Introduction
Throughout history, arsenic has been recognized as
both an environmental hazard and a therapeutic chemical (1). Arsenic trioxide (ATO) is widely
considered an effective anti-cancer treatment medicine (2) and it is the leading treatment
medicine for acute promyelocytic leukemia. Human populations
exposed to high level of arsenic are at risk of developing skin,
bladder, liver and lung cancers (3). Despite the well-known toxicity of
ATO, it has been used to treat various diseases for centuries
(4).
The kidney is an important organ that not only can
maintain blood pressure and eliminate waste, but also can maintain
and regulate body fluid, such as acid-base balance (5). Kidney damage or nephrotoxicity leads
to the impairment of detoxification and excretion functions, which
can be confirmed by renal markers such as blood urea nitrogen
(BUN), creatinine (CRE) and CRE clearance (6).
Research has shown that ATO can cause kidney damage.
Indeed, oxidation, inflammation and apoptosis have all been linked
to ATO-induced nephrotoxicity (7,8). The
primary mechanism by which ATO causes nephrotoxicity is oxidative
stress (9). Additionally,
oxidative stress can increase reactive oxygen species (ROS)
production and malondialdehyde (MDA) level while decreasing
superoxide dismutase (SOD), catalase (CAT) and glutathione (GSH)
activities, leading to apoptosis (10,11).
Thus, antioxidative agents may mitigate or prevent ATO-induced
nephrotoxicity.
Toll-like receptor-4 (TLR4) acts as a
lipopolysaccharide sensor to induce inflammation by activating
factors that induce inflammation (12). TLR4 may activate the intercellular
nuclear factor-κB (NF-κB), given the relationship between innate
and acquired immunity (13). The
NF-κB pathway is further activated, which leads to the entry of the
p-p65 protein into the nucleus and increases the expressions of the
pro-inflammatory factors (14,15).
ATO is capable of stimulating the formation of ROS
(16). Overproduction of ROS is
thought to activate the NF-κB pathway and result in the
upregulation of pro-inflammatory mediators (17,18).
On the whole, increased oxidative stress and consequent
inflammation may result in a greater number of cells undergoing
apoptosis (19). NF-κB is
implicated in regulating numerous inflammatory genes and
ATO-induced various stimuli can stimulate NF-κB (20). In conclusion, oxidative stress is
recognized to serve a significant role in ATO-induced toxicity
(21).
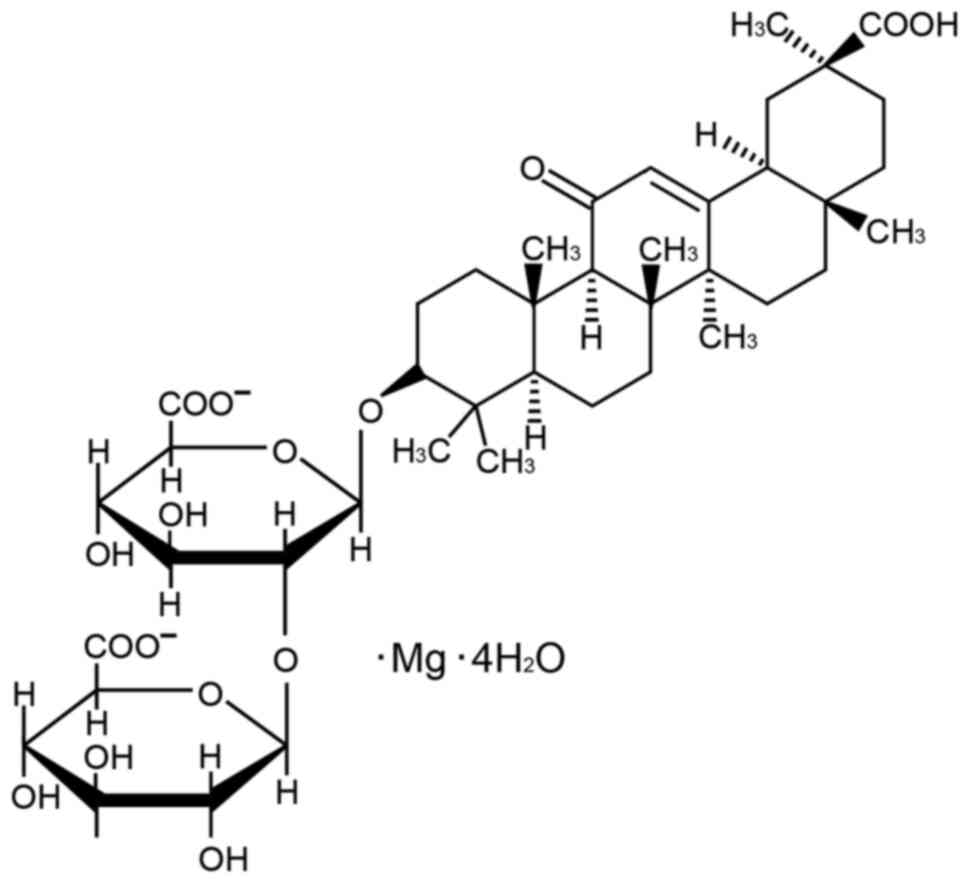
Magnesium isoglycyrrhizinate (MgIG; Fig. 1) is derived from 18-glycyrrhizic
acid, which is a major component of natural licorice
(Glycyrrhiza glabra L.) and has been shown to protect the
liver and possesses anti-inflammatory, anti-oxidative and
anti-apoptotic properties (22,23).
In pharmacological experiments, MgIG has been demonstrated to
suppress the inflammatory process, decrease pathological damage to
hepatocytes and enhance the overall function of hepatocytes
(24). Furthermore, MgIG protects
the heart by inhibiting the TLR4/NF-κB signaling pathway, which is
responsible for activating inflammatory factors (25). Licorice and its active components
have been found to be protective against kidney damage (26). Nevertheless, no studies to date
have examined the possible effects and mechanisms of MgIG on the
nephrotoxicity caused by ATO.
The present study constructed a mouse model of
ATO-induced kidney injury and examined the effects of MgIG on renal
morphological and renal function to investigate the effects and
potential mechanisms of MgIG in ATO-treated mice. In addition, the
regulation of MgIG in the TLR4/NF-κB signaling pathways was
investigated.
Materials and methods
Chemicals and drugs
MgIG for injection was purchased from the Chia Tai
Tianqing Pharmaceutical Group Co., Ltd. ATO parenteral solution was
purchased from Beijing SL Pharmaceutical Co., Ltd. All additional
analytical-grade reagents were acquired from MilliporeSigma.
Animals
A total of 50 male Kunming mice (weight: 22.0±2.0 g,
age: 6-7 weeks) were acquired from the Hebei Medical University
Center for Laboratory Animals. The mice were fed at a normative
temperature (22±2˚C) and humidity (50±10%). The mice were fed a
standard pellet diet and water ad libitum. Animal studies at
Hebei Medical University of Chinese Medicine (Shijiazhuang, China)
were conducted in compliance with the Animal Care and Ethical
Committee (approval no. DWLL2020005) and the United Kingdom Animal
(Scientific Procedures) Act 1986. The Reporting of In Vivo
Experiments guidelines were used to guide the current study
(27).
Experimental design
Male mice were randomly divided into 5 groups
(n=10): The control group (CONT; saline i.p. 10 mg/kg/day), the ATO
group (ATO; i.p. 5 mg/kg/day) (28,29),
the MgIG only group (MgIG; i.p. 50 mg/kg/day), the low-dose MgIG
group (L-MgIG; i.p. 25 mg/kg/day) (30,31)
and the high-dose MgIG group (H-MgIG; i.p. 50 mg/kg/day). With the
exception of the CONT and MgIG groups, all mice were injected i.p.
with ATO (5 mg/kg/day) for a total of 7 days. All 50 mice were
still alive during the experiment and the body weight and status of
the mice were checked daily. The criteria of humane endpoints for
euthanasia were body weight loss (>10%) and anorexia in contrast
with the controls. Following the last administration, the urine of
the mice in each group was collected, after which the mice were
anesthetized with sodium pentobarbital (50 mg/kg) and blood was
drawn from the eyeballs. Then, the mice were sacrificed by
intraperitoneal injection of an overdose of sodium pentobarbital
(200 mg/kg) (28,32). Animal death was confirmed by the
loss of signs of life, such as toe pinch response, heartbeat and
breathing. After mice were sacrificed, kidney tissues were rapidly
collected for subsequent experiments.
Urine and serum chemistry
analysis
Blood (3,500 x g; 10 min; 4˚C) and urine (1,500 x g;
5 min; 4˚C) samples were obtained by centrifugation. The
supernatant was then collected into Doff tubes. The serum and urine
were refrigerated at -20˚C until use. The expression activities of
BUN (cat. no. C013-2-1; Jiancheng Institute of Bioengineering) and
CRE (cat. no. C011-2-1; Jiancheng Institute of Bioengineering) were
determined by using colorimetric assay. Creatinine clearance was
calculated according to the equation: Creatinine clearance
(µl/min/g body weight)=[urinary CRE(µmol/l) x urine
volume(µl/min)/serum CRE(µmol/l) x body weight(g)] (33,34).
Histopathological analysis
The kidney samples of mice were bisected, trimmed
and fixed in 10% formalin at room temperature for 48 h. The samples
were dehydrated in graded ethanol concentrations and then embedded
in paraffin. Paraffin sections (4-µm) were stained with hematoxylin
for 5 min and eosin for 3 min at room temperature. Light microscopy
(Leica DM4000B; Leica Microsystems GmbH) was used to examine
histological alterations. Score 1, 2, 3, 4 and 5 represent the
kidney injury area <10, 10-25, 25-50, 50-75 and >75%,
respectively. The renal injury was assessed by scoring tubular cell
swelling, cellular vacuolization and others under the light
microscopy in ≥10 different horizons and the average score
calculated.
