Introduction
Cadmium (Cd) is one of the global pollutants of the
environment (1). Tobacco smoke,
Cd-contaminated food and industrial contamination are the main
sources of Cd toxicity in humans (2,3).
Chronic exposure to Cd has a major effect on humans, causing damage
to multiple organs including kidneys, liver, lung, pancreas and
testes (3-5).
In addition, studies have reported that the vascular endothelial
cell (VEC) is another main target of Cd (6,7).
Accumulated Cd impairs endothelial function at a variety of
molecular levels, including cell adhesion molecules, metal ion
transporters and protein kinase signaling pathways (7,8). As
blood vessels are widely distributed in organs, functional damage
of VECs further causes toxicity in the parenchymal cells of human
organs (8,9). Therefore, it is of great significance
to investigate the underlying mechanism of Cd toxicity on VECs.
Vascular endothelial (VE) cadherin, a principal
adhesion protein, is composed of a large ectodomain with five
cadherin repeats, a transmembrane domain and a cytoplasmic tail
(10). Cell-to-cell adhesion is
established by interactions of ectodomain of VE-cadherin (11). The cytoplasmic tail of VE-cadherin
is regulated by a range of phosphatases and kinases (12). Phosphorylation of the VE-cadherin
cytoplasmic tail leads to VE-cadherin dissociation from the
junctions (11). VE-cadherin is
produced by various stimuli to regulate vascular permeability
(13-16).
It has been reported that VE-cadherin mRNA expression is
reduced in chick embryo model treated with 50 µM Cd (17). In addition, low concentration of Cd
induces membrane dissociation of VE-cadherin in human renal
glomerular endothelial cells and human umbilical vein endothelial
cells (HUVECs) (18,19). Further research is required on the
effect of Cd on VE-cadherin.
Rho is a member of guanosine triphosphatase (GTPase)
family (20). Rho-associated
coiled-coil kinase (ROCK), including ROCK1 and ROCK2, is a
serine/threonine kinase downstream of Rho GTPases (20). ROCK pathway regulates various
cellular functions, including contraction, cytoskeleton
organization, cell-to-cell adhesion and permeability (21). Lipopolysaccharide has been reported
to activate the RhoA/ROCK signaling pathway to weaken cell
junctions by reducing the expression of VE-cadherin and altering
distribution of VE-cadherin (22).
However, whether Cd regulates VE-cadherin by ROCK pathway is
uncertain.
In the present study, the results showed that Cd
increased VE-cadherin expression in HUVECs in a dose-dependent
manner. It also found that 10 µM Cd inhibited ROCK pathway.
Narciclasine, an activator of ROCK pathway, reversed Cd-induced
VE-cadherin expression. In addition, the expression of VE-cadherin
is unchanged in HUVECs treated with 10 µM narciclasine in the
absence or presence of 10 µM CdCl2. ROCK pathway
inhibitor Y27632 increased VE-cadherin expression in HUVECs in a
dose-dependent manner. With pretreatment of 20 µM Y27632, 10 µM Cd
did not alter VE-cadherin expression. The present study
demonstrated a role for the ROCK pathway in regulating the
VE-cadherin expression induced by Cd in HUVECs.
Materials and methods
Reagents and antibodies
CdCl2 was purchased from Millipore Sigma
and dissolved in phosphate buffered saline (PBS). Narciclasine
(MedChem Express) and Y27632 (Selleck Chemicals) were dissolved in
dimethyl sulfoxide (DMSO). The primary antibody against VE-cadherin
was purchased from Abcam. The primary antibodies against myosin
phosphatase-targeting subunit (MYPT), phosphorylated (p)-MYPT
(Ser507) and GAPDH were purchased from Cell Signaling Technology,
Inc. The secondary antibody was goat anti-rabbit IgG (cat. no.
7074; Cell Signaling Technology, Inc.).
Cell Culture
HUVECs were purchased from American Type Culture
Collection and maintained in Dulbecco's modified Eagle's medium
(DMEM) supplemented with 10% fetal bovine serum (FBS; Lonza Group
Ltd.), 10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin at
37˚C in 5% CO2.
Reverse transcription-quantitative
(RT-q) PCR
HUVECs were seeded at a density of 5x105
cells/ml in a 65-mm dish with different treatment. RNA extraction,
cDNA synthesis and qPCR were performed according to the
manufacturer's protocols. Total RNA from the treated HUVECs was
isolated with the E.Z.N.A. Total RNA kit II (Omega Bio-Tek, Inc.).
Complementary cDNA was synthesized using the RevertAid First strand
cDNA Synthesis kit (Thermo Fisher Scientific, Inc.). Diluted cDNA
(4.6 µl; 50 ngcDNA) and 5.4 µl of primer and supermix mixture (SYBR
premix Ex Tap™ II and dH2O; Takara Biotechnology Co.,
Ltd.) were used in each RT-qPCR reaction. The RT-qPCR process was
performed on the CFX96 Real-Time System (Bio-Rad Laboratories,
Inc.). Reaction conditions were: 95˚C for 5 min, 40 cycles of 95˚C
for 10 sec and 60˚C for 32 sec. All PCR reactions were repeated
three times and the mRNA levels were normalized to β-actin.
Relative quantitative values were obtained from cycle threshold
(Ct) and the 2-∆∆Cq method (23). The human VE-cadherin PCR primers
were 5'-CAGCCCAAAGTGTGTGAGAA-3' (sense) and
5'-CGGTCAAACTGCCCATACTT-3' (antisense). The human β-actin PCR
primers were 5'-TTGCCGACAGGATGCAGAA-3' (sense) and
5'-GCCGATCCACACGGAGTACT-3' (antisense).
Western blotting
Western blotting was performed as previously
described (24). Cells were
homogenized with in RIPA buffer at 4˚C. Protein concentrations were
measured using Pierce BCA Assay kit (Thermo Fisher Scientific,
Inc.). Supernatants of cell lysates were mixed with loading buffer
and heated at 95˚C for 5 min. Samples containing 25 µg protein was
separated by 10% SDS-PAGE and transferred to 0.45 µm polyvinylidene
difluoride (PVDF) membrane. Membranes were blocked with 5% non-fat
milk for 2 hat room temperature and incubated with primary antibody
against VE-cadherin (1:1,000; cat. no. ab33168; Abcam), MYPT
(1:1,000; cat. no. 2634; Cell Signaling Technology, Inc.),
phosphorylated (p)-MYPT (1:1,000; cat. no. 3040; Cell Signaling
Technology, Inc.) and GAPDH (1:3,000, cat. no. 2118; Cell Signaling
Technology, Inc.) at 4˚C overnight. After washing with TBS-T (0.5%
Tween), the membranes were incubated with secondary antibodies for
2 h at room temperature and then washed with TBS-T. The secondary
antibody was HRP-linked goat anti-rabbit IgG antibody (1:8,000;
cat. no. 7074; Cell Signaling Technology, Inc.). Following the
manufacturer's instructions, specific binding was revealed by an ECL
kit (Pierce; Thermo Fisher Scientific, Inc.). Densitometry analysis
was performed with ImageJ software 1.48 (National Institute
Health).
Immunofluorescence
HUVECs were grown into monolayer on
fibronectin-coated glass chamber slides and were then treated with
10 µM CdCl2 for 12 h. Then, cells were washed with PBS
and fixed with 4% paraformaldehyde for 15 min. After washing three
times with PBS for 10 min, the cells were stained with a primary
antibody against human VE-cadherin (cat. no. a33168; Abcam) at a
dilution of 1:500 overnight at 4˚C and were incubated with the
Alexa Fluor 546 donkey anti-rabbit secondary antibody (1:200; cat.
no. A10040; Thermo Fisher Scientific, Inc.) for 2 h at room
temperature. After washing three times with PBS for 10 min at room
temperature, samples were imaged using an Olympus BX51 fluorescence
microscope (Olympus Corporation) with an excitation wavelength of
546 nm (magnification, x200). The chamber slide with the monolayer
was divided in 16 equal areas (4x4) and one field was randomly
chosen in each area. The image mostly close to the average staining
intensity was chosen as representative image.
Statistical data analysis
All the data were analyzed using GraphPad Prism 5.0
(GraphPad Software, Inc.). Data are presented as means ± SD.
Statistical significance was assessed using one-way analysis of
variance followed by Tukey's post hoc test or Student's t-test. A
statistical difference of P<0.05 was considered significant.
Results
High dose Cd increases VE-Cadherin
expression
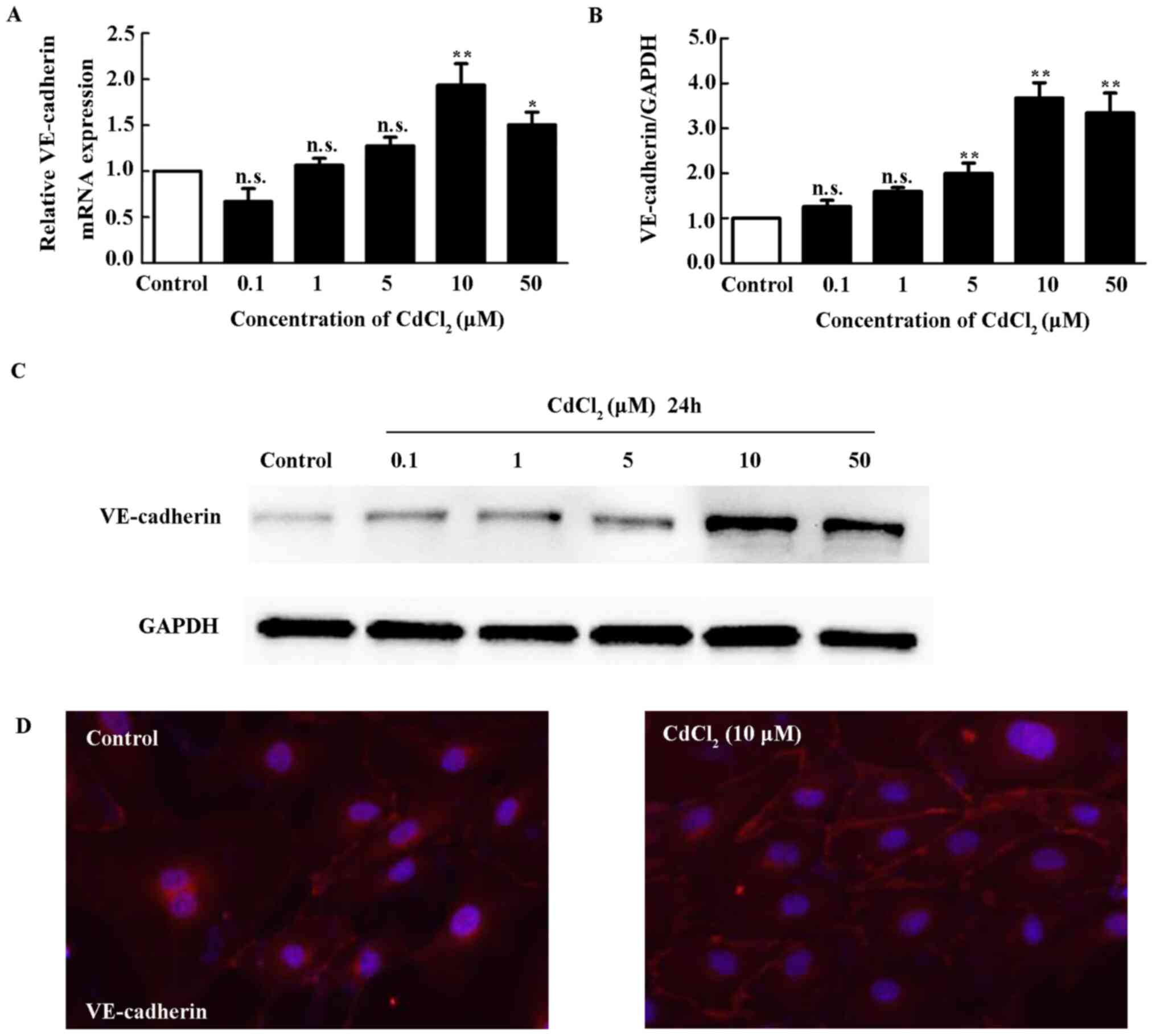
The present study analyzed the mRNA and protein
expression of VE-cadherin in HUVECs treated with different
concentrations of Cd for 12 h. Fig.
1A showed that relative VE-cadherin mRNA expression was
not changed at concentrations of 0.1, 1 and 5 µM of Cd. However, it
was significantly upregulated by 10 and 50 µM Cd. Western blotting
showed that Cd increases VE-cadherin protein expression following
treatment with Cd at concentrations of 1, 5, 10 and 50 µM (Fig. 1B and C). A previous study showed that Cd
disrupts VE-cadherin mediated cell-to-cell adhesion of HUVECs
(25). Immunofluorescent staining
with VE-cadherin antibody on HUVECs treated with Cd was performed.
As shown in Fig. 1D, VE-cadherin is
mainly distributed in cytoplasm membrane under normal condition.
After treatment with 10 µM Cd, the fluorescence became stronger,
suggesting that higher levels of VE-cadherin were detected at
cell-to-cell junctions between cells.
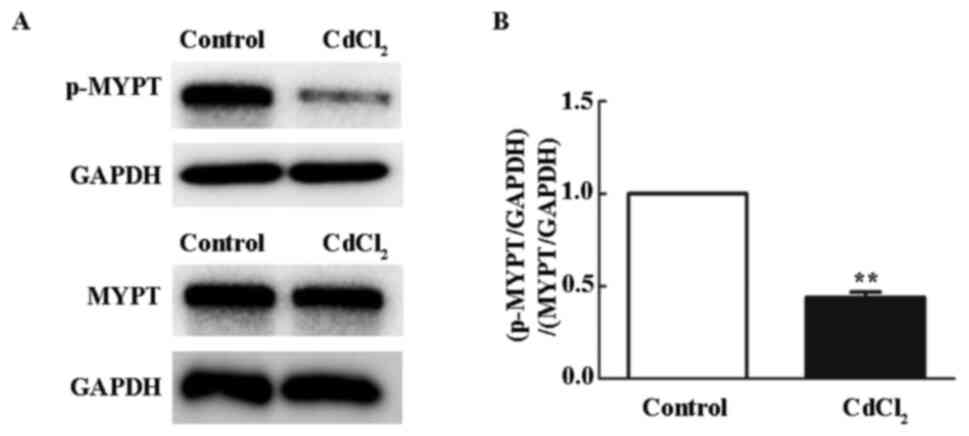
Cd inhibits ROCK activity in
HUVECs
To examine whether Cd affected ROCK activity, the
phosphorylation levels of ROCK downstream substrate, myosin-binding
subunit of myosin phosphatase (MYPT), were evaluated by western
blotting. HUVECs treated with 10 µM Cd showed a significant
decrease in p-MYPT after 12 h (Fig.
2A and B), suggesting that Cd
inhibited the Rho/ROCK pathway.
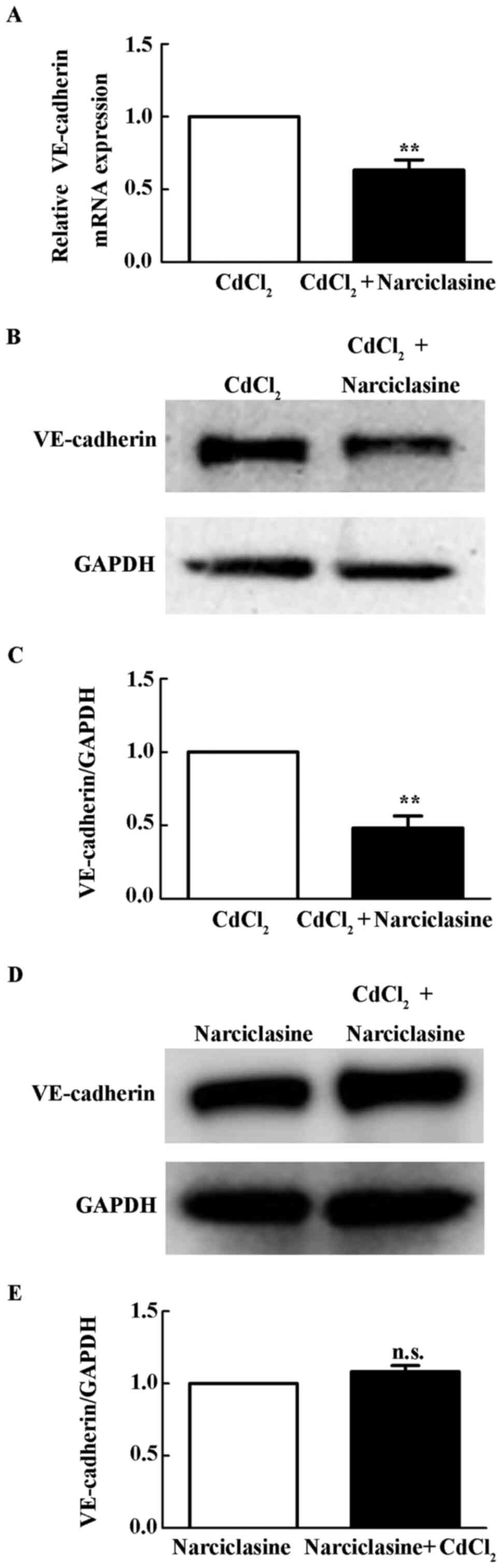
ROCK activation reduces Cd-induced
increase of VE-cadherin expression
Narciclasine is an activator of the Rho/ROCK pathway
(26). The present study examined
whether narciclasine inhibited Cd-induced VE-cadherin expression.
As shown in Fig. 3A-C, 10 µM
narciclasine reduced mRNA and protein level of VE-cadherin upon Cd
treatment (P<0.01). With pretreatment of 10 µM narciclasine, 10
µM Cd did not increases VE-cadherin protein in HUVECs (Fig. 3D), suggesting Cd induced VE-cadherin
expression through inhibition of ROCK signaling.
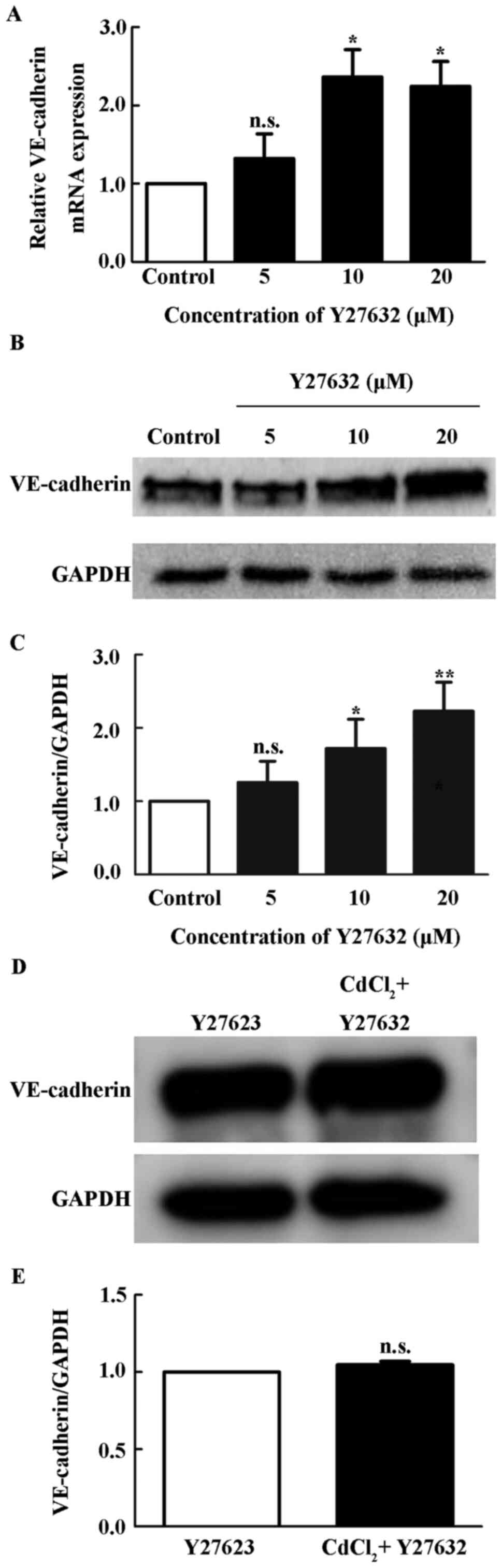
Inhibition of ROCK upregulates the
expression of VE-cadherin
To examine the effect of ROCK pathway on
VE-cadherin, HUVECs were treated with different concentrations of
ROCK inhibitors Y27632 for 12 h. Y27632 increased the expression of
VE-cadherin mRNA at 10 and 20 µM (P<0.05; Fig. 4A). Y27632 also increased the protein
levels of VE-cadherin (P<0.05; Fig.
4B and C), suggesting that ROCK
pathway negatively regulated VE-cadherin expression. With
pretreatment of 10 µM Y27632, Cd did not significantly alter the
level of VE-cadherin in HUVECs (Fig.
4D).
Discussion
Cd exposure has been reported to cause dysfunction
of VECs (8,27). Depending on the dose of exposure, Cd
differentially affects vascular VECs, including permeability,
apoptosis and proliferation (7,28,29).
The present study demonstrated that Cd upregulated expression of
VE-cadherin via inhibition of ROCK activities.
The regulation of signaling pathways in response to
Cd toxicity is dependent on Cd concentration (30). A previous study demonstrated that
low-dose Cd (4 µM) impairs adherens junctions by inducing
VE-cadherin and β-catenin redistribution, causing hyperpermeability
in HUVEC monolayers (19,31). In the present study, Cd increased
VE-cadherin expression in HUVECs in a concentration-dependent
manner. The effect of Cd (10 µM) on VE-cadherin was the more
remarked than other concentration. In a previous study, treatment
of HUVECs with Cd reduces VE-cadherin localization to cell
junctions in a concentration-dependent manner (32). Similarly, in the present study,
lower levels of VE-cadherin were noted at cell-to-cell junctions
between cells following 10 µM Cd treatment. However, Cd (10 µM) has
been reported to induce VEC hyperpermeability, suggesting that the
increased expression of VE-cadherin induced by 10 µM Cd fails to
rescue vascular hyperpermeability (33). One reason may be that apoptosis and
senescence of VECs during 10 µM Cd exposure leads to enhanced
vessel wall permeability to cytokines, growth factors, lipids and
immune cells (34).
The present study demonstrated that 10 µM of Cd
inhibited ROCK activity. The ROCK pathway increases vascular
permeability by causing junction protein remodeling and endothelial
barrier dysfunction (35,36). ROCK inhibits the expression of tight
junction components, including occludin and claudin-1 (37,38).
The present study found that ROCK also negatively regulated the
expression of VE-cadherin in HUVECs, In addition, Cd did not
increase the expression of VE-cadherin in the presence of ROCK
inhibitor Y27632, suggesting that ROCK mediated Cd-induced
VE-cadherin expression. The results of the present study are
consistent with previous studies. For example, the ROCK pathway
inhibitor partially limits the increased monolayer permeability in
lethal toxin-treated VECs through restoration of VE-cadherin
expression and membrane localization (39). Inhibition of ROCK decreases the
tension across VE-cadherin adhesion and VE-cadherin dissociation
rate, resulting in accumulation of VE-cadherin in adherens
junctions (40). FPND, a ROCK
inhibitor, protects vascular integrity through cytoskeletal
rearrangement and enhancement of cell-to-cell junctions in VECs via
the ROCK1 and VE-cadherin signaling pathways (41).
In conclusion, the results of the present study
suggested that ROCK inhibition contributes to Cd-induced expression
of VE-cadherin in endothelial cells. It increases our understanding
of Cd-induced vascular dysfunction.
Acknowledgements
The authors thank Dr Jing Liu of Shandong Provincial
Qianfoshan Hospital (Jinan, China) for scientific discussion and
technical support.
Funding
Funding: This study was supported by a grant from Science and
Technology Development Plan of Shandong Province (grant no.
2013GSF11805).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MG and HW conceived and designed the experiments.
XiaoruiL, XiaoL and RS performed the experiments. XiaoruiL, XiaoL
and RS analyzed the data. XiaoruiL, MG and HW wrote the paper.
XiaoruiL and MG confirm the authenticity of all the raw data. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Apykhtina OL, Dybkova SM, Sokurenko LM and
Chaikovsky YB: Cytotoxic and genotoxic effects of cadmium sulfide
nanoparticles. Exp Oncol. 40:194–199. 2018.PubMed/NCBI
|
|
2
|
Surolia R, Karki S, Kim H, Yu Z, Kulkarni
T, Mirov SB, Carter AB, Rowe SM, Matalon S, Thannickal VJ, et al:
Heme oxygenase-1-mediated autophagy protects against pulmonary
endothelial cell death and development of emphysema in
cadmium-treated mice. Am J Physiol Lung Cell Mol Physiol.
309:L280–l292. 2015.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Angeli JK, Cruz Pereira CA, de Oliveira
Faria T, Stefanon I, Padilha AS and Vassallo DV: Cadmium exposure
induces vascular injury due to endothelial oxidative stress: The
role of local angiotensin II and COX-2. Free Radic Biol Med.
65:838–848. 2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Chen H, Lu Y, Cao Z, Ma Q, Pi H, Fang Y,
Yu Z, Hu H and Zhou Z: Cadmium induces NLRP3 inflammasome-dependent
pyroptosis in vascular endothelial cells. Toxicol Lett. 246:7–16.
2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Chen X, Li J, Cheng Z, Xu Y, Wang X, Li X,
Xu D, Kapron CM and Liu J: Low dose cadmium inhibits proliferation
of human renal mesangial cells via activation of the JNK pathway.
Int J Environ Res Public Health. 13(990)2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Liu F, Wang B, Li L, Dong F, Chen X, Li Y,
Dong X, Wada Y, Kapron CM and Liu J: Low-dose cadmium upregulates
VEGF expression in lung adenocarcinoma cells. Int J Environ Res
Public Health. 12:10508–10521. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Prozialeck WC, Edwards JR and Woods JM:
The vascular endothelium as a target of cadmium toxicity. Life Sci.
79:1493–1506. 2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Nagarajan S, Rajendran S, Saran U, Priya
MK, Swaminathan A, Siamwala JH, Sinha S, Veeriah V, Sonar P, Jadhav
V, et al: Nitric oxide protects endothelium from cadmium mediated
leakiness. Cell Biol Int. 37:495–506. 2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Fujiwara Y, Yamamoto C, Yoshida E, Kumagai
Y and Kaji T: Heparan sulfate chains potentiate cadmium
cytotoxicity in cultured vascular endothelial cells. Arch Toxicol.
90:259–267. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lampugnani MG, Resnati M, Raiteri M,
Pigott R, Pisacane A, Houen G, Ruco LP and Dejana E: A novel
endothelial-specific membrane protein is a marker of cell-cell
contacts. J Cell Biol. 118:1511–1522. 1992.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Lagendijk AK and Hogan BM: VE-cadherin in
vascular development: A coordinator of cell signaling and tissue
morphogenesis. Curr Top Dev Biol. 112:325–352. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Dejana E and Vestweber D: The role of
VE-cadherin in vascular morphogenesis and permeability control.
Prog Mol Biol Transl Sci. 116:119–144. 2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Du L, Dong F, Guo L, Hou Y, Yi F, Liu J
and Xu D: Interleukin-1β increases permeability and upregulates the
expression of vascular endothelial-cadherin in human renal
glomerular endothelial cells. Mol Med Rep. 11:3708–3714.
2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Li L, Chen X, Dong F, Liu Q, Zhang C, Xu
D, Allen TD and Liu J: Dihydroartemisinin up-regulates VE-cadherin
expression in human renal glomerular endothelial cells. J Cell Mol
Med. 22:2028–2032. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhang C, Liu Q, Dong F, Li L, Du J, Xie Q,
Hu H, Yan S, Zhou X, Li C, et al: Catalpol downregulates vascular
endothelial-cadherin expression and induces vascular
hyperpermeability. Mol Med Rep. 13:373–378. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Liu J, Dong F, Jeong J, Masuda T and Lobe
CG: Constitutively active Notch1 signaling promotes
endothelialmesenchymal transition in a conditional transgenic mouse
model. Int J Mol Med. 34:669–676. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Gheorghescu A and Thompson J: Delayed
vasculogenesis and impaired angiogenesis due to altered Ang-2 and
VE-cadherin levels in the chick embryo model following exposure to
cadmium. Pediatr Surg Int. 32:175–186. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li L, Dong F, Xu D, Du L, Yan S, Hu H,
Lobe CG, Yi F, Kapron CM and Liu J: Short-term, low-dose cadmium
exposure induces hyperpermeability in human renal glomerular
endothelial cells. J Appl Toxicol. 36:257–265. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
19
|
Dong F, Guo F, Li L, Guo L, Hou Y, Hao E,
Yan S, Allen TD and Liu J: Cadmium induces vascular permeability
via activation of the p38 MAPK pathway. Biochem Biophys Res Commun.
450:447–452. 2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Etienne-Manneville S and Hall A: Rho
GTPases in cell biology. Nature. 420:629–635. 2002.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang J, Liu X and Zhong Y:
Rho/Rho-associated kinase pathway in glaucoma (Review). Int J
Oncol. 43:1357–1367. 2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Huang Y, Tan Q, Chen R, Cao B and Li W:
Sevoflurane prevents lipopolysaccharide-induced barrier dysfunction
in human lung microvascular endothelial cells: Rho-mediated
alterations of VE-cadherin. Biochem Biophys Res Commun.
468:119–124. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Guo L, Dong F, Hou Y, Cai W, Zhou X, Huang
AL, Yang M, Allen TD and Liu J: Dihydroartemisinin inhibits
vascular endothelial growth factor-induced endothelial cell
migration by a p38 mitogen-activated protein kinase-independent
pathway. Exp Ther Med. 8:1707–1712. 2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Prozialeck WC: Evidence that E-cadherin
may be a target for cadmium toxicity in epithelial cells. Toxicol
Appl Pharmacol. 164:231–249. 2000.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lefranc F, Sauvage S, Van Goietsenoven G,
Mégalizzi V, Lamoral-Theys D, Debeir O, Spiegl-Kreinecker S, Berger
W, Mathieu V, Decaestecker C and Kiss R: Narciclasine, a plant
growth modulator, activates Rho and stress fibers in glioblastoma
cells. Mol Cancer Ther. 8:1739–1750. 2009.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wang X, Dong F, Wang F, Yan S, Chen X,
Tozawa H, Ushijima T, Kapron CM, Wada Y and Liu J: Low dose cadmium
upregulates the expression of von Willebrand factor in endothelial
cells. Toxicol Lett. 290:46–54. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wei T, Jia J, Wada Y, Kapron CM and Liu J:
Dose dependent effects of cadmium on tumor angiogenesis.
Oncotarget. 8:44944–44959. 2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chen X, Li L, Liu F, Hoh J, Kapron CM and
Liu J: Cadmium induces glomerular endothelial cell-specific
expression of complement factor H via the -1635 AP-1 binding site.
J Immunol. 202:1210–1218. 2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Messner B, Turkcan A, Ploner C, Laufer G
and Bernhard D: Cadmium overkill: Autophagy, apoptosis and necrosis
signalling in endothelial cells exposed to cadmium. Cell Mol Life
Sci. 73:1699–1713. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhang H, Li L, Wang Y, Dong F, Chen X, Liu
F, Xu D, Yi F, Kapron CM and Liu J: NF-ĸB signaling maintains the
survival of cadmium-exposed human renal glomerular endothelial
cells. Int J Mol Med. 38:417–422. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Woods JM, Leone M, Klosowska K, Lamar PC,
Shaknovsky TJ and Prozialeck WC: Direct antiangiogenic actions of
cadmium on human vascular endothelial cells. Toxicol In Vitro.
22:643–651. 2008.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wolf MB and Baynes JW: Cadmium and mercury
cause an oxidative stress-induced endothelial dysfunction.
Biometals. 20:73–81. 2007.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Dong Z, Wang L, Xu J, Li Y, Zhang Y, Zhang
S and Miao J: Promotion of autophagy and inhibition of apoptosis by
low concentrations of cadmium in vascular endothelial cells.
Toxicol In Vitro. 23:105–110. 2009.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Eisa-Beygi S and Wen XY: Could
pharmacological curtailment of the RhoA/Rho-kinase pathway reverse
the endothelial barrier dysfunction associated with Ebola virus
infection? Antiviral Res. 114:53–56. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Liu J, Wada Y, Katsura M, Tozawa H, Erwin
N, Kapron CM, Bao G and Liu J: Rho-associated coiled-coil kinase
(ROCK) in molecular regulation of angiogenesis. Theranostics.
8:6053–6069. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Grothaus JS, Ares G, Yuan C, Wood DR and
Hunter CJ: Rho kinase inhibition maintains intestinal and vascular
barrier function by upregulation of occludin in experimental
necrotizing enterocolitis. Am J Physiol Gastrointest Liver Physiol.
315:G514–G528. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Yang M, Chen XM, Du XG, Cao FF, Vijaya
Luxmi S and Shen Q: Continuous blood purification ameliorates
endothelial hyperpermeability in SAP patients with MODS by
regulating tight junction proteins via ROCK. Int J Artif Organs.
36:700–709. 2013.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Warfel JM and D'Agnillo F: Anthrax lethal
toxin-mediated disruption of endothelial VE-cadherin is attenuated
by inhibition of the Rho-associated kinase pathway. Toxins (Basel).
3:1278–1293. 2011.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Daneshjou N, Sieracki N, van Nieuw
Amerongen GP, Conway DE, Schwartz MA, Komarova YA and Malik AB:
Rac1 functions as a reversible tension modulator to stabilize
VE-cadherin trans-interaction. J Cell Biol. 208:23–32.
2015.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Li S, Ai N, Shen M, Dang Y, Chong CM, Pan
P, Kwan YW, Chan SW, Leung GPH, Hoi MPM, et al: Discovery of a ROCK
inhibitor, FPND, which prevents cerebral hemorrhage through
maintaining vascular integrity by interference with VE-cadherin.
Cell Death Discov. 3(17051)2017.PubMed/NCBI View Article : Google Scholar
|