temperature for 1
h, the membranes were incubated at 4˚C overnight with the following
primary antibodies: Anti-androgen receptor (AR; cat. no. 5153;
1:1,000; Cell Signaling Technology, Inc.); Anti-p53 (cat. no. ab26;
1:1,000; Abcam), anti-acetylated p53 (Lys379; cat. no. 2570;
1:1,000; Cell Signaling Technology, Inc.), anti-HDAC1 (cat. no.
34589; 1:1,000; Cell Signaling Technology, Inc.), anti-sirtuin 1
(SIRT1; cat. no. 8469; 1:1,000; Cell Signaling Technology, Inc.)
and anti-GAPDH (cat. no. ab8245; 1:1,000; Abcam). After washing by
TBS with 0.05% Tween-20, the membranes were incubated with
appropriate horseradish peroxidase-conjugated secondary antibodies
(cat. no. ab7090 and ab97040; 1:5,000; Abcam) at room temperature
for 1 h. Protein bands were visualized using an ECL kit (Beyotime
Institute of Biotechnology).
Measurement of HDAC activity
GC nuclear extracts were obtained and quantified
using a Nuclear Extract kit (Beyotime Institute of Biotechnology)
and a BCA protein assay kit (Beyotime Institute of Biotechnology),
respectively. HDAC activity in the nuclear extracts was determined
using an HDAC Activity Colorimetric Assay kit (cat. no. K331;
BioVision, Inc.) according to the manufacturer's instructions. The
absorbance was measured at a wavelength of 405 nm using a
microplate reader.
Statistical analysis
All experiments were performed in triplicate. Data
are presented as the mean ± SEM. Comparisons between two groups
were analyzed using the Student's unpaired t-test. Comparisons
among multiple groups were analyzed using one-way ANOVA followed by
Bonferroni's post hoc test. For correlation analyses, Pearson's
correlation coefficient was used. Statistical analyses were
performed using SPSS 13.0 software (SPSS, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-874-3p is upregulated in GCs
isolated from patients with PCOS
miR-874-3p expression in the GCs of patients with or
without PCOS were assessed. A total of 16 patients with PCOS and 11
patients with PCOS were recruited in the present study. The
clinical features of the patients are presented in Table I. There was no significant
difference in age between the two groups. However, compared with
patients without PCOS, patients with PCOS displayed significantly
higher basal testosterone levels, basal luteinizing hormone (LH)
levels and LH/FSH ratios. The rate of apoptosis of GCs isolated
from patients with PCOS was significantly higher compared with
patients without PCOS (Fig. S1).
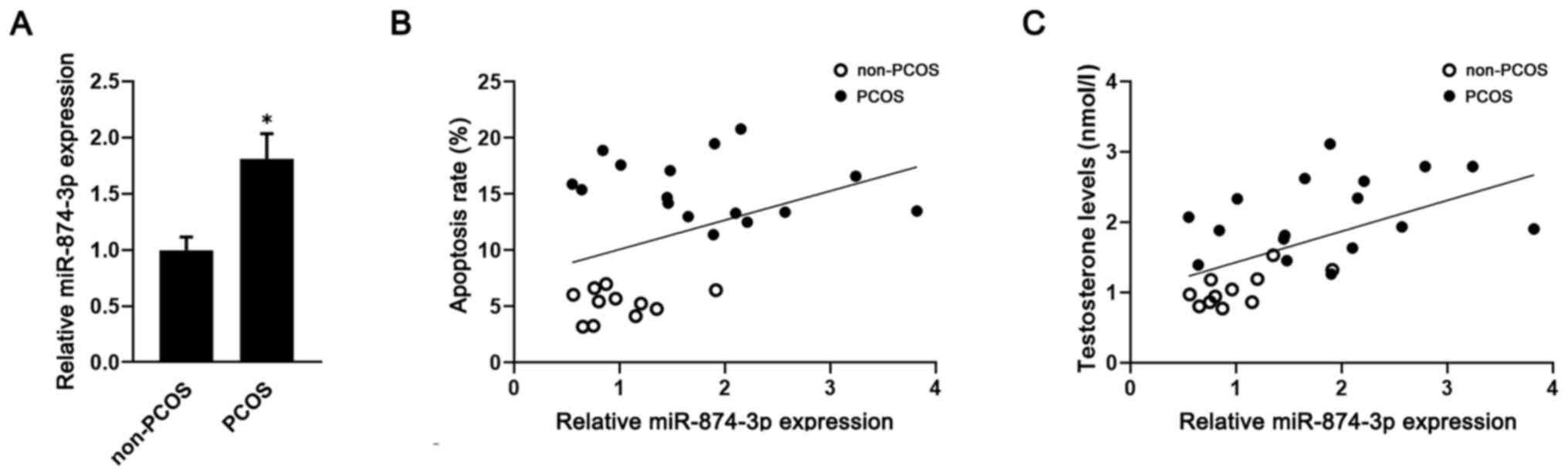
Meanwhile, the RT-qPCR results demonstrated that miR-874-3p
expression was significantly upregulated in GCs isolated from
patients with PCOS compared with patients without PCOS (Fig. 1A). Moreover, miR-874-3p expression
levels were positively correlated with GC apoptosis (r=0.39;
Fig. 1B) and testosterone levels
(r=0.55; Fig. 1C) in both patients
with PCOS and patients without PCOS.