Measurement of oxidative stress and
antioxidant enzymes
The collected serum and kidney tissues were used to
detect the activities of GSH, SOD, CAT, MDA. The expression
activities of GSH (cat. no. A006-2-1; Jiancheng Institute of
Bioengineering), SOD (cat. no. A001-3-2; Jiancheng Institute of
Bioengineering), CAT (cat. no. A007-1-1; Jiancheng Institute of
Bioengineering) and MDA (cat. no. A003-1-1; Jiancheng Institute of
Bioengineering) were determined by using colorimetric assay.
Measurement of ROS levels in the
kidney
Ethoxylation is commonly used as a method to monitor
the production of reactive ROS in cells (35). Dihydroethidium (DHE; cat. no.
G1045l Wuhan Servicebio Technology Co., Ltd.) fluorescence was used
to evaluate ROS production in the kidney. The kidney specimens were
flash-frozen in liquid nitrogen. Then, the freezing microtome
(Cryotome E; Thermo Fisher Scientific, Inc.) was used to cut
slices. Frozen slices (5-µm) were incubated for 5 min with a
spontaneous fluorescence-quenching reagent. The slides were then
rinsed for 10 min with flowing water and the DHE was then dropped
into the indicated region and incubated for 30 min at 37˚C in the
dark. After washing with PBS (pH 7.4) in a Rocker device, the
slides were incubated in a dark location with a DAPI solution for
10 min at room temperature. A total of 10 microscopic fields in
each section were examined in a blinded method. A fluorescence
microscope (Nikon Eclipse C1; Nikon Corporation) was used to
observe and capture images at x200 magnification.
Inflammatory cytokine analysis
The kidney samples were promptly snap-frozen at
-196˚C in liquid nitrogen for standby use. Interleukin-6 (IL-6;
cat. no. 88-7064-88; MultiSciences Biotech Co., Ltd.),
interleukin-1β (IL-1β; cat. no. 88-7013-88; MultiSciences Biotech
Co., Ltd.) and tumor necrosis factor alpha (TNF-α; cat. no.
88-7324-88; Thermo Fisher Scientific, Inc.) levels in the kidney
were measured by relevant ELISA kits.
Bax, Bcl-2, Caspase-3, TLR4, NF-κB and
p-NF-κB by western blotting analysis
RIPA lysis buffer (Wuhan Servicebio Technology Co.,
Ltd.) was used to decompose the kidney samples. Centrifugation
(12,000 x g; 10 min; 4˚C) was used to collect the total protein in
tissue that concentration was determined with the BCA kit (catalog
no. G2026; Wuhan Servicebio Technology Co., Ltd.). Sodium dodecyl
sulfate polyacrylamide gel electrophoresis (10%) was used to
separate equal amounts of protein (15 µg). The proteins were
transferred to polyvinylidene difluoride membranes that were soaked
for overnight at 37˚C in a blocking solution containing 5% skimmed
milk. Then, the membranes were incubated with anti-Bcl-2 associated
X protein (Bax; cat. no. GB11690, 1:1,000; Wuhan Servicebio
Technology Co., Ltd.), anti-B-cell lymphoma 2 (Bcl-2; cat. no.
PAA778Mu01, 1:1,000; Cloud-Clone Corp.), anti-caspase-3 (cat. no.
66470-2-lg, 1:1,000; ProteinTech Group, Inc.), anti-NF-κB (cat. no.
GM1003, 1:2,000; Wuhan Servicebio Technology Co., Ltd.),
anti-p-NF-κB (cat. no. GB11142-1, 1:2,000; Wuhan Servicebio
Technology Co., Ltd.), anti-TLR4 (cat. no. GB11519, 1:1,000; Wuhan
Servicebio Technology Co., Ltd.) and β-actin (cat. no. GB12001,
1:1,000; Wuhan Servicebio Technology Co., Ltd.) antibodies at 4˚C
for 12 h. Afterwards, the membranes were rinsed three times with
PBS. Membranes were incubated for 2 h at 37˚C in darkness with
secondary antibodies of goat anti-mouse IgG-HRP (cat. no. GB23301;
1:3,000; Wuhan Servicebio Technology Co., Ltd.) or goat anti-rabbit
IgG-HRP (cat. no. GB23303; 1:3,000; Wuhan Servicebio Technology
Co., Ltd.) to detect the protein bands. After subjecting the
membranes to a chemiluminescent substrate (ECL; catalog no. G2014;
Wuhan Servicebio Technology Co., Ltd.) and to autoradiographic film
exposure, the optical density value of the target band was analyzed
using Alpha software (Alpha Innotech).
Statistical analysis
Origin Pro 9.1 (OriginLab Corp.) software was used
to perform statistical analyses. Data are demonstrated as mean ±
SEM and one-way analysis of variance (ANOVA) and Tukey's post hoc
test were used to assess statistically significant differences
between groups. Non-parametric data were analyzed by the
Kruskal-Wallis test and the Dunn-Sidak post hoc test (36). P<0.05 was considered to indicate
a statistically significant difference.
Results
Effects of MgIG on alterations in
biochemical indices
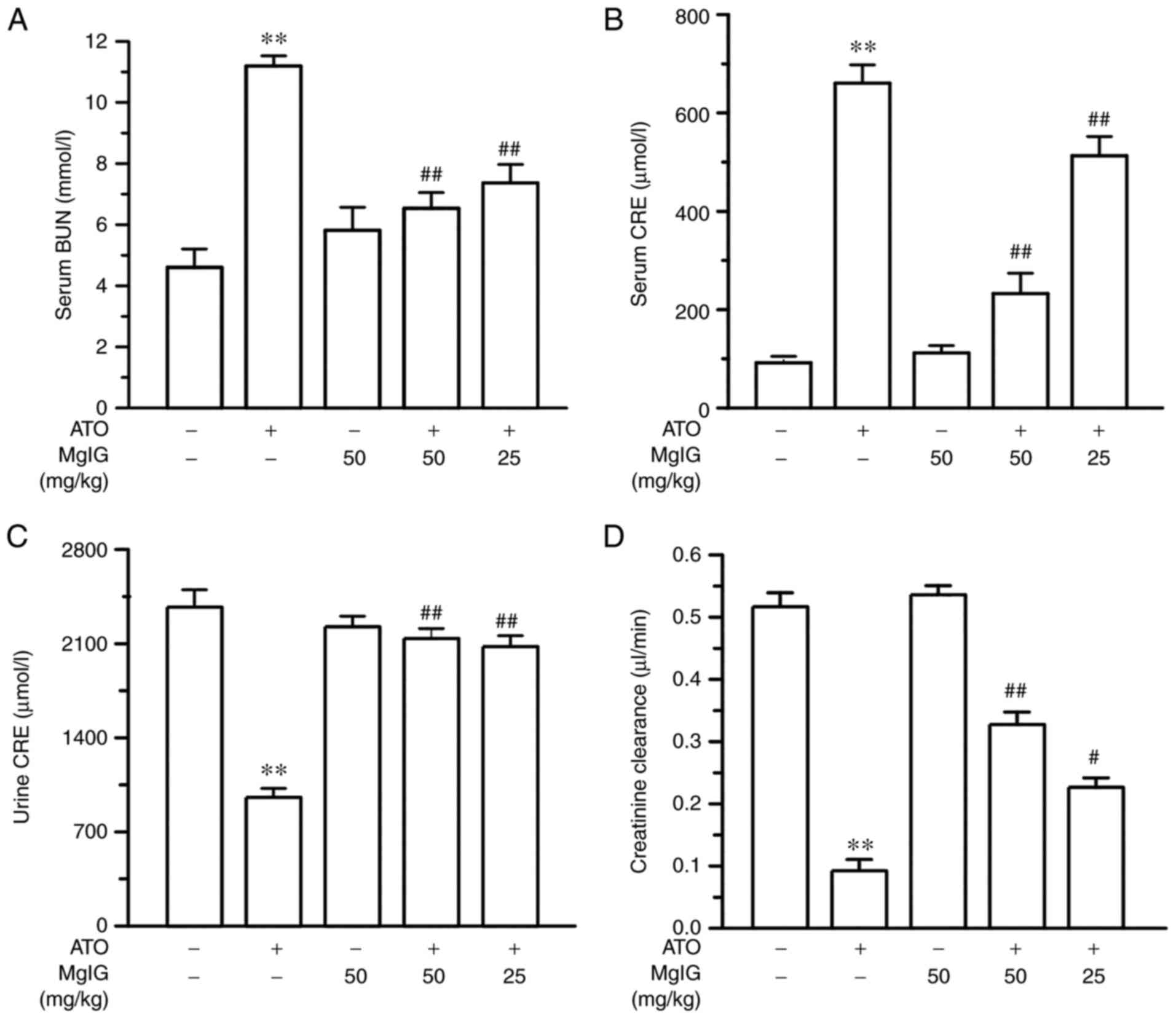
In Fig. 2, the
levels of serum BUN, serum CRE, urinary CRE and CRE clearance were
measured to examine the effect of MgIG on ATO-induced
nephrotoxicity. Serum BUN and CRE levels were notably augmented and
urinary CRE and CRE clearance levels were reduced in the ATO group
(P<0.01) compared with the CONT group. However, the serum BUN
and CRE levels of the H-MgIG and L-MgIG groups were markedly
decreased but urinary CRE and CRE clearance levels were augmented
in contrast with the ATO group (P<0.05 or P<0.01). There was
no significant difference in levels of serum BUN, serum CRE,
urinary CRE and CRE clearance between the CONT group and the MgIG
group (P>0.05). MgIG may have an effect of renal protection.
