Introduction
Carotid stenosis is the narrowing or contraction of
the internal surface of the carotid artery (1). In patients with severe carotid
stenosis, the incidence of cerebral ischemic events within 2 years
is as high as 26% (2). Carotid
stenosis is a major risk factor for stroke that leads to brain
damage (3), and it is a common
cause of ischaemic stroke and transient ischaemic attack (4). In >60% of cases of cerebral
infarction, the cause is carotid stenosis, and severe cerebral
infarction may lead to disability or even death of patients
(5). Carotid stenosis seriously
threatens the health of affected individuals. Over the past decade,
carotid angioplasty and stenting (CAS) has been recognized as a
safe and effective treatment for patients with carotid stenosis
(6). However, restenosis remains a
serious and unresolved problem following CAS treatment (7). Therefore, exploring the molecules
related to the occurrence and development of restenosis is of great
significance to prevent restenosis and improve the efficacy of CAS
in the treatment of carotid stenosis. Vascular smooth muscle cells
(VSMCs) are the major cell type in blood vessels. Unlike numerous
other mature cell types in the adult body, VSMCs cannot terminally
differentiate and therefore retain a significant plasticity
(8). VSMC proliferation is an
important feature of restenosis and one of the important
pathological mechanisms of restenosis (9). Thus, exploring the regulatory
mechanisms of VSMC proliferation and testing methods of interfering
with VSMC proliferation are expected to provide novel ideas and
targets for preventing restenosis.
Several studies have indicated that circulating
microRNA (miRNA/miR)-145 is significantly associated with vascular
restenosis after stent implantation (10,11).
Targeted regulation of miR-145 by long-noncoding (lnc)RNA small
nucleolar RNA host gene 1 (SNHG1) (12-14).
As a potential marker of restenosis, miR-145 has the ability to
inhibit VSMC proliferation and migration (15). Yang and Zi (16) reported that SNHG1 participates in
the cerebrovascular pathological process of stroke. SNHG1 also has
the function of regulating endothelial cell function (17). However, the regulatory effect of
SNHG1 on VSMC function, an important mechanism in the pathological
process of restenosis, has not been reported. Therefore, the aim of
the present study was to analyze the expression level of SNHG1 in
patients with restenosis after carotid artery stenosis stenting,
and to explore the regulatory effect of SNHG1 on the proliferation
and migration of VSMCs.
Materials and methods
Patient inclusion and sample
collection
The present study included 20 patients with
restenosis and 30 patients without restenosis underwent CAS for
carotid stenosis at Cangzhou Central Hospital (Cangzhou, China)
between February 2014 and October 2016. The degree of stenosis was
determined using the criteria of the North American Symptomatic
Carotid Endarterectomy trial (18).
Patients were excluded if any of the following applied: i) Acute
infection, cough, fever or diarrhea; ii) chronic inflammatory
diseases; iii) malignant tumors; iv) severe kidney or liver
diseases; v) poor adherence. Blood samples were collected from
subjects after CAS, the blood samples were all centrifuged for
serum isolation immediately after collection to avoid hemolysis in
this experiment and stored at -80˚C for future use. The demographic
and clinicopathological characteristics of the patients were
recorded and listed in Table I,
including age, sex, body mass index, smoking status, drinking
status, diabetes, hypertension, hyperlipidemia, total cholesterol
(TC), triglyceride, high-density lipoprotein cholesterol and
low-density lipoprotein cholesterol (LDL-C). Written informed
consent was obtained from patients or their relatives for the use
of blood samples and clinical data. Experimental procedures were
approved by the guidelines of the Ethics Committee of Cangzhou
Central Hospital (Cangzhou, China; approval no. CZCH14h0280).
 | Table IBaseline characteristics of the study
subjects. |
Table I
Baseline characteristics of the study
subjects.
| Variable | non-ISR (n=30) | ISR (n=20) | P-value |
|---|
| Age (years) | 64.5±7.2 | 67.5±8.5 | 0.199 |
| Male sex | 26 | 17 | 0.868 |
| BMI
(kg/m2) | 24.7±2.5 | 25.1±3.1 | 0.640 |
| Smoking status | 12 | 9 | 0.726 |
| Alcohol
consumption | 12 | 10 | 0.166 |
| Diabetes | 8 | 10 | 0.018 |
| Hypertension | 20 | 16 | 0.304 |
| Hyperlipidemia | 13 | 11 | 0.419 |
| TC (mg/dl) | 149.0±36.3 | 169.88±36.9 | 0.046 |
| TG (mg/dl) | 143.0±48.5 | 134.6±48.0 | 0.548 |
| HDL-C (mg/dl) | 40.1±6.7 | 41.7±7.1 | 0.433 |
| LDL-C (mg/dl) | 96.6±19.9 | 84.2±18.5 | 0.031 |
Cell culture and transfection
The human carotid artery smooth muscle cell
(hHCtASMC) line was purchased from the Cell Applications, Inc. and
was cultured with Medium 231 and Smooth Muscle Growth Supplement
(Thermo Fisher Scientific, Inc.) in an incubator containing air
with 5% CO2 at 37˚C.
Cell transfection was used to achieve the in
vitro regulation of SNHG1 and miR-145 in the present study.
