Introduction
Proliferative vitreoretinopathy (PVR) is a rare
ocular inflammatory disease that can result in vision loss or even
blindness (1,2). Treatment strategies for PVR has
drastically improved over the past decade owing to advances in
surgical techniques, such as gauge vitrectomy, scleral buckling, as
well as three-dimensional mediated ophthalmic surgery (3). However, PVR remains the most common
cause of recurring retinal detachment, occurring in 5-10% of all
cases (4). Therefore, it is of
great significance to understand the mechanism underlying PVR
development to devise novel therapeutic strategies.
PVR development is dependent on the proliferation
and migration of retinal pigment epithelial (RPE) cells, which
forms the main component of PVR membranes (5). Mounting evidence has demonstrated
that RPE cells can serve fibrotic roles in promoting PVR
development (6-8).
Mesenchymal-like RPE cells that underwent epithelial-to mesenchymal
transition (EMT) have enhanced capability of proliferation,
migration and invasion (9,10). This population of RPE cells is
associated with the progression of PVR (9,10).
Therefore, investigating the regulation of EMT in RPE cells may
provide a potential therapeutic target for PVR treatment.
Palmitic acid (PA) is a long-chain saturated fatty
acid found in palm oil that has multiple reported biological and
nutritional effects (11,12). Wang et al (13) previously reported that saturated PA
can induce myocardial inflammation by binding to the toll-like
receptor 4 accessory protein myeloid differentiation factor 2. In
colorectal cancer, PA can increase tumor cell proliferation in a
β2-adrenergic receptor-dependent manner (14). In addition, PA can inhibit
macrophage-mediated EMT in colorectal cancer (15). A previous study also demonstrated
that PA can protect RPE cells against 4-hydroxynonenal-mediated
stress and light-mediated retinal degeneration in mice (16). However, the function of PA in RPE
cells and the development of PVR remain poorly understood at
present.
MicroRNAs (miRNAs/miRs) belong to a group of small
non-coding RNAs that are involved in various biological processes,
including PVR (17,18). Accumulating evidence has
demonstrated that miRNAs can induce or inhibit the EMT process in
RPE cells (19,21). In addition, miRNAs, such as miR-182
and miR-34a, have been reported to modulate RPE cell proliferation
and migration (20,22). Consequently, augmenting
understanding into the miRNA-mediated regulation and function in
RPE cells can potentially facilitate the development of miRNA
manipulators to reverse the progression of PVR.
According to the aforementioned literature, miRNAs
and their associated mechanisms might play critical role in the
regulation of RPE cell activity during PVR. The present study aimed
to explore the underlying mechanism of PA mediated miR-124
dysregulation on the EMT in ARPE-19 cells. In addition, the
underlying target of miR-124 was investigated to further reveal the
biofunction of miR-124 in the EMT of ARPE-19 cells. According to
these investigations, we hope to provide more information in the
understanding and therapeutic insights in PVR diseases.
Materials and methods
Cell culture and treatment
The human RPE cell line ARPE-19 was purchased from
American Type Culture Collection. The cells were cultured in DMEM
supplemented with 10% FBS (both Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin (Thermo
Fisher Scientific, Inc.) in a 5% CO2 incubator at 37˚C.
PA was purchased from MilliporeSigma and dissolved in 100% ethanol.
The ARPE-19 cells were treated with PA at the indicated
concentrations (0, 50, 100 200, and 400 µM) at 37˚C for 48 h and
cells treated with equal volume of 100% ethanol was used as the
control group.
Transfection
miR-124 mimic (50 nM; 5'-CGUGUUCACAGCGGACCUUGAU-3'),
negative control mimic (NC mimic; 50 nM;
5'-AUUGGAACGAUACAGAGAAGAUU-3'), miR-124 inhibitor (50 nM;
5'-AUCAAGGUCCGCUGUGAACACG-3') and the NC inhibitor (50 nM;
5'-UUGUACUACACAAAAGUACUG-3') were purchased from Invitrogen (Thermo
Fisher Scientific, Inc.). For knocking down LIN7C in ARPE-19 cells,
short interfering (si)RNAs targeting LIN7C (100 nM; si-LIN7C #1,
5'-GGCTACTGTTGCTGCATTTGC-3'; si-LIN7C #2,
5'-GACCTAATACATTTCAAAACTTG-3'; and si-LIN7C #3,
5'-GGGAATTTGAGAAATATTTCATT-3' cloned into pLKO vector) and
non-targeting sequence, si-NC, (100 nM; 5'-TAAGGCTATGAAGAGATAC-3')
were obtained from Guangzhou RiboBio Co., Ltd. Overexpression
plasmid pcDNA3.1-LINC7C (100 nM) and pcDNA3.1 (control plasmid; 100
nM) were purchased from Shanghai GenePharma Co., Ltd. Transfection
was performed in ARPE-19 cells with 70% confluency using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's guidelines at
37˚C for 48 h. Following incubation at 37˚C for 48 h, cells were
harvested for the following experiments.
Reverse transcription-quantitative
(RT-qPCR)
RNA was extracted from the transfected/PA treated
ARPE-19 cells using AccuRef Cell/Tissue Total RNA isolation kit
(cat. no. AM0041; AccuRef Diagnostics; Applied StemCell, Inc.) and
reverse-transcribed into complementary DNA using PrimeScript™ RT
reagent kit (Takara) at 37˚C for 15 min, 85˚C for 5 sec and 4˚C for
15 min. qPCR was performed using SYBR® Green PCR Master
Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.) with the
following conditions: 95˚C for 10 min, 40 cycles of 95˚C for 10 sec
and 57˚C for 20 sec, and 72˚C for 15 sec. The relative expression
levels of miRNAs and genes were calculated using the
2-ΔΔCq method (23) and
GAPDH/U6 were used as the internal controls. The primers designed
by the Primer Premier 5 software (PREMIER) and used in the study
are listed in Table I.
 | Table IPrimers used in the present
study. |
Table I
Primers used in the present
study.
| Protein | Forward
(5'-3') | Reverse
(5'-3') |
|---|
| E-cadherin |
GCTCGCTGAACTCCTCTGA |
TCGCCGCCACCATACATA |
| ZO-1 |
CGGTCCTCTGAGCCTGTAAG |
GGATCTACATGCGACGACAA |
| α-SMA |
CCACTGCTGCTTCCTCTTC |
CGCCGACTCCATTCCAAT |
| Fibronectin |
GTGTCCTCCTTCCATCTTC |
CAGACTGTCGGTACTCACG |
| LIN7C |
AACGCTACTGCAAAGGCTACT |
TGAATCCAAGGCCCTCTTCTG |
| MRPS33 |
TTCTCTGCTCATCACACGGC |
GGCAAGGAGTTAGAGTTCCGTAT |
| PAGR1 |
GCGAAGAGGAGAGATCCGATG |
GTGGGCATGTGTGGTTTTTCC |
| TSPAN8 |
AGTGCCCCAGGAGCTATGA |
AGATTTCTGTATCCACGGACATTTA |
| NABP1 |
CGCGCCTGTCCCAATATGA |
TTGGTCACGCGTCCTATCTC |
| SPIN1 |
CTCCCTGATAGAGTTGCGACA |
AGACCATTCCCCTCCACTCA |
| AZI2 |
CTTCGAAGAAACCGGAAGCC |
GCATCCATGACAACCAGAAGC |
| NOA1 |
TTTCCTCTGCAGGTTGGGTT |
GGTGTATAGCCTCGGAGATGC |
| TXNRD1 |
CGCCGTAGGTCAGCTAAAGAT |
GAAGCAGGGCTCTGGAGTCT |
| GAPDH |
ACGGCAAGTTCAACGGCACAG |
GAAGACGCCAGTAGACTCCACGAC |
| miR-124 |
CGUGUUCACAGCGGACCUUGAU | Universal PCR
Reverse Primer (cat. no. B532451; Sangon Biotech Co., Ltd.) |
| miR-23b |
ATCACATTGCCAGGGATTACCAC | Universal PCR
Reverse Primer (cat. no. B532451; Sangon Biotech Co., Ltd.) |
| miR-221 |
AGCTACATTGTCTGCTGGGTTTC | Universal PCR
Reverse Primer (cat. no. B532451; Sangon Biotech Co., Ltd.) |
| miR-124 |
TAAGGCACGCGGTGAATGCCAA | Universal PCR
Reverse Primer (cat. no. B532451; Sangon Biotech Co., Ltd.) |
| U6 |
CGCTTCGGCAGCACATATACT |
GAATTTGCGTGTCATCCTTGC |
Western blotting
The cultured ARPE-19 cells were lysed using 1X CST
lysis buffer (Cell Signaling Technology, Inc.) supplemented with
the proteinase inhibitor cocktail (Pierce; Thermo Fisher
Scientific, Inc.). Total proteins were extracted and the protein
concentration was determined using a BCA protein quantification kit
(Pierce; Thermo Fisher Scientific, Inc.). Equal amounts of proteins
(25 µg per lane or 10 µg per lane for LIN7C overexpression assay)
were separated using 12% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes (MilliporeSigma). The membranes
were blocked with 5% non-fat milk at room temperature for 1 h and
then incubated with primary antibodies overnight at 4˚C. The
following primary antibodies from Abcam were used: Zonula occludens
protein 1 (ZO-1; 1:1,000; cat. no. ab216880), E-cadherin (1:10,000;
cat. no. ab40772), α-smooth muscle actin (SMA; 1:500; cat. no.