Effects of MgIG on the
histopathological changes
As shown in Fig. 3,
light microscopy was used to visualize hematoxylin and
eosin-stained kidney slices. No morphological changes were found in
the kidney tissues of the CONT and MgIG groups. By contrast, the
ATO group showed tubular cell swelling, interstitial edema,
inflammatory cell infiltration in the renal interstitial tissue,
glomeruli dilation and hyperemia, partial epithelial cell necrosis
(P<0.01). The mice who received H-MgIG and L-MgIG treatment had
reduced areas of necrosis and inflammatory infiltration (P<0.05
or P<0.01). In other words, treatment of MgIG may ameliorated
ATO-induced kidney damage in a dose-dependent manner.
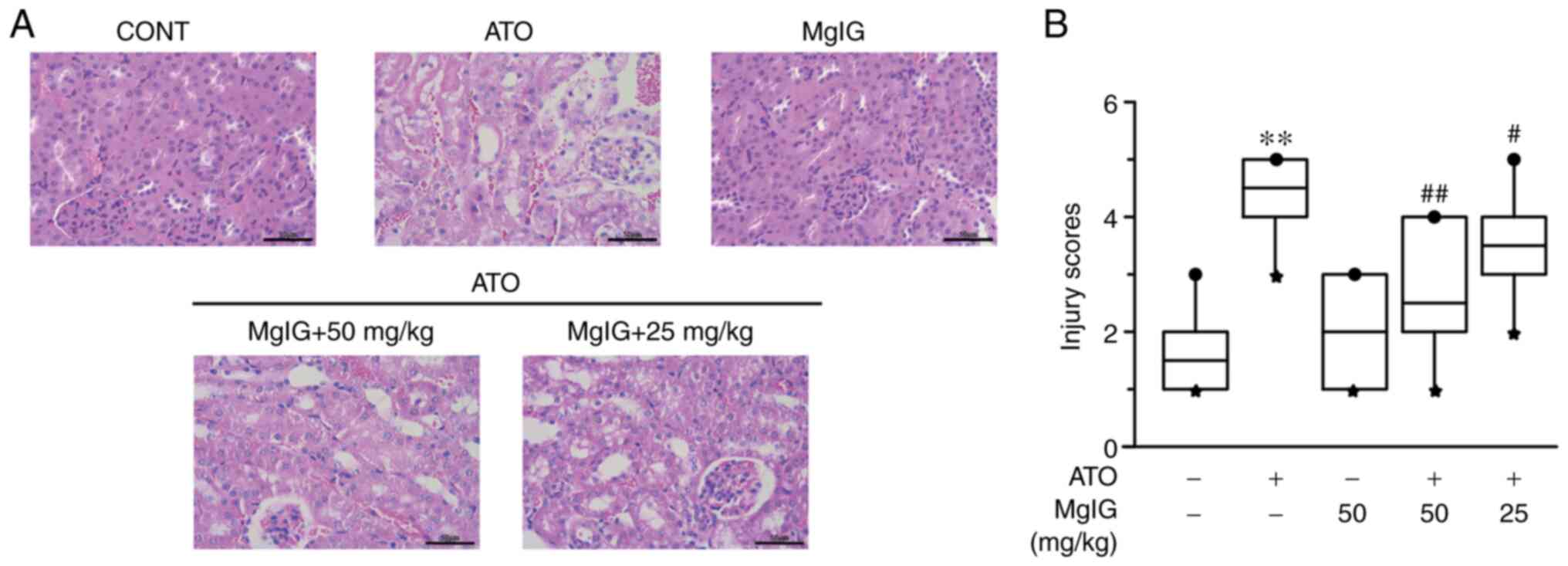 | Figure 3Effects of MgIG on histopathological
variations in mice, as observed by hematoxylin and eosin staining
[(A) magnification, x400; scale bar=50 µm]. The CONT group shows
the standard structure of glomerular capillaries and tubular
epithelium; the ATO group exhibits the inflammatory cell
infiltration in the renal interstitial, glomeruli dilation and
hyperemia; the MgIG treatment group shows a standard structure; and
the H-MgIG and L-MgIG groups alleviate renal morphological
alterations. (B) A boxplot was used to depict the kidney injury
scores, in which the band inside the box represents the median, and
the bottom and top of the box represent the lower and upper
quartiles, respectively. The circle and star represent the largest
and smallest data respectively. Values are demonstrated as median ±
range, n=10. **P<0.01 vs. CONT group;
##P<0.01, #P<0.05 vs. ATO group. MgIG,
magnesium isoglycyrrhizinate; CONT, control group; ATO, arsenic
trioxide group; H-MgIG, high MgIG group (50 mg/kg/day); L-MgIG, low
MgIG group (25 mg/kg/day). |
Effects of MgIG on the expression
levels of antioxidant enzymes
As shown in Figs. 4
and 5, the ATO group showed
markedly decreased the activities of SOD, CAT and GSH in serum and
kidney tissues (P<0.01) and the content of MDA was increased, in
contrast with the CONT group. Compared with the ATO group, the
activities of SOD, CAT and GSH in the groups of MgIG therapy were
enhanced (P<0.05 or P<0.01) and the level of MDA was
decreased (P<0.01). In contrast to the control group, mice that
only received 50 mg/kg MgIG showed no prominent difference in the
serum and kidney tissues expression levels of SOD, MDA, CAT and
GSH. Therefore, MgIG treatment may relieve ATO-induced oxidative
stress.
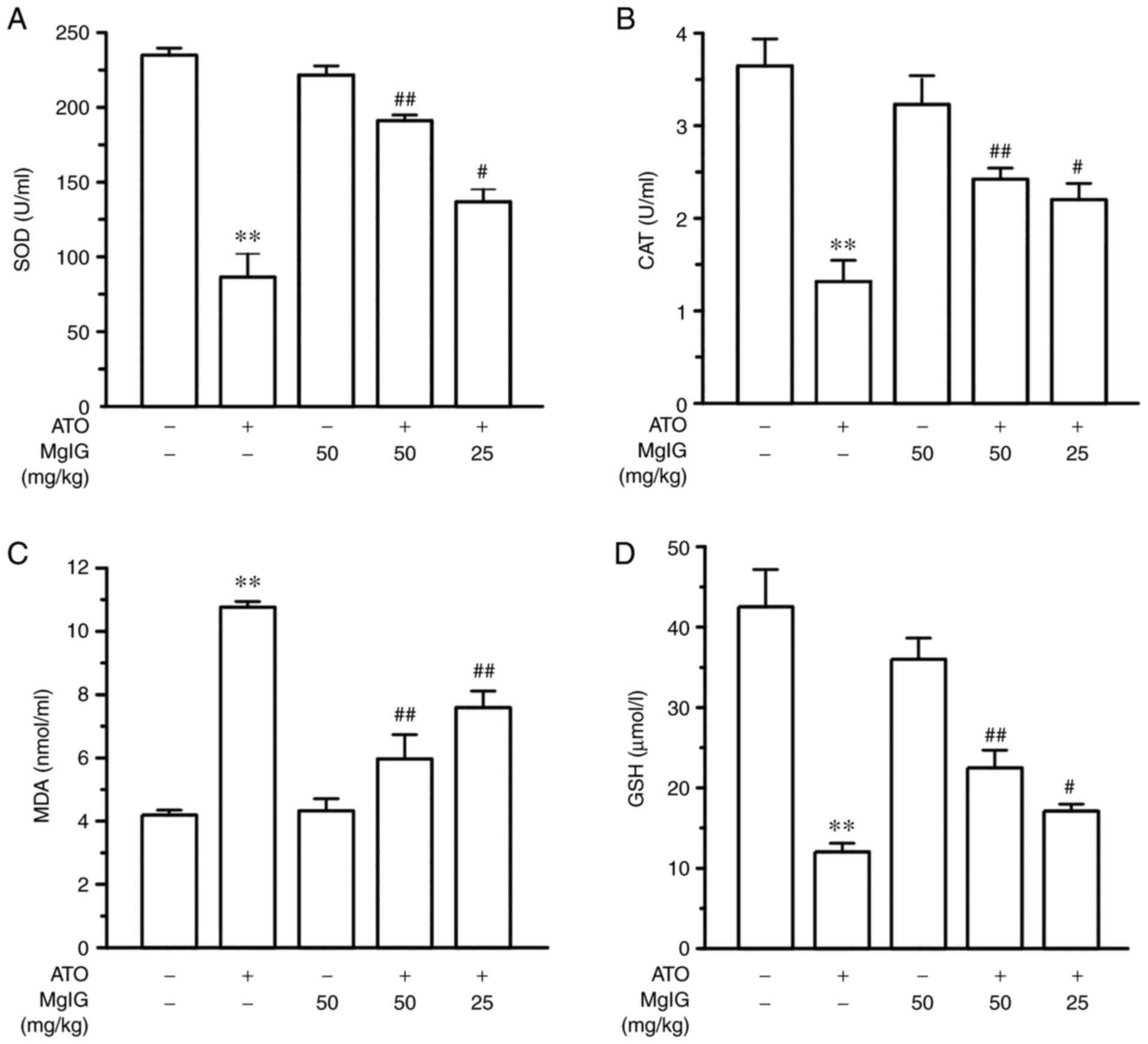 | Figure 4Effects of MgIG on the expression
levels of oxidative stress markers (A) SOD, (B) CAT, (C) MDA and
(D) GSH in serum. Values are demonstrated as mean ± SEM. n=10.