 | Table IClinical features of patients with or
without PCOS. |
Table I
Clinical features of patients with or
without PCOS.
| | Non-PCOS (n=11) | PCOS (n=16) | P-value |
|---|
| Age (year) | 30.18±3.16 | 30.50±2.90 | >0.05 |
| BMI
(kg/m2) | 23.84±1.91 | 23.76±1.97 | >0.05 |
| Basal T
(nmol/l)a | 1.04±0.24 | 2.05±0.53 | 0.03 |
| Basal E2
(pmol/l) | 136.57±47.29 | 153.66±49.18 | >0.05 |
| Basal PRL
(µIU/ml) | 278.33±87.45 | 301.72±90.33 | >0.05 |
| Basal FSH
(mIU/ml) | 6.95±1.02 | 6.77±1.21 | >0.05 |
| Basal LH
(mIU/ml)a | 4.55±1.37 | 12.51±5.93 | 0.01 |
| LH/FSH
ratioa | 0.68±0.28 | 1.57±0.58 | 0.02 |
miR-874-3p serves a role in
testosterone-induced GC apoptosis
Since hyperandrogenic conditions are a key
pathogenic factor that mediate GC apoptosis in PCOS (9), the role of miR-874-3p in
testosterone-induced GC apoptosis was investigated. Compared with
the control group (0 µM testosterone), testosterone significantly
increased GC apoptosis at concentrations ≥2.5 µM (Fig. 2A and B). Moreover, miR-874-3p expression levels
were significantly increased in testosterone-treated GCs compared
with vehicle-treated GCs (Fig. 2C).
The protein expression levels of androgen receptor were also
notably upregulated in testosterone-treated GCs compared with
vehicle-treated GCs (Fig. 2D). To
examine whether the induction of miR-874-3p was mediated by the
androgen receptor, GCs were treated with testosterone in the
presence or absence of the androgen receptor antagonist flutamide.
In GCs, testosterone-induced miR-874-3p upregulation was
significantly inhibited by flutamide (Fig. 2E). Furthermore, GCs were transduced
with lentiviral vectors expressing miR-874-3p mimic or inhibitor.
Compared with the mimic NC group, miR-874-3p mimic significantly
increased miR-874-3p expression levels, whereas compared with the
inhibitor NC group, miR-874-3p inhibitor significantly decreased