ab124964), and fibronectin (1:1,000; cat. no. ab2413). In addition,
LIN7C (1:1,000; cat. no. SAB2101347) was purchased from
Sigma-Aldrich (Merck KGaA). To control sample loading, the
membranes were stripped with the western blotting stripping buffer
(AccuRef Diagnostics; Applied StemCell, Inc.) and re-probed with
the anti-mouse GAPDH antibody from Abcam (1:10,000; cat. no.
ab181602) at room temperature for 1.5 h. The membranes were washed
and incubated with a HRP-conjugated goat anti-mouse (1:5,000; cat.
no. ab47827; Abcam) or anti-rabbit (1:5,000; cat. no. ab7090;
Abcam) secondary antibody at room temperature for 1 h and then
visualized using an enhanced chemiluminescence kit (Pierce; Thermo
Fisher Scientific, Inc.). With GAPDH as the internal control, the
relative expression of protein bands was quantified using Image J
Software (Version 1.5; National Institutes of Health).
Transwell assay
Cell migration was analyzed with a Transwell assay
using a Transwell chamber (8-µm pores; Corning, Inc.).
Transfected/PA-treated ARPE-19 cells were resuspended in a
serum-free DMEM and added into the upper chamber at a density of
2x104 per well. DMEM supplemented with 10% FBS was then
added into the lower chamber. After incubation for 24 h at 37˚C,
cells that migrated through the membrane were fixed by 4%
paraformaldehyde at room temperature for 15 min and stained with
0.1% crystal violet at room temperature for 15 min (Sigma-Aldrich;
Merck KGaA). The migrating cells were then imaged in a light
microscope at 200x and counted manually.
Prediction of miRNA targets
The targets of miR-124 were predicted by TargetScan
(https://www.targetscan.org/vert_72/),
miRDB (http://mirdb.org/index.html), and
microT (https://mrmicrot.imsi.athenarc.gr/?r=mrmicrot/index)
online Tools. Then, the shared targeted genes were screened using
the Venn analysis (https://bioinfogp.cnb.csic.es/tools/venny/) and
presented in a Venn diagram.
Luciferase reporter assay
The ARPE-19 cells were seeded into 24-well plates at
a density of 5.0x105 per well. Subsequently, 24 h later,
luciferase reporter vector psiCHECK2 (Promega Corporation)
containing the wild-type or mutant (0.25 µg each) 3'-untranslated
regions (UTR) of LIN7C, purchased from Guangzhou RiboBio Co., Ltd,
was transiently co-transfected with miR-124 mimics (100 nM) or NC
(100 nM) into ARPE-19 cells using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol at 37˚C for 24 h. At 24 h post-transfection, the relative
luciferase activity was analyzed using a Dual-Luciferase Reporter
Assay System with Renilla luciferase activity as the
internal control (Promega Corporation).
Prediction of miR-124 using
bioinformatics
The underlying targets of miR-124 were predicted
using TargetScan (targetscan.org/vert_72/), miRDB (http://mirdb.org/index.html), and microT (https://dianalab.e-ce.uth.gr/html/dianauniverse/index.php?r=microT_CDS)
online databases with their default parameters. Then, the shared
targets among these three databases were screened using Venn
analysis.
Statistical analysis
All experiments were performed in triplicate and the
results are presented as the mean ± standard deviation. Statistical
analysis was performed using GraphPad prism (version 8.0; GraphPad
Software, Inc.). Unpaired student's t-test or one-way
analysis of variance followed by Tukey's post hoc test was
performed to analyze the differences among ≥ two groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
PA treatment induces EMT and enhances
miR-124 expression in ARPE-19 cells
To investigate the function of PA in EMT and the
underlying mechanism, ARPE-19 cells were treated with PA at
different concentrations (0, 50, 100, 200 and 400 µM) for 48 h. The
results revealed that PA treatment decreased the protein expression
levels of E-cadherin and ZO-1 whilst increasing the protein
expression levels of fibronectin and α-SMA in a dose-dependent
manner (Fig. 1A). Similarly,
treatment with 200 µM PA significantly suppressed the mRNA
expression levels of E-cadherin and ZO-1 but increased those of
fibronectin and α-SMA in ARPE-19 cells compared with the control
(Fig. 1B). RT-qPCR found that
although miR-424, miR-221 and miR-23b expression was significantly
enhanced upon PA treatment, the increase in miR-124 expression was
the highest expression compared with that in the control group
(Fig. 1C). Taken together, these
data suggested that miR-124 may be involve in PA-induced EMT of
ARPE-19 cells; therefore, the present study focused on the role of
miR-124 in EMT and its regulation by PA treatment.
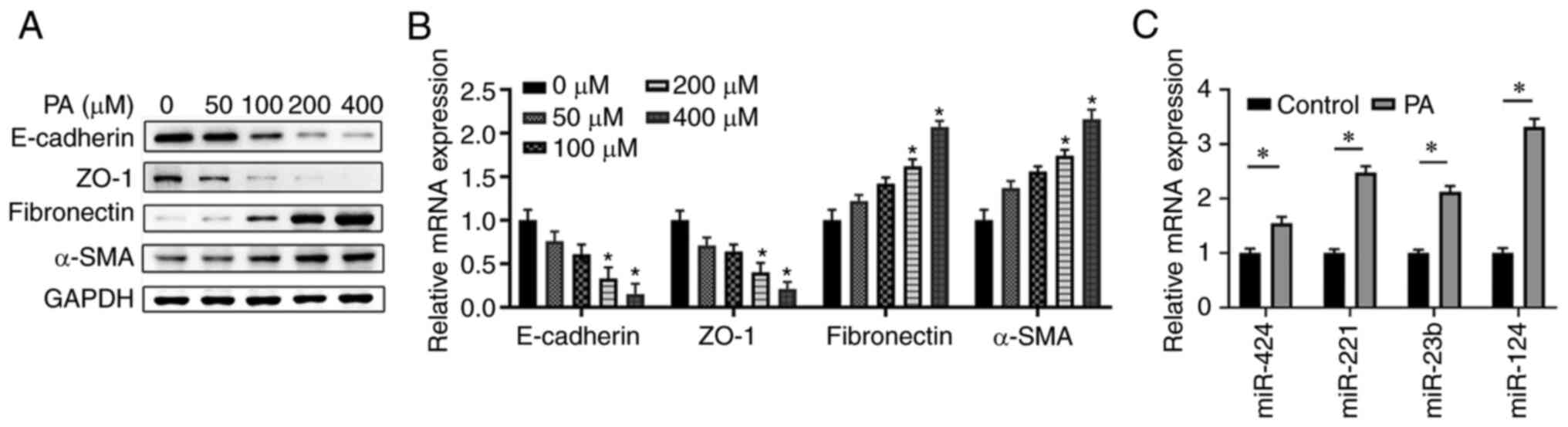 | Figure 1PA treatment induces EMT and
increases miR-124 expression in ARPE-19 cells. ARPE-19 cells were
treated with PA at different concentrations (0, 50, 100, 200 and
400 µM) for 48 h. (A) Protein and (B) mRNA expression levels of the
key molecules in EMT (E-cadherin, ZO-1, fibronectin and α-SMA) were
measured using western blotting and RT-qPCR. (C) ARPE-19 cells were
treated with 200 µM PA for 48 h, before the expression of miR-424,
miR-221, miR-23b and miR-124 were analyzed using RT-qPCR.
*P<0.05 vs. 0 µM/control group. PA, palmitic acid;
EMT, epithelial mesenchymal transition; miR, microRNA; RT-qPCR,
reverse transcription-quantitative PCR; ZO-1, zonula occludens
protein 1; α-SMA, α-smooth muscle actin. |
Overexpression of miR-124 enhances
PA-induced EMT and cell migration in ARPE-19 cells
Next, the possible function of miR-124 in EMT was
explored, where ARPE-19 cells were transfected with either the NC
or miR-124 mimics. Overexpression of miR-124, which was confirmed
by RT-qPCR compared with that in corresponding control cells
transfected with the NC mimic (Fig.
2A), mediated a similar effect to PA treatment. Specifically,
miR-124 mimic transfection markedly decreased the expression of
E-cadherin and ZO-1 but enhanced the expression of fibronectin and
α-SMA on both mRNA and protein levels compared with those in the
control group (Fig. 2B and
C). Subsequently, overexpression
of miR-124 together with PA treatment resulted in the lowest
expression levels of E-cadherin and ZO-1 but the highest expression
levels of fibronectin and α-SMA in ARPE-19 cells (Fig. 2B and C). All of these observations were
significant compared with those in the PA-only group (Fig. 2B and C). In addition, it was observed that
miR-124 mimics transfection or PA treatment markedly enhanced
ARPE-19 cell migration compared with that in the control (Fig. 2D and E). By contrast, the overexpression of
miR-124 significantly enhanced the cell migration of ARPE-19 cells
treated with PA compared with that in cells treated with PA only
(Fig. 2D and E). These findings suggest that miR-124
accelerated migration ability of ARPE-19 cells to promote
PA-induced EMT.
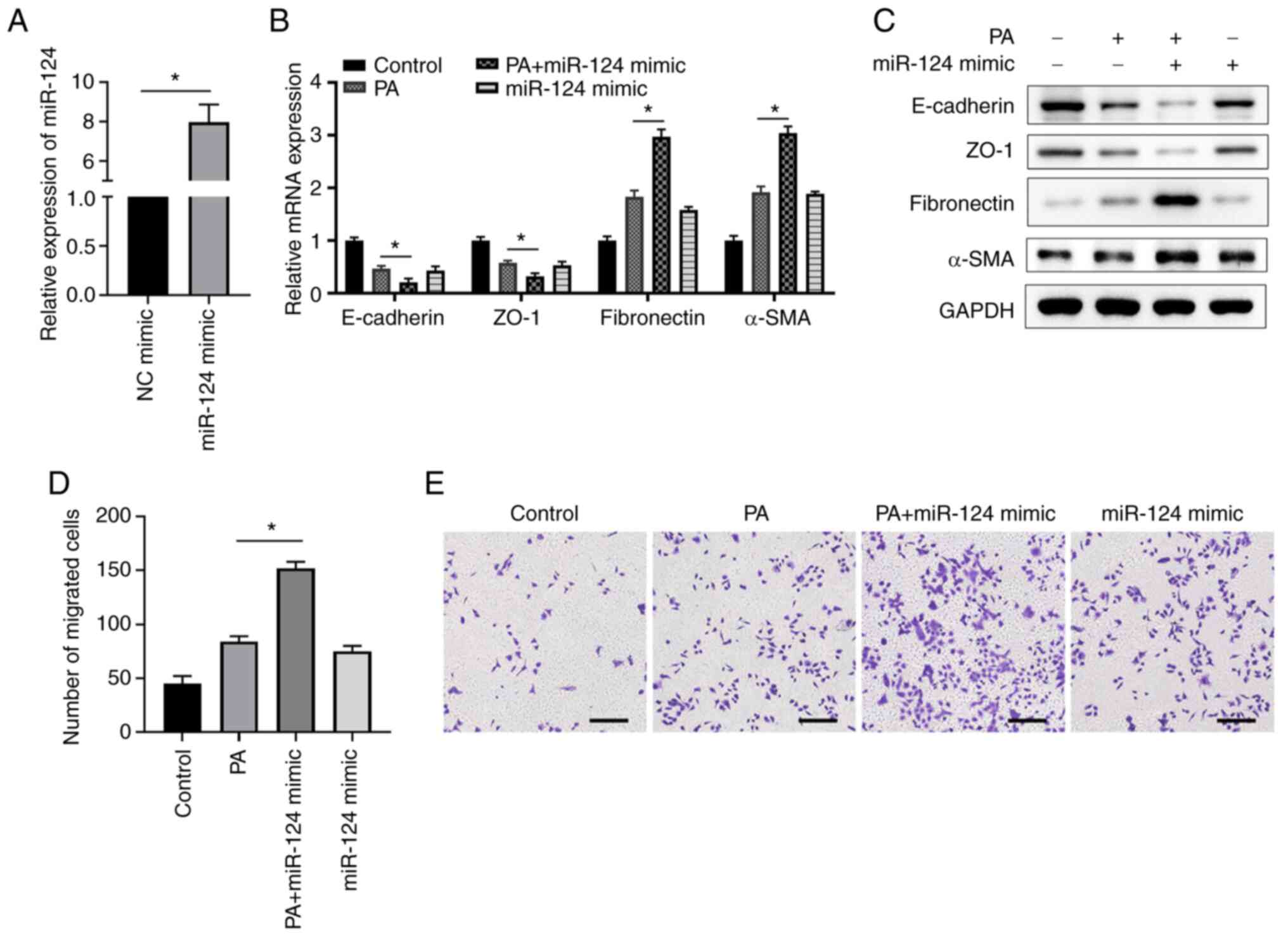 | Figure 2miR-124 overexpression enhances
PA-induced EMT and cell migration in ARPE-19 cells. (A) ARPE-19
cells were transfected with either the miR-124 mimics or NC, before
the expression levels of miR-124 were determined at 48 h after
transfection by RT-qPCR. (B) mRNA and (C) protein expression levels
of the key molecules in EMT (E-cadherin, ZO-1, fibronectin and
α-SMA) were measured using RT-qPCR and western blotting,
respectively. (D and E) ARPE-19 cells were transfected with miR-124
mimics or NC and/or cultured in the presence or absence of 200 µM
PA. Cells migration was analyzed using Transwell assay. (D) Number
of migrated cells in the different groups was calculated and (E)
representative images of Transwell cell migration assays are
presented; scale bar=200 µm. *P<0.05. PA, palmitic
acid; EMT, epithelial mesenchymal transition; miR, microRNA;
RT-qPCR, reverse transcription-quantitative PCR; ZO-1, zonula
occludens protein 1; α-SMA, α-smooth muscle actin; NC, negative
control. |
Knocking down miR-124 expression
suppresses PA-induced EMT and cell migration in ARPE-19 cells
Furthermore, a loss-of-function assay was performed
using a miR-124 inhibitor to investigate the function of miR-124.
Transfection with the miR-124 inhibitor significantly suppressed
the expression of miR-124 compared with that in cells transfected
with the NC inhibitor (Fig. 3A).
PA treatment decreased the expression of E-cadherin and ZO-1 whilst
enhancing the expression of fibronectin and α-SMA compared with
those in the control group, transfection with the miR-124 inhibitor
resulted in the opposite effects (Fig.
3B and C). In the PA + miR-124
inhibitor group, miR-124 inhibitor transfection was also found to
antagonize the effect of PA treatment, resulting in the E-cadherin,
ZO-1, fibronectin and α-SMA expression levels in ARPE-19 cells
returning to those comparable with the control group (Fig. 3B and C). In addition, transfection with the
miR-124 inhibitor was revealed to markedly inhibit ARPE-19 cell
migration compared with the control, whereas miR-124 inhibition
significantly reversed the PA-enhanced ARPE-19 cell migration
(Fig. 3D and E). These findings indicate that PA
enhances the EMT of ARPE-19 cells via miR-124 and inhibition of
miR-124 abrogated EMT of ARPE-19 cells induced by PA.
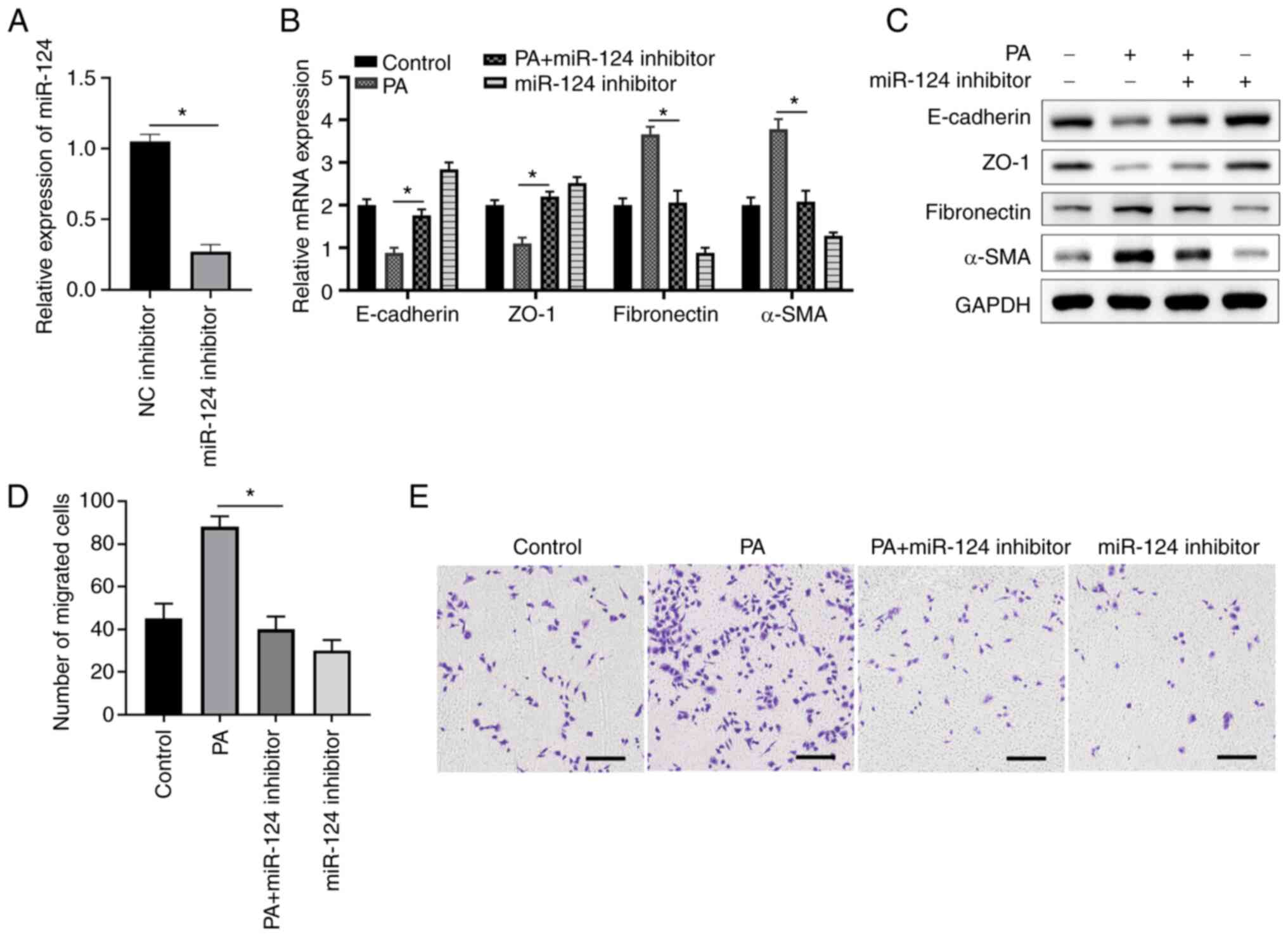 | Figure 3miR-124 knockdown suppresses
PA-induced EMT and cell migration in ARPE-19 cells. (A) ARPE-19
cells were transfected with either the miR-124 inhibitor or NC,
before the expression levels of miR-124 were determined at 48 h
post-transfection using RT-qPCR. (B) mRNA and (C) protein levels of
the key molecules in EMT (E-cadherin, ZO-1, fibronectin and α-SMA)
were determined using RT-qPCR and western blotting, respectively.
(D and E) ARPE-19 cells were transfected with miR-124 inhibitor or
NC and/or cultured in the presence or absence of 200 µM PA before
cell migration was analyzed using a Transwell assay. (D) Number of
migrated cells in different groups were calculated and (E)
representative images of Transwell cell migration assays are
presented; scale bar=200 µm. *P<0.05. PA, palmitic
acid; EMT, epithelial mesenchymal transition; miR, microRNA;
RT-qPCR, reverse transcription-quantitative PCR; ZO-1, zonula
occludens protein 1; α-SMA, α-smooth muscle actin; NC, negative
control. |
LIN7C is a direct target of miR-124 in
ARPE-19 cells
To explore the mechanism by which miR-124 regulates
EMT in ARPE-19 cells, bioinformatics analyses were performed to
predict the potential targets of miR-124 using TargetScan, miRDB,
and micro-T (Fig. 4A). A total of
nine potential targets were predicted by all three tools (Fig. 4A). ARPE-19 cells were then
transfected with miR-124 mimics or NC, before the relative
expression of these potential target genes were analyzed 48 h
later. As presented in Fig. 4B,
transfection of miR-124 mimics significantly suppressed the
expression of LIN7C compared with that in the NC group. There were
also complementary binding sequences between miR-124 and the 3'-UTR
of LIN7C (Fig. 4C). Luciferase
reporter assay subsequently confirmed that miR-124 mimic could
significantly decrease the luciferase activity of LIN7C-Wt but had
no effect on the activity of LIN7C-Mut, suggesting a direct
interaction between miR-124 and LIN7C (Fig. 4C). PA treatment was also confirmed
to suppress LIN7C expression in a dose-dependent manner, with this
reduction reaching significance at 200 and 400 µM compared with
that in the 0 µM group (Fig. 4D).
Furthermore, inhibition of miR-124 expression markedly enhanced the
expression of LIN7C in ARPE-19 cells compared with the NC inhibitor
group, whilst miR-124 inhibitor transfection reversed the
suppressive effects of PA treatment on LIN7C expression (Fig. 4E and F). All of these findings indicate that
LIN7C is a target of miR-124 in ARPE-19 cells.
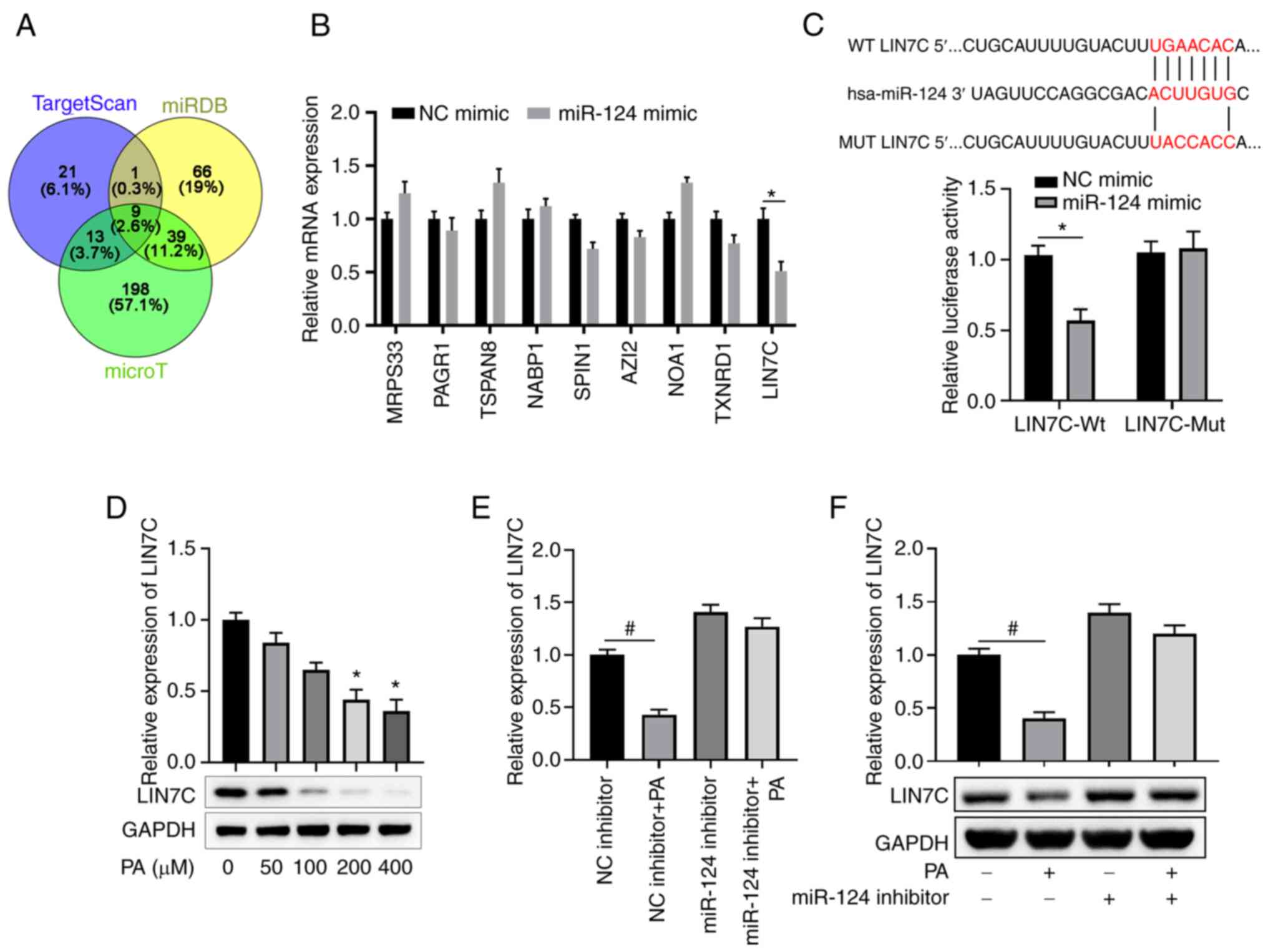 | Figure 4LIN7C is a direct target of miR-124
in ARPE-19 cells. (A) Venn diagram presents the numbers of
candidate mRNA targets of miR-124 predicted using three public
algorithms TargetScan, miRDB and micro-T. (B) ARPE-19 cells were
transfected with NC or miR-124 mimics. The mRNA expression levels
of the indicated candidate targets of miR-124 were analyzed using
RT-qPCR 48 h later. (C) Diagrams show the putative binding sites
between miR-124 and corresponding WT or MUT sites of LIN7C 3'UTR.
ARPE-19 cells were co-transfected with reporter vector containing
WT or MUT LIN7C 3'-UTR, together with NC or miR-124 mimics. Dual
luciferase activity was detected in ARPE-19 cells at 48 h post
transfection. (D) Protein expression levels of LIN7C in ARPE-19
cells treated with PA at the indicated concentrations were
determined by western blotting. (E and F) ARPE-19 cells were
transfected with NC or miR-124 inhibitor with or without 200 µM PA
treatment. At 48 h later, (E) mRNA and (F) expression protein
levels of LIN7C in each treatment group were measured using RT-qPCR
and western blot assays, respectively. *P<0.05 vs. NC
mimic; #P<0.05 vs. NC inhibitor. LIN7C, lin-7 homolog
C; miR, microRNA; NC, negative control; RT-qPCR, reverse
transcription-quantitative PCR; WT, wild-type; MUT, mutant; UTR,
untranslated region; PA, palmitic acid. |
LIN7C regulates EMT and cell migration
in ARPE-19 cells
To study the effect of LIN7C on EMT, the
overexpression vector for LIN7C was constructed and transfected
into ARPE-19 cells to overexpress LIN7C, which was successful
compared with that in cells transfected with the empty pcDNA3.1
vector (Fig. 5A). By contrast,
siRNAs targeting LIN7C were transfected into ARPE-19 cells to
silence the expression of LIN7C. The result suggested that si-LIN7C
2# and 3# significantly decrease LIN7C expression, and si-LIN7C 3#
presented a more effective effect than si-LIN7C 2# (Fig. 5B). Therefore, si-LIN7C 2# was
selected and used for the following experiments. The overexpression
of LIN7C was demonstrated to suppress EMT, with markedly enhanced
expression levels of E-cadherin/ZO-1 and markedly decreased
expression levels of fibronectin/α-SMA compared with those in the
empty vector control group (Fig.
5C and D). Knockdown of LIN7C
significantly enhanced EMT, namely significantly inhibiting
expression of E-cadherin and ZO-1 but enhancing expression of
fibronectin and α-SMA compared with the si-NC group (Fig. 5C and D). Furthermore, overexpression of LIN7C
was demonstrated to markedly inhibit cell migration compared with
that in the empty vector group, whilst its knockdown significantly
enhanced cell migration compared with that in the si-NC group
(Fig. 5E and F). Taken together, these results indicate
that silencing LIN7C significantly promoted migration of ARPE-19
cells.
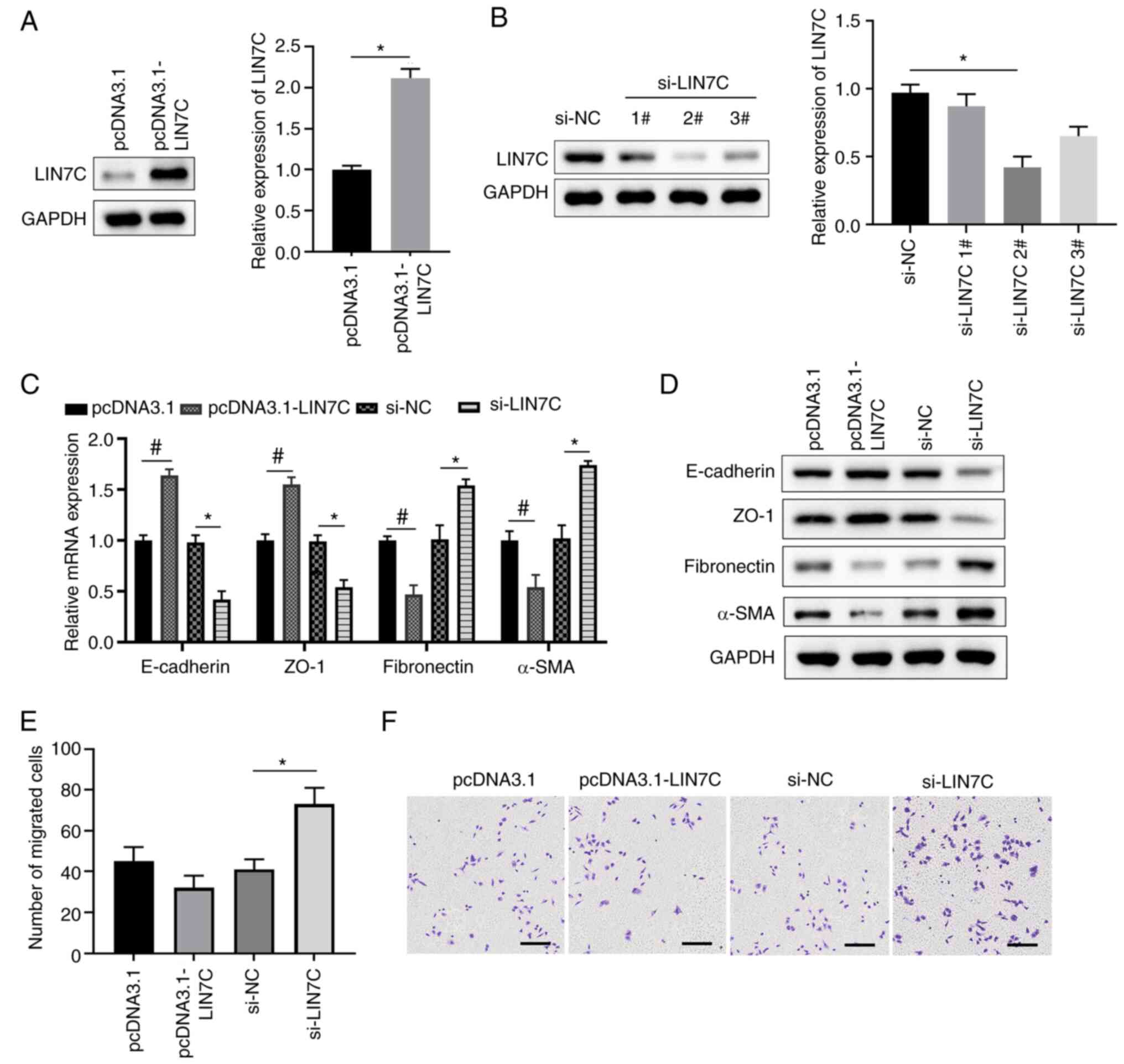 | Figure 5LIN7C regulates EMT and cell
migration in ARPE-19 cells. (A) ARPE-19 cells were transfected with
control empty plasmids or LIN7C-overexpressing plasmids. Protein
and mRNA expression levels of LIN7C in ARPE-19 cells were
determined using western blotting and RT-qPCR, respectively. (B)
ARPE-19 cells were transfected with si-NC or LIN7C-specific siRNA
oligos. Protein and mRNA expression levels of LIN7C in ARPE-19
cells were then determined using western blotting and RT-qPCR
assays, respectively. (C-F) ARPE-19 cells were transfected with
control empty plasmids or LIN7C-overexpressing plasmids, si-NC or
si-LIN7C #2. Expression levels of the key molecules in EMT
(E-cadherin, ZO-1, fibronectin and α-SMA) were determined using (C)
RT-qPCR for mRNA and (D) western blotting for proteins. At 48 h
post-transfection, ARPE-19 cells were cultured in Transwell
chambers and migratory cells were stained with crystal-violet. (E)
The number of migrated cells in one field of view in each group is
quantified and (F) representative images of the Transwell cell
migration assays are presented; scale bar, 200 µm.
*P<0.05 and #P<0.05. EMT, epithelial
mesenchymal transition; RT-qPCR, reverse transcription-quantitative
PCR; ZO-1, zonula occludens protein 1; α-SMA, α-smooth muscle
actin; NC, negative control; LIN7C, lin-7 homolog C; NC, negative
control; si, short-interfering. |
Overexpression of LIN7C abrogates
PA-induced EMT and cell migration in ARPE-19 cells
To further investigate the role of LIN7C in EMT, the
APRE-19 cells were transfected either with the control vector or
the overexpression vector for LIN7C, with or without PA treatment.
PA treatment was found to induce EMT compared with the pcDNA3.1
group, as demonstrated by decreased expression levels of E-cadherin
and ZO-1 and enhanced expression levels of fibronectin and α-SMA
(Fig. 6A and B). By contrast, overexpression of LIN7C
markedly reversed the PA-induced EMT in ARPE-19 cells compared with
that in the PA-only group (Fig. 6A
and B). Overexpression of LIN7C
also significantly suppressed cell migration compared with that in
the empty vector group, whereas PA treatment produced the opposite
effect. However, LIN7C overexpression significantly reduced
PA-induced cell migration in ARPE-19 cells compared with the PA
group (Fig. 6C and D). In addition, overexpression of miR-124
markedly reversed the upregulation of E-cadherin and ZO-1 and the
downregulation of fibronectin and α-SMA previously mediated by
LIN7C overexpression, which were in turn enhanced by PA treatment
(Fig. 6E and F). In addition, overexpression of miR-124
significantly attenuated the inhibitory effects of LIN7C
overexpression on the migration of ARPE-19 cells, which were
significantly potentiated by PA treatment (Fig. 6G and H). Collectively, these data suggest that
LIN7C could antagonize the EMT promotion function of PA in ARPE-19
cells.
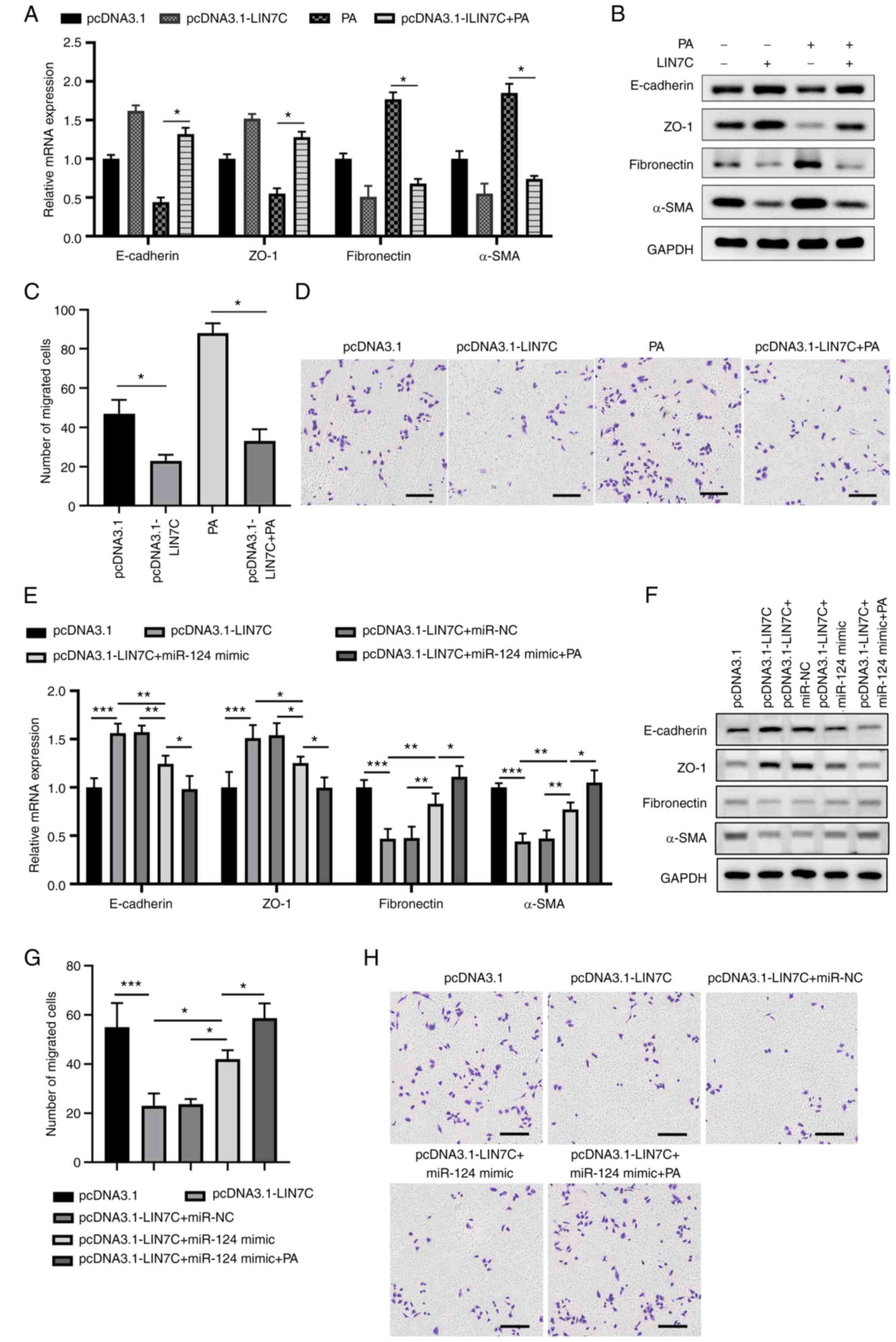 | Figure 6Overexpression of LIN7C abrogates
PA-induced EMT and cell migration in ARPE-19 cells. ARPE-19 cells
were transfected with control vector, pLIN7C and/or treated with or
without 200 µM PA. EMT-associated markers and migration in the
cells were determined 48 h later. Expression levels of key
molecules in EMT (E-cadherin, ZO-1, fibronectin and α-SMA) were
determined using (A) RT-qPCR for mRNA and (B) western blotting for
proteins. (C and D) ARPE-19 cells were transfected with control or
pLIN7C and/or cultured in the presence or absence of 200 µM PA.
Cell migration was analyzed using a Transwell assay. (C) Number of
migrated cells in different groups were calculated and (D)
representative images from Transwell cell migration assays are
presented; scale bar, 200 µm. (E-H) ARPE-19 cells were transfected
with pcDNA3.1 or pcDNA3.1-LIN7 and miR-NC or miR-124 followed by
200 µM PA treatment, before EMT-associated marker expression and
cell migration were measured 48 h later. Expression levels of the
key molecules in EMT (E-cadherin, ZO-1, fibronectin and α-SMA) were
determined using (E) RT-qPCR for mRNA and (F) western blotting for
proteins. (G) Number of migrated cells in the different groups were
calculated and (H) representative images of Transwell cell
migration assays are presented; scale bar, 200 µm.
*P<0.05, **P<0.01,
***P<0.001. PA, palmitic acid; EMT, epithelial
mesenchymal transition; miR, microRNA; RT-qPCR, reverse
transcription-quantitative PCR; ZO-1, zonula occludens protein 1;
α-SMA, α-smooth muscle actin; NC, negative control; LIN7C, lin-7
homolog C. |
Discussion
Accumulating evidence has demonstrated that cell
proliferation, migration and EMT of RPE cells are important for PVR
development (24,25). Furthermore, blocking EMT in RPE
cells as been previously shown to prevent PVR pathogenesis
(26). The present study revealed
that PA treatment induced EMT and enhanced miR-124 expression in
ARPE-19 cells. miR-124 overexpression synergistically enhanced the
PA-induced EMT and cell migration in ARPE-19 cells. Furthermore,
LIN7C was established as a direct target of miR-124, whereby the
overexpression of LIN7C abrogated the PA-induced EMT in ARPE-19
cells. Therefore, the PA/miR-124/LIN7C functional cascade was
suggested to mediate EMT and cell migration in RPE cells.
PA has been reported to regulate miRNA expression,
such as miR-221 and miR-130a (27,28).
Therefore, the present study performed a PA-associated miRNA
microarray to analyze the differentially expressed miRNAs in
ARPE-19 cells with or without PA treatment. The present study
demonstrated that the expression of multiple miRNAs have been
demonstrated to be regulated by PA treatment, where miR-124 was
among the most upregulated miRNAs in ARPE-19 cells. Chu-Tan et
al (29) previously revealed
that the dysregulated expression of miR-124 is associated with
retinal inflammation and photoreceptor death, such that miR-124
overexpression can alleviate retinal degeneration via regulating
CCL2. The regulatory effects of miR-124 on EMT has been
investigated by previous studies on triple-negative breast cancer,
nasopharyngeal carcinoma, non-small cell lung cancer and clear cell
renal carcinoma (30-33).
Consistently, the present study revealed that the miR-124 mimics
enhanced EMT in ARPE-19 cells whilst the miR-124 inhibitor
suppressed PA-induced EMT. In addition, miR-124 enhanced PA-induced
ARPE-19 cell migration, which may further aggravate PVR
progression.
miR-124 has been studied as a potential tumor
suppressor in various tumors (30,31,34).
In triple-negative breast cancer and clear cell renal carcinoma,
miR-124 was found to negatively regulate the expression of zinc
finger E-box-binding homeobox 2 (30,31,32).
Calpain small subunit 4 has been identified to be a target of
miR-124, whilst long non-coding RNA metastasis-associated lung
adenocarcinoma transcript 1 was documented to sponge miR-124
expression in nasopharyngeal carcinoma cells (31). The present study revealed that
miR-124 regulated LIN7C expression by directly binding to the
3'-UTR of LIN7C. LIN7C has been reported to be as a dynamic marker
for polarity maturation in the zebra fish retinal epithelium
(35). However, to the best of our
knowledge, the function of LIN7C in PVR development has not been
studied previously. The present study hypothesized that LIN7C
regulated EMT and cell migration in ARPE-19 cells. Overexpression
of LIN7C antagonized the effect of PA treatment on EMT induction.
Therefore, the miR-124/LIN7C axis, in conjunction with PA
treatment, may serve a key role in the EMT and pathogenesis of PVR.
Nevertheless, how LIN7C modulated the EMT process and the
underlying mechanism of miR-124/LIN7C in PVR warrant further
investigation.
In summary, the present study demonstrated that PA
treatment induced EMT and promoted the migration of RPE cells by
upregulating miR-124 expression whilst inhibiting LIN7C expression.
These findings suggest that targeting miR-124/LIN7C may prove to be
useful in preventing and treating PVR.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study available from the corresponding author on reasonable
request.
Authors' contributions
XDH and LGL conceived and designed the experiments.
XGJ and MY performed the experiments. WJC analyzed and interpreted
the data. XDH and LGL confirm the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pastor JC: Proliferative
vitreoretinopathy: An overview. Surv Ophthalmol. 43:3–18.
1998.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Di Lauro S, Kadhim MR, Charteris DG and
Pastor JC: Classifications for proliferative vitreoretinopathy
(PVR): An analysis of their use in publications over the last 15
years. J Ophthalmol. 2016(7807596)2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Coffee RE, Jiang L and Rahman SA:
Proliferative vitreoretinopathy: Advances in surgical management.
Int Ophthalmol Clin. 54:91–109. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Idrees S, Sridhar J and Kuriya AE:
Proliferative vitreoretinopathy: A review. Int Ophthalmol Clin.
59:221–240. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Charteris DG: Proliferative
vitreoretinopathy: Pathobiology, surgical management, and
adjunctive treatment. Br J Ophthalmol. 79:953–960. 1995.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Lee SC, Kwon OW, Seong GJ, Kim SH, Ahn JE
and Kay ED: Epitheliomesenchymal transdifferentiation of cultured
RPE cells. Ophthalmic Res. 33:80–86. 2001.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Mudhar HS: A brief review of the
histopathology of proliferative vitreoretinopathy (PVR). Eye
(Lond). 34:246–250. 2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zou H, Shan C, Ma L, Liu J, Yang N and
Zhao J: Polarity and epithelial-mesenchymal transition of retinal
pigment epithelial cells in proliferative vitreoretinopathy. Peer
J. 8(e10136)2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
He H, Kuriyan AE, Su CW, Mahabole M, Zhang
Y, Zhu YT, Flynn HW, Parel JM and Tseng SC: Inhibition of
proliferation and epithelial mesenchymal transition in retinal
pigment epithelial cells by heavy chain-hyaluronan/pentraxin 3. Sci
Rep. 7(43736)2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Tamiya S, Liu L and Kaplan HJ:
Epithelial-mesenchymal transition and proliferation of retinal
pigment epithelial cells initiated upon loss of cell-cell contact.
Invest Ophthalmol Vis Sci. 51:2755–2763. 2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Carta G, Murru E, Banni S and Manca C:
Palmitic acid: Physiological role, metabolism and nutritional
implications. Front Physiol. 8(902)2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Mancini A, Imperlini E, Nigro E,
Montagnese C, Daniele A, Orrù S and Buono P: Biological and
nutritional properties of palm oil and palmitic acid: Effects on
health. Molecules. 20:17339–17361. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wang Y, Qian Y, Fang Q, Zhong P, Li W,
Wang L, Fu W, Zhang Y, Xu Z, Li X and Liang G: Saturated palmitic
acid induces myocardial inflammatory injuries through direct
binding to TLR4 accessory protein MD2. Nat Commun.
8(13997)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Fatima S, Hu X, Huang C, Zhang W, Cai J,
Huang M, Gong RH, Chen M, Ho AHM, Su T, et al: High-fat diet
feeding and palmitic acid increase CRC growth in β2AR-dependent
manner. Cell Death Dis. 10(711)2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
de Araujo Junior RF, Eich C, Jorquera C,
Schomann T, Baldazzi F, Chan AB and Cruz LJ: Ceramide and palmitic
acid inhibit macrophage-mediated epithelial-mesenchymal transition
in colorectal cancer. Mol Cell Biochem. 468:153–168.
2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Gutierrez MA, Davis SS, Rosko A, Nguyen
SM, Mitchell KP, Mateen S, Neves J, Garcia TY, Mooney S, Perdew GH,
et al: A novel AhR ligand, 2AI, protects the retina from
environmental stress. Sci Rep. 6(29025)2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Osada H and Takahashi T: MicroRNAs in
biological processes and carcinogenesis. Carcinogenesis. 28:2–12.
2007.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kaneko H and Terasaki H: Biological
involvement of microRNAs in proliferative vitreoretinopathy. Transl
Vis Sci Technol. 6(5)2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Usui-Ouchi A, Ouchi Y, Kiyokawa M, Sakuma
T, Ito R and Ebihara N: Upregulation of Mir-21 levels in the
vitreous humor is associated with development of proliferative
vitreoretinal disease. PLoS One. 11(e0158043)2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang L, Dong F, Reinach PS, He D, Zhao X,
Chen X, Hu DN and Yan D: MicroRNA-182 suppresses HGF/SF-induced
increases in retinal pigment epithelial cell proliferation and
migration through targeting c-Met. PLoS One.
11(e0167684)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Takayama K, Kaneko H, Hwang SJ, Ye F,
Higuchi A, Tsunekawa T, Matsuura T, Iwase T, Asami T, Ito Y, et al:
Increased ocular levels of microRNA-148a in cases of retinal
detachment promote epithelial-mesenchymal transition. Invest
Ophthalmol Vis Sci. 57:2699–2705. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Hou Q, Zhou L, Tang J, Ma N, Xu A, Tang J,
Zheng D, Chen X, Chen F, Dong XD and Tu L: LGR4 is a direct target
of microRNA-34a and modulates the proliferation and migration of
retinal pigment epithelial ARPE-19 cells. PLoS One.
11(e0168320)2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Li X, Zhao M and He S: RPE
epithelial-mesenchymal transition plays a critical role in the
pathogenesis of proliferative vitreoretinopathy. Ann Transl Med.
8(263)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Tamiya S and Kaplan HJ: Role of
epithelial-mesenchymal transition in proliferative
vitreoretinopathy. Exp Eye Res. 142:26–31. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Nagasaka Y, Kaneko H, Ye F, Kachi S, Asami
T, Kato S, Takayama K, Hwang SJ, Kataoka K, Shimizu H, et al: Role
of Caveolin-1 for blocking the epithelial-mesenchymal transition in
proliferative vitreoretinopathy. Invest Ophthalmol Vis Sci.
58:221–229. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Huang F, Chen J, Wang J, Zhu P and Lin W:
Palmitic acid induces microRNA-221 expression to decrease glucose
uptake in HepG2 cells via the PI3K/AKT/GLUT4 pathway. Biomed Res
Int. 2019(8171989)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gu X, Wang XQ, Lin MJ, Liang H, Fan SY,
Wang L, Yan X, Liu W and Shen FX: Molecular interplay between
microRNA-130a and PTEN in palmitic acid-mediated impaired function
of endothelial progenitor cells: Effects of metformin. Int J Mol
Med. 43:2187–2198. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chu-Tan JA, Rutar M, Saxena K, Aggio-Bruce
R, Essex RW, Valter K, Jiao H, Fernando N, Wooff Y, Madigan MC, et
al: MicroRNA-124 dysregulation is associated with retinal
inflammation and photoreceptor death in the degenerating retina.
Invest Ophthalmol Vis Sci. 59:4094–4105. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ji H, Sang M, Liu F, Ai N and Geng C:
miR-124 regulates EMT based on ZEB2 target to inhibit invasion and
metastasis in triple-negative breast cancer. Pathol Res Pract.
215:697–704. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Shi B, Wang Y and Yin F:
MALAT1/miR-124/Capn4 axis regulates proliferation, invasion and EMT
in nasopharyngeal carcinoma cells. Cancer Biol Ther. 18:792–800.
2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Chen J, Zhong Y and Li L: miR-124 and
miR-203 synergistically inactivate EMT pathway via coregulation of
ZEB2 in clear cell renal cell carcinoma (ccRCC). J Transl Med.
18(69)2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wu J, Weng Y, He F, Liang D and Cai L:
LncRNA MALAT-1 competitively regulates miR-124 to promote EMT and
development of non-small-cell lung cancer. Anticancer Drugs.
29:628–636. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Li Y, Yan J, Wang Y, Wang C, Zhang C and
Li G: LINC00240 promotes gastric cancer cell proliferation,
migration and EMT via the miR-124-3p / DNMT3B axis. Cell Biochem
Funct. 38:1079–1088. 2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Luz M and Knust E: Fluorescently tagged
Lin7c is a dynamic marker for polarity maturation in the zebrafish
retinal epithelium. Biol Open. 2:867–871. 2013.PubMed/NCBI View Article : Google Scholar
|