**P<0.01 vs. CONT group; ##P<0.01,
#P<0.05 vs. ATO group. MgIG, magnesium
isoglycyrrhizinate; SOD, superoxide dismutase; CAT, catalase; MDA,
malondialdehyde; GSH, glutathione; CONT, control group; ATO,
arsenic trioxide group. |
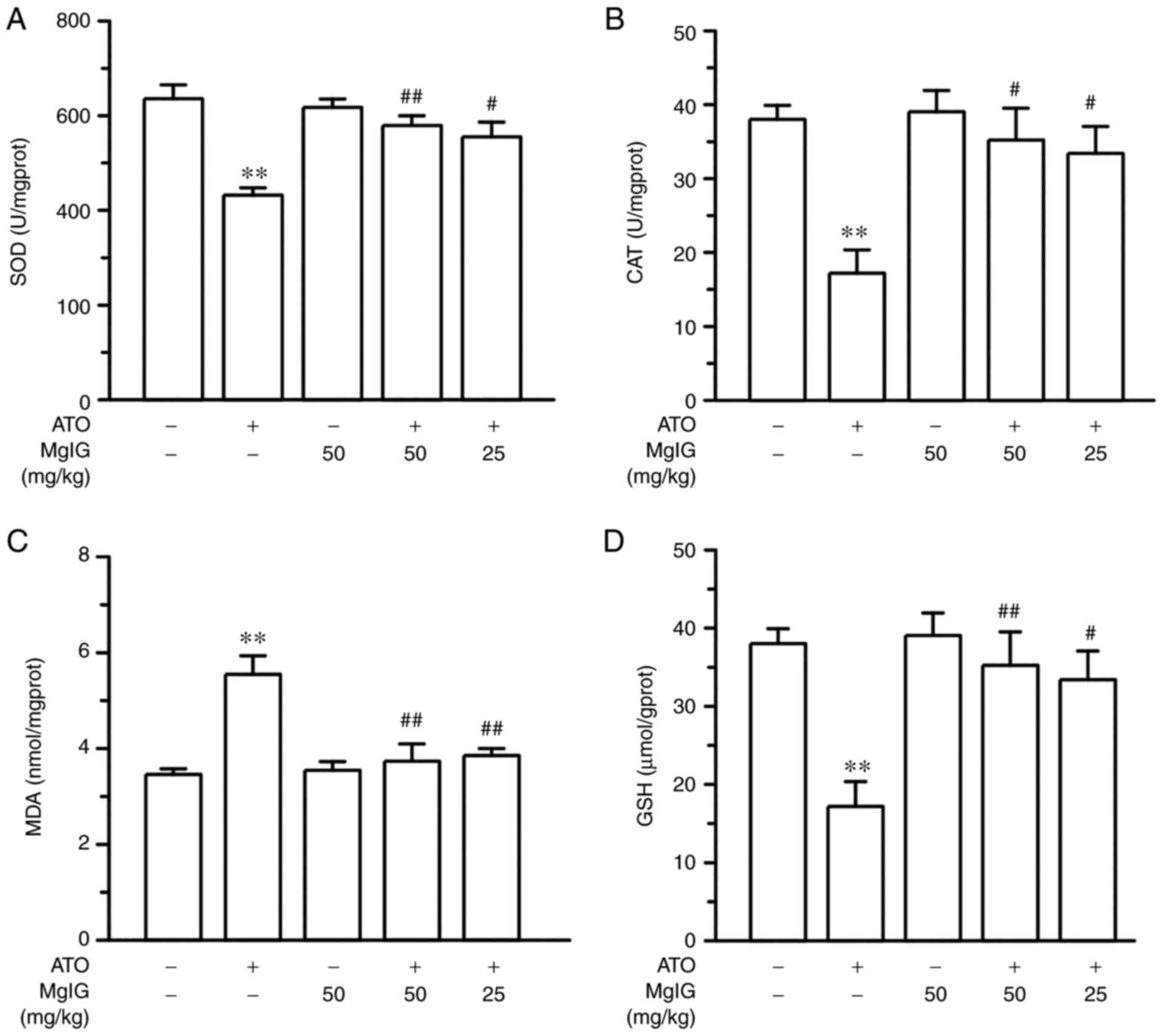 | Figure 5Effects of MgIG on the expression
levels of oxidative stress markers (A) SOD, (B) CAT, (C) MDA and
(D) GSH in renal tissues. Values are demonstrated as mean ± SEM.
n=10. **P<0.01 vs. CONT group;
##P<0.01, #P<0.05 vs. ATO group. MgIG,
magnesium isoglycyrrhizinate; SOD, superoxide dismutase; CAT,
catalase; MDA, malondialdehyde; GSH, glutathione; CONT, control
group; ATO, arsenic trioxide group. |
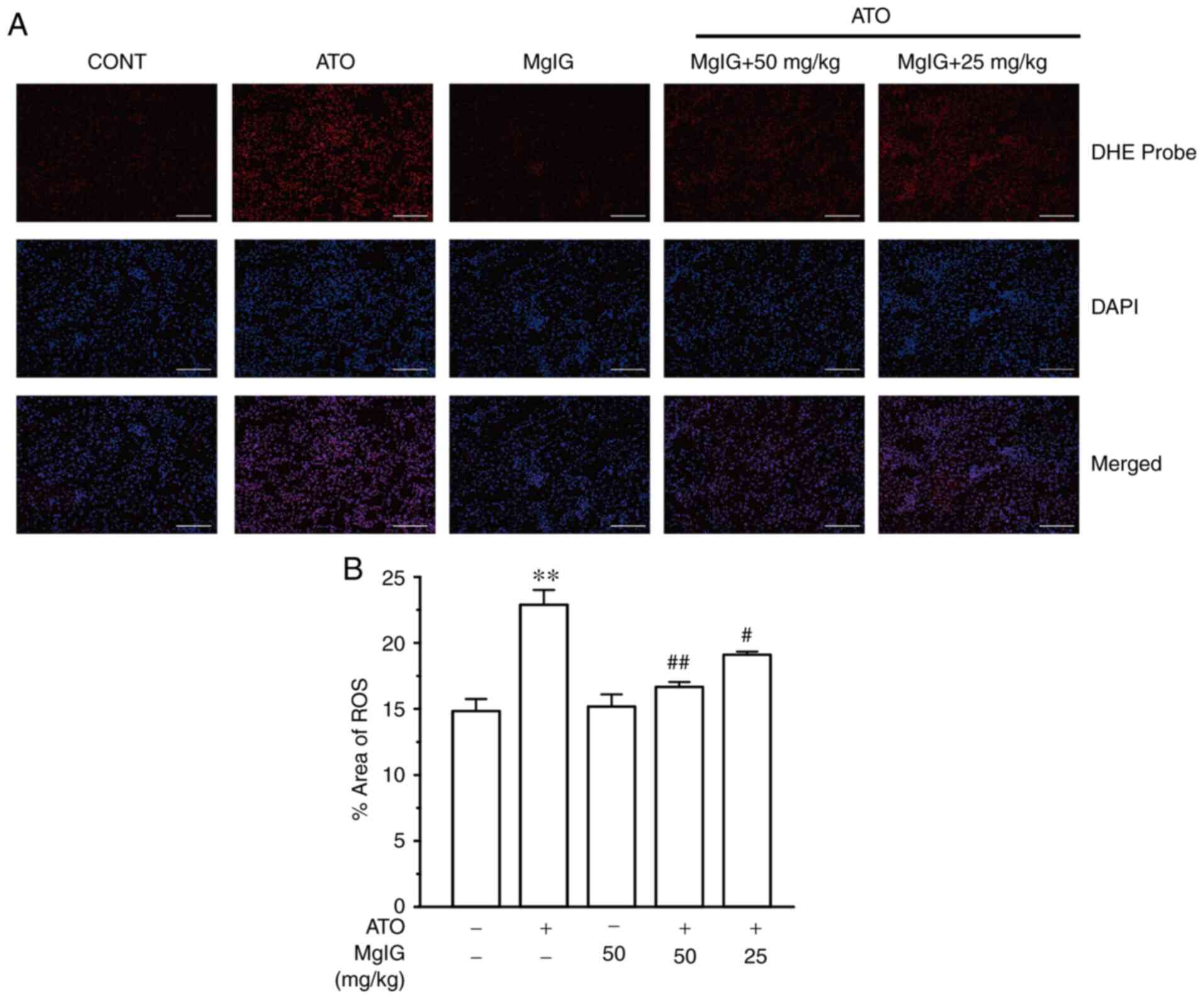
Effects of MgIG on ROS production in
ATO-treated mice
As seen in Fig. 6,
the kidney slices of mice were stained with a fluorescent dye that
identified ROS. The ATO group had substantially greater ROS
generation than the CONT group (P<0.01), whereas no significant
changes were found between the CONT group and the MgIG group. In
contrast to the ATO group, the H-MgIG and L-MgIG groups had lower
ROS generation (P<0.05 or P<0.01). Thus, MgIG treatment may
reduce the ROS production caused by ATO.