Short interfering (si)RNA targeting SNHG1 (si-SNHG1), negative
control siRNA (si-NC), mimics NC, miR-145 mimics, inhibitor NC and
miR-145 inhibitor were synthesized from GenePharma and were
transfected into hHCtASMCs using Lipofectamine 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturers'
protocol . Cells treated with only transfection reagen ts were set
as a mock group. After 48 h of transfection, the cells were used
for further analyses. The transfected sequences were as follows
(from 5' to 3'): si-SNHG1, 5'-CUUAAAGUGUUAGCAGACATT-3'; si-NC,
5'-UUCUCCGAACGUGUCACGUTT-3'; miR-145 mimic,
5'-GUCCAGUUUUCCCAGGAAUCCCU-3'; miR-145 inhibitor,
5'-AGGGAUUCCUGGGAAAACUGGAC-3'; mimics NC,
5'-UUCUCCGAACGUGUCACGU-3'; inhibitor NC,
5'-CAGUACUUUUGUGUAGUACAA-3'.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was obtained from fresh serum samples and
cells with TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and the RNA was reversely transcribed into single-stranded
complementary DNA with a PrimeScript reverse transcriptase reagent
kit (Takara Bio, Inc.) following the manufacturer's protocol. The
expression of SNHG1 and miR-145 was measured by real-time qPCR,
which was performed using a SYBR-Green I Master Mix kit
(Invitrogen; Thermo Fisher Scientific, Inc.) on a 7500 Real-Time
PCR System (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The following thermocycling conditions were used: Initial
denaturation at 95˚C for 10 min; followed by 40 cycles of 95˚C for
20 sec, 58˚C for 15 sec and 72˚C for 20 sec. U6 was used as an
endogenous control for miR-145 and GAPDH was used as an endogenous
control for SNHG1. The oligonucleotide primer sequences were as
follows: SNHG1 forward, 5'-CCCCATGATGGTTCCTCAGTT-3' and reverse,
5'-GGAAAGCAAGTGCAGGTTAGTC-3'; GAPDH forward,
5'-TGCACCACCAACTGCTTAGC-3' and reverse,
5'-GGCATGCACTGTGGTCATGAG-3'; miR-145 forward,
5'-GCCGAGGUCCAGUUUUCCCC-3' and reverse, 5'-CTCAACTGGTGTCGTGGA-3';
U6 forward, 5'-CTCGCTTCGGCAGCACA-3' and reverse,
5'-AACGCTTCACGAATTTGCGT-3'. The final expression value was
calculated using the 2-ΔΔCq method (19).
Cell proliferation analysis
A Cell Counting Kit 8 (CCK-8) assay was used to
analyze the cell proliferation. Cells were seeded into a 96-well
plate at 5x103 cells/well and incubated for 0, 24, 48 or
72 h. Subsequently, as per the manufacturer's protocol, 10 µl CCK-8
reagent (Beyotime Institute of Biotechnology) was added to each
well, followed by further incubation for 2 h. The optical density
at 450 nm was determined using a microplate reader.
Transwell assay
Transwell chambers (Corning, Inc.) 8.0 µm pore size
filter membranes (Corning, Inc.) without Matrigel coating (for
migration assay) were used to analyze the migration ability of
hHCtASMCs. Cells (2x105 cells/well) in serum-free
culture medium were seeded into the upper chambers, while the lower
chambers were filled with medium containing 2% FBS (Gibco; Thermo
Fisher Scientific, Inc.). After incubation at 37˚C for 48 h, the
cells that had migrated to the lower chambers were stained with
0.1% crystal violet for 10 min at room temperature and the cell
number in five random fields was counted under an inverted
microscope (Olympus Corp.).
Luciferase reporter assay
The bioinformatics database, StarBase v.2.0
(http://starbase.sysu.edu.cn/starbase2/), was used to
decipher miRNA-target interactions. After determining that miR-145
contained a binding site of SNHG1, a luciferase reporter assay was
used to confirm the interaction between miR-145 and SNHG1. The
wild-type (WT) and mutant-type (MUT) of the SNHG1 sequence that
contained the binding site for miR-145 were individually combined
into the pGL3-luciferase basic reporter vector (Promega Corp.). The
MUT sequence was obtained using a QuickMutation kit (Beyotime
Institute of Biotechnology). The respective combined vectors were
then co-transfected into hHCtASMCs with miR-145 mimics or mimics NC
using Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific,
Inc.). Changes in luciferase activity were analyzed by Dual
Luciferase Reporter Assay System (Promega Corp.) and normalized to
Renilla luciferase activity.
Statistical analysis
Values are expressed as the mean ± standard
deviation and analyzed with SPSS 21.0 (IBM Corp.) and GraphPad 7.0
(GraphPad Software, Inc.). Differences between groups were analyzed
with Student's t-test or one-way ANOVA with Tukey's
multiple-comparison test. Pearson correlation analysis was used to
analyze the correlation between the expression of SNHG1 and
miR-145. Logistic regression analysis was used to analyze the
correlation between SNHG1, miR-145 and the occurrence of
restenosis. P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression of SNHG1 and miR-145 in
patients with restenosis
The relative expression levels of SNHG1 and miR-145
in the serum of patients with and without restenosis was determined
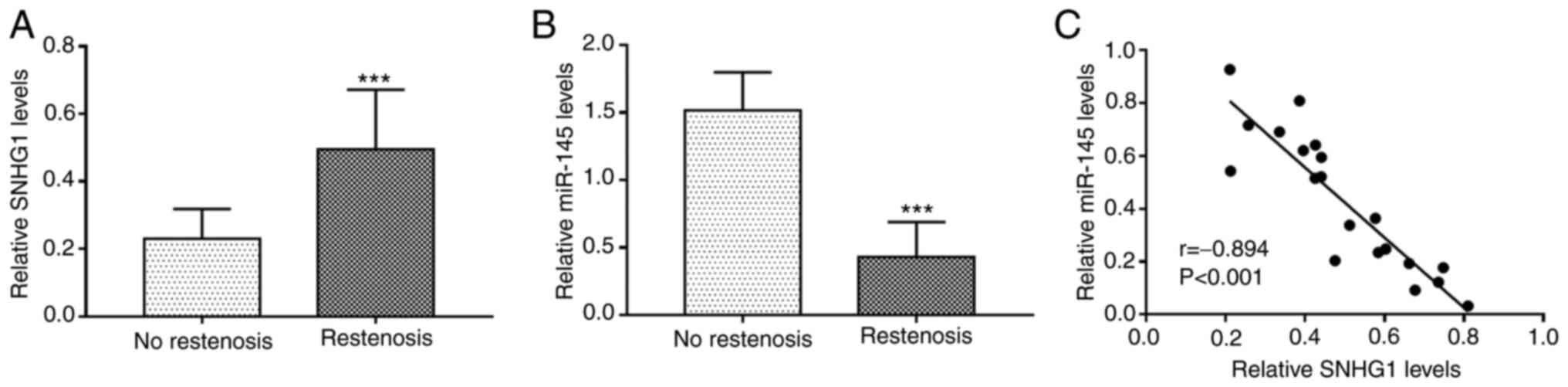
by RT-qPCR. The results revealed that the expression of SNHG1 was
significantly upregulated (P<0.001; Fig. 1A), while that of miR-145 was
downregulated (P<0.001; Fig. 1B)
in the serum of patients with restenosis compared with the patients
without restenosis. There was a significant negative correlation
between the expression levels of SNHG1 and miR-145 in serum of
patients with restenosis (r=-0.894; P<0.001; Fig. 1C).