miR-874-3p expression levels (Fig.
2F). Moreover, compared with the control group, miR-874-3p
mimic significantly increased GC apoptosis and miR-874-3p inhibitor
significantly decreased GC apoptosis (Fig. 2G and H). The results indicated that miR-874-3p
may serve a role in testosterone-induced GC apoptosis.
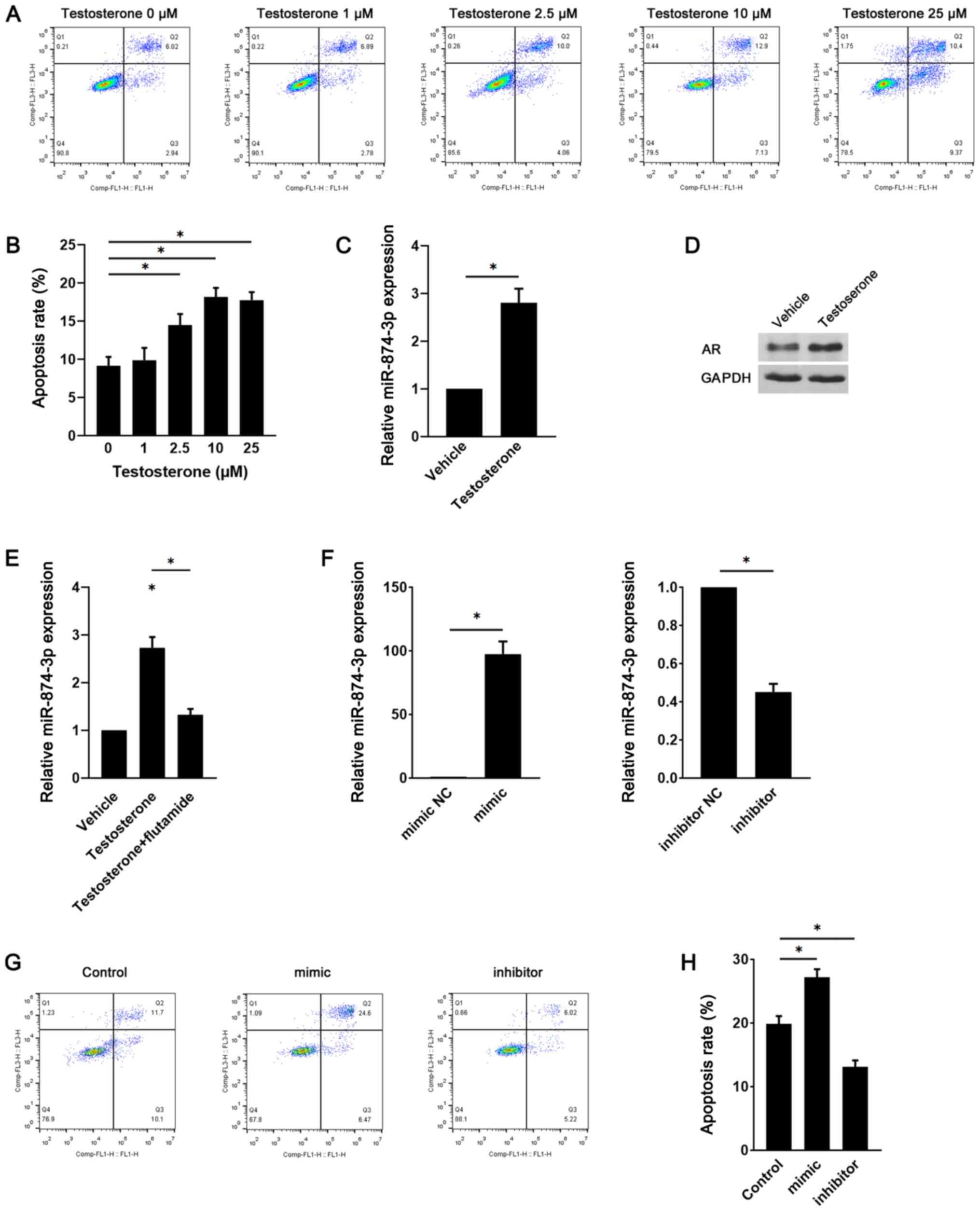 | Figure 2miR-874-3p serves a role in
testosterone-induced GC apoptosis. Following treatment with
different doses of testosterone for 24 h, mouse GC apoptosis was
(A) determined via flow cytometry and (B) quantified. (C) Following
treatment with vehicle or 10 µM testosterone for 24 h, miR-874-3p
expression levels were detected via RT-qPCR. (D) AR protein
expression levels were detected via western blotting. (E) Following
treatment with 10 µM testosterone in the presence or absence of 1
µM flutamide for 24 h, miR-874-3p expression levels in mouse GCs
were detected via RT-qPCR. (F) Transduction efficiency of
miR-874-3p mimic and miR-874-3p inhibitor in mouse GCs. Following
transduction with lentiviral vectors expressing miR-874-3p mimic or
miR-874-3p inhibitor for 1 day, then treatment with 10 µM
testosterone for 24 h, mouse GC apoptosis was (G) determined via
flow cytometry and (H) quantified. *P<0.05. miR,
microRNA; GC, granulosa cell; RT-qPCR, reverse
transcription-quantitative PCR; AR, androgen receptor; NC, negative
control. |
HDAC1 is a target of miR-874-3p during
GC apoptosis
HDAC1 was reported to be a target of miR-874-3p
(12). Therefore, HDAC1 expression
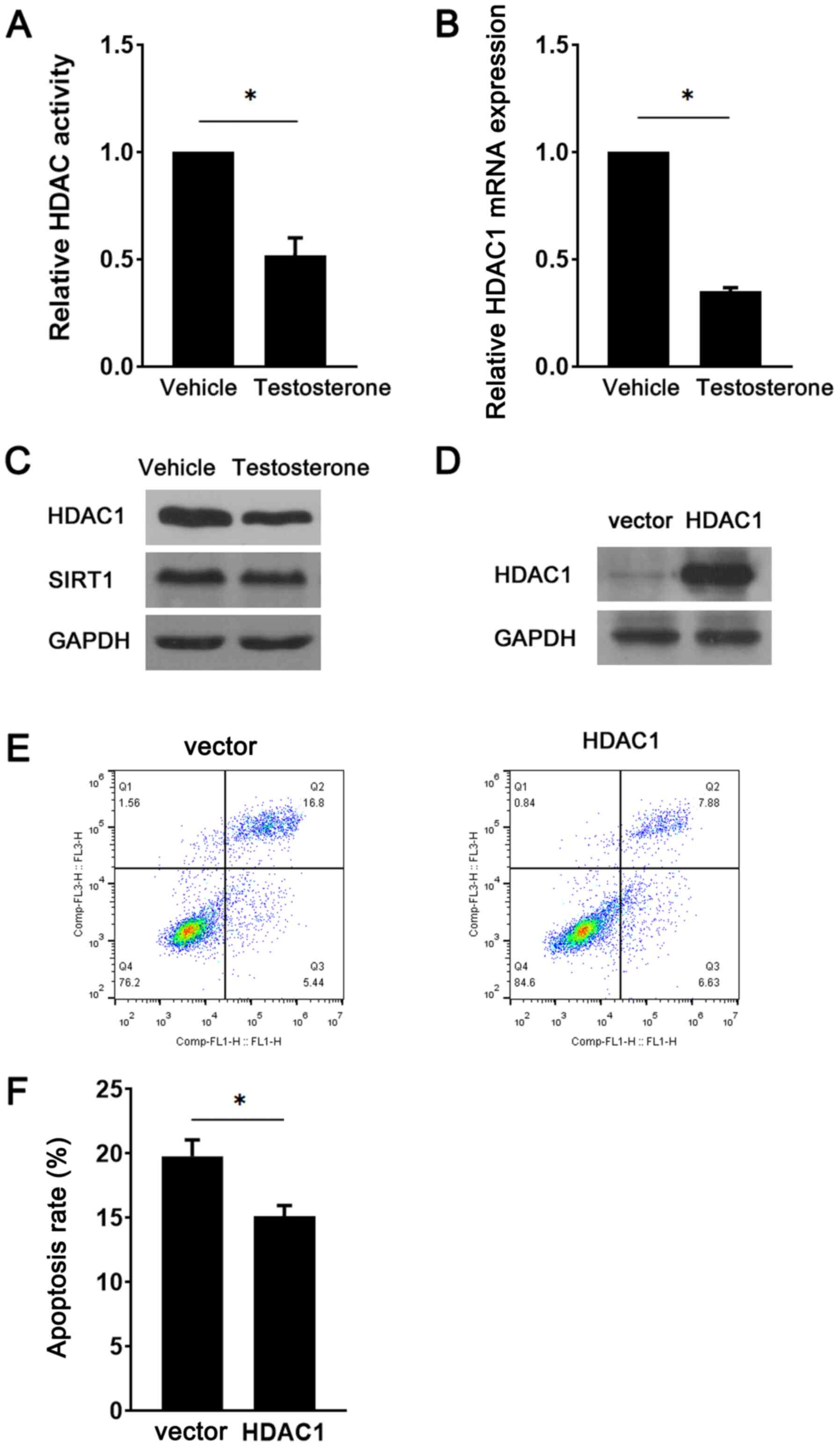
during GC apoptosis was examined. Compared with the vehicle group,
testosterone treatment significantly decreased HDAC activity
(Fig. 3A). Similarly, HDAC1
expression was markedly downregulated in testosterone-treated GCs
compared with vehicle-treated GCs (Fig.
3B and C). However, there were