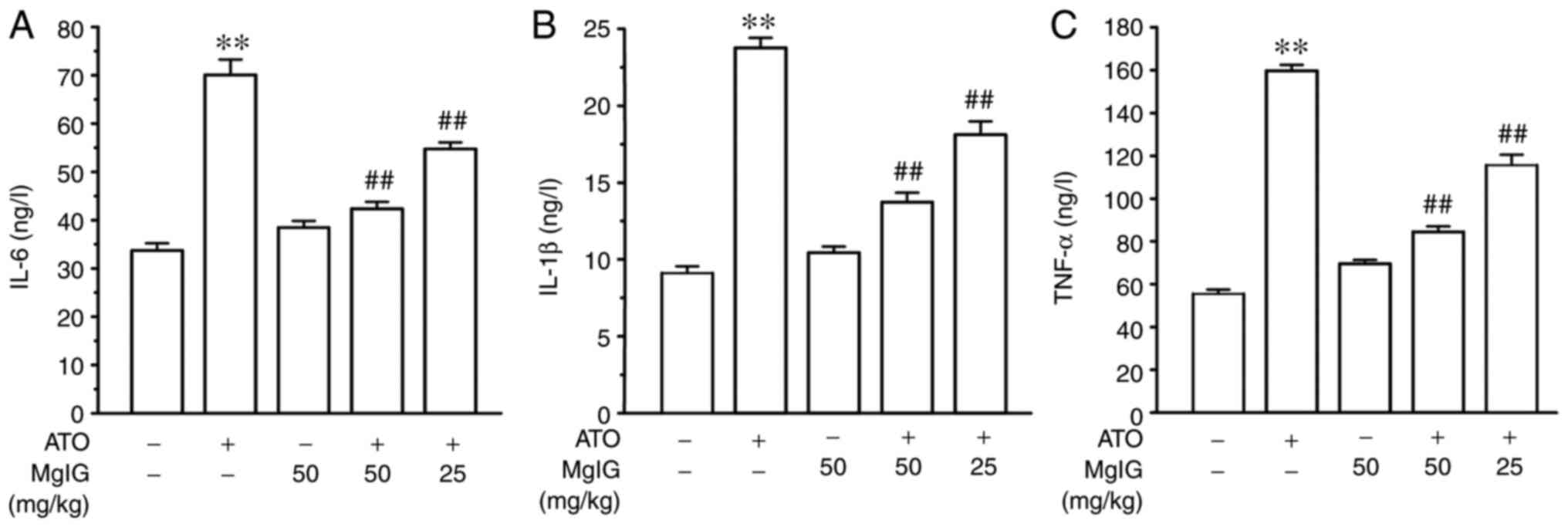
Effects of MgIG on IL-6, IL-1β and
TNF-α expression
Compared with the CONT group, the ATO group showed
increase in IL-6, IL-1β and TNF-α expressions, as shown in Fig. 7 (P<0.01). MgIG alone treated
mice did not show significant differences when compared to the CONT
group. Nevertheless, the levels of IL-6, IL-1β and TNF-α in the
H-MgIG and L-MgIG groups were decreased compared with the ATO
groups (P<0.01). MgIG may have inhibitory effects on ATO-induced
inflammatory reaction.
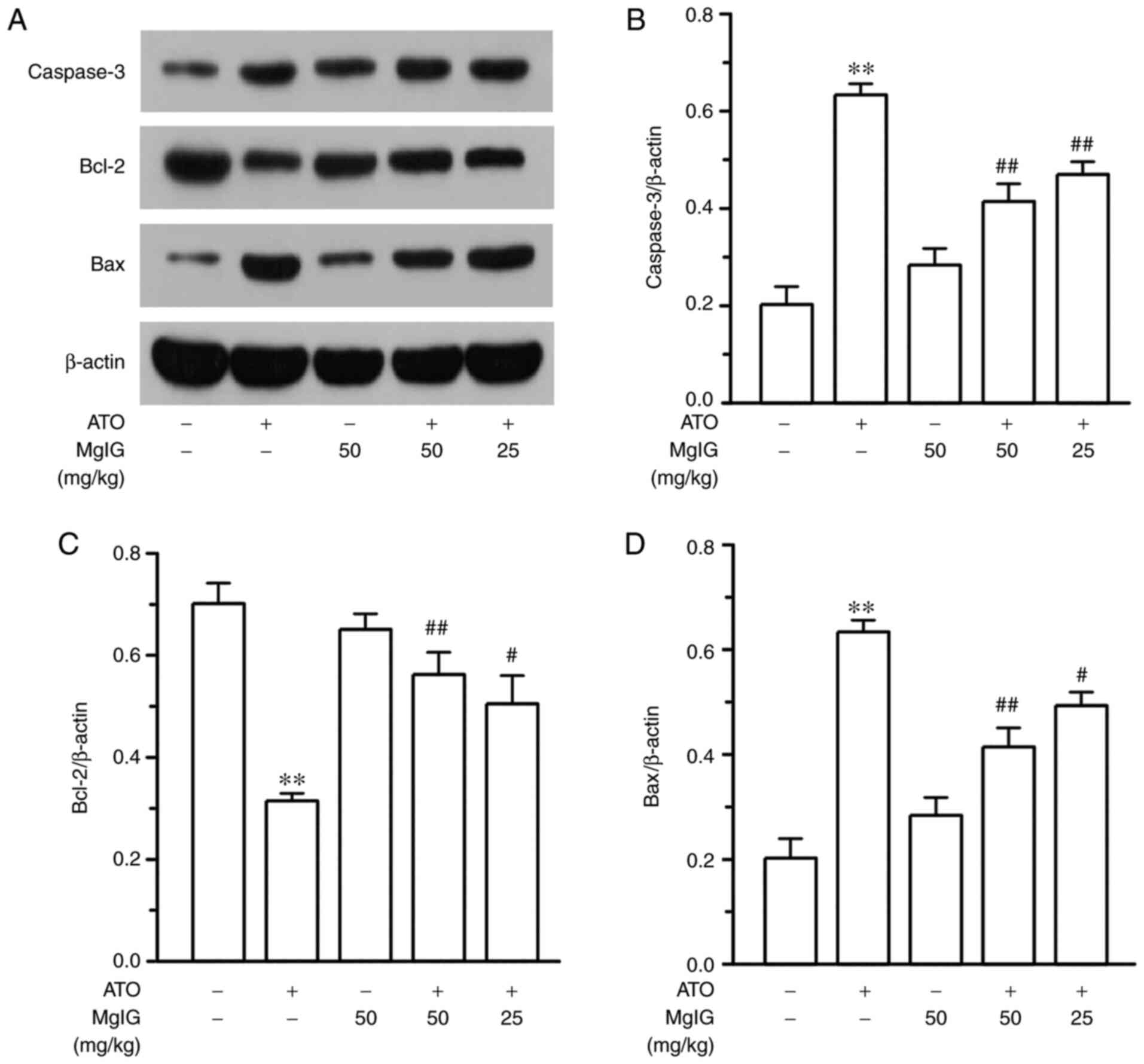
Effects of MgIG on caspase-3, Bcl-2
and Bax expression
In the ATO group, protein expression levels of
caspase-3 and Bax were upregulated, while Bcl-2 protein expression
was downregulated, compared with the CONT group as show in Fig. 8 (P<0.01). However, the
expression levels of caspase-3 and Bax were decreased and Bcl-2
expression level was significantly elevated in the L-MgIG group and
H-MgIG group compared with the ATO group (P<0.05 or P<0.01).
The expression levels of apoptotic proteins in MgIG alone treated
mice did not show significant differences when compared to the CONT
group. Thus, MgIG may alleviate the increase in the expression of
pro-apoptosis indicators induced by ATO.
Effects of MgIG on TLR4/NF-κB
expression
As shown in Fig. 9,
compared with the CONT group, the expressions of TLR4, p-NF-κB,
NF-κB and p-NF-κB/NF-κB in the ATO group were markedly higher
(P<0.01). TLR4, p-NF-κB, NF-κB and p-NF-κB/NF-κB expression
levels in the H-MgIG and L-MgIG groups were lower compared with ATO
group (P<0.05 or P<0.01). The expressions of TLR4, p-NF-κB,
NF-κB and p-NF-κB/NF-κB in the MgIG group and CONT group
demonstrated no significant difference.