Association of SNHG1 and miR-145 with
restenosis
The clinical data of all patients, including age,
sex, BMI, smoking and drinking status, hypertension,
hyperlipidemia, TC, TG, HDL-C, LDL-C and the expression levels of
SNHG1 and miR-145 were included in the univariate and multivariate
logistic regression analysis. The continuous data, including age,
TC, TG, HDL-C, LDL-C, SNHG1 and miR-145, were classified based on
median value, and 24 kg/m2 was used as a threshold for
BMI. The results indicated that SNHG1 (P=0.008), miR-145 (P=0.002),
diabetes (P=0.049), TC (P=0.044) and LDL-C (P=0.032) were
independently associated with the occurrence of restenosis and are
thus potential risk factors for the occurrence of restenosis in
patients following CAS (Table
II).
| Table IIResults of the logistic analysis for
patients with restenosis. |
Table II
Results of the logistic analysis for
patients with restenosis.
| | Univariate
analysis | Multivariate
analysis |
|---|
| Variable | OR | 95% CI | P-value | OR | 95% CI | P-value |
|---|
| Age (≥65 years vs.
<65 years) | 1.531 | 0.862-2.371 | 0.268 | 1.325 | 0.856-2.369 | 0.286 |
| Gender (male vs.
female) | 1.226 | 0.701-1.894 | 0.794 | 1.123 | 0.952-1.236 | 0.758 |
| BMI (≥24
kg/m2 vs. <24 kg/m2) | 1.112 | 0.891-2.555 | 0.121 | 1.523 | 0.758-2.569 | 0.162 |
| Smoking status (yes
vs. no) | 1.336 | 0.740-2.397 | 0.549 | 1.156 | 0.652-3.296 | 0.521 |
| Alcohol consumption
(yes vs. no) | 1.401 | 0.812-2.306 | 0.521 | 2.212 | 0.785-4.621 | 0.596 |
| Diabetes (yes vs.
no) | 1.505 | 1.038-2.377 | 0.041 | 1.526 | 1.068-2.768 | 0.049 |
| Hypertension (yes
vs. no) | 1.520 | 1.013-1.886 | 0.046 | 2.425 | 0.856-3.965 | 0.132 |
| Hyperlipidemia (yes
vs. no) | 1.131 | 0.801-2.089 | 0.407 | 1.132 | 0.925-1.995 | 0.856 |
| TC (high vs.
low) | 1.721 | 1.124-2.579 | 0.031 | 2.569 | 1.451-3.874 | 0.044 |
| TG (high vs.
low) | 1.524 | 0.951-2.185 | 0.082 | 1.589 | 0.802-3.552 | 0.356 |
| HDL-C (high vs.
low) | 1.208 | 0.712-1.941 | 0.507 | 1.124 | 0.936-1.696 | 0.784 |
| LDL-C (low vs.
high) | 1.896 | 1.375-3.752 | 0.016 | 2.232 | 1.521-4.132 | 0.032 |
| miR-145 (low vs.
high) | 2.597 | 1.812-3.997 | <0.001 | 2.896 | 1.812-4.712 | 0.002 |
| SNHG1 (high vs.