no notable alterations in the expression levels of SIRT1, a HDAC
family member, between the vehicle and testosterone groups
(Fig. 3C). To determine the role of
HDAC1 downregulation in testosterone-induced GC apoptosis, a
lentiviral vector was used to overexpress HDAC1 in the presence of
testosterone. The western blotting results indicated that HDAC1
protein expression levels were notably increased in the HDAC1
overexpression group compared with the vector group, demonstrating
successful transduction (Fig. 3D).
GC apoptosis was significantly decreased in the HDAC1
overexpression group compared with the vector group (Fig. 3E and F). Subsequently, the present study
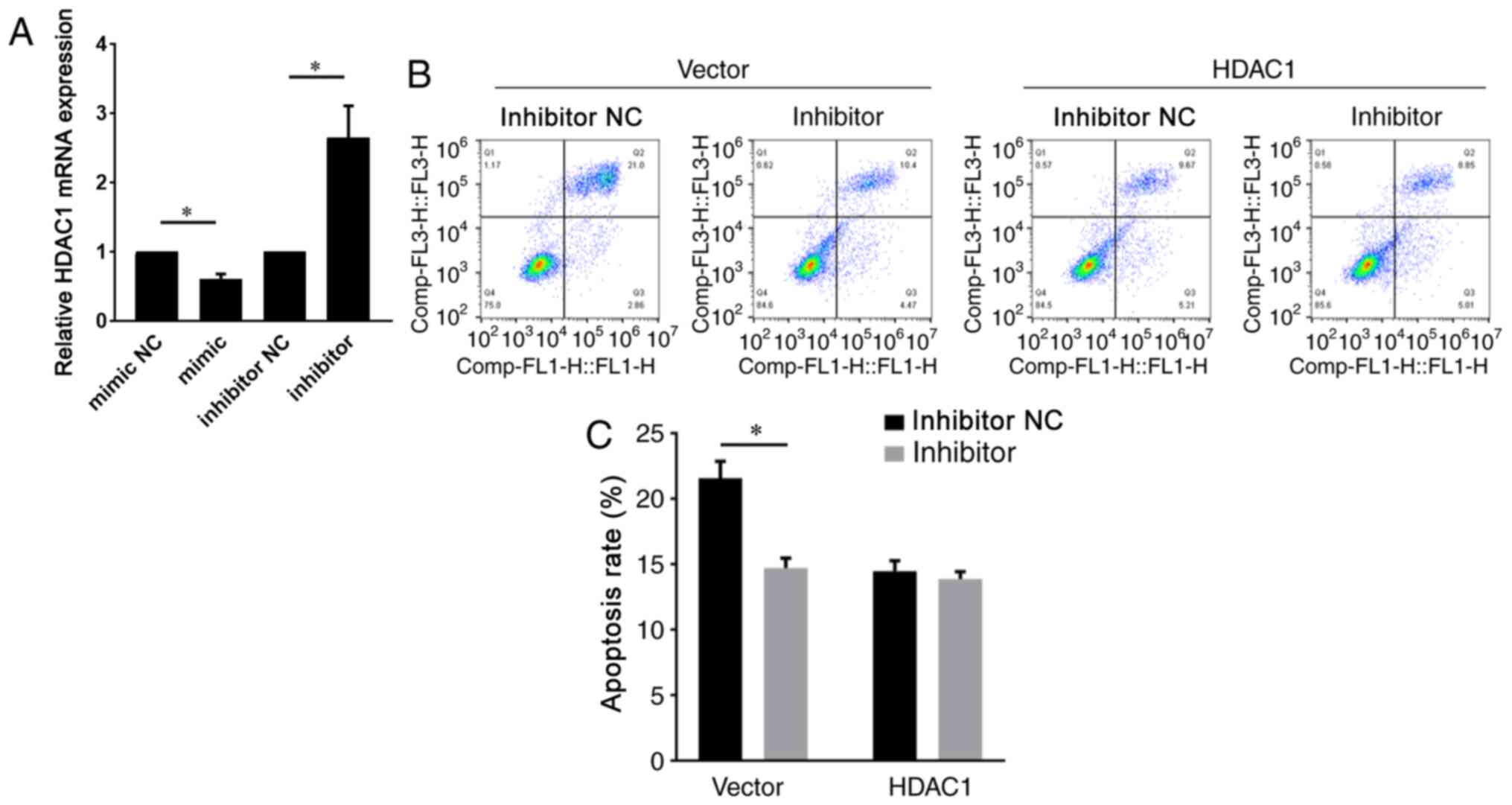
investigated whether miR-874-3p regulated HDAC1 in GCs. Compared
with the control group, miR-874-3p mimic significantly decreased
HDAC1 expression, whereas miR-874-3p inhibitor significantly
increased HDAC1 expression in GCs (Fig.
4A). By contrast, miR-874-3p inhibitor could not attenuate the
apoptosis of GCs with HDAC1 overexpression further (Fig. 4B and C), suggesting that miR-874-3p mediated GC
apoptosis via HDAC1. Collectively, the aforementioned results
indicated that miR-874-3p downregulated HDAC1 to promote GC
apoptosis.
miR-874-3p promotes p53 acetylation
and accumulation by suppressing HDAC1
To investigate how miR-874-3p mediated GC apoptosis,
the expression of several apoptosis-related proteins was examined.
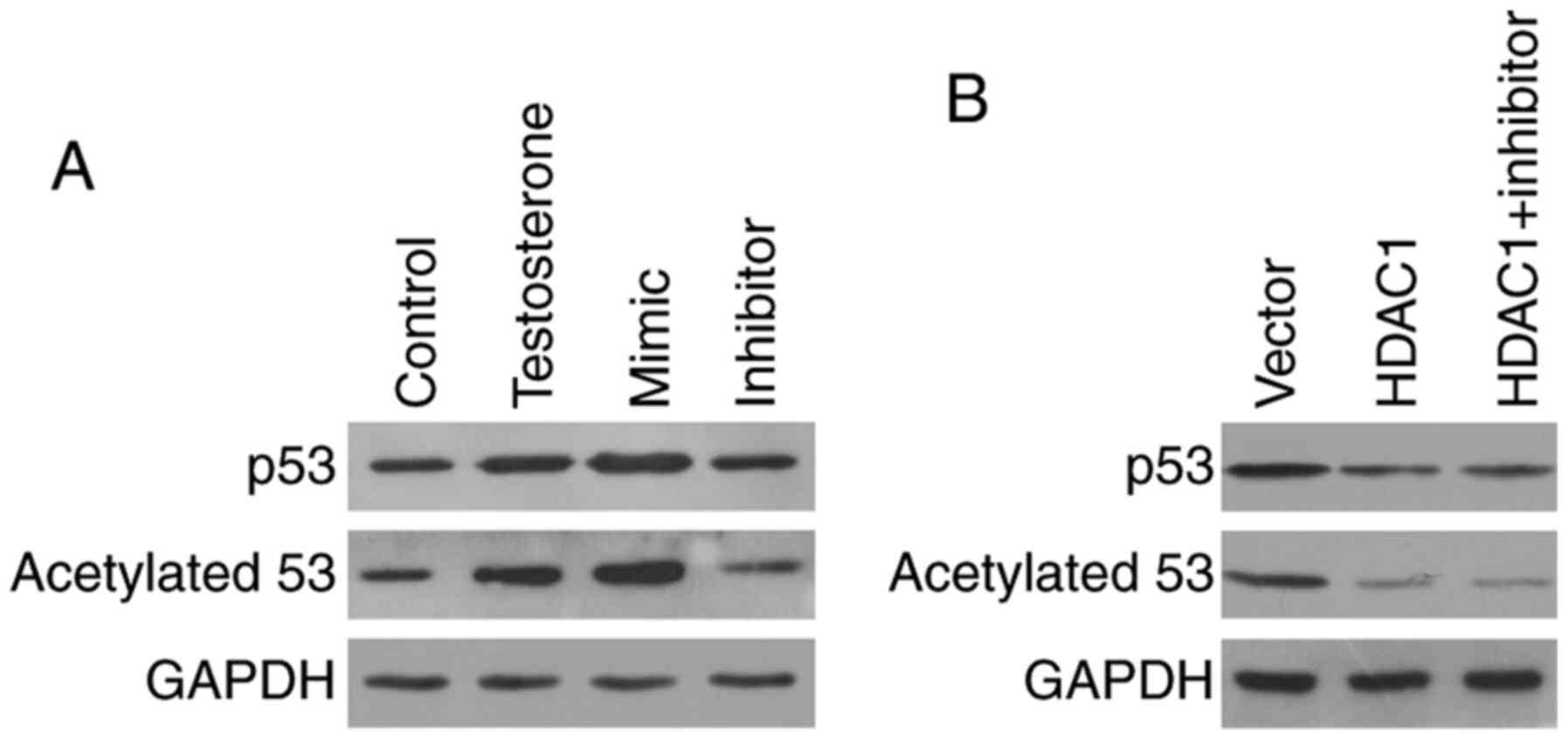
Compared with the control group, the protein expression levels of
the proapoptotic protein p53 were notably upregulated in
testosterone-treated GCs. Moreover, compared with the control
group, p53 protein expression levels were markedly increased
miR-874-3p mimic, but notably decreased by miR-874-3p inhibitor
(Fig. 5A). Since acetylation serves
a positive role in p53 protein accumulation (21), the expression levels of acetylated