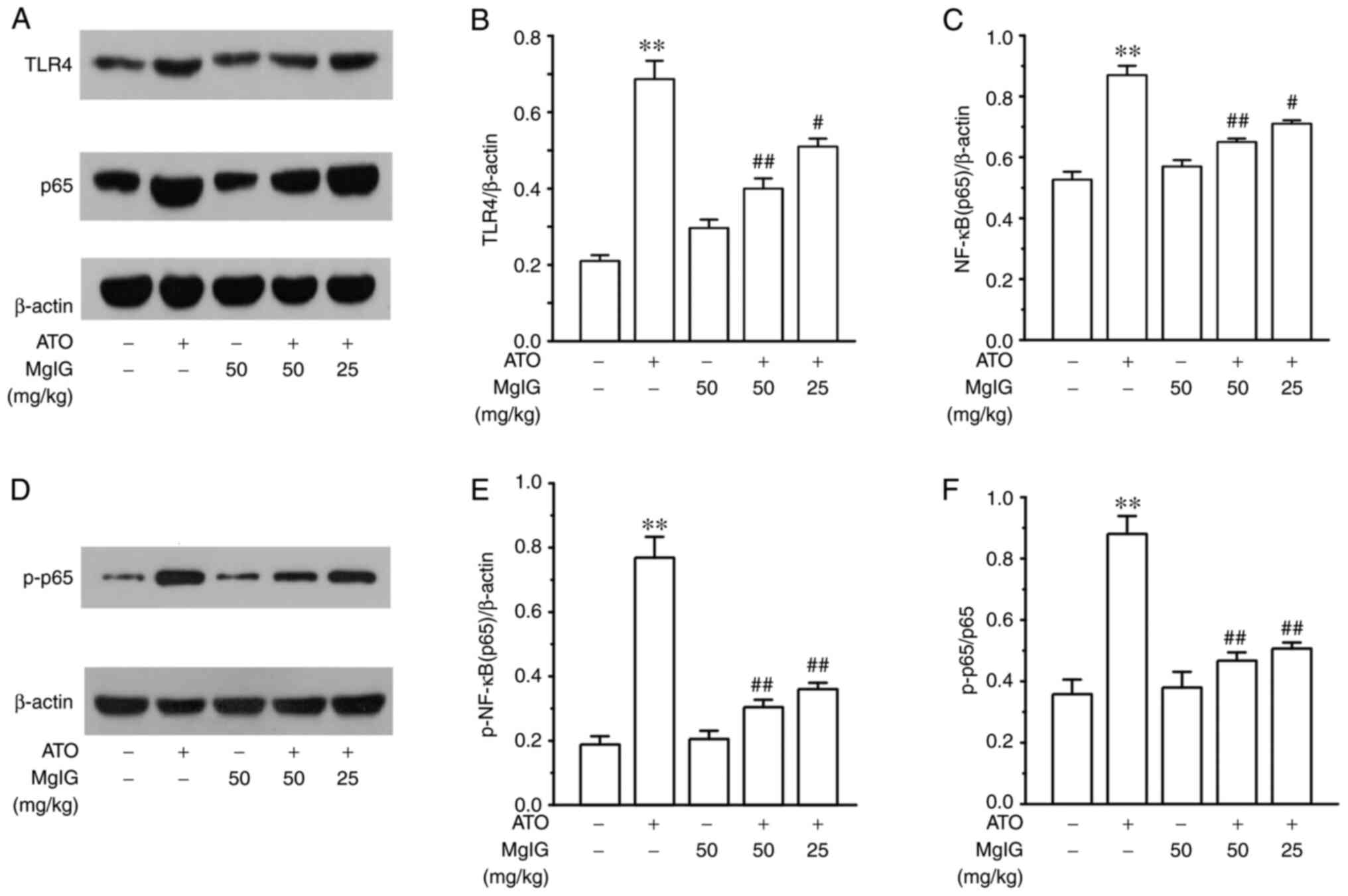 | Figure 9Effects of MgIG on the expression
levels of TLR4, NF-κB (p65) and p-NF-κB (p-p65). (A and D) For each
group, the intensity of (B) TLR4, (C) NF-κB (p65), (E) p-NF-κB
(p-p65) and (F) p-p65/p65 were analyzed by normalizing to β-actin.
Values are demonstrated as mean ± SEM. n=3. **P<0.01
vs. CONT group; ##P<0.01, #P<0.05 vs.
ATO group. MgIG, magnesium isoglycyrrhizinate; TLR4, Toll-like
receptor-4; NF-κB, nuclear factor-κB; p-, phosphorylated; CONT,
control group; ATO, arsenic trioxide group. |
Discussion
Arsenic, a trivalent inorganic arsenic, also is a
worldwide environmental pollutant and a human carcinogen (6,37,38).
As MgIG has good lipophilicity, it easily penetrates the cell
membrane to bind to the receptor protein and the target cell
receptor of steroid hormones, thereby exerting hormonal effects
(such as remission lipid metabolism disorder) (24,39).
The present study demonstrated that MgIG can
ameliorate ATO-induced kidney damage in mice. In addition to
modifying the ATO-mediated increase in BUN and CRE, oxidative
stress and inflammatory cytokines were alleviated following MgIG
treatment. The hematoxylin and eosin-stained kidney tissues of
ATO-treated mice revealed abnormal kidney morphology and the kidney
tissues of medication treatment groups showed slight anomaly, which
indicted MgIG may possess the protective effect on kidney
injury.
The kidney is the main organ of body used for
arsenic excretion and also a principal site for arsenic
accumulation. The present study found that MgIG could improve the
pathological damage of the kidney and mitigate the abnormal
variation of serum BUN, serum CRE, urinary CRE and CRE clearance.
These markers are typically end-products of nitrogenous compounds
and protein metabolism and they are used as biochemical indicators
for detecting renal function (19,40,41).
The considerable restoration of serum BUN, serum CRE, urinary CRE
and CRE clearance activity levels showed that MgIG protected the
kidney from the damage from ATO. However, the content of serum BUN,
serum CRE, urinary CRE and CRE clearance in MgIG alone administered
mice did not change abnormally. In conclusion, treatment of MgIG
could alleviate ATO-induced nephrotoxicity.
Studies report that one of the mechanisms of
nephrotoxicity induced by ATO is the excessive production of
oxidative stress (42). In
addition, its anticancer effects are also associated with abnormal
oxidative stress (43). One of the
ways to mitigate the oxidative stress caused by superfluous ROS is
to augment the synergistic effects of antioxidant enzymes (GSH, SOD
and CAT) (44). This is consistent
with the conclusion of the present study that MgIG enhanced the
activities of antioxidant enzymes and decreased the content of MDA
and the production of ROS. In addition, the activities of GSH, SOD
and CAT and the production of ROS and MDA in the MgIG group showed
no significant change compared with those of the CONT group. As ATO
can stimulate ROS production during the process of metabolic
activation and ROS can devastate the structure of cells, it causes
significant inflammation and ultimately leads to cell apoptosis
(45). Therefore, ATO can further
exacerbate nephrotoxicity by inducing oxidative stress and
inflammation (46). The present
study showed that the ATO group had the highest ROS content, which
generated kidney damage. ROS can also activate all types of
signaling pathways (such as TLR4/NF-κB pathway) (18).
Pro-inflammatory cytokines serve a significant
position in the pathogenesis of various inflammatory illnesses
(47). TNF-α is involved in
cisplatin-induced nephrotoxicity (48). The activation of NF-κB can
stimulate the transcription of IL-6, IL-1β and TNF-α genes, thereby
aggravating the inflammatory response (47,49).
The present study found that the levels of IL-6, IL-1β and TNF-α in
renal tissues were considerably elevated in the ATO group compared
with the CONT group, while treatment with MgIG lessened the
expressions of inflammatory factors, illustrating the
anti-inflammatory effect of MgIG (50,51).
Caspases and Bcl-2 family proteins (such as
Bcl-2-like proteins, Bax-like proteins) are regulators of apoptotic
signaling pathway (52). Bcl-2 is
a membrane protein with anti-apoptotic effect and can restrain the
activation of caspase-3. Bax is a death-promoting molecule and
induces apoptosis (53). In the
present study the ATO group demonstrated increased expression
levels of caspase-3 and Bax and a decreased level of Bcl-2.
However, the expression of apoptotic protein did not significantly
change in the mice treated with MgIG alone. Thus, MgIG treatment
slowed down the ATO-induced apoptosis by restoring the caspase-3,
Bcl-2 and Bax expression to normal levels.
The activation of TLR can initiate the congenital
inflammatory response as TLR4 can bind to a variety of ligands to
trigger an inflammatory response (54,55).
NF-κB is a downstream effector of the TLR4 signaling pathway and
mediates a variety of inflammatory processes (56). In addition, activation of the NF-κB
pathway leads to enhancement of p-p65 and increased expressions of
IL-6, IL-1β and TNF-α, which eventually leads to the generation of
inflammatory reactions in kidney tissue (57). ATO caused a notably increase the
expression of NF-κB in kidney tissue. However, the mice treated
with MgIG did not show a significantly change in the expression of
TLR4, p-p65 and p65. Nevertheless, treatment with MgIG
downregulated the TLR4 and NF-κB expression and inhibited the
production of pro-inflammatory cytokines. Thus, the protective
effect of MgIG in mice with acute renal injury may be due to its
inhibition of TLR4 and NF-κB signaling.
In summary, the present study found that MgIG may
protect against kidney toxicity through its antioxidant,
anti-inflammation and anti-apoptosis properties. The results
demonstrated that MgIG significantly mitigated ATO-induced
nephrotoxicity by decreasing oxidative stress and inflammation,
possibly via restraining the TLR4/NF-κB signaling pathway (Fig. 10). The results indicated that MgIG
could be an effective therapy against ATO-induced nephrotoxicity.
However, in clinical practice, the combination of MgIG with ATO
requires further research.