low) | 2.366 | 1.672-3.413 | 0.001 | 2.751 | 1.685-4.556 | 0.008 |
Knockdown of SNHG1 inhibits the
proliferation and migration of hHCtASMCs
In the present study, the regulatory effect of SNHG1
on the proliferation and migration of hHCtASMCs, which is
associated with restenosis, was analyzed. By transfection of
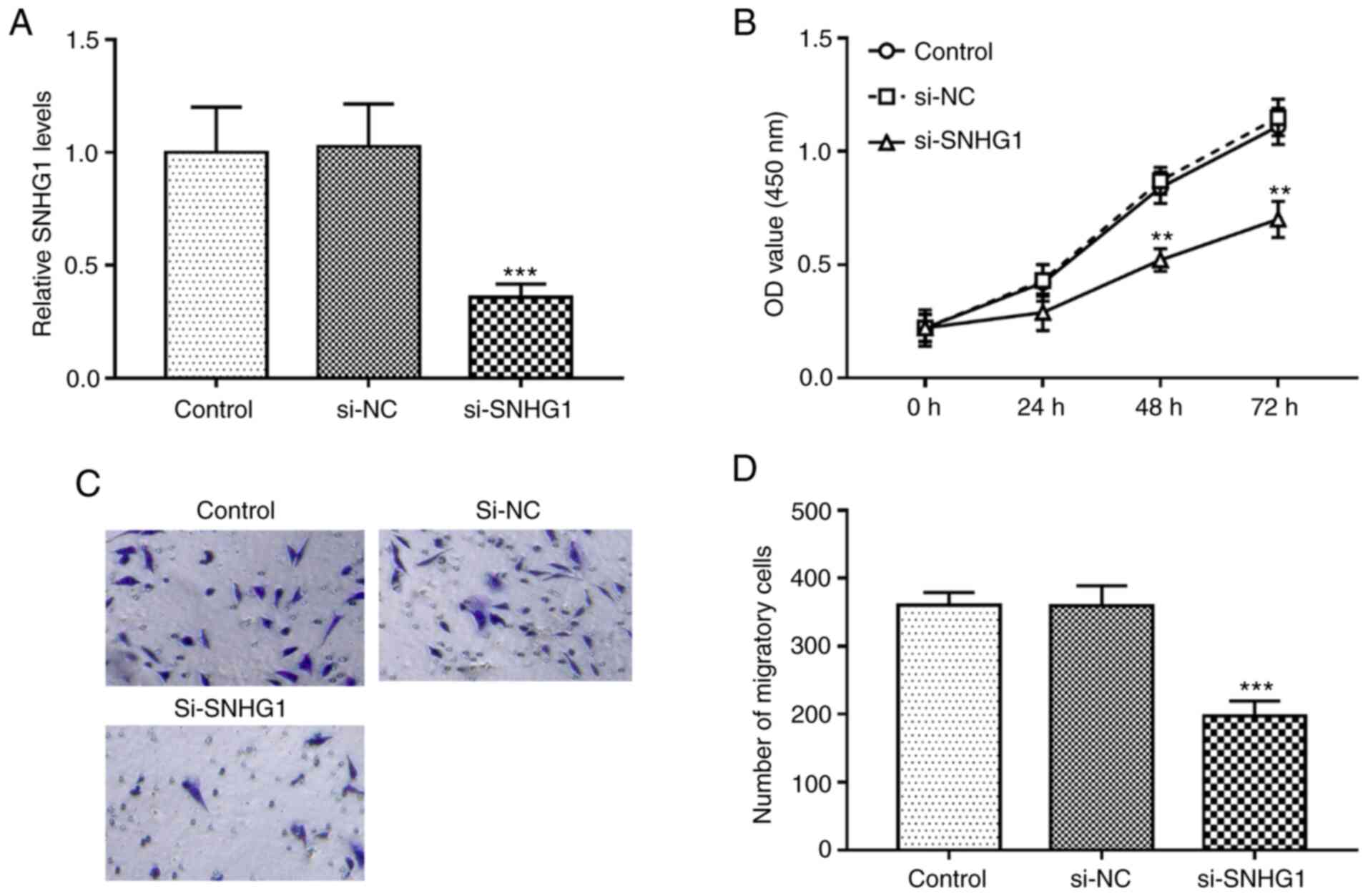
si-SNHG1, the expression levels of SNHG1 in hHCtASMCs were
successfully downregulated (P<0.001; Fig. 2A). The results of the in
vitro assays indicated that cell proliferation and migration
were significantly inhibited after silencing of SNHG1 (all
P<0.01; Fig. 2B-D).
SNHG1 directly inhibits miR-145
expression in hHCtASMCs
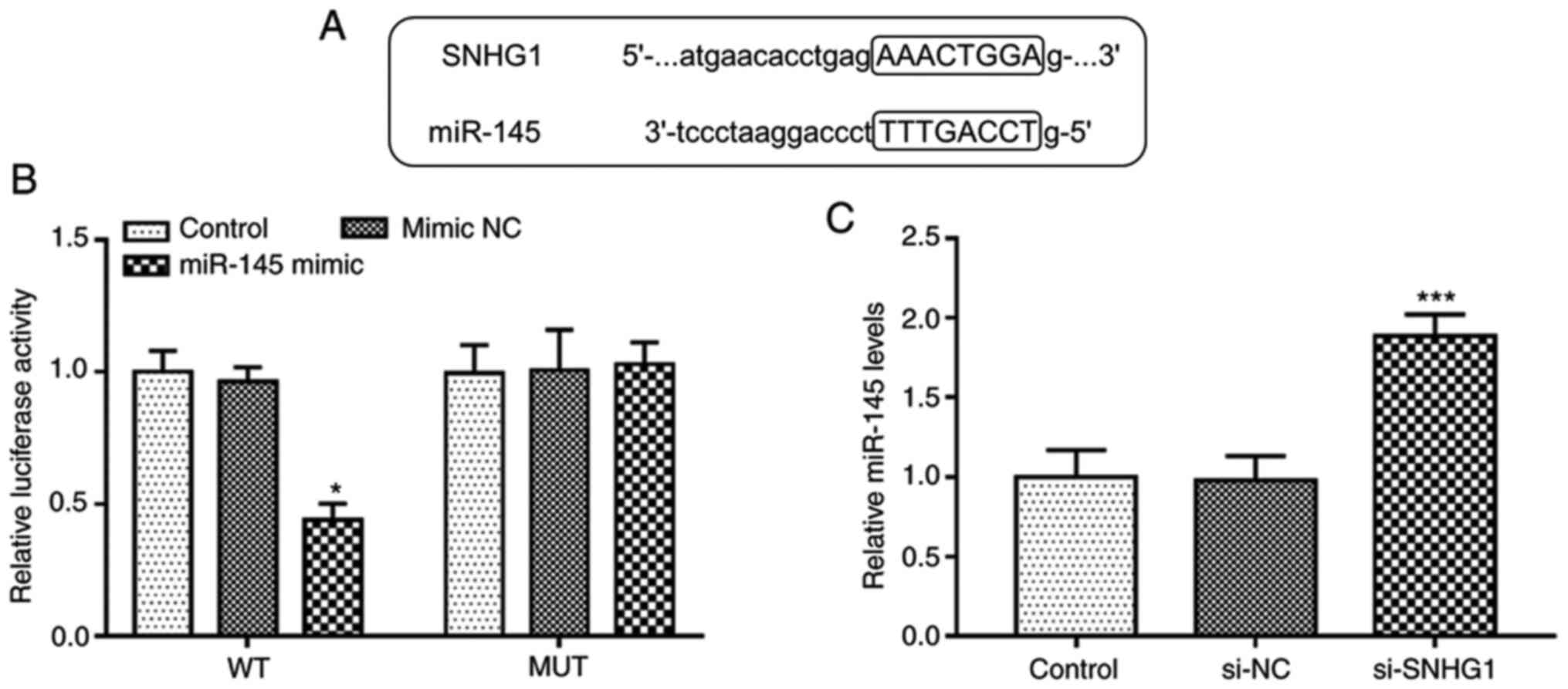
The complementary sequences of miR-145 and SNHG1 at
StarBase v.2.0 web are presented in Fig. 3A. To confirm the accuracy of this
prediction, a luciferase reporter assay was performed in hHCtASMCs.