p53 were also detected. The western blotting results demonstrated
that p53 acetylation was markedly increased by testosterone
treatment compared with the control group. Moreover, miR-874-3p
mimic increased p53 acetylation, whereas miR-874-3p inhibitor
decreased p53 acetylation, compared with the testosterone group
(Fig. 5A). To determine whether
miR-874-3p regulated p53 acetylation via HDAC1, GCs were treated
with miR-874-3p inhibitor in the presence of HDAC1. HDAC1
overexpression markedly decreased p53 acetylation compared with the
vector group. However, in HDAC1-overepression GCs, miR-874-3p
inhibitor did not further decrease p53 acetylation and expression
(Fig. 5B). The results suggested
that miR-874-3p promoted p53 acetylation and expression via
inhibition of HDAC1.
Discussion
To the best of our knowledge, the present study was
the first to investigate miR-874-3p expression in PCOS and its role
in GC apoptosis. Recently, the role of miRNAs in the pathogenesis
of PCOS has attracted increasing attention (22). Altered miRNA expression profiles in
patients with PCOS have been regarded as biomarkers and therapeutic
targets for PCOS (22,23). Testosterone-induced GC apoptosis is
involved in the pathological development of PCOS (7). However, the role of miRNAs in
hyperandrogen-induced GC apoptosis is not completely understood.
The present study demonstrated that miR-874-3p was positively
correlated with testosterone levels, and miR-874-3p expression was
upregulated by testosterone treatment compared with the vehicle
group, which suggested that miR-874-3p may serve as a diagnostic
marker for PCOS. Moreover, gain- and loss-of-function experiments
indicated that miR-874-3p mediated the proapoptotic effects of
testosterone, suggesting that miR-874-3p may serve a role in
testosterone-induced GC apoptosis. Since miR-874-3p is a newly