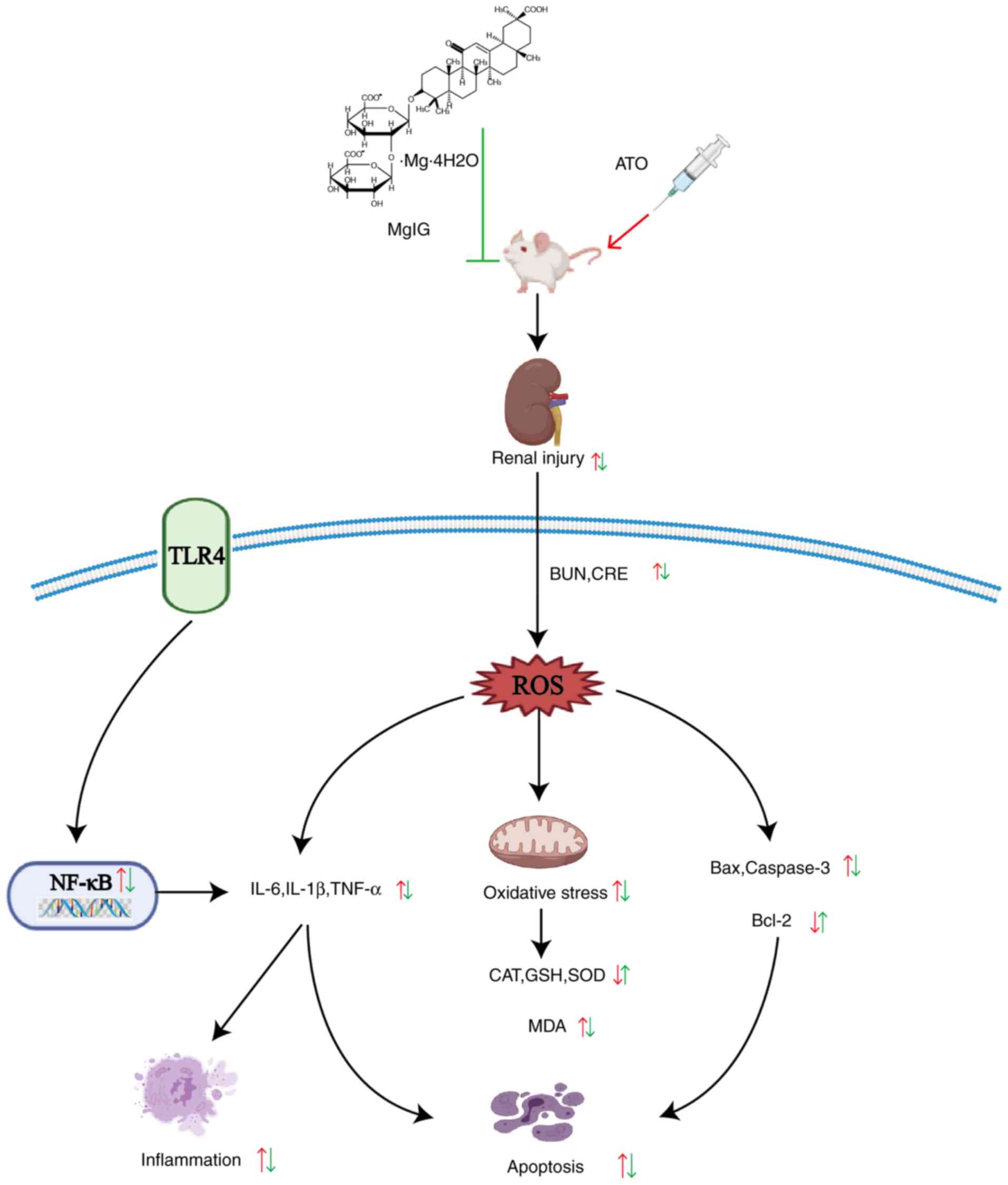 | Figure 10Mechanism of MgIG on ATO-treated
nephrotoxicity. MgIG, magnesium isoglycyrrhizinate; ATO, arsenic
trioxide; TLR4, Toll-like receptor-4; BUN, blood urea nitrogen;
CRE, creatinine; NF-κB, nuclear factor-κB; ROS, reactive oxygen
species; IL, interleukin; TNF, tumor necrosis factor; CAT,
catalase; GSH, glutathione; SOD, superoxide dismutase; MDA,
malondialdehyde; Bax, Bcl-2 associated X; Bcl-2, B-cell lymphoma
2. |
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Research
Foundation of Administration of Traditional Chinese Medicine of
Hebei Province, China (grant nos. 2019135 and 2020188).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZWe, ZWu, XC and SG were involved in the conception
and planning of the current study. ZWe, XS, QH and YZ performed the
experiments. ZWe, QH and XS interpreted the data. ZWe, YZ, YW and
XH were involved in the data analysis. YZ, YW, XH and ZWu provided
guidance for software and figures. ZWe wrote the original draft.
XS, QH, YZ, YW, XH, ZWu, XC and SG reviewed and edited the
manuscript. XH, ZWu and XC supervised the project. SG identified
resources. ZWe and SG confirm the authenticity of all the raw data.
All authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All animal procedures were authorized by Hebei
Medical University of Chinese Medicine's Animal Care and Ethical
Committee (approval no. DWLL2020005).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Watanabe T and Hirano S: Metabolism of
arsenic and its toxicological relevance. Arch Toxicol. 87:969–979.
2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Antman KH: Introduction: The history of
arsenic trioxide in cancer therapy. Oncologist. 6 (Suppl 2):S1–S2.
2001.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bao L and Shi H: Potential molecular
mechanisms for combined toxicity of arsenic and alcohol. J Inorg
Biochem. 104:1229–1233. 2010.PubMed/NCBI View Article : Google Scholar
|
|
4
|
You BR and Park WH: Arsenic trioxide
induces human pulmonary fibroblast cell death via increasing ROS
levels and GSH depletion. Oncol Rep. 28:749–757. 2012.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Simon JP, Parthasarathy M, Nithyanandham
S, Katturaja RK, Namachivayam A and Prince SE: Protective effect of
the ethanolic and methanolic leaf extracts of madhuca longifolia
against diclofenac-induced toxicity in female wistar albino rats.
Pharmacol Rep. 71:983–993. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Katturajan R and Evan Prince S: A role of
connexin 43 on the drug-induced liver, kidney, and gastrointestinal
tract toxicity with associated signaling pathways. Life Sci.
280(119629)2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhang J, Pan X, Li N, Li X, Wang Y, Liu X,
Yin X and Yu Z: Grape seed extract attenuates arsenic-induced
nephrotoxicity in rats. Exp Ther Med. 7:260–266. 2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yu M, Xue J, Li Y, Zhang W, Ma D, Liu L
and Zhang Z: Resveratrol protects against arsenic trioxide-induced
nephrotoxicity by facilitating arsenic metabolism and decreasing
oxidative stress. Arch Toxicol. 87:1025–1035. 2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhang W, Liu Y, Ge M, Jing J, Chen Y,
Jiang H, Yu H, Li N and Zhang Z: Protective effect of resveratrol
on arsenic trioxide-induced nephrotoxicity in rats. Nutr Res Pract.
8:220–226. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Shi H, Shi X and Liu K: Oxidative
mechanism of arsenic toxicity and carcinogenesis. Mol Cell Biochem.
255:67–78. 2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lushchak VI: Free radicals, reactive
oxygen species, oxidative stress and its classification. Chem Biol
Interact. 224:164–175. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Avlas O, Fallach R, Shainberg A, Porat E
and Hochhauser E: Toll-like receptor 4 stimulation initiates an
inflammatory response that decreases cardiomyocyte contractility.
Antioxid Redox Signal. 15:1895–1909. 2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tang J, Xu L, Zeng Y and Gong F: Effect of
gut microbiota on LPS-induced acute lung injury by regulating the
TLR4/NF-kB signaling pathway. Int Immunopharmacol.
91(107272)2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Mir SM, Ravuri HG, Pradhan RK, Narra S,
Kumar JM, Kuncha M, Kanjilal S and Sistla R: Ferulic acid protects
lipopolysaccharide-induced acute kidney injury by suppressing
inflammatory events and upregulating antioxidant defenses in Balb/c
mice. Biomed Pharmacother. 100:304–315. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Feng D, Wang Y, Liu Y, Wu L, Li X, Chen Y,
Chen Y, Chen Y, Xu C, Yang K and Zhou T: DC-SIGN reacts with TLR-4
and regulates inflammatory cytokine expression via NF-κB activation
in renal tubular epithelial cells during acute renal injury. Clin
Exp Immunol. 191:107–115. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhong G, Wan F, Ning Z, Wu S, Jiang X,
Tang Z, Huang R and Hu L: The protective role of autophagy against
arsenic trioxide-induced cytotoxicity and ROS-dependent pyroptosis
in NCTC-1469 cells. J Inorg Biochem. 217(111396)2021.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Farooq MA, Niazi AK, Akhtar J, Saifullah
Farooq M, Souri Z, Karimi N and Rengel Z: Acquiring control: The
evolution of ROS-induced oxidative stress and redox signaling
pathways in plant stress responses. Plant Physiol Biochem.
141:353–369. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Roy A, Manna P and Sil PC: Prophylactic
role of taurine on arsenic mediated oxidative renal dysfunction via
MAPKs/NF-kappaB and mitochondria dependent pathways. Free Radic
Res. 43:995–1007. 2009.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Nandi D, Patra RC and Swarup D: Oxidative
stress indices and plasma biochemical parameters during oral
exposure to arsenic in rats. Food Chem Toxicol. 44:1579–1584.
2006.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang K, Zhao P, Guo G, Guo Y, Tian L, Sun
X, Li S, He Y, Sun Y, Chai H, et al: Arsenic trioxide attenuates
NF-κB and cytokine mRNA levels in the livers of cocks. Biol Trace
Elem Res. 170:432–437. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zheng B, Yang Y, Li J, Li J, Zuo S, Chu X,
Xu S, Ma D and Chu L: Magnesium isoglycyrrhizinate alleviates
arsenic trioxide-induced cardiotoxicity: Contribution of Nrf2 and
TLR4/NF-κB signaling pathway. Drug Des Devel Ther. 15:543–556.
2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yang YZ, Liu ZH, Wang SC, Zhang XQ, Xu HJ,
Yang L and Kong LD: Magnesium isoglycyrrhizinate alleviates
fructose-induced liver oxidative stress and inflammatory injury
through suppressing NOXs. Eur J Pharmacol.
883(173314)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhao Z, Tang Z, Zhang W, Liu J and Li B:
Magnesium isoglycyrrhizinate protects against
renal-ischemia-reperfusion injury in a rat model via
anti-inflammation, anti-oxidation and anti-apoptosis. Mol Med Rep.
16:3627–3633. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zheng J, Wu G, Hu GX, Peng YZ and Xiong
XJ: Protective effects against and potential mechanisms underlying
the effect of magnesium isoglycyrrhizinate in hypoxia-reoxygenation
injury in rat liver cells. Genet Mol Res. 14:15453–15461.
2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ma D, Zhang J, Zhang Y, Zhang X, Han X,
Song T, Zhang Y and Chu L: Inhibition of myocardial hypertrophy by
magnesium isoglycyrrhizinate through the TLR4/NF-κB signaling
pathway in mice. Int Immunopharmacol. 55:237–244. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Liao Y, Tan RZ, Li JC, Liu TT, Zhong X,
Yan Y, Yang JK, Lin X, Fan JM and Wang L: Isoliquiritigenin
attenuates UUO-induced renal inflammation and fibrosis by
inhibiting mincle/Syk/NF-Kappa B signaling pathway. Drug Des Devel
Ther. 14:1455–1468. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Percie du Sert N, Ahluwalia A, Alam S,
Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U,
Emerson M, et al: Reporting animal research: Explanation and
elaboration for the ARRIVE guidelines 2.0. PLoS Biol.
18(e3000411)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liu M, Zheng B, Liu P, Zhang J, Chu X,
Dong C, Shi J, Liang Y, Chu L, Liu Y and Han X: Exploration of the
hepatoprotective effect and mechanism of magnesium
isoglycyrrhizinate in mice with arsenic trioxide-induced acute
liver injury. Mol Med Rep. 23(438)2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Li SG, Xu SZ, Niu Q, Ding YS, Pang LJ, Ma
RL, Jing MX, Wang K, Ma XM, Feng GL, et al: Lutein alleviates
arsenic-induced reproductive toxicity in male mice via Nrf2
signaling. Hum Exp Toxicol. 35:491–500. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Jiang W, Chen Q, Li P, Lu Q, Pei X, Sun Y,
Wang G and Hao K: Magnesium Isoglycyrrhizinate attenuates
lipopolysaccharide-induced depressive-like behavior in mice. Biomed
Pharmacother. 86:177–184. 2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Jiang W, Guo H, Su D, Xu H, Gu H and Hao
K: Ameliorative effect of magnesium isoglycyrrhizinate on hepatic
encephalopathy by epirubicin. Int Immunopharmacol.
75(105774)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Laferriere CA, Leung VS and Pang DS:
Evaluating intrahepatic and intraperitoneal sodium pentobarbital or
ethanol for mouse euthanasia. J Am Assoc Lab Anim Sci. 59:264–268.
2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kim HY, Yokozawa T, Nakagawa T and Sasaki
S: Protective effect of gamma-aminobutyric acid against
glycerol-induced acute renal failure in rats. Food Chem Toxicol.
42:2009–2014. 2004.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Tonomura Y, Yamamoto E, Kondo C, Itoh A,
Tsuchiya N, Uehara T and Baba T: Amphotericin B-induced
nephrotoxicity: Characterization of blood and urinary biochemistry
and renal morphology in mice. Hum Exp Toxicol. 28:293–300.
2009.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Elliott AC and Hynan LS: A SAS(®) macro
implementation of a multiple comparison post hoc test for a
Kruskal-Wallis analysis. Comput Methods Programs Biomed. 102:75–80.
2011.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Yazdani M: Concerns in the application of
fluorescent probes DCDHF-DA, DHR 123 and DHE to measure reactive
oxygen species in vitro. Toxicol In Vitro. 30:578–582.
2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ferreccio C, Smith AH, Durán V, Barlaro T,
Benítez H, Valdés R, Aguirre JJ, Moore LE, Acevedo J, Vásquez MI,
et al: Case-control study of arsenic in drinking water and kidney
cancer in uniquely exposed Northern Chile. Am J Epidemiol.
178:813–818. 2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Surdu S, Fitzgerald EF, Bloom MS, Boscoe
FP, Carpenter DO, Haase RF, Gurzau E, Rudnai P, Koppova K, Févotte
J, et al: Occupational exposure to arsenic and risk of nonmelanoma
skin cancer in a multinational European study. Int J Cancer.
133:2182–2191. 2013.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhao XJ, Yang YZ, Zheng YJ, Wang SC, Gu
HM, Pan Y, Wang SJ, Xu HJ and Kong LD: Magnesium isoglycyrrhizinate
blocks fructose-induced hepatic NF-κB/NLRP3 inflammasome activation
and lipid metabolism disorder. Eur J Pharmacol. 809:141–150.
2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Gora RH, Kerketta P, Baxla SL, Toppo R,
Prasad R, Patra PH and Roy BK: Ameliorative effect of tephrosia
purpurea in arsenic-induced nephrotoxicity in rats. Toxicol Int.
21:78–83. 2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wang X, Zhao H, Shao Y, Wang P, Wei Y,
Zhang W, Jiang J, Chen Y and Zhang Z: Nephroprotective effect of
astaxanthin against trivalent inorganic arsenic-induced renal
injury in wistar rats. Nutr Res Pract. 8:46–53. 2014.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Dutta S, Saha S, Mahalanobish S, Sadhukhan
P and Sil PC: Melatonin attenuates arsenic induced nephropathy via
the regulation of oxidative stress and inflammatory signaling
cascades in mice. Food Chem Toxicol. 118:303–316. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Kumar S, Yedjou CG and Tchounwou PB:
Arsenic trioxide induces oxidative stress, DNA damage, and
mitochondrial pathway of apoptosis in human leukemia (HL-60) cells.
J Exp Clin Cancer Res. 33(42)2014.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Schieber M and Chandel NS: ROS function in
redox signaling and oxidative stress. Curr Biol. 24:R453–R462.
2014.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Wang Y, Zhao H, Shao Y, Liu J, Li J and
Xing M: Copper or/and arsenic induce oxidative stress-cascaded,
nuclear factor kappa B-dependent inflammation and immune imbalance,
trigging heat shock response in the kidney of chicken. Oncotarget.
8:98103–98116. 2017.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Lushchak VI: Classification of oxidative
stress based on its intensity. EXCLI J. 13:922–937. 2014.PubMed/NCBI
|
|
47
|
Kumar P, Sulakhiya K, Barua CC and Mundhe
N: TNF-α, IL-6 and IL-10 expressions, responsible for disparity in
action of curcumin against cisplatin-induced nephrotoxicity in
rats. Mol Cell Biochem. 431:113–122. 2017.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Jiang L, Liu Y, He P, Chen J, Liu S and
Tan N: Geraniin ameliorates cisplatin-induced nephrotoxicity in
mice. Free Radic Res. 50:813–819. 2016.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Abou-Hany HO, Atef H, Said E, Elkashef HA
and Salem HA: Crocin mediated amelioration of oxidative burden and
inflammatory cascade suppresses diabetic nephropathy progression in
diabetic rats. Chem Biol Interact. 284:90–100. 2018.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Xie C, Li X, Wu J, Liang Z, Deng F, Xie W,
Zhu M, Zhu J, Zhu W, Geng S and Zhong C: Anti-inflammatory activity
of magnesium isoglycyrrhizinate through inhibition of phospholipase
A2/Arachidonic acid pathway. Inflammation. 38:1639–1648.
2015.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Lin G, Nnane IP and Cheng TY: The effects
of pretreatment with glycyrrhizin and glycyrrhetinic acid on the
retrorsine-induced hepatotoxicity in rats. Toxicon. 37:1259–1270.
1999.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Wang YL, Wang CY, Zhang BJ and Zhang ZZ:
Shenfu injection suppresses apoptosis by regulation of Bcl-2 and
caspase-3 during hypoxia/reoxygenation in neonatal rat
cardiomyocytes in vitro. Mol Biol Rep. 36:365–370. 2009.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Pratheeshkumar P and Kuttan G: Oleanolic
acid induces apoptosis by modulating p53, Bax, Bcl-2 and caspase-3
gene expression and regulates the activation of transcription
factors and cytokine profile in B16F. J Environ Pathol Toxicol
Oncol. 30:21–31. 2011.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Lagos D, Vart RJ, Gratrix F, Westrop SJ,
Emuss V, Wong PP, Robey R, Imami N, Bower M, Gotch F and Boshoff C:
Toll-like receptor 4 mediates innate immunity to Kaposi sarcoma
herpesvirus. Cell Host Microbe. 4:470–483. 2008.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Du Y, Qian B, Gao L, Tan P, Chen H, Wang
A, Zheng T, Pu S, Xia X and Fu W: Aloin preconditioning attenuates
hepatic ischemia/reperfusion injury via inhibiting TLR4/MyD88/NF-κB
signal pathway in vivo and in vitro. Oxid Med Cell Longev.
2019(3765898)2019.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Elsharkawy AM and Mann DA: Nuclear
factor-kappaB and the hepatic inflammation-fibrosis-cancer axis.
Hepatology. 46:590–597. 2007.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Song J, Fan H, Li H, Ding H, Lv Q and Hou
S: Zingerone ameliorates lipopolysaccharide-induced acute kidney
injury by inhibiting Toll-like receptor 4 signaling pathway. Eur J
Pharmacol. 772:108–114. 2016.PubMed/NCBI View Article : Google Scholar
|