The experimental results indicated that upregulation of miR-145
suppressed the luciferase activity in the WT-SNHG1 group
(P<0.05; Fig. 3B), while it was
not affected in the MUT group. Furthermore, RT-qPCR, indicated that
silencing of SNHG1 significantly increased the expression levels of
miR-145 in hHCtASMCs (P<0.05; Fig.
3C). It was thus suggested that SNHG1 is able to directly
interfere with the expression of miR-145 in hHCtASMCs.
miR-145 mediates the effects of SNHG1
on hHCtASMC proliferation and migration
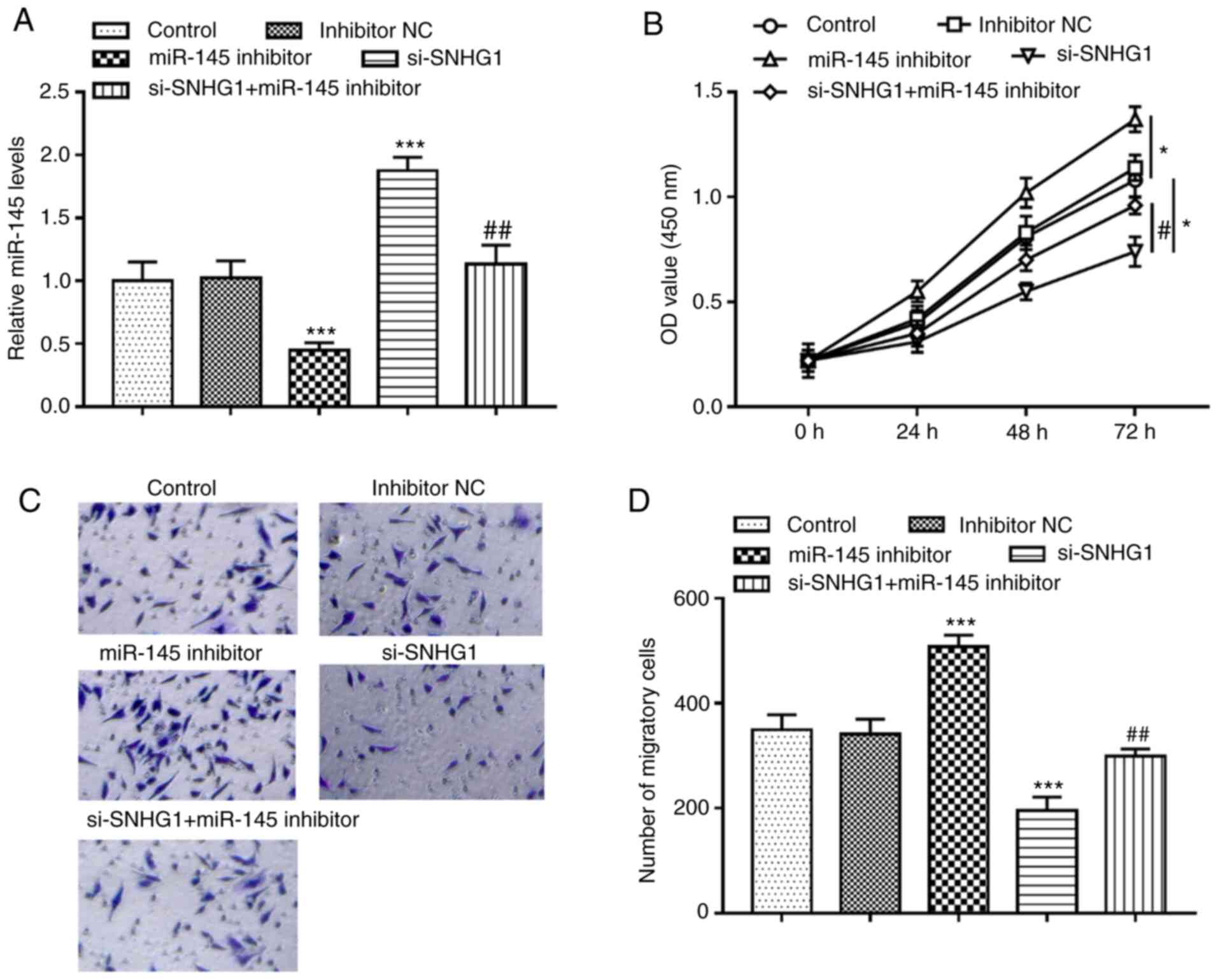
miR-145 inhibitor significantly inhibited the
expression of miR-145, but knockout of SNHG1 promoted the
expression of miR-145 in hHCtASMCs. When hHCtASMCs were
co-transfected with si-SNHG1 and miR-145 inhibitor, it was
indicated that miR-145 inhibitor was able to abolish the increase
of miR-145 expression caused by knockout of SNHG1 (all P<0.01;
Fig. 4A). Analysis of the cells'
activities under different treatment conditions revealed that
silencing of miR-145 significantly promoted cell proliferation and
migration but knockout of SNHG1 suppressed cell proliferation and
migration, while in the case of co-regulation of SNHG1 and miR-145,
it was observed that miR-145 inhibitor significantly reversed the
inhibitory effect of SNHG1 silencing on the proliferation and
migration of hHCtASMCs (all P<0.05; Fig. 4B-D), indicating that miR-145
mediated the regulatory effect of SNHG1 on cell functions of
hHCtASMCs.