identified miRNA, the present study furthered the current
understanding of the role of miR-874-3p.
HDACs are enzymes that catalyze the deacetylation of
lysine residues of both histone and non-histone proteins (24). Abnormal HDAC activity is involved in
a number of diseases, including cancer, neurological disease,
metabolic disorders, cardiac disease and inflammatory diseases
(25). To the best of our
knowledge, the present study was the first to demonstrate that HDAC
activity and HDAC1 expression were notably decreased in
testosterone-treated GCs compared with vehicle-treated GCs. A
previous study also reported that the expression of SIRT1, another
HDAC family member, was also downregulated in a PCOS model
(26). Moreover, females receiving
HDAC inhibitor displayed polycystic ovaries (27). Therefore, low HDAC activity may
serve as a mechanism underlying the development of PCOS. However,
in the present study, SIRT1 expression was not altered in
testosterone-treated GCs compared with vehicle-treated GCs,
suggesting that HDAC1 might be the primary target of
testosterone/miR-874-3p and that SIRT1 might be regulated by other
factors in PCOS. The present study indicated that the androgen
receptor antagonist flutamide inhibited testosterone-induced
upregulation of miR-874-3p, an upstream factor of HDAC1.
Interestingly, it has also been reported that HDAC1 can regulate
the function of the androgen receptor (28,29).
Thus, there might be a feedback mechanism controlling the effects
of HDAC1 and androgen receptor. In the present study, HDAC1 was
overexpressed in GCs and the results indicated that HDAC1 protected
against GC apoptosis. Therefore, HDAC1 may serve as a potential
therapeutic target for PCOS.
The p53 protein is a proapoptotic protein that
serves a central role in the cellular response to DNA damage and
other genomic aberrations (30).
