Introduction
Sepsis is a potentially lethal condition that is
commonly encountered in intensive care units (ICUs) (1-4).
Septic shock is a subset of sepsis that is associated with a
greater risk of mortality than sepsis alone (5). The usual host reaction is complex,
aiming to detect and control pathogen incursion, and to initiate
immediate tissue repair. The body activates both cellular and the
humoral immunity, which release massive quantities of
pro-inflammatory and anti-inflammatory mediators, leading to
systemic inflammatory response syndrome (SIRS) (6,7).
Aggravation of these mechanisms can cause a series of events that
may lead to endothelial injury, tissue hypoperfusion, intravascular
coagulation, multiple organ dysfunction syndrome and possibly death
(8).
Despite improvements in critical care medicine,
sepsis and septic shock continue to be among the leading causes of
death and a serious challenge to clinicians and scientists
(1,9). Some of the most important
approaches in sepsis investigation consist of focusing on agents
that may modify systemic inflammation; however, these efforts have
not had much success. Another important feature of sepsis is
cellular apoptosis, which can lead to organ failure (10-13).
Parenchymal cells of the lung and the liver, as well as intestinal
epithelial cells have been reported to exhibit higher levels of
apoptotic death in animal models of sepsis following microvascular
dysfunction and tissue hypoxia compared with non-septic animals
(14,15). Unlike genomic DNA, which is static,
RNA expression can dynamically change between healthy and diseased
states, and thus can provide real-time information regarding
cellular function. miRNAs have been recognized as a class of gene
expression regulators that serve significant roles in normal cell
function and in disease development, including the pathogenesis of
cardiovascular disorders, cancer and inflammation (16). miRNAs are 19-24 nucleotide-long,
endogenous, non-coding RNAs that function as post-transcriptional
suppressors of gene expression by interfering with target mRNA
translation or stability; notably, miRNAs affect multiple target
genes (17). There is growing
evidence that miRNA dysregulation corresponds with the clinical
symptoms of sepsis (18).
Increases in the levels of circulating miRNAs originating from
lipopolysaccharide (LPS)-stimulated monocytes and monocyte-derived
dendritic cells during sepsis have also been reported (19,20).
In addition, the pathophysiological responses associated with
sepsis, such as inflammation, shock or even ileus in the CLP model
may contribute to the upregulation of circulating miRNAs. It has
also been shown that miRNA dysregulation can correspond to clinical
symptoms and severity of sepsis (18,21,22).
Therefore, miRNAs could be potential therapeutic targets for sepsis
management. In the present study, the expression levels of miRNAs
(miR-16, miR-21, miR-27a and miR-34a) were examined, shortlisted
from a group of miRNAs which have been shown to
upregulate/downregulate during sepsis in multiple studies (16,18,23-25).
Thymoquinone (TQ) has been reported to possess
notable immunomodulatory effects, as well as other pharmacological
properties, including anti-inflammatory activity, improvement of
microvascular function and regulation of endothelial nitric oxide
synthase (26,27). In the present study, it was
hypothesized that TQ could regulate miRNA expression levels,
protect organs and improve survival in sepsis. Furthermore, changes
in sepsis markers, such as cytokines, C-reactive protein (CRP),
VEGF, endothelial cell-specific molecule-1 (ESM-1 or Endocan-1),
procalcitonin (PCT) and D-dimer, were detected in a cecal ligation
and puncture (CLP) model (28).
Materials and methods
Animals and CLP model
A total of 60 male Wistar albino rats (Rattus
norvegicus), weighing 200-220 g, age, 8-10 weeks, were used in
the present study. The animals were housed in filter-top cages in a
temperature-controlled environment (23±2˚C, 40-60% humidity with a
12-hour light/dark cycle) and were provided access to standard rat
chow and water ad libitum. The animals were allowed to
acclimate for 1 week before the experiments were conducted. The
present study was approved by the Ethics Committee of the College
of Pharmacy, King Saud University (Riyadh, Saudi Arabia; approval
no. KSU-SE-19-17).
Under aseptic conditions, the rats (n=10/group) were
anesthetized with an intraperitoneal injection of ketamine (80
mg/kg) and xylazine (10 mg/kg). The animal was placed on a surgical
tray 5 min after injection, its abdomen was shaved and an ~1-cm cut
was made into the abdomen. The cecum was exposed, ligated and
punctured at two points with a 21G needle; in the sham group, the
cecum was not punctured. Subsequently, the cecum was placed back
into the body and the incision was sutured. At 1 h following CLP,
each animal received 1 ml sterile saline intraperitoneally for
resuscitation, and the animals were allowed ad libitum
access to food and water. At 24 h after CLP, a single intramuscular
dose of TQ (1 mg/kg; prepared in 10% DMSO) was administered in the
thigh muscles of the hind limbs to the animals in the treatment
group (CLP + TQ). The sham and CLP control groups were treated only
with 10% DMSO. For the survival study, animal mortality was
assessed up to 7 days after TQ treatment. In another set of
experiments, blood samples (0.5 ml) were collected in sterile tubes
from the tail vein prior to TQ administration, and 12, 24 and 36 h
post-treatment. The blood was centrifuged (3,000 x g, 4˚C for 10
min) immediately after collection, and plasma and serum were stored
at -80˚C. The animals were subsequently euthanized with ketamine
(100 mg/kg) and xylazine (10 mg/kg). The death of the animals was
confirmed by lack of movement, absence of heartbeat and respiration
over a sufficient period of time. The liver, kidney and lung
tissues were harvested, dissected and stored in formalin for
histopathological examination.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from the serum samples using
miRNeasy kit (Qiagen, Inc.) according to the manufacturer's
protocol. The quality and quantity of total RNA were checked using
a spectrophotometer. To remove any DNA contamination, the total RNA
was treated with DNase enzyme from a DNase 1 kit (Millipore Sigma).
Total RNA, including miRNA, was reverse transcribed into cDNA using
MystiCq microRNA cDNA Synthesis Mix (Millipore Sigma) according to
the manufacturer's protocol. Prior to miRNA quantification by
RT-qPCR, conventional PCR was run for all the primers used in the
present study to check amplification specificity. Primer details
are presented in Table I. In
conventional PCR, all the primers produced only one amplification
band corresponding to their expected amplicon size (data not
shown). qPCR was subsequently performed using SYBR Green (Roche
Molecular Diagnostics) according to the manufacturer's protocol; U6
small nuclear ribonucleoprotein was used as a reference gene. For
each miRNA, the reaction mixture consisted of 10 µl 2X SYBR Green
master mix, 0.25 µl each reverse and forward primer, 1 µl cDNA and
8.5 µl nuclease-free water. The following thermocycling conditions
were used: 40 cycles of denaturation at 94˚C for 4 min, annealing
at various temperatures as provided in Table I, and extension at 72˚C for 15 sec.
To confirm the product specificity, melting curve analysis was
performed. The relative expression levels of the different miRNAs
were determined using the 2-ΔΔCq method (29). Relative expression levels of the
miRNAs detected in the present study are presented as fold
change.
 | Table IList of primers used for PCR. |
Table I
List of primers used for PCR.
| Gene | Primer sequence
(5'-3') | Annealing
temperature (˚C) | Expected size
(bp) |
|---|
| U6 | F:
CTCGCTTCGGCAGCACA | 55 | 89 |
| | R:
AACGCTTCACGAATTTGCGT | | |
| miR-16 | F:
CCGCTCTAGCAGCACGTAAA | 60 | 82 |
| | R:
CCCTGTCACACTAAAGCAGC | | |
| miR-21 | F:
GTACCACCTTGTCGGGTAGC | 55 | 82 |
| | R:
ATGTCAGACAGCCCATCGAC | | |
| miR-27a | F:
CCTGTGGAGCAGGGCTTAG | 60 | 73 |
| | R:
GCGGAACTTAGCCACTGTGA | | |
| miR-34a | F:
TGGCAGTGTCTTAGCTGGTT | 56 | 81 |
| | R:
AACGTGCAGCACTTCTAGGG | | |
Biomarkers and biochemical analyses of
plasma
ELISA was used to estimate the concentrations of
inflammatory cytokines, including TNF-α (cat. no. PRTA00), IL-1α
(RRA00), IL-2 (SR2000), IL-6 (SR6000B), and IL-10 (SR1000),
according to the manufacturer's protocols (R&D Systems, Inc.).
A quantitative sandwich ELISA was used to assess the levels of
sepsis biomarkers, including CRP (EK0978), and VEGF (EK0540; all
Boster Biological Technology, Pleasanton, CA, USA) and ESM-1
(MBS762527; MyBioSource, Inc. San Diego, CA, USA), according to the
manufacturer's protocols. The concentration of PCT (CSB-E13419r)
was measured using a specific ELISA kit (CUSABIO, Houston, TX,
USA). Similarly, D-dimer (CSB-E12984r-1) levels were determined
using an ELISA kit (Cosmo Bio Co., Ltd.); changes in color were
measured using a spectrophotometer at 450 nm and were quantified
using constructed standard curves. Pertinent biochemical
parameters, such as alanine transaminase (ALT, aspartate
transaminase, (AST) alkaline phosphatase (ALP), serum creatinine
and blood urea nitrogen (BUN), were measured using colorimetric
methods (HUMAN Diagnostics Worldwide, Wiesbaden, Germany).
Histopathological examination
Liver, kidney and lung tissues were sliced into
small pieces and fixed in 10% formalin for 24 h at room
temperature. The fixed tissues were then embedded in paraffin.
Subsequently, the tissues were cut into 4-5-µm sections,
deparaffinized and rehydrated with methanol as previously described
(30). Morphological changes
induced by sepsis with or without TQ treatment were assessed by
evaluating the organ tissues using H&E staining; tissues were
observed under a light microscope as previously described (31).
Statistical analysis
All of the data analysis was carried out using
GraphPad Prism version 6 (GraphPad Software, Inc.) and SPSS 20.0
software package (IBM Corp.). Variables are presented as the mean ±
SEM. Animal survival was assessed using Kaplan-Meier analysis and
log-rank test. For the miRNA expression analysis, the relative
expression of the different miRNAs was determined by the
2-ΔΔCq method, normalized with ΔCq=Average
CqmiRNA-Average CqU6. The relative expression
levels of miRNAs are presented as fold change, calculated from the
mean Cq values for each group. Differences in the miRNA expression,
biochemical parameters and inflammatory biomarkers between the
groups were evaluated using one-way ANOVA followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of TQ on animal survival and
biochemical estimation
Animals that underwent CLP started displaying signs
of infection including lethargy, piloerection, huddling and a
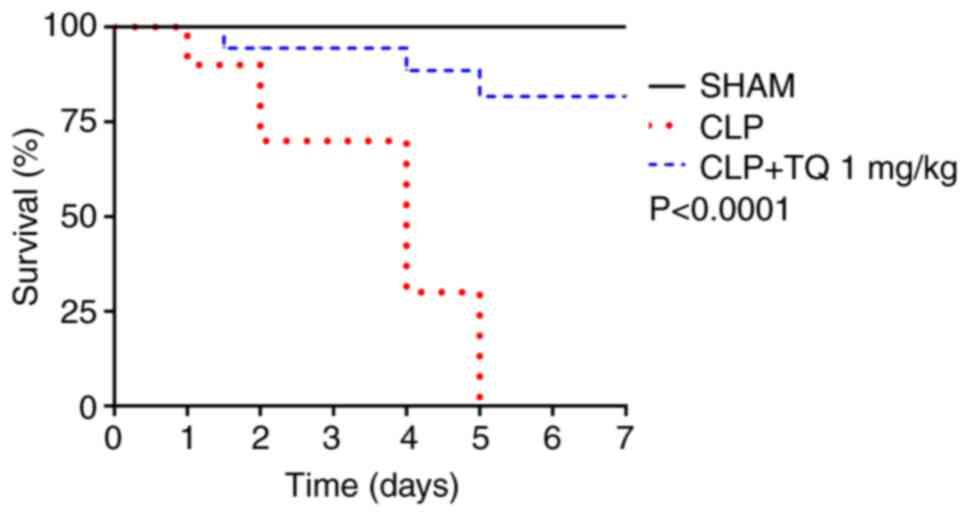
decrease in water and food uptake within 12 h of surgery. As shown
in Fig. 1, 24 h post-CLP, death
started to occur in the CLP groups but significantly decreased when
TQ was administered in the CLP+TQ group. The mortality rate reached
100% in the rats in the untreated CLP group at 5 days, whereas
animals in the TQ treatment group exhibited an increase in survival
rates of up to 80%; no deaths were reported in the sham group
during this time. Results from the biochemical analyses used to
assess organ function showed a significant increase in liver
enzymes (i.e. ALT, AST and ALP) in the CLP group, which were
reduced within 12 h of TQ treatment. Similarly, TQ administration
significantly improved the kidney function as shown by a reduction
in serum creatinine and BUN compared with that in the CLP control
(Table II).
| Table IIComparison of biochemical parameters
between different experimental groups. |
Table II
Comparison of biochemical parameters
between different experimental groups.
| | Time after TQ,
h |
|---|
| Parameter | Group | 0a | 12 | 24 | 36 |
|---|
| ALP, IU/l | Sham | 118.79±4.73 | 131.85±5.94 | 129.41±5.37 | 121.32±5.18 |
| | CLP |
292.66±6.94b |
358.25±12.20b |
388.56±18.20b |
418.74±10.68b |
| | CLP + TQ |
278.08±5.57b |
191.12±4.99b,c |
236.01±5.50b,c |
260.44±6.10b,c |
| AST, IU/l | Sham | 121.45±3.76 | 129.19±4.65 | 119.79±3.26 | 124.52±4.84 |
| | CLP |
238.89±5.13b |
275.14±7.64b |
289.04±8.86b |
320.86±2.96b |
| | CLP + TQ |
224.35±8.38b,c |
157.76±6.48b,c |
177.35±7.59b,c |
187.49±6.86b,c |
| ALT, IU/l | Sham | 29.66±1.94 | 34.87±2.49 | 31.76±2.09 | 27.06±2.78 |
| | CLP |
53.37±2.01b |
58.15±1.62b |
62.42±1.82b |
69.32±1.39b |
| | CLP + TQ |
51.31±0.58b |
38.12±1.55c |
41.02±1.08b,c |
46.51±1.27b,c |
| Creatinine,
mg/dl | Sham | 1.30±0.12 | 1.27±0.22 | 1.32±0.43 | 1.30±0.59 |
| | CLP | 2.15±1.22 |
2.39±0.84b |
2.66±1.02b |
3.13±0.60b |
| | CLP + TQ |
2.18±0.51b |
1.58±0.65c |
1.75±.22b,c |
1.91±0.78b,c |
| BUN, mg/dl | Sham | 49.07±0.65 | 51.98±0.77 | 53.69±0.52 | 52.85±0.61 |
| | CLP |
91.16±0.38b |
101.95±0.65b |
125.85±0.76b |
129.37±0.35b |
| | CLP + TQ |
95.87±0.20b,c | 67.65
±0.66b,c |
74.51±0.42b,c |
81.75±0.91b,c |
Effect of TQ on miRNA and biomarker
estimation
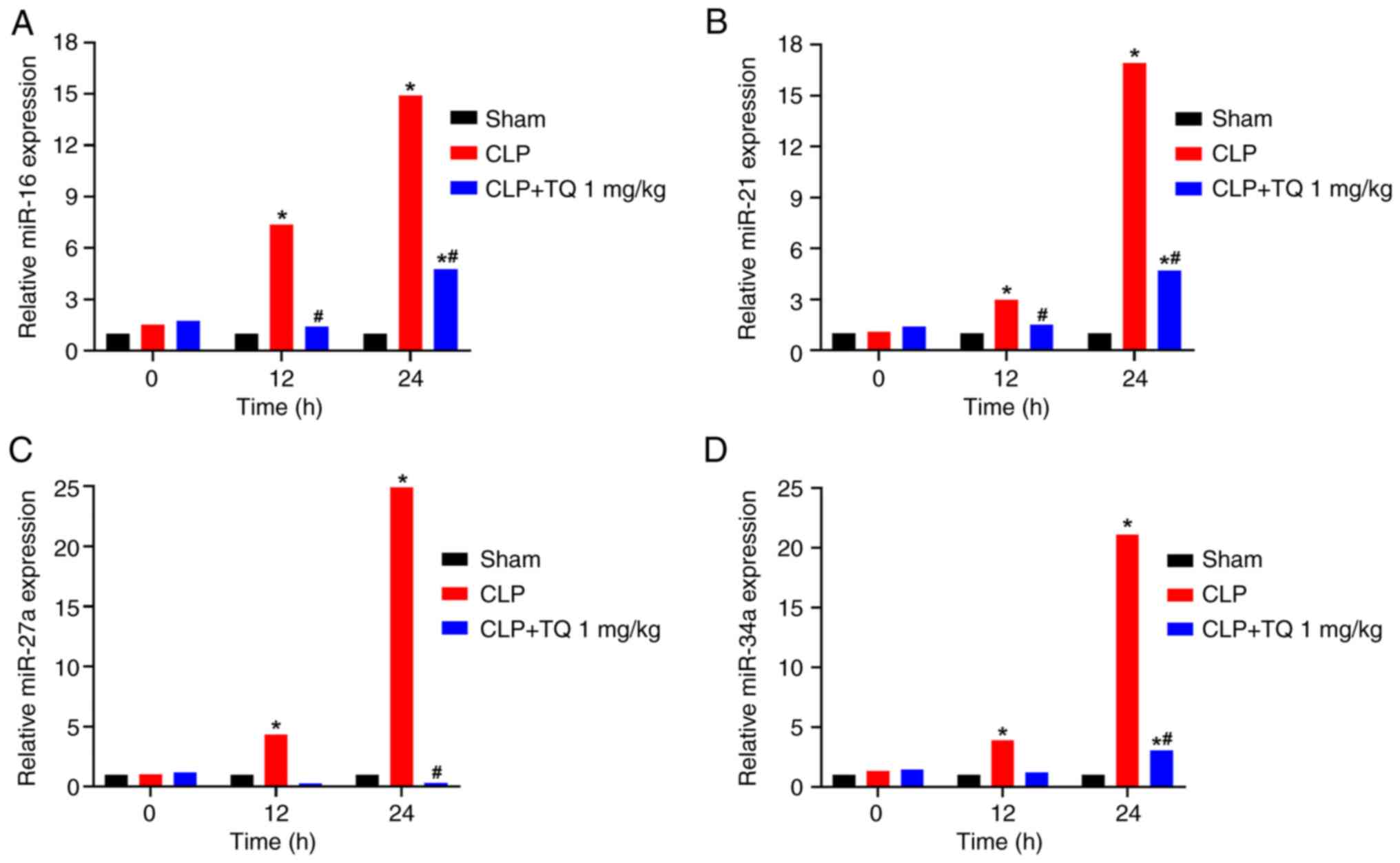
The expression levels of miR-16, miR-21, miR-27a and
miR-34a were significantly increased at 12 and 24 h after CLP
compared with the sham group (Fig.
2). Treatment with TQ significantly downregulated the miRNA
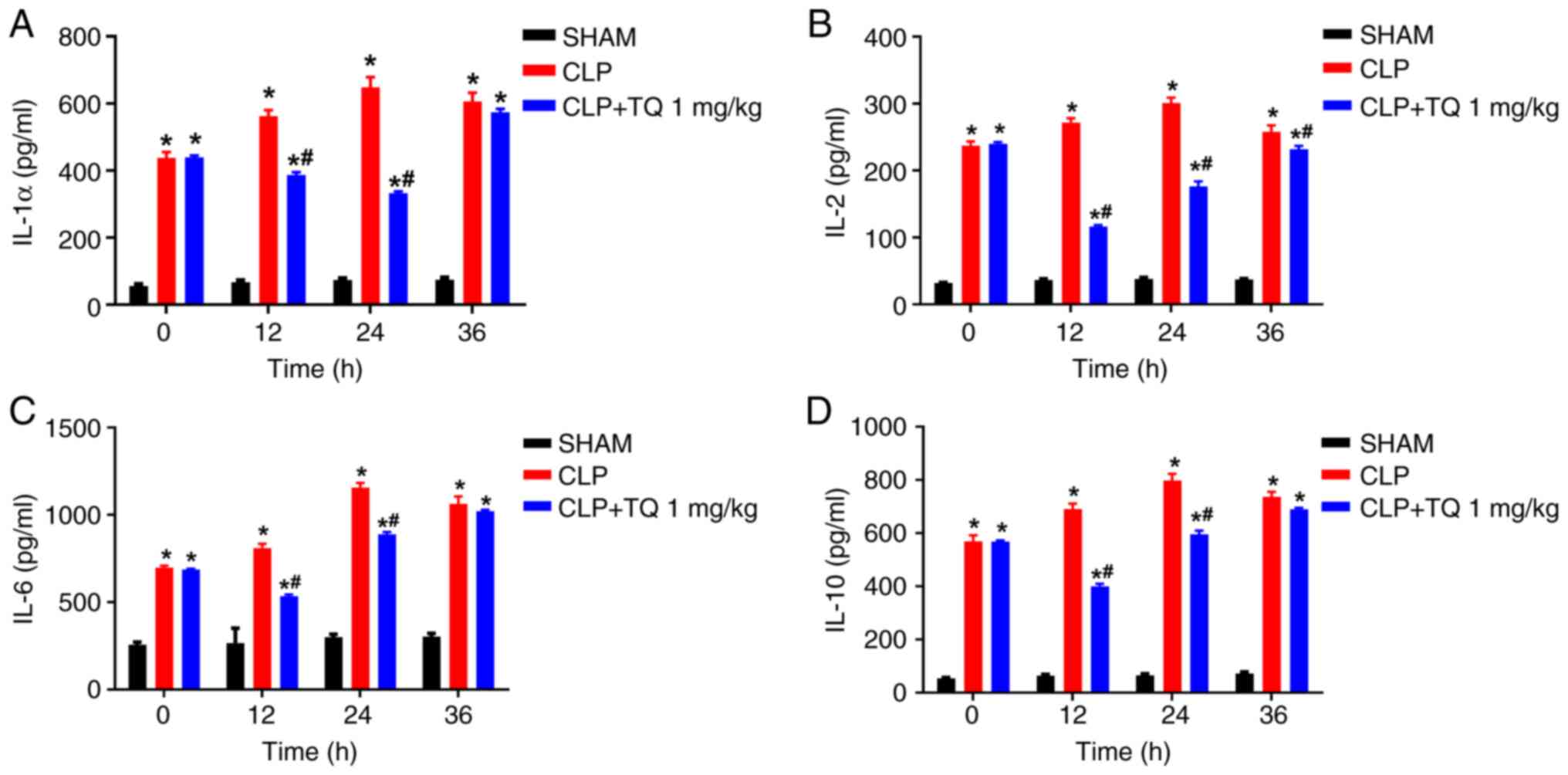
expression levels by 40-80%. Similarly, the concentrations of
IL-1α, IL-2, IL-6, IL-10 and TNF-α, were significantly increased in
the CLP group compared with those in the sham group, and TQ
treatment resulted in a significant reduction in the levels of the
inflammatory cytokines in a time dependent manner and effect of TQ
seems to diminish by the 36 h (Figs.
3 and 4A; P<0.05).
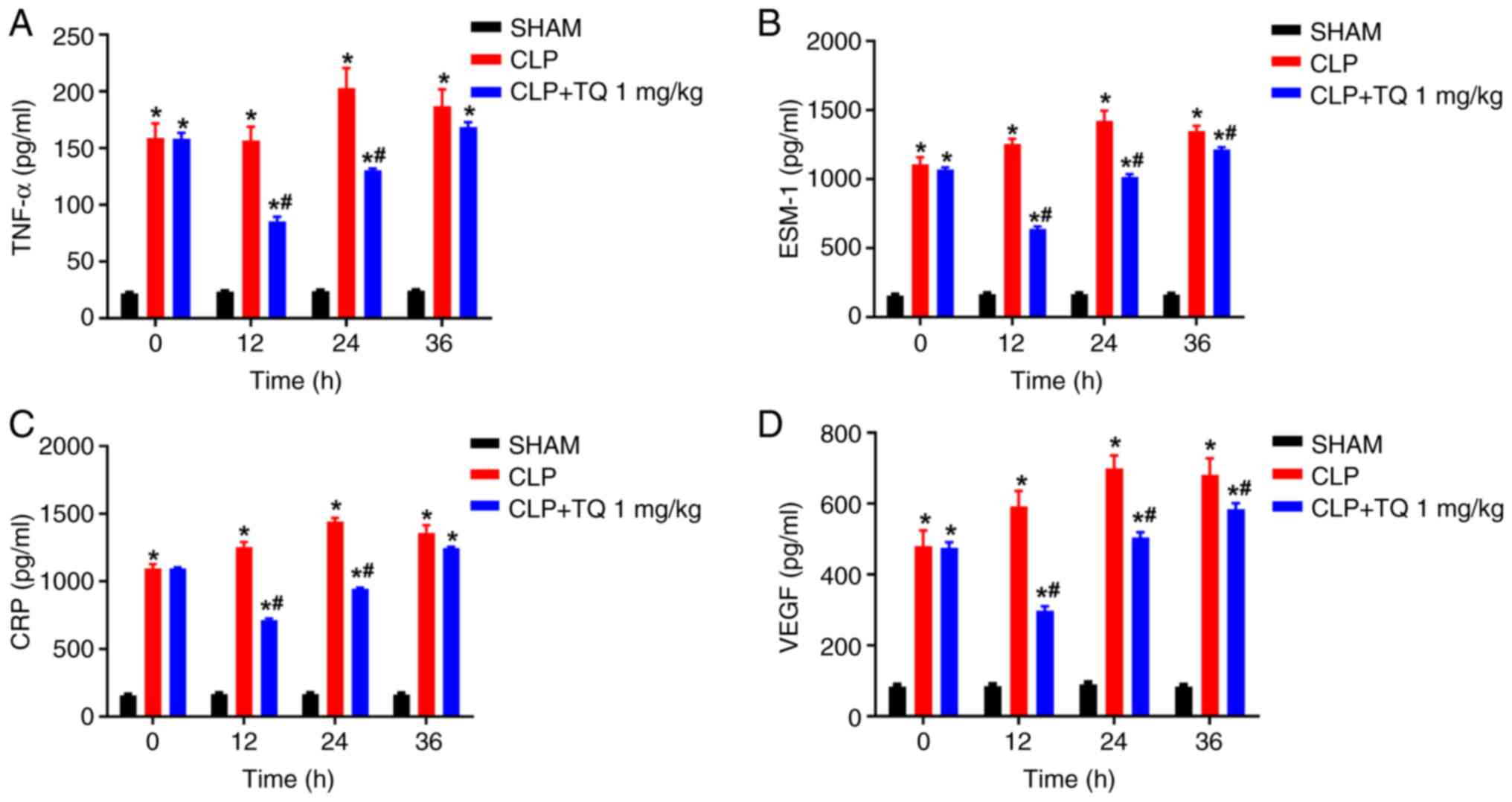
The effect of CLP on sepsis biomarkers CRP, VEGF and
ESM-1 was also determined. CLP resulted in a notable increase in
the concentrations of the sepsis biomarkers, which was reversed by
TQ administration (Fig. 4B-D;
P<0.05). TQ treatment also mitigated the increased levels of
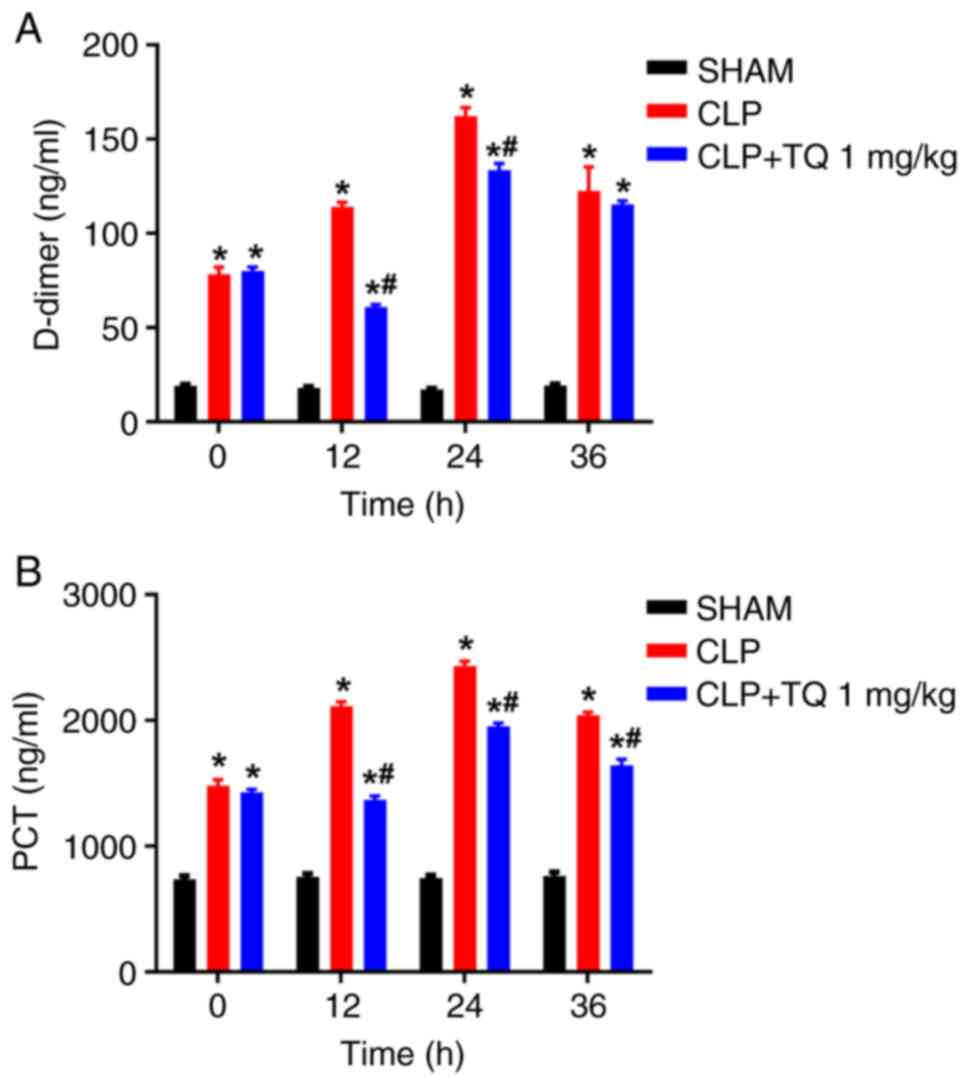
D-dimer and PCT in septic model animals (Fig. 5A and B), which was consistent with its
antiseptic effect.
Histopathology observations
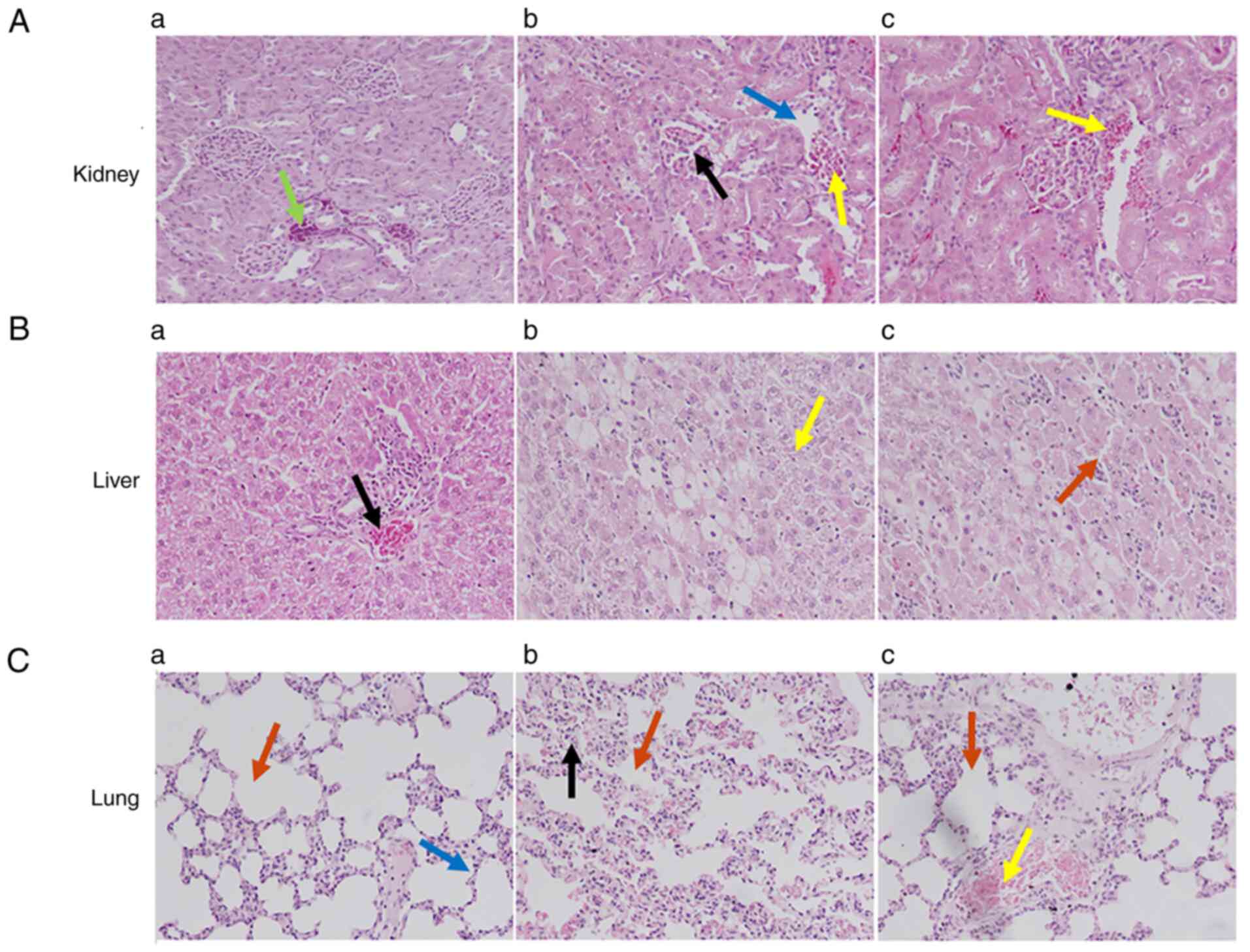
The histological evaluation (Fig. 6) of rat renal tissues from the sham
group showed normal architecture of the renal cortex, renal
glomeruli and renal tubules (Fig.
6Aa). By contrast, animals having undergone the CLP procedure
exhibited deterioration of the renal cortex and medullary tubules,
and some interstitial hemorrhages with mononuclear cell
infiltration (Fig. 6Ab). In
addition, epithelial tubular necrotic areas and cellular atrophy
were observed. CLP model animals treated with TQ exhibited reduced
deterioration of the renal cortex, renal glomeruli and renal
tubules (Fig. 6Ac); in addition,
reduced dilatation of renal glomeruli and cortical tubules was
observed. No signs of deterioration in cellular organelles and no
inferential hemorrhages were found (Fig. 6Ac).
Similarly, the rat liver from the sham group
exhibited normal liver composition with a prominent nucleus, well
preserved cytoplasm, central vein and a compact arrangement of
hepatocytes without any fatty lobulation (Fig. 6Ba). By contrast, animals in the
untreated CLP group exhibited acute cellular swelling, congestion
of the central vein, medium centrilobular necrosis of hepatocytes
and apoptotic bodies with sinusoidal dilatation (Fig. 6Bb). An acute massive focal
infiltration of mononuclear cells in the portal area, and
sinusoidal infiltration in the central zones with vacuolization and
steatosis were also evident (Fig.
6Bb). CLP animals treated with TQ displayed distinct features,
such as pentagonal or hexagonal lobules, with central veins and
borderline hepatic triads or tetrads, engrained in connective
tissues (Fig. 6Bc). A mild degree
of centrilobular necrosis of hepatocytes was observed but no
apoptotic bodies were seen, whereas slight sinusoidal dilatation
and mild congestion of central vein were detected. A small number
of mononuclear cells and sinusoidal intrusion in the central zones
with less vacuolization and steatosis was detected but without
signs of edematous tissue (Fig.
6Bc).
Lung tissues from animals in the sham group
displayed normal structure and composition, with a well-preserved
alveolar space and pulmonary interstitium, and no inflammatory cell
infiltration into the alveolar cavity (Fig. 6Ca). Evaluation of lung tissues of
the animals in the CLP group showed acute edema, emphysema and
pulmonary interstitial hyperemia, resulting in impaired alveolar
architecture with penetrations of mononuclear cells causing
alveolar congestion and hyperemia in the pulmonary capillaries
(Fig. 6Cb). By contrast, tissues
from CLP model animals treated with TQ displayed limited
inflammatory cell infiltration and restoration of the normal
alveolar structure (Fig. 6Cc).
Discussion
Sepsis is a detrimental condition with a high
mortality rate in ICUs; therefore, scientists and clinicians have
focused on identifying new therapeutic modalities to combat its
negative outcomes (32,33). The CLP animal model has been
extensively used to study new approaches for sepsis management
(34). The constant source of
bacteria in this model is the punctured cecum and endotoxin, which
is the main constituent of the external membrane of bacterial cell
walls, activates several pathophysiological events of gram-negative
sepsis (35). The overproduction
of reactive oxygen species (ROS) produced under these conditions
has been implicated in tissue injury (36). Subsequently, CLP triggers
pathological changes in the lung, liver and kidney tissues,
signifying varying degrees of organ injury. In the present study,
it was demonstrated that treatment of CLP model animals with TQ
improved animal survival by up to 80% over a period of 7 days. This
coincided with a significant reduction in the expression levels of
specific miRNAs, levels of circulating inflammatory cytokines and
early-stage sepsis biomarkers, as well as preservation of liver,
lung and kidney functions.
Among the circulating miRNAs that have been reported
to be upregulated following CLP, miR-16 serves a vital role in
sepsis. Notably, deletion of miR-16 in myeloid cells has been shown
to significantly decrease Escherichia coli-associated
mortality in several sepsis models (37). Consistently, miR-16 overexpression
can decrease both phagocytosis and production of mitochondrial ROS.
Additionally, lack of miR-16 has been reported to enhance secretion
of cytokines and chemokines from bone-marrow-derived macrophages at
the early stages of infection (38). Furthermore, Gao and Yu (39) identified IκB kinase-β (IKKβ) as a
direct target of miR-16, the expression of which was negatively
regulated by miR-16 at the mRNA and protein expression levels,
indicating that miR-16 may suppress the inflammatory response by
inhibiting the IKKβ/NF-κB signaling pathway. Notably, bacterial
infection-induced miR-16 upregulation may cause significant organ
damage and cell death. Another study demonstrated a positive
correlation between circulating miR-16 in serum and death of
patients infected with sepsis (23). Therefore, it may be concluded that
miR-16 has a crucial role in tissue damage and cell death in sepsis
and associated SIRS.
Another miRNA that is highly expressed in different
immune cells, including monocytes, macrophages, T and B
lymphocytes, and dendritic cells, is miR-21(25). Numerous studies have shown that
different inflammatory stimuli, such as LPS, lipids (prostaglandin
E2 and resolvin A1) and cytokines, can trigger miR-21
expression (25,40,41).
miR-21 has also been shown to be upregulated in acute sepsis and
sustained in late sepsis in patients and mice (41-43).
McClure et al (44)
demonstrated that administration of miR-21 antagomir to BALB/c mice
improved animal survival and decreased the bacterial load following
CLP. Determination of the functions of miR-21 have been difficult
owing to its multiple mRNA target interactions, as well as its
complex regulation in response to extracellular signals (25). Furthermore, miR-21 has been
associated with numerous key processes of inflammation, such as
detection and response to homeostatic disturbances throughout the
body as well as coordinating these responses appropriately, thus
serving a dynamic function in inflammatory responses (25,45).
Differing from other mediators, the presence of miR-21 is not
restricted to pro-inflammatory or immunosuppressive states; it can
act as a crucial signal to mediate the homeostasis between both
states (25,45).
miR-27a is another miRNA that has been reported to
serve an important role in the regulation of inflammatory responses
in sepsis (24). Inhibition of
miR-27a has been shown to downregulate the expression levels of
TNF-α and IL-6 by reducing the phosphorylation levels of NF-κB p65
subunit and through the inhibition of its DNA-binding activity
(24). Moreover, miR-27a
neutralization has been suggested to upregulate peroxisome
proliferator-activated receptor γ, possibly inhibiting the
production of monocyte inflammatory cytokines and activating
macrophages to boost innate defense against pathogens, thereby
downregulating TNF-α expression, relieving inflammation and
increasing the survival rate of patients with sepsis (46,47).
A previous study has reported that accumulation of
ROS and reduced antioxidant enzyme activities increase oxidative
stress and serve important roles in the progression of sepsis,
leading to mitochondrial dysfunction and organ failure (48). miR-34a regulates oxidative stress
and autophagy through the inhibition of silent information
regulator T1 (SIRT1) and autophagy gene 4B signaling. Differential
miR-34a expression has also been suggested as a potential
prognostic biomarker, in addition to providing insight into the
mechanisms of endothelial dysfunction of septic shock. Notably,
miR-34a can target and inhibit BCL-2 and SIRT1, which are important
negative regulators of endothelium apoptosis and cellular
senescence (49,50). Therefore, increased circulating
miR-34a levels in response to cytokine stimulation may contribute
to septic shock-induced endothelial dysfunction through its effects
on apoptosis and senescence.
Collectively, targeting these miRNAs, particularly
after the initial phase of infection, may provide a novel
therapeutic and/or diagnostic tool for sepsis. The present study
demonstrated that CLP-induced experimental sepsis resulted in a
time-dependent upregulation in the expression levels of these
selected miRNAs, which was subsequently mitigated by treatment with
TQ.
Multiple cytokine biomarkers have been identified
over the past few decades for the diagnosis and treatment of
sepsis, of which TNF-α, IL-1, IL-2, IL-6 and IL-10 are some of the
most important mediators of inflammation (51,52).
TNF-α serves a role in apoptosis, cell survival, inflammation and
immunity (53). IL-1 is a
prototypical pro-inflammatory cytokine that aids in the stimulation
of local and systemic responses. TNF-α and IL-1 have been
demonstrated to augment inflammatory cascades through the
stimulation of macrophages, which release pro-inflammatory
cytokines, such as IL-6 and IL-8, as well as ROSs, reactive
nitrogen species and lipid mediators that are vital in
sepsis-induced organ failure (54,55).
Results from the present study demonstrated that TQ treatment
reduced TNF-α and IL-1α levels during sepsis, which may explain its
protective effects on CLP-induced sepsis. These results are
consistent with our previous findings showing beneficial effects of
TQ using a slightly different sepsis model (27,56).
IL-2 has varying and sometimes opposing functions
during inflammation, contributing to both the initiation and the
termination of the immune response (55). The increased plasma levels of IL-2
in response to CLP infection may act as a prognostic marker for
septic shock (55). IL-2 is
released from T and B lymphocytes, possibly contributing to the
pathogenesis of sepsis (57). In
the present study, TQ treatment significantly decreased the levels
of IL-2 in septic animals. Another important cytokine mediator that
enhances the production of acute phase reactants in the liver is
IL-6(58). IL-6 is rapidly
produced in response to infections and tissue injuries,
contributing to host defense by stimulating the acute phase
responses, hematopoiesis and immune reactions. The present study
results confirmed that TQ could regulate IL-6 concentration in a
CLP model. IL-10 is a key anti-inflammatory cytokine, which is
released in response to TNF-α and IL-1α (59-61).
Notably, the present study also demonstrated that TQ reduced IL-10
levels at an early time point, which might be a consequence of the
initial downregulation of the release of pro-inflammatory cytokines
in septic animals.
Precise assessment of sepsis remains a challenge in
most ICU settings; therefore, there is a high demand for accurate
and early diagnostic biomarkers (62). An early diagnostic biomarker for
sepsis must have a rapid turnaround time and be widely available
for effective therapeutic potential. CRP, a well-known marker of
inflammation, has been suggested to bind the phospholipid
components of microorganisms, thereby enabling eradication by
macrophages (63,64). During systemic inflammation of a
microbial origin, the levels of CRP are increased, as observed in
the present study. TQ treatment was able to decrease CRP levels in
the CLP model rats, thus indicating its ability to ameliorate the
levels of early phase reactant proteins.
VEGF was originally considered to be only a potent
stimulator of endothelial permeability, but has since been reported
to enhance proliferation and survival of endothelial cells
(65,66). Previous studies have shown that a
number of features of VEGF make it a strong candidate to control
inflammation (66-68).
An association between higher circulating levels of VEGF and severe
human septic shock has also been reported (69,70).
Notably, sepsis or septic shock is associated with a time-dependent
surge in the circulating levels of VEGF (71,72).
The present findings revealed that the circulating levels of VEGF
were significantly increased following CLP, whereas TQ
administration substantially decreased these levels. These findings
suggested that TQ may also exert its effect on sepsis, at least
partially, by controlling the production of VEGF which are in
support of our previous finding using a different sepsis model
(27,56).
Endotoxins, such as LPS, induce endothelial cell
contraction and the development of intercellular gaps, thus
increasing permeability of blood vessels (73,74).
In addition, it has been demonstrated that ESM-1 can cause vascular
responses in in vivo models of inflammation, as well as
increase barrier permeability and the passage of leukocytes by
upregulating cytokines during sepsis (75). ESM-1 release has been shown to be
neutralized by ESM-1 antibodies in a CLP-induced mouse model of
sepsis (75). The treatment of
rats with TQ in the present study markedly reduced ESM-1 release,
providing consistent evidence of its promising effect for the
management of sepsis and septic shock (56).
PCT is a peptide precursor of calcitonin hormone
that has been identified as a biomarker of bacterial infection
(76,77). Moreover, it can be used in
evaluating a suitable treatment response, determining severity of
sepsis, and estimating morbidity and mortality rates (77-82).
Similarly, increased D-dimer levels are another sign of systemic
thrombosis, which has been identified as an effective predictor of
mortality in severe sepsis (83-86).
In the present study, CLP increased PCT and D-dimer levels in a
time-dependent manner. Treatment with TQ reduced the levels of both
markers, particularly at the 12-h time point, and improved the
outcome of sepsis, including animal survival.
From a histopathological perspective, liver, kidney
and lung tissue samples of the sham group had normal features,
whereas rats subjected to CLP exhibited substantial
histopathological alterations. For example, liver sections of the
CLP group displayed penetration of inflammatory cells along with
necrotic damage; this may be due to increased ROS and lipid
peroxidation (12,87). TQ has been shown to attenuate ROS
and lipid peroxidation, as well as organ injury caused by various
agents (27,88). In the present study, TQ treatment
improved both morphological and histological features of the
tissue. TQ significantly mitigated kidney and lung injuries in CLP
model rats. In particular, renal tissue of septic rats exhibited
degeneration in the renal cortex and medullary tubules, which was
ameliorated by TQ. Similarly, lung tissue exhibited damage to the
alveolar structure with infiltration of mononuclear cells; these
pathological changes were restored to normal alveolar architecture
with TQ treatment.
In conclusion, the present study demonstrated the
effects of TQ in sepsis management by reducing morbidity and
mortality in a CLP model. These potential effects are thought to be
due to immunomodulation and control of inflammatory status,
including effects on the expression of relevant miRNAs, under
septic conditions.
Acknowledgements
The authors extend their sincere appreciation to the
Deanship of Scientific Research at King Saud University (Riyadh,
Saudi Arabia) for carrying out this work through research group no.
RG-1441-337.
Funding
Funding: The present study was supported by the Deanship of
Scientific Research, KSU (RG-1441-337).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
KMA and BLJ wrote the manuscript, and KMA designed
the experiments. AA and MR carried out animal experiments and
analyzed the data. MUR and BLJ carried out the microRNA study. KMA,
AA and BLJ confirm the authenticity of all the raw data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the College of Pharmacy, King Saud University (Riyadh,
Saudi Arabia; approval no. KSU-SE-19-17).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fleischmann C, Scherag A, Adhikari NKJ,
Hartog CS, Tsaganos T, Schlattmann P, Angus DC and Reinhart K:
International Forum of Acute Care Trialists. Assessment of global
incidence and mortality of hospital-treated sepsis. Current
Estimates and Limitations. Am J Respir Crit Care Med. 193:259–272.
2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Martin GS, Mannino DM, Eaton S and Moss M:
The epidemiology of sepsis in the United States from 1979 through
2000. New Engl J Med. 348:1546–1554. 2003.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Angus DC and Wax RS: Epidemiology of
sepsis: An update. Crit Care Med. 29 (7 Suppl):S109–S116.
2001.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Bone RC, Balk RA, Cerra FB, Dellinger RP,
Fein AM, Knaus WA, Schein RM and Sibbald WJ: Definitions for sepsis
and organ failure and guidelines for the use of innovative
therapies in sepsis. Chest. 101:1644–1655. 1992.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Singer M, Deutschman CS, Seymour CW,
Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche
JD, Coopersmith CM, et al: The third international consensus
definitions for sepsis and septic shock (Sepsis-3). JAMA.
315:801–810. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Koyama I, Matsunaga T, Harada T, Hokari S
and Komoda T: Alkaline phosphatases reduce toxicity of
lipopolysaccharides in vivo and in vitro through dephosphorylation.
Clin Biochem. 35:455–461. 2002.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Männel DN: Advances in sepsis research
derived from animal models. Int J Med Microbiol. 297:393–400.
2007.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bentala H, Verweij WR, der Vlag AH-V, van
Loenen-Weemaes AM, Meijer DKF and Poelstra K: Removal of phosphate
from lipid a as a strategy to detoxify lipopolysaccharide. Shock.
18:561–566. 2002.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Annane D, Buisson CB, Cariou A, Martin C,
Misset B, Renault A, Lehmann B, Millul V, Maxime V and Bellissant
E: APROCCHSS Investigators for the TRIGGERSEP Network. Design and
conduct of the activated protein C and corticosteroids for human
septic shock (APROCCHSS) trial. Ann Intensive Care.
6(43)2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Ayala A and Chaudry IH: IMMUNE dysfunction
in murine polymicrobial sepsis. Shock. 5 (Suppl 1):S27–S38.
1996.PubMed/NCBI
|
|
11
|
Lang JD and Matute-Bello G: Lymphocytes,
apoptosis and sepsis: Making the jump from mice to humans. Crit
Care. 13(109)2009.PubMed/NCBI View
Article : Google Scholar
|
|
12
|
Matsuda H, Ishikado A, Nishida N, Ninomiya
K, Fujiwara H, Kobayashi Y and Yoshikawa M: Hepatoprotective,
superoxide scavenging, and antioxidative activities of aromatic
constituents from the bark of Betula platyphylla var. japonica.
Bioorg Med Chem Lett. 8:2939–2944. 1998.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wesche DE, Lomas-Neira JL, Perl M, Chung
CS and Ayala A: Leukocyte apoptosis and its significance in sepsis
and shock. J Leuk Biol. 78:325–337. 2005.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Coopersmith CM, Chang KC, Swanson PE,
Tinsley KW, Stromberg PE, Buchman TG, Karl IE and Hotchkiss RS:
Overexpression of Bcl-2 in the intestinal epithelium improves
survival in septic mice. Crit Care Med. 30:195–201. 2002.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Coopersmith CM: Inhibition of intestinal
epithelial apoptosis and survival in a murine model of
pneumonia-induced sepsis. JAMA. 287:1716–1721. 2002.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Sonkoly E and Pivarcsi A: MicroRNAs in
inflammation. Int Rev Immunol. 28:535–561. 2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Ratti M, Lampis A, Ghidini M, Salati M,
Mirchev MB, Valeri N and Hahne JC: MicroRNAs (miRNAs) and long
non-coding RNAs (lncRNAs) as new tools for cancer therapy: First
steps from bench to bedside. Target Oncol. 15:261–278.
2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ardekani AM and Naeini MM: The role of
microRNAs in human diseases. Avicenna J Med Biotechnol. 2:161–179.
2010.PubMed/NCBI
|
|
19
|
Taganov KD, Boldin MP, Chang KJ and
Baltimore D: NF-kappaB-dependent induction of microRNA miR-146, an
inhibitor targeted to signaling proteins of innate immune
responses. Proc Natl Acad Sci. 103:12481–12486. 2006.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ceppi M, Pereira PM, Dunand-Sauthier I,
Barras E, Reith W, Santos MA and Pierre P: MicroRNA-155 modulates
the interleukin-1 signaling pathway in activated human
monocyte-derived dendritic cells. Proc Natl Acad Sci.
106:2735–2740. 2009.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Essandoh K and Fan GC: Role of
extracellular and intracellular microRNAs in sepsis. Biochim
Biophys Acta. 1842:2155–2162. 2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
23
|
Wang HJ, Zhang PJ, Chen WJ, Feng D, Jia YH
and Xie LX: Four serum microRNAs identified as diagnostic
biomarkers of sepsis. J Trauma Acute Care Surg. 73:850–854.
2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang Z, Ruan Z, Mao Y, Dong W, Zhang Y,
Yin N and Jiang L: miR-27a is up regulated and promotes
inflammatory response in sepsis. Cell Immunol. 290:190–195.
2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Sheedy FJ: Turning 21: Induction of miR-21
as a key switch in the inflammatory response. Front Immunol.
6(19)2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Woo CC, Kumar AP, Sethi G and Tan KHB:
Thymoquinone: Potential cure for inflammatory disorders and cancer.
Biochem Pharmacol. 83:443–451. 2012.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Alkharfy KM, Ahmad A, Raish M and
Vanhoutte PM: Thymoquinone modulates nitric oxide production and
improves organ dysfunction of sepsis. Life Sci. 143:131–138.
2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Hubbard WJ, Choudhry M, Schwacha MG, Kerby
JD, Rue LW III, Bland KI and Chaudry IH: Cecal ligation and
puncture. Shock. 24:52–57. 2005.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Bancroft J and Gamble M: Theory and
practice of histological techniques. Churchill Livingstone Pub,
Edinburgh, 2002.
|
|
31
|
Drury R and Wallington E: Carlton's
histological techniques, 4th ed. 1967. Oxford University Press, New
York, Toronto, 1967.
|
|
32
|
Calandra T, Glauser MP, Schellekens J and
Verhoef J: Treatment of gram-negative septic shock with human igg
antibody to escherichia coli J5: A prospective, double-blind,
randomized trial. J Infect Dis. 158:312–319. 1988.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Frazier WJ and Hall MW: Immunoparalysis
and adverse outcomes from critical illness. Pediatr Clin North Am.
55:647–668. 2008.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Dejager L, Pinheiro I, Dejonckheere E and
Libert C: Cecal ligation and puncture: The gold standard model for
polymicrobial sepsis? Trends Microbiol. 19:198–208. 2011.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Menezes G, Amaral S, Alvarenga D and Cara
D: Surgical procedures to an experimental polymicrobial sepsis:
Cecal Ligation and Puncture. Braz J Vet Pathol. 1:77–80. 2008.
|
|
36
|
Mittal M, Siddiqui MR, Tran K, Reddy SP
and Malik AB: Reactive oxygen species in inflammation and tissue
injury. Antiox Redox Signal. 20:1126–1167. 2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Precone V, Stornaiuolo G, Amato A,
Brancaccio G, Nardiello S and Gaeta GB: Different changes in
mitochondrial apoptotic pathway in lymphocytes and granulocytes in
cirrhotic patients with sepsis. Liver Int. 33:834–842.
2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Moon HG, Yang J, Zheng Y and Jin Y:
MiR-15a/16 regulates macrophage phagocytosis after bacterial
infection. J Immunol. 193:4558–4567. 2014.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Gao Y and Yu Z: MicroRNA-16 inhibits
interleukin-13-induced inflammatory cytokine secretion and mucus
production in nasal epithelial cells by suppressing the IκB kinase
β/nuclear factor-κB pathway. Mol Med Rep. 18:4042–4050.
2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Löffler D, Brocke-Heidrich K, Pfeifer G,
Stocsits C, Hackermüller J, Kretzschmar AK, Burger R, Gramatzki M,
Blumert C, Bauer K, et al: Interleukin-6-dependent survival of
multiple myeloma cells involves the Stat3-mediated induction of
microRNA-21 through a highly conserved enhancer. Blood.
110:1330–1333. 2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Sheedy FJ, Palsson-McDermott E, Hennessy
EJ, Martin C, O'Leary JJ, Ruan Q, Johnson DS, Chen Y and O'Neill
LAJ: Negative regulation of TLR4 via targeting of the
proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat
Immunol. 11:141–147. 2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
McClure C, Brudecki L, Ferguson DA, Yao
ZQ, Moorman JP, McCall CE and Gazzar ME: MicroRNA 21 (miR-21) and
miR-181b couple with nfi-a to generate myeloid-derived suppressor
cells and promote immunosuppression in late sepsis. Infect Immun.
82:3816–3825. 2014.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Goodwin AJ, Guo C, Cook JA, Wolf B,
Halushka PV and Fan H: Plasma levels of microRNA are altered with
the development of shock in human sepsis: An observational study.
Crit Care. 19:2015.PubMed/NCBI View Article : Google Scholar
|
|
44
|
McClure C, Ali E, Youssef D, Yao ZQ,
McCall CE and El Gazzar M: NFI-A disrupts myeloid cell
differentiation and maturation in septic mice. J Leukoc Biol.
99:201–211. 2016.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Lu TX and Rothenberg ME: Diagnostic,
functional, and therapeutic roles of microRNA in allergic diseases.
J Allergy Clin Immunol. 132:3–13; quiz 14. 2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Jiang C, Ting AT and Seed B: PPAR-gamma
agonists inhibit production of monocyte inflammatory cytokines.
Nature. 391:82–86. 1998.PubMed/NCBI View
Article : Google Scholar
|
|
47
|
Chawla A: Control of macrophage activation
and function by PPARs. Circul Res. 106:1559–1569. 2010.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Mantzarlis K, Tsolaki V and Zakynthinos E:
Role of oxidative stress and mitochondrial dysfunction in sepsis
and potential therapies. Oxid Med Cell Longev.
2017(5985209)2017.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Ackermann EJ, Taylor JK, Narayana R and
Bennett CF: The role of antiapoptotic Bcl-2 family members in
endothelial apoptosis elucidated with antisense oligonucleotides. J
Biol Chem. 274:11245–11252. 1999.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Potente M and Dimmeler S: Emerging roles
of SIRT1 in vascular endothelial homeostasis. Cell Cycle.
7:2117–2122. 2008.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Zhang JM and An J: Cytokines,
inflammation, and pain. Int Anesthesiol Clin. 45:27–37.
2007.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Schulte W, Bernhagen J and Bucala R:
Cytokines in sepsis: Potent immunoregulators and potential
therapeutic targets-an updated view. Mediators Inflamm.
2013(165974)2013.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Parameswaran N and Patial S: Tumor
necrosis factor-α signaling in macrophages. Crit Rev Eukaryot Gene
Expr. 20:87–103. 2010.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Cohen J: The immunopathogenesis of sepsis.
Nature. 420:885–891. 2002.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Fong Y, Tracey KJ, Moldawer LL, Hesse DG,
Manogue KB, Kenney JS, Lee AT, Kuo GC, Allison AC and Lowry SF:
Antibodies to cachectin/tumor necrosis factor reduce interleukin 1
beta and interleukin 6 appearance during lethal bacteremia. J Exp
Med. 170:1627–1633. 1989.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Alkharfy KM, Ahmad A, Jan BL and Raish M:
Thymoquinone reduces mortality and suppresses early acute
inflammatory markers of sepsis in a mouse model. Biomed
Pharmacother. 98:801–805. 2018.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Hoyer KK, Dooms H, Barron L and Abbas AK:
Interleukin-2 in the development and control of inflammatory
disease. Immunol Rev. 226:19–28. 2008.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Tanaka T, Narazaki M and Kishimoto T: IL-6
in inflammation, immunity, and disease. Cold Spring Harb Perspect
Biol. 6:a016295. 2014.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Pestka S, Krause CD, Sarkar D, Walter MR,
Shi Y and Fisher PB: Interleukin-10andrelatedcytokines
andreceptors. Ann Rev Immunol. 22:929–979. 2004.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Couper KN, Blount DG and Riley EM: IL-10:
The master regulator of immunity to infection. J Immunol.
180:5771–5777. 2008.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Howard M, Muchamuel T, Andrade S and Menon
S: Interleukin 10 protects mice from lethal endotoxemia. J Exp Med.
177:1205–1208. 1993.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Calfee CS and Pugin J: The search for
diagnostic markers in sepsis. Am J Respir Crit Care Med. 186:2–4.
2012.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Benzaquen LR, Yu H and Rifai N: High
sensitivity c-reactive protein: An emerging role in cardiovascular
risk assessment. Crit Rev Clin Lab Sci. 39:459–497. 2002.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Gabay C and Kushner I: Acute-phase
proteins and other systemic responses to inflammation. N Engl J
Med. 340:448–454. 1999.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Senger DR, Galli SJ, Dvorak AM, Perruzzi
CA, Harvey VS and Dvorak HF: Tumor cells secrete a vascular
permeability factor that promotes accumulation of ascites fluid.
Science. 219:983–985. 1983.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Leung D, Cachianes G, Kuang W, Goeddel D
and Ferrara N: Vascular endothelial growth factor is a secreted
angiogenic mitogen. Science. 246:1306–1309. 1989.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Hotchkiss RS and Karl IE: The
pathophysiology and treatment of sepsis. N Engl J Med. 348:138–150.
2003.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Voelkel NF, Cool C, Taraceviene-Stewart L,
Geraci MW, Yeager M, Bull T, Kasper M and Tuder RM: Janus face of
vascular endothelial growth factor: The obligatory survival factor
for lung vascular endothelium controls precapillary artery
remodeling in severe pulmonary hypertension. Crit Care Med. 30 (5
Suppl):S251–S256. 2002.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Pickkers P, Sprong T, Eijk LV, Hoeven HVD,
Smits P and Deuren MV: Vascular endothelial growth factor is
increased during the first 48 hours of human septic shock and
correlates with vascular permeability. Shock. 24:508–512.
2005.PubMed/NCBI View Article : Google Scholar
|
|
70
|
van der Flier M, van Leeuwen HJ, van
Kessel KP, Kimpen JL, Hoepelman AI and Geelen SP: Plasma vascular
endothelial growth factor in severe sepsis. Shock. 23:35–38.
2005.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Yano K, Liaw PC, Mullington JM, Shih SC,
Okada H, Bodyak N, Kang PM, Toltl L, Belikoff B, Buras J, et al:
Vascular endothelial growth factor is an important determinant of
sepsis morbidity and mortality. J Exp Med. 203:1447–1458.
2006.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Thickett DR, Armstrong L, Christie SJ and
Millar AB: Vascular endothelial growth factor may contribute to
increased vascular permeability in acute respiratory distress
syndrome. Am J Respir Crit Care Med. 164:1601–1605. 2001.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Marshall JC, Vincent JL, Fink MP, Cook DJ,
Rubenfeld G, Foster D, Fisher CJ Jr, Faist E and Reinhart K:
Measures, markers, and mediators: Toward a staging system for
clinical sepsis. A report of the fifth toronto sepsis roundtable,
toronto, ontario, canada, october 25-26, 2000. Crit Care Med.
31:1560–1567. 2003.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Peters K, Unger RE, Brunner J and
Kirkpatrick CJ: Molecular basis of endothelial dysfunction in
sepsis. Cardiovasc Res. 60:49–57. 2003.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Lee W, Ku SK, Kim SW and Bae JS: Endocan
elicits severe vascular inflammatory responses in vitro and in
vivo. J Cell Physiol. 229:620–630. 2014.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Becker KL, Snider R and Nylen ES:
Procalcitonin assay in systemic inflammation, infection, and
sepsis: Clinical utility and limitations. Crit Care Med.
36:941–952. 2008.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Nakamura A, Wada H, Ikejiri M, Hatada T,
Sakurai H, Matsushima Y, Nishioka J, Maruyama K, Isaji S, Takeda T
and Nobori T: Efficacy of procalcitonin in the early diagnosis of
bacterial infections in a critical care unit. Shock. 31:586–591.
2009.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Çetinkaya M, Özkan H, Köksal N, Çelebi S
and Hacımustafaoğlu M: Comparison of serum amyloid A concentrations
with those of C-reactive protein and procalcitonin in diagnosis and
follow-up of neonatal sepsis in premature infants. J Perinatol.
29:225–231. 2008.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Kim KE and Han JY: Evaluation of the
clinical performance of an automated procalcitonin assay for the
quantitative detection of bloodstream infection. Korean J Lab Med.
30:153–159. 2010.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Ugarte H, Silva E, Mercan D, De Mendonca A
and Vincent JL: Procalcitonin used as a marker of infection in the
intensive care unit. Crit Care Med. 27:498–504. 1999.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Deis JN, Creech CB, Estrada CM and Abramo
TJ: Procalcitonin as a marker of severe bacterial infection in
children in the emergency department. Pediatr Emerg Care. 26:51–60.
2010.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Schneider CP, Yilmaz Y, Kleespies A, Jauch
KW and Hartl WH: Accuracy of procalcitonin for outcome prediction
in unselected postoperative critically ill patients. Shock.
31:568–573. 2009.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Amaral A, Opal SM and Vincent JL:
Coagulation in sepsis. Intensive Care Med. 30:1032–1040.
2004.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Fu Y, Jiang H, Li LX, Chen J, Niu Q and Li
RX: Correlation of coagulation indicators with inflammatory markers
for sepsis in the patients with hematological malignancies.
Zhongguo Shi Yan Xue Ye Xue Za Zhi. 22:1381–1385. 2014.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
85
|
Zhan ZG and Li CS: Prognostic value of
D-dimer in patients with sepsis in emergency department: A
prospective study. Zhongguo Wei Zhong Bing Ji Jiu Yi Xue.
24:135–139. 2012.PubMed/NCBI(In Chinese).
|
|
86
|
Rodelo JR, De la Rosa G, Valencia ML,
Ospina S, Arango CM, Gómez CI, García A, Nuñez E and Jaimes FA:
d-dimer is a significant prognostic factor in patients with
suspected infection and sepsis. Am J Emerg Med. 30:1991–1999.
2012.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Kono H, Asakawa M, Fujii H, Maki A,
Amemiya H, Yamamoto M, Matsuda M and Matsumoto Y: Edaravone, a
novel free radical scavenger, prevents liver injury and mortality
in rats administered endotoxin. J Pharmacol Exp Ther. 307:74–82.
2003.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Nagi MN, Alam K, Badary OA, al-Shabanah
OA, al-Sawaf HA and al-Bekairi AM: Thymoquinone protects against
carbon tetrachloride hepatotoxicity in mice via an antioxidant
mechanism. Biochem Mol Biol Int. 47:153–159. 1999.PubMed/NCBI View Article : Google Scholar
|