Discussion
Carotid stenosis is a major risk factor for stroke
that leads to brain damage (3). In
the last decade, CAS has been developed into a viable alternative
treatment to carotid endarterectomy for patients with symptomatic
moderate and high-grade stenosis (20). However, restenosis still remains an
unresolved issue following CAS treatment (7). Thus, in order to develop more
effective therapeutic approaches for preventing restenosis, an
improved understanding of the molecular mechanisms of restenosis is
important. One of the important characteristics of restenosis is
abnormal proliferation of VSMCs (21). Therefore, it is urgent to understand
the molecular mechanisms underlying restenosis and develop specific
drugs that target VSMCs. lncRNAs are critical regulatory factors
for VSMC function (22). For
instance, lncRNA-taurine upregulated gene 1 knockdown attenuated
oxidized LDL-induced injury through regulating the proliferation
and apoptosis of VSMCs and human umbilical vein endothelial cells
via the miR-148b/insulin-like growth factor 2 axis, providing a
novel mechanism for the pathogenesis of atherosclerosis (23). In occlusive vascular disease, lncRNA
nuclear enriched abundant transcript 1 was discovered as a novel
therapeutic target (24). Tao et
al (25) indicated that
maternally expressed gene 3 may be a negative regulator of spiral
artery remodeling via suppressing extravillous trophoblast (EVT)
invasion and EVT-mediated VSMC loss. However, the effect of lncRNAs
on the proliferation and migration of VSMCs during restenosis has
remained largely elusive.
SNHG1 is an lncRNA that is located at 11q12.3 and
has 11 exons (26). It has been
indicated that SNHG1 is able to participate in cell biological
activities, including cell proliferation, migration, invasion and
apoptosis (27). SNHG1 has been
found to be involved in the pathogenesis of certain diseases. For
instance, lncRNA SNHG1 exerted protective effects against
oxygen/glucose deprivation-induced injury via sponging miR-338,
thus upregulating hypoxia-inducible factor (HIF)-1α/VEGF-A in brain
microvascular endothelialscells (16). Zhang et al (28) indicated that SNHG1 exerted a
neuroprotective effect mediated by HIF-1α/VEGF signaling through
acting as a competing endogenous (ce)RNA for miR-18a. These results
revealed a novel function of SNHG1, contributed to a broad
understanding of ischemic stroke and provided novel therapeutic
options for this disease. However, the results of the present study
demonstrated an opposite role of SNHG1 in the progression of
restenosis after CAS. It is revealed that the inhibition of SNHG1
served a protective role against restenosis by inhibiting carotid
artery smooth muscle cell proliferation and migration. The effect
of SNHG1 on the pathologic changes of blood vessels may be
dependent on the different pathological states and the types of
cells affected by SNHG1, such as endothelial cells or smooth muscle
cells. In the present study, SNHG1 was significantly upregulated in
serum samples of patients with restenosis. In the present study,
serum samples but not plasma samples were used, as plasma contains
fibrinogen, which may affect certain proteinsindicators (29). In numerous studies of the roles of
ncRNAs in the human circulation, serum samples may better reflect
the exact differential expression of these RNAs in various
pathologicalsstates (30-32).
Furthermore, the blood samples were all centrifuged for serum
isolation immediately after collection to avoid hemolysis in this
experiment. In addition, the in vitro experimental results
indicated that knockout of SNHG1 significantly inhibited hHCtASMC
migration and proliferation. The results of the logistic
multivariate regression analysis suggested that SNHG1 was
independently associated with the occurrence of restenosis and was
a potential risk factor for the occurrence of restenosis.
Therefore, SNHG1 may have a regulatory role in the development of
restenosis.
In recent years, the concept of ceRNA has been
proposed as a novel regulatory mechanism in a variety of
pathological processes, meaning that lncRNA may function as a ceRNA
to sponge miRNA and competitively interact withsmiRNAs (33). In addition, miRNAs have an important
role in the diagnosis of various diseases. Diagnosis of human
diseases at an early stage contributes to an increased probability
of cure and blood tests are easier and faster to perform than
tissue tests for early disease screening. Thus, blood tests are
considered comparatively more feasible and noninvasive, and may be
performed morlyfrequently (34). In
the present study, miR-145 binding sites on SNHG1 were discovered.
Based on available studies, downregulation of miR-145 in a wide
range of diseases, including immune-mediated neuroinflammators
diseases (35), colorectal cancer
(CC) (36), non-small cell lung
cancer (NSC) (37) and breasr
cancer (38), may provide novel
diagnostic approaches. Previous studies have indicated that the
SNHG1/miR-145 axis has an important role in a variety of diseases.