The present study demonstrated that compared with the control
group, p53 accumulation was markedly increased by testosterone
treatment, which might explain the proapoptotic role of
testosterone. However, another study reported that testosterone
reduced p53 expression levels during oxidative stress damage
(31). The contradiction between
the results of the present study and the aforementioned study
suggested that testosterone might differentially regulate p53
expression depending on the pathological circumstances and cell
type. Acetylation is one of the characteristics of stabilized p53,
and upon deacetylation, p53 expression is maintained at low levels
by degradation (21). The results
of the present study indicated that both testosterone treatment and
miR-874-3p overexpression induced p53 acetylation, whereas
miR-874-3p knockdown and HDAC1 overexpression lead to p53
deacetylation. The results suggested that
testosterone/miR-874-3p/HDAC1 signaling regulated p53 acetylation.
Moreover, miR-874-3p inhibitor did not further decrease p53
acetylation and expression in HDAC1-overexpression GCs, indicating
that miR-874-3p regulated p53 expression by suppressing
HDAC1-mediated deacetylation of p53 but not via other mechanisms.
Previous studies demonstrated that HDAC1 mediated the deacetylation
of p53 (32,33), confirming that p53 could be
deacetylated by HDAC1. Collectively, the aforementioned results
indicated that testosterone/miR-874-3p promoted p53 acetylation and
expression to induce GC apoptosis.
The present study demonstrated that miR-874-3p was
upregulated in PCOS and promoted testosterone-induced GC apoptosis
by suppressing HDAC1-mediated p53 deacetylation. Therefore, the
present study furthered the current understanding of the
pathogenesis of PCOS.
Supplementary Material
Cell apoptosis is increased in
granulosa cells isolated from patients with PCOS. Cell apoptosis in
(A) patients without PCOS and (B) patients with PCOS was determined
via flow cytometry and (C) quantified. *P<0.05 vs.
non-PCOS. PCOS, polycystic ovary syndrome.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Natural Science
Foundation of China (grant no. 81601255).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW and CL designed the experiments, drafted the
manuscript and authenticated the data in this study. YW, ZW and CL
contributed to patient recruitment and data collection. YW, ZW, LW,
SL and XQ performed cell culture, reverse
transcription-quantitative PCR, western blotting and flow cytometry
experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of Zaozhuang Maternal and Child Health Care Hospital and
the Animal Ethics Committee of Zaozhuang Maternal and Child Health
Care Hospital [ZFYYLL2016(001) and ZFYYLL2016(002)]. Written
informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Escobar-Morreale HF: Polycystic ovary
syndrome: Definition, aetiology, diagnosis and treatment. Nat Rev
Endocrinol. 14:270–284. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Neven ACH, Laven J, Teede HJ and Boyle JA:
A summary on polycystic ovary syndrome: Diagnostic criteria,
prevalence, clinical manifestations, and management according to
the latest international guidelines. Semin Reprod Med. 36:5–12.
2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Cai L, Ma X, Liu S, Liu J, Wang W, Cui Y,
Ding W, Mao Y, Chen H, Huang J, et al: Effects of upregulation of
Hsp27 expression on oocyte development and maturation derived from
polycystic ovary syndrome. PLoS One. 8(e83402)2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Shen H and Wang Y: Activation of
TGF-β1/Smad3 signaling pathway inhibits the development of ovarian
follicle in polycystic ovary syndrome by promoting apoptosis of
granulosa cells. J Cell Physiol. 234:11976–11985. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yeung CK, Wang G, Yao Y, Liang J, Tenny
Chung CY, Chuai M, Lee KK and Yang X: BRE modulates granulosa cell
death to affect ovarian follicle development and atresia in the
mouse. Cell Death Dis. 8(e2697)2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Mikaeili S, Rashidi BH, Safa M, Najafi A,
Sobhani A, Asadi E and Abbasi M: Altered FoxO3 expression and
apoptosis in granulosa cells of women with polycystic ovary
syndrome. Arch Gynecol Obstet. 294:185–192. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Azhary JMK, Harada M, Takahashi N, Nose E,
Kunitomi C, Koike H, Hirata T, Hirota Y, Koga K, Wada-Hiraike O, et
al: Endoplasmic reticulum stress activated by androgen enhances
apoptosis of granulosa cells via induction of death receptor 5 in
PCOS. Endocrinology. 160:119–132. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zheng Q, Li Y, Zhang D, Cui X, Dai K, Yang
Y, Liu S, Tan J and Yan Q: ANP promotes proliferation and inhibits
apoptosis of ovarian granulosa cells by NPRA/PGRMC1/EGFR complex
and improves ovary functions of PCOS rats. Cell Death Dis.
8(e3145)2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhao KK, Cui YG, Jiang YQ, Wang J, Li M,
Zhang Y, Ma X, Diao FY and Liu JY: Effect of HSP10 on apoptosis
induced by testosterone in cultured mouse ovarian granulosa cells.
Eur J Obstet Gynecol Reprod Biol. 171:301–306. 2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Pal I, Safari M, Jovanovic M, Bates SE and
Deng C: Targeting translation of mRNA as a therapeutic strategy in
cancer. Curr Hematol Malig Rep. 14:219–227. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Feng X, Xue H, Guo S, Chen Y, Zhang X and
Tang X: miR-874-3p suppresses cell proliferation and invasion by
targeting ADAM19 in nasopharyngeal carcinoma. Panminerva Med: Jul
19, 2019 (Epub ahead of print). doi:
10.23736/S0031-0808.19.03682-6.
|
|
12
|
Yuan RB, Zhang SH, He Y, Zhang XY and
Zhang YB: miR-874-3p is an independent prognostic factor and
functions as an anti-oncomir in esophageal squamous cell carcinoma
via targeting STAT3. Eur Rev Med Pharmacol Sci. 22:7265–7273.
2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yan Y, Song X, Li Z, Zhang J, Ren J, Wu J,
Li Y, Guan Y and Wang J: Elevated levels of granzyme B correlated
with miR-874-3p downregulation in patients with acute myocardial
infarction. Biomark Med. 11:761–767. 2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kushwaha P, Khedgikar V, Sharma D, Yuen T,
Gautam J, Ahmad N, Karvande A, Mishra PR, Trivedi PK, Sun L, et al:
MicroRNA 874-3p exerts skeletal anabolic effects epigenetically
during weaning by suppressing Hdac1 expression. J Biol Chem.
291:3959–3966. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Huo W and Li H, Zhang Y and Li H:
Epigenetic silencing of microRNA-874-3p implicates in erectile
dysfunction in diabetic rats by activating the Nupr1/Chop-mediated
pathway. FASEB J. 34:1695–1709. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wang XQ, Bai HM, Li ST, Sun H, Min LZ, Tao
BB, Zhong J and Li B: Knockdown of HDAC1 expression suppresses
invasion and induces apoptosis in glioma cells. Oncotarget.
8:48027–48040. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Demyanenko SV, Dzreyan VA, Neginskaya MA
and Uzdensky AB: Expression of histone deacetylases HDAC1 and HDAC2
and their role in apoptosis in the penumbra induced by
photothrombotic stroke. Mol Neurobiol. 57:226–238. 2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Rotterdam ESHRE/ASRM-Sponsored PCOS
consensus workshop group. Revised 2003 consensus on diagnostic
criteria and long-term health risks related to polycystic ovary
syndrome (PCOS). Hum Reprod. 19:41–47. 2004.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ferrero H, Delgado-Rosas F, Garcia-Pascual
CM, Monterde M, Zimmermann RC, Simón C, Pellicer A and Gómez R:
Efficiency and purity provided by the existing methods for the
isolation of luteinized granulosa cells: A comparative study. Hum
Reprod. 27:1781–1789. 2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Brooks CL and Gu W: The impact of
acetylation and deacetylation on the p53 pathway. Protein Cell.
2:456–462. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chen B, Xu P, Wang J and Zhang C: The role
of miRNA in polycystic ovary syndrome (PCOS). Gene. 706:91–96.
2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Chen Z, Ou H, Wu H, Wu P and Mo Z: Role of
microRNA in the pathogenesis of polycystic ovary syndrome. DNA Cell
Biol. 38:754–762. 2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang P, Wang Z and Liu J: Role of HDACs in
normal and malignant hematopoiesis. Mol Cancer.
19(5)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Seto E and Yoshida M: Erasers of histone
acetylation: The histone deacetylase enzymes. Cold Spring Harb
Perspect Biol. 6(a018713)2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Tao X, Zhang X, Ge SQ, Zhang EH and Zhang
B: Expression of SIRT1 in the ovaries of rats with polycystic ovary
syndrome before and after therapeutic intervention with exenatide.
Int J Clin Exp Pathol. 8:8276–8283. 2015.PubMed/NCBI
|
|
27
|
Isojarvi JI, Laatikainen TJ, Pakarinen AJ,
Juntunen KT and Myllyla VV: Polycystic ovaries and hyperandrogenism
in women taking valproate for epilepsy. N Engl J Med.
329:1383–1388. 1993.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gaughan L, Logan IR, Cook S, Neal DE and
Robson CN: Tip60 and histone deacetylase 1 regulate androgen
receptor activity through changes to the acetylation status of the
receptor. J Biol Chem. 277:25904–25913. 2002.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Welsbie DS, Xu J, Chen Y, Borsu L, Scher
HI, Rosen N and Sawyers CL: Histone deacetylases are required for
androgen receptor function in hormone-sensitive and
castrate-resistant prostate cancer. Cancer Res. 69:958–966.
2009.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ou HL and Schumacher B: DNA damage
responses and p53 in the aging process. Blood. 131:488–495.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Pronsato L and Milanesi L: Effect of
testosterone on the regulation of p53 and p66Shc during oxidative
stress damage in C2C12 cells. Steroids. 106:41–54. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Luo J, Su F, Chen D, Shiloh A and Gu W:
Deacetylation of p53 modulates its effect on cell growth and
apoptosis. Nature. 408:377–381. 2000.PubMed/NCBI View
Article : Google Scholar
|
|
33
|
Chen S, Yao X, Li Y, Saifudeen Z,
Bachvarov D and El-Dahr SS: Histone deacetylase 1 and 2 regulate
Wnt and p53 pathways in the ureteric bud epithelium. Development.
142:1180–1192. 2015.PubMed/NCBI View Article : Google Scholar
|