Introduction
In recent years, the incidence of type 2 diabetes
mellitus (T2DM) has gradually increased (1). T2DM-associated disability and
fatality rates are increasing and are mainly caused due to its
complications, including diabetic eye disease, diabetic nephropathy
and diabetic peripheral nephropathy (2,3).
These issues are considered to be vascular complications of T2DM.
At present, the main clinical treatment strategies for diabetic
vascular complications include the use of antihypertensive and
hypoglycemic drugs to prevent and delay their development (4). For example, the aldose reductase
inhibitor epalrestat is used to control blood glucose levels
(5). Renin-angiotensin system
inhibitors, including angiotensin-converting enzyme inhibitors or
angiotensin receptor blockers, are commonly used first-line
therapeutic agents for the treatment of diabetic nephropathy
(6). Losartan and enalapril are
used to reduce the progression of diabetic retinopathy (7).
Endothelial dysfunction serves an important role in
diabetic vascular complications and mitochondrial dysfunction is
the main cause of endothelial dysfunction in patients with diabetes
(8,9). Therefore, previous study have
addressed the treatment of T2DM and its complications by improving
endothelial function via the enhancement of mitochondrial
biosynthesis, maintenance of mitochondrial function and the
mitigation of mitochondrial dysfunction (10).
Mitochondrial biogenesis serves a crucial role in
cellular mitochondrial function (11). Mitochondrial biogenesis is caused
by the initiation of mitochondrial DNA (mtDNA) replication, and the
expression of mitochondrial proteins encoded by the nuclear and
mitochondrial genomes (12).
Peroxisome proliferator-activated receptor γ coactivator 1
(PGC-1)α/β are considered to be the central regulators of
mitochondrial biogenesis (13).
PGC-1α transactivates nuclear respiratory factor 1 (NRF1), which
activates mitochondrial transcription factor A (TFAM).
Subsequently, TFAM regulates the transcription and replication of
mtDNA. The 5'AMP-activated protein kinase (AMPK)/PGC-1α signaling
pathway is closely associated with mitochondrial biogenesis
(14).
Salvianolic acid A (SAA) is one of the main active
water-soluble constituents of the plant, Salvia miltiorrhiza
(15). SAA has been demonstrated
to serve an important role in the treatment of vascular
complications of T2DM (16). It
has previously been reported that SAA may improve endothelial
vascular function (17); however,
the mechanism by which SAA ameliorates endothelial dysfunction
remains unclear. Qiang et al (17) demonstrated that SAA can improve
mitochondrial function of liver cells and skeletal muscle cells in
T2DM mice via the activation of AMPK. Therefore, the aim of the
present study was to investigate whether SAA could promote the
mitochondrial biosynthesis of human umbilical vein endothelial
cells (HUVECs) and improve mitochondrial function via activation of
the AMPK/PGC-1α signaling pathway, thereby improving endothelial
vascular function and ultimately treating diabetic
complications.
Materials and methods
Cell culture
Primary HUVECs were purchased from The Cell Bank of
Type Culture Collection of The Chinese Academy of Sciences. The
cellular experiments were performed using 1st-5th generation cells.
The cells were cultured in RPMI-1640 medium (Procell Life Science
& Technology Co., Ltd.) containing 10% FBS (Biological
Industries) and 1% penicillin-streptomycin (Procell Life Science
& Technology Co., Ltd.). The cells were cultured in an
incubator at 37˚C in a 5% CO2 humidifying
atmosphere.
Drug treatment
SAA (purity, 98%) was provided as a lyophilized
powder by The Institute of Materia Medica, Chinese Academy of
Medical Sciences. The cells were cultured in petri dishes in a
medium of 37˚C humidified with 5% CO2. Following their
attachment to the wells, different concentrations of SAA
(10-3, 10-2 and 10-1 µM) were
added and incubated at 37˚C for 24 h. To determine the role of SAA
in mitochondrial biogenesis, the inhibitor group was treated with
2.5 µM AMPK inhibitor compound C (MedChemExpress) at 37˚C for 30
min prior to treatment with 10-2 µM SAA. The AMPK
activator 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR; 0.5
mM; MedChemExpress) at 37˚C for 30 min as a positive control.
Mitochondrial quantification
Cells were treated at the above dosing concentration
at 37˚C for 24 h and were subsequently examined via electron
microscopy. Briefly, the cells were fixed in 2.5% glutaraldehyde
(Beijing Solarbio Science & Technology Co., Ltd.) for 4 h and
then rinsed with phosphate buffer and immobilized with 2% osmium
tetroxide at 4˚C for 1.5 h. Dehydrated with a series of graded
ethanol to 70% ethanol, in a saturated uranium acetate solution
prepared with 70% ethanol or acetone, dyed for 2 h or more, cleaned
with epoxy propane, embedded in Spurrs low viscosity resin and
sectioned. Thin sections with thickness of 70 nm were observed via
transmission electron microscopy (Olympus Corporation). Image-Pro
Plus (version 5.0; Media Cybernetics, Inc.) software was used to
quantify the number of mitochondria in each image and the data were
expressed as mitochondrial density (number of
mitochondria/cytoplasmic region). At least 20 cells were assessed
per treatment.
Adenosine triphosphate (ATP)
assay
The ATP levels of cells were determined via a
colorimetric method using an ATP test kit (Elabscience
Biotechnology, Inc.). Creatine kinase catalyzes the reaction of
creatine and ATP to generate creatine phosphate; creatine phosphate
content is detected using a colorimetric method to reflect the ATP
content (18). The cells were
cultured in 6-well plates in 37˚C humidified with 5% CO2
until adherent. Following adherence, the cells were treated with
different concentrations of SAA (10-3, 10-2
and 10-1 µM) and incubated at 37˚C for 24 h. The
collected cells were added with 0.3 ml boiling deionized water for
each 106 cells, tightly covered and mixed, then inserted
into the floating float and placed in the boiling water bath for 10
min, then mixed in vortex for 1 min at room temperature of 1,000 x
g and centrifuged for 10 min. The ATP levels of the supernatant
were assessed according to the instructions provided by the
manufacturer. The optical density was measured at 636 nm using a
microplate reader and the ATP levels were determined using the
manufacturer's formula.
Western blotting
Following treatment with SAA, the cells were lysed
using RIPA buffer (Beyotime Institute of Biotechnology) containing
a 1% protease inhibitor cocktail (MedChemExpress) on ice for 15
min. The protein concentration was assessed using a BCA kit
(Elabscience Biotechnology, Inc.). Protein samples (20 µg were
separated with 10% SDS-PAGE (Epizyme). Following separation, the
protein was transferred to a PVDF membrane (Beijing Solarbio
Science & Technology Co., Ltd) and blocked with 5% skimmed milk
at room temperature for 1.5 h. After blocking, the membrane was
incubated with primary antibodies against the following: AMPK
(1:1,000; cat. no. 5831; Cell Signaling Technology, Inc.),
phosphorylated (p)-AMPK (1:1,000; cat. no. 2535; Cell Signaling
Technology, Inc.), acetyl-CoA carboxylase (ACC; 1:1,000; cat. no.
3676; Cell Signaling Technology, Inc.), p-ACC (1:1,000; cat. no.
11818; Cell Signaling Technology, Inc.), PGC-1α (1:1,000; cat. no.
2178; Cell Signaling Technology, Inc.), NRF1 (1:1,000; cat. no.
46743; Cell Signaling Technology, Inc.), TFAM (1:1,000; cat. no.
8076; Cell Signaling Technology, Inc.), complex Ⅲ (1:1,000; cat.
no. ab182330; Abcam), complex Ⅳ (1:1,000; cat. no. ab14705; Abcam)
and GAPDH (1:2,000; cat. no. E-AB-20059; Elabscience Biotechnology,
Inc.) at 4˚C overnight. Subsequently, the membrane was incubated
with horseradish peroxidase-conjugated secondary antibodies (cat.
nos. E-AB-1003 and E-AB-1001; 1:10,000; Elabscience Biotechnology,
Inc.) for 2 h at room temperature. A ChemiDoc™ Imager (VILBER,
Ltd.) was used to detect the chemiluminescence signals by dropping
chemiluminescence solution (Elabscience Biotechnology, Inc.)
Image-pro Plus (Version 5.0; Media Cybernetics, Inc.) software was
used to quantify protein expression in each image.
RNA isolation and reverse
transcription-quantitative (RT-q)PCR
Total intracellular RNA was extracted from HUVECs
using RNA simple Total RNA kit (Tiangen Biotech Co., Ltd.). A
Nanodrop spectrophotometer (Thermo Fisher Scientific, Inc.) was
used to determine the concentration and quality of the extracted
RNA. A total of 1 µg RNA was reverse-transcribed into cDNA using an
Evo M-MLV RT Mix kit with gDNA Clean for qPCR (GATC Biotech AG).
The cDNA was analyzed via qPCR (QuantStudio3, Thermo Fisher
Scientific, Inc.) using SYBR-Green Master Mix (GATC Biotech AG) to
determine the mRNA expression levels of the target genes. The
thermocycling conditions were as follows: 1 cycle of 95˚C for 10
min; 95˚C for 10 sec; 60˚C for 15 sec; 72˚C for 20 sec (40 cycles),
and 72˚C for 10 min. The starting template was quantitated using
the CT value estimated by the real-time PCR recorder. β-actin was
used as the internal control .The qPCR primers used were as
follows: PGC-1α forward (F), 5'-ATCTACTGCCTGGGGACCTT-3' and reverse
(R), 5'-ATGTGTCGCCTTCTTGCTCT-3'; NRF1 F, 5'-CGCAGCACCTTTGGAGAA-3'
and R, 5'-CCCGACCTGTGGAATACTTG-3'; TFAM F,
5'-GGCACAGGAAACCAGTTAGG-3' and R, 5'-CAGAACACCGTGGCTTCTAC-3'; and
β-actin F, 5'-ACGGCCAAGTCATCACTATTG-3' and R,
5'-AGCCACCGATCCACACAGA-3'.
Determination of mtDNA copy
number
Total DNA was extracted from cells using TIANamp
Genomic DNA kit (Tiangen Biotech Co., Ltd.). Using the
aforementioned method, qPCR was used to quantify mtDNA by assessing
the proportion of the mitochondrial D-loop region and the 18S
ribosomal (r)RNA expression levels; 18S rRNA was used to represent
nuclear DNA (nDNA). The primers used in the experiment were as
follows: Mitochondrial D-loop F, 5'-ATGGCCAACCTCCTACTCCT-3' and R,
5'-GCGGTGATGTAGAGGGTGAT-3'; and 18S F, 5'-CATTCGAACGTCTGCCCTATC-3'
and R, 5'-CCTGCTGCCTTCCTTGGA-3'.
Mitochondrial mass assessment
The cells were incubated in a confocal glass dish in
37˚C humidified with 5% CO2 until adherent. Following
adherence, the cells were treated with different concentrations of
SAA (10-3, 10-2 and 10-1 µM) and
incubated at 37˚C for 24 h. After washing with PBS, Mitotracker red
(Shanghai Yeasen Biotechnology Co., Ltd.) at a final concentration
of 50 nM was added and cells were incubated at 37˚C for 30 min in
the dark. Then the nuclei were stained with 10 µg/ml DAPI (Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
for 5 min. Staining was assessed using a confocal microscope (Zeiss
AG). The integrated optical density of the stained cells was
analyzed using Image-Pro Plus (version 5.0) software to assess the
mitochondrial mass.
Statistical analysis
All results are presented as the mean ± SEM of at
least three independent experiments. Statistical analysis was
performed using GraphPad Prism (version 5.0; GraphPad Software
Inc.). One-way ANOVA followed by Tukey's post hoc test was used to
assess the statistical significance of the differences between more
than two groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
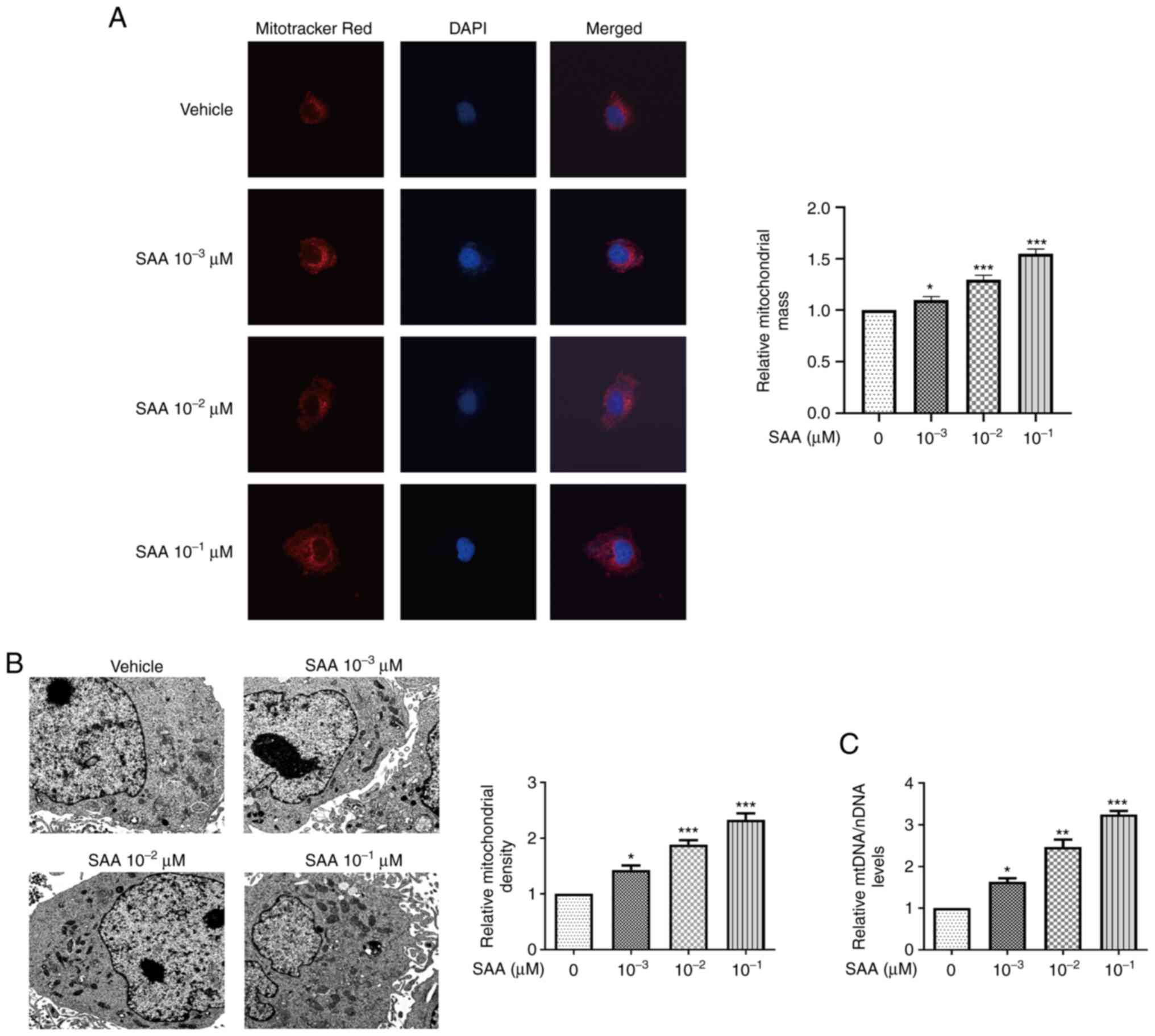
SAA promotes mitochondrial
biogenesis
To assess the effects of SAA on mitochondrial
biogenesis, Mitotracker red staining was used to detect the
mitochondrial mass of HUVECs. SAA treatment significantly increased
the mitochondrial mass in a dose-dependent manner compared with
cells treated with vehicle only (Fig.
1A). The number of mitochondria was also observed using
electron microscopy. Compared with the vehicle group, the number of
mitochondria in the SAA group significantly increased in a
dose-dependent manner, with the highest number of mitochondria in
cells treated with 10-1 µM SAA (Fig. 1B). Increased copy number of mtDNA
indicated mitochondrial biogenesis (Fig. 1C). The mtDNA levels were increased
by 1.8, 2.4, and 3.2 times with SAA treatment at 10-3,
10-2 and 10-1 µM, respectively. These results
indicated that SAA treatment significantly increased mtDNA levels
in HUVECs compared with the vehicle-only group. The results
suggested that SAA promotes mitochondrial biogenesis.
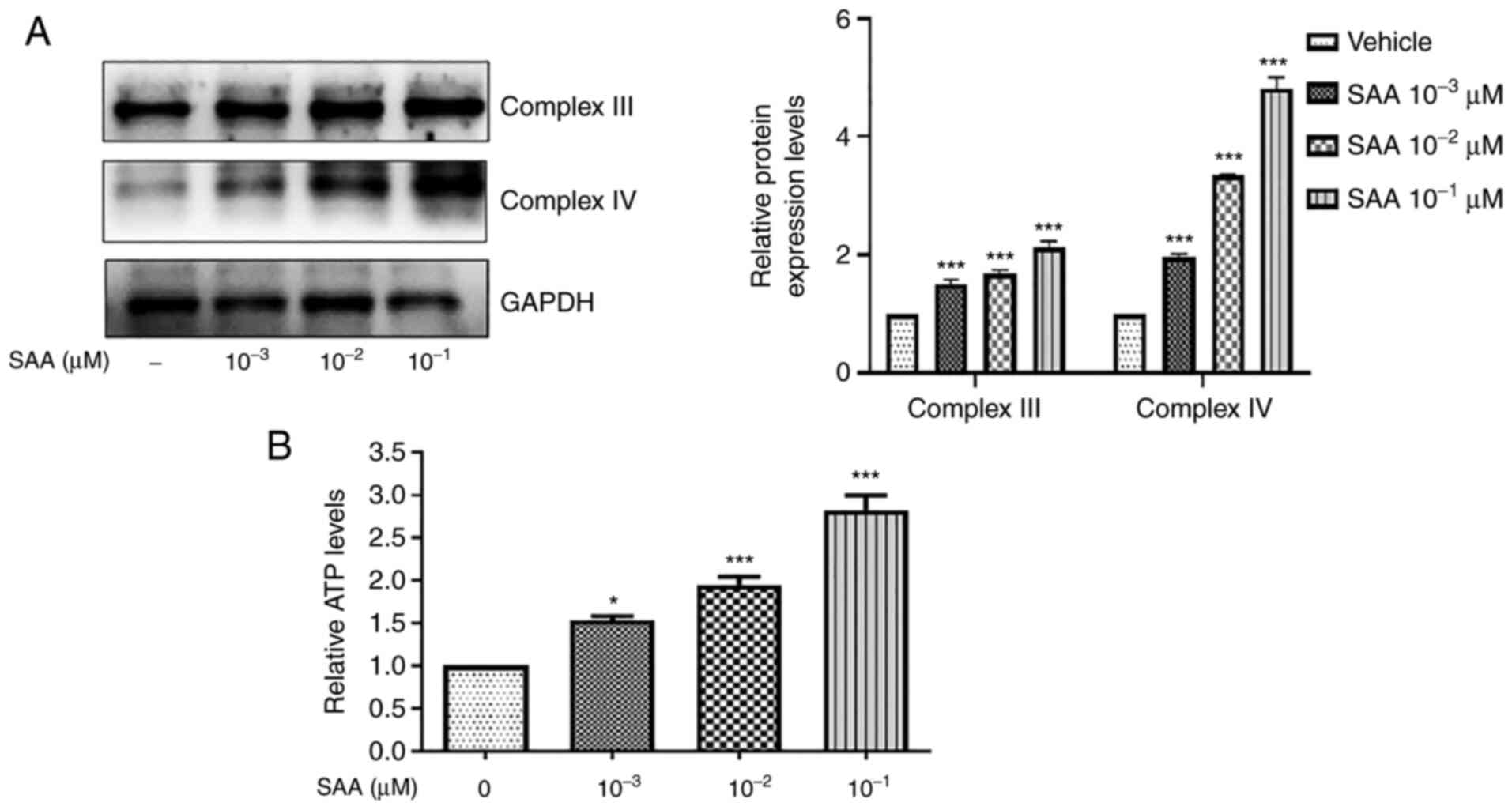
SAA enhances mitochondrial
function
Mitochondrial biogenesis increases mitochondrial
function and intracellular ATP is produced by the mitochondria
(19). To assess whether SAA
treatment increased mitochondrial function, intracellular ATP
levels were quantified in HUVECs and the protein expression levels
of the related mitochondrial complex Ⅲ and Ⅳ were determined via
western blotting. As presented in Fig.
2A, the protein expression levels of complex Ⅲ and Ⅳ were
significant increased following SAA treatment. In addition,
compared with the vehicle group, intracellular ATP levels increased
significantly after SAA treatment in a dose-dependent manner, which
suggested that SAA potentially enhanced mitochondrial function
(Fig. 2B).
SAA promotes the expression of PGC-1α,
NRF1 and TFAM
It has previously been reported that mitochondrial
biogenesis is associated with the PGC-1α/NRF1/TFAM signaling
pathway (20). It was therefore
hypothesized that the promotion of mitochondrial biogenesis by SAA
may also be related to the PGC-1α/NRF1/TFAM signaling pathway.
Therefore, the effect of SAA on the mRNA and protein expression
levels of PGC-1α, NRF1 and TFAM in HUVECs was investigated. The
results indicated that the mRNA and protein expression levels of
PGC-1α, NRF1 and TFAM were all significantly increased following
the treatment of HUVECs with SAA at concentrations of
10-3, 10-2 and 10-1 µM compared
with the vehicle group (Fig. 3).
These results suggested that SAA increased the expression of PGC-1
α, NRF1, TFAM, which are associated with mitochondrial
biogenesis.
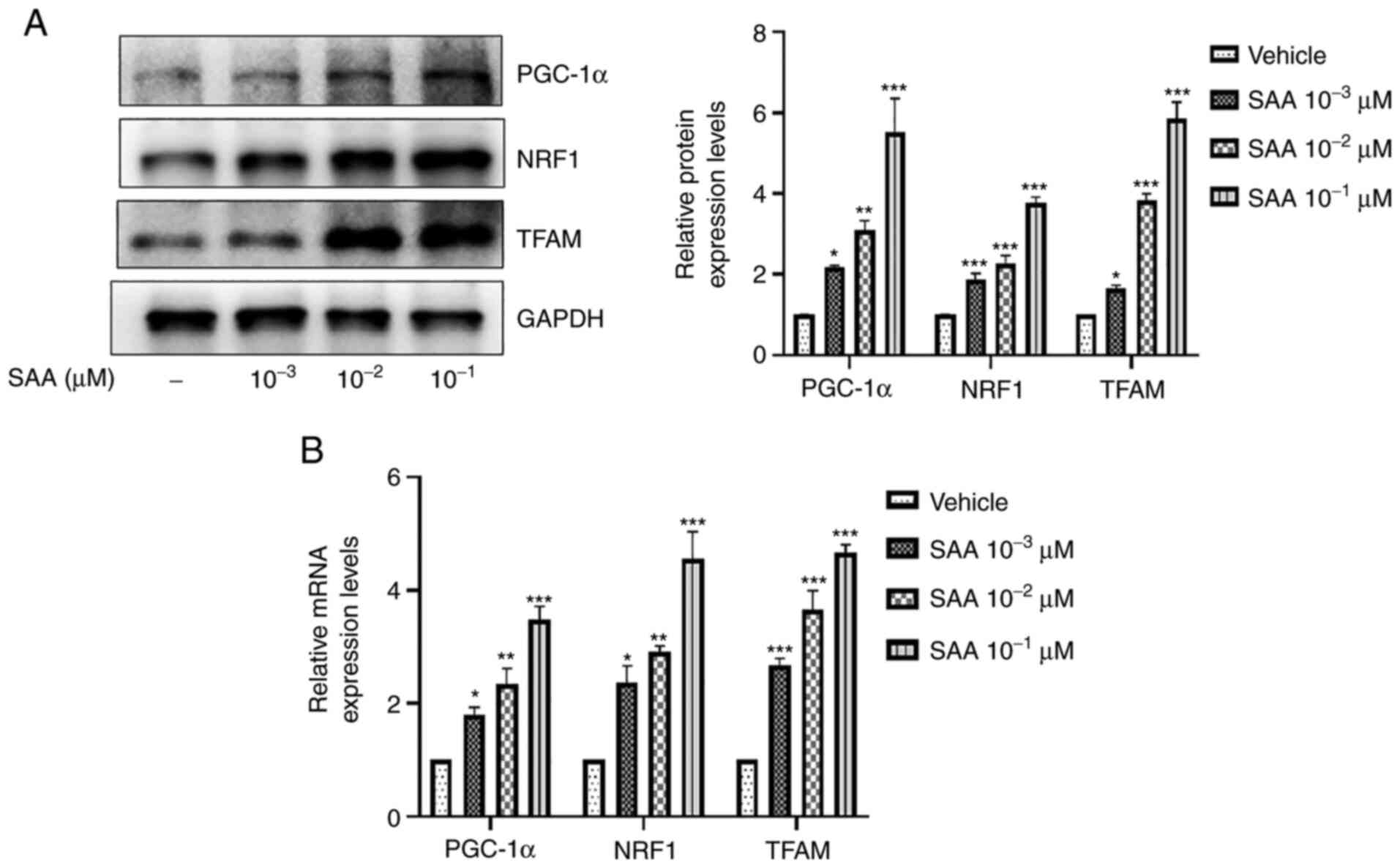 | Figure 3SAA promotes the expression of
PGC-1α, NRF1 and TFAM. (A) Following SAA treatment, western
blotting was performed to assess PGC-1α, NRF1 and TFAM protein
expression levels. (B) mRNA expression levels of PGC-1α, NRF1, and
TFAM were determined using reverse transcription-quantitative PCR.
*P<0.05, **P<0.01,
***P<0.001 vs. vehicle. SAA, salvianolic acid A;
PGC-1α, peroxisome proliferator-activated receptor γ coactivator
1-α; NRF1, nuclear respiratory factor 1; TFAM, mitochondrial
transcription factor A. |
SAA promotes AMPK signaling
pathway-mediated mitochondrial biogenesis
As mentioned above, SAA significantly increased the
expression levels of PGC-1α, NRF1 and TFAM, which thereby promoted
mitochondrial biogenesis. PGC-1α expression is dependent mainly on
upstream AMPK activation (21).
Therefore, it was hypothesized that the effects of SAA on
mitochondrial biogenesis may be caused via the activation of AMPK.
The results demonstrated that the AMPK specific inhibitor compound
C significantly reduced the expression levels of PGC-1α, and its
downstream target proteins NRF1 and TFAM compared with cells only
treated with SAA (10-2 µM) (Fig. 4A and B). Moreover, the SAA-mediated enhancement
of mtDNA levels was also significantly reduced following treatment
with compound C (2.5 µM) compared with cells treated with SAA
(10-2 µM; Fig. 4C).
AICAR, which is an AMPK activator, was used as a positive control.
The results showed that SAA significantly increased phosphorylation
levels of AMPK and ACC compared with SAA untreated group. However,
compound C significantly inhibited the phosphorylation of AMPK and
ACC by SAA (Fig. 4D).