For instance Lu et al (12)
suggested that downregulation of SNHG1 inhibited NSCLC cell
viability, proliferation, migration and invasion, but this
inhibition was alleviated by miR-145-5p, which indicated that SNHG1
promoted NSCLC progression by regulating the miR-145-5p/metadherin
(MTDH) axis. In CRC, SNHG1 promotes cell proliferation by acting as
a sponge of miR-145, suggesting that SNHG1/miR-145 may be a
potential target for CRt treatment (13). The present data confirmed that
miR-145 was overexpressed in patients with restenosis. Furthermore,
inhibition of miR-145 in hHCtASMCs promoted cell migration and
proliferation, and the expression level of SNHG1 was negatively
correlated with miR-145 in patient serum. Furthermore, a luciferase
reporter assay confirmed that SNHG1 was able to specifically
associate with miR-145 in hHCtASMCs. SNHG1 affected hHCtASMC
proliferation and migration by regulating miR-145. Furthermore,
inhibition of miR-145 partially reversed the inhibition of hHCtASMC
proliferation and migration by SNHG1 knockout. Thus, based on
previous studies and the results of the present experiments, SNHG1
may be involved in the progression of restenosis by directly
regulating miR-145.
However, there are certain limitations to the
present study, including small sample size, no deconvolution
analysis to reduce cell-type heterogeneity, as well as the lack of
cell experiments under pathological conditions, animal experiments
and further mechanistic analyses. Therefore, further studies are
required to explore the effect of the SNHG1/miR-145 axis on the
function of VSMCs under pathological conditions, such as hypoxia,
as well as to confirm the role of the SNHG1/miR-145 axis in
restenosis in vivo.
Overall, the present study indicated that the
SNHG1/miR-145 axis regulates the proliferation and migration of
hHCtASMCs, and both SNHG1 and miR-145 are independently associated
with and potential risk factors for restenosis. The present results
suggested that the SNHG1/miR-145 axis may serve as a meaningful
non-invasive biomarker for the prevention and treatment of
restenosis and provide novel insight into the pathogenesis of
restenosis.
Acknowledgements
Not applicable.
Funding
The current study was supported by the Cangzhou Science and
Technology Plan Project grant no. 151302109).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HM analyzed and interpreted the data. HM and AD
performed the experiments and wrote and revised the manuscript. HM
and AD checked and confirmed the authenticity of the raw data, and
read and approved the final manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from each
patient and the experimental procedures were approved by the
guidelines of the Ethics Committee of Cangzhou Central Hospital
(Cangzhou, China; approval no. CZCH14h0280).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Berman SS, Bernhard VM, Erly WK, McIntyre
KE, Erdoes LS and Hunter GC: Critical carotid artery stenosis:
Diagnosis, timing of surgery, and outcome. J Vasc Surg. 20:499–508.
1994.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Puccinelli F, Roffi M, Murith N and
Sztajzel R: Management of carotid artery stenosis. Rev Med Suisse.
13:894–849. 2017.PubMed/NCBI(In French).
|
|
3
|
Kelly R: Selections from current
literature: Prevention of stroke in non-rheumatic atrial
fibrillation and carotid artery stenosis. Fam Pract. 9:231–236.
1992.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Cheng SF, Brown MM, Simister RJ and
Richards T: Contemporary prevalence of carotid stenosis in patients
presenting with ischaemic stroke. Br J Surg. 106:872–878.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ijas P, Nuotio K, Vikatmaa P and Soinne L:
Carotid artery stenosis as predictor of the risk of cerebral and
cardiac infarction. Duodecim. 130:2193–2200. 2014.PubMed/NCBI(In Finnish).
|
|
6
|
Fanelli F, Boatta E, Cannavale A, Corona
M, Lucatelli P, Wlderk A, Cirelli C and Salvatori FM: Carotid
artery stenting: Analysis of a 12-year single-center experience. J
Endovasc Ther. 19:749–756. 2012.PubMed/NCBI View Article : Google Scholar
|
|
7
|
AbuRahma AF, Abu-Halimah S, Hass SM,
Nanjundappa A, Stone PA, Mousa A, Lough E and Dean LS: Carotid
artery stenting outcomes are equivalent to carotid endarterectomy
outcomes for patients with post-carotid endarterectomy stenosis. J
Vasc Surg. 52:1180–1187. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Frismantiene A, Philippova M, Erne P and
Resink TJ: Smooth muscle cell-driven vascular diseases and
molecular mechanisms of VSMC plasticity. Cell Signal. 52:48–64.
2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhang MJ, Zhou Y, Chen L, Wang YQ, Wang X,
Pi Y, Gao CY, Li JC and Zhang LL: An overview of potential
molecular mechanisms involved in VSMC phenotypic modulation.
Histochem Cell Biol. 145:119–130. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yuan Y, Liu X, Hao S, He Q and Shen Z:
Plasma levels of miR-143 and miR-145 are associated with coronary
in-stent restenosis within 1 year of follow-up after drug-eluting
stent implantation. Ann Transl Med. 8(756)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
He M, Gong Y, Shi J, Pan Z, Zou H, Sun D,
Tu X, Tan X, Li J, Li W, et al: Plasma microRNAs as potential
noninvasive biomarkers for in-stent restenosis. PLoS One.
9(e112043)2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Lu Q, Shan S, Li Y, Zhu D, Jin W and Ren
T: Long noncoding RNA SNHG1 promotes non-small cell lung cancer
progression by up-regulating MTDH via sponging miR-145-5p. FASEB J.
32:3957–3967. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tian T, Qiu R and Qiu X: SNHG1 promotes
cell proliferation by acting as a sponge of miR-145 in colorectal
cancer. Oncotarget. 9:2128–2139. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lan X and Liu X: lncRNA SNHG1 functions as
a ceRNA to antagonize the effect of miR-145a-5p on the
down-regulation of NUAK1 in nasopharyngeal carcinoma cell. J Cell
Mol Med. 23:2351–2361. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Li L, Mao D, Li C and Li M: miR-145-5p
inhibits vascular smooth muscle cells (VSMCs) proliferation and
migration by dysregulating the transforming growth factor-b
signaling cascade. Med Sci Monit. 24:4894–4904. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yang X and Zi XH: lncRNA SNHG1 alleviates
OGD induced injury in BMEC via miR-338/HIF-1α axis. Brain Res.
1714:174–181. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Liang S, Ren K, Li B, Li F, Liang Z, Hu J,
Xu B and Zhang A: lncRNA SNHG1 alleviates
hypoxia-reoxygenation-induced vascular endothelial cell injury as a
competing endogenous RNA through the HIF-1α/VEGF signal pathway.
Mol Cell Biochem. 465:1–11. 2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Gasecki AP, Hachinski VC, Mendel T and
Barnett HT: Endarterectomy for symptomatic carotid stenosis. Review
of the European and North American symptomatic carotid surgery
trials. Nebr Med J. 77:121–123. 1992.PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Moresoli P, Habib B, Reynier P, Secrest
MH, Eisenberg MJ and Filion KB: Carotid stenting versus
endarterectomy for asymptomatic carotid artery stenosis: A
systematic review and meta-analysis. Stroke. 48:2150–2157.
2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Luo T, Cui S, Bian C and Yu X: Crosstalk
between TGF-β/smad3 and BMP/BMPR2 signaling pathways via miR-17-92
cluster in carotid artery restenosis. Mol Cell Biochem.
389:169–176. 2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhang B, Dong Y and Zhao Z: lncRNA MEG8
regulates vascular smooth muscle cell proliferation, migration and
apoptosis by targeting PPARα. Biochem Biophys Res Commun.
510:171–176. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wu X, Zheng X, Cheng J, Zhang K and Ma C:
lncRNA TUG1 regulates proliferation and apoptosis by regulating
miR-148b/IGF2 axis in ox-LDL-stimulated VSMC and HUVEC. Life Sci.
243(117287)2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ahmed ASI, Dong K, Liu J, Wen T, Yu L, Xu
F, Kang X, Osman I, Hu G, Bunting KM, et al: Long noncoding RNA
NEAT1 (nuclear paraspeckle assembly transcript 1) is critical for
phenotypic switching of vascular smooth muscle cells. Proc Natl
Acad Sci USA. 115:E8660–E8667. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tao H, Liu X, Liu X, Liu W, Wu D, Wang R
and Lv G: lncRNA MEG3 inhibits trophoblast invasion and
trophoblast-mediated VSMC loss in uterine spiral artery remodeling.
Mol Reprod Dev. 86:686–695. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhang M, Wang W, Li T, Yu X, Zhu Y, Ding
F, Li D and Yang T: Long noncoding RNA SNHG1 predicts a poor
prognosis and promotes hepatocellular carcinoma tumorigenesis.
Biomed Pharmacother. 80:73–79. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Thin KZ, Tu JC and Raveendran S: Long
non-coding SNHG1 in cancer. Clin Chim Acta. 494:38–47.
2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhang L, Luo X, Chen F, Yuan W, Xiao X,
Zhang X, Dong Y, Zhang Y and Liu Y: lncRNA SNHG1 regulates
cerebrovascular pathologies as a competing endogenous RNA through
HIF-1alpha/VEGF signaling in ischemic stroke. J Cell Biochem.
119:5460–5472. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhang A, Sun H and Wang X: Serum
metabolomics as a novel diagnostic approach for disease: A
systematic review. Anal Bioanal Chem. 404:1239–1245.
2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Xie J and Cao Y: Expression of TGF-β1 and
miR-99a in serum of patients with early spontaneous abortion and
correlation with hormone levels during pregnancy. Exp Ther Med.
17:4593–4597. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhang X, Hu Y, Gong C and Zhang C:
Overexpression of miR-518b in non-small cell lung cancer serves as
a biomarker and facilitates tumor cell proliferation, migration and
invasion. Oncol Lett. 20:1213–1220. 2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Li H, Qiu F, Tian F, Shi X, Gao A, Song L
and Liu J: Changes of miR-155 expression in serum of uremic
patients before and after treatment and risk factors analysis. Exp
Ther Med. 20:3352–3360. 2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The rosetta stone of a hidden RNA
language? Cell. 146:353–358. 2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Shao C, Yang F, Qin Z, Jing X, Shu Y and
Shen H: The value of miR-155 as a biomarker for the diagnosis and
prognosis of lung cancer: A systematic review with meta-analysis.
BMC Cancer. 19(1103)2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Sharaf-Eldin WE, Kishk NA, Gad YZ, Hassan
H, Ali MA, Zaki MS, Mohamed MR and Essawi ML: Extracellular
miR-145, miR-223 and miR-326 expression signature allow for
differential diagnosis of immune-mediated neuroinflammatory
diseases. J Neurol Sci. 383:188–198. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Sheng N, Tan G, You W, Chen H, Gong J,
Chen D, Gong J, Chen D, Zhang H and Wang Z: MiR-145 inhibits human
colorectal cancer cell migration and invasion via PAK4-dependent
pathway. Cancer Med. 6:1331–1340. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Yu W, Ding J, He M, Chen Y, Wang R, Han Z,
Xing EZ, Zhang C and Yeh S: Estrogen receptor beta promotes the
vasculogenic mimicry (VM) and cell invasion via altering the
lncRNA-MALAT1/miR-145-5p/NEDD9 signals in lung cancer. Oncogene.
38:1225–1238. 2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Eades G, Wolfson B, Zhang Y, Li Q, Yao Y
and Zhou Q: lincRNA-RoR and miR-145 regulate invasion in
triple-negative breast cancer via targeting ARF6. Mol Cancer Res.
13:330–338. 2015.PubMed/NCBI View Article : Google Scholar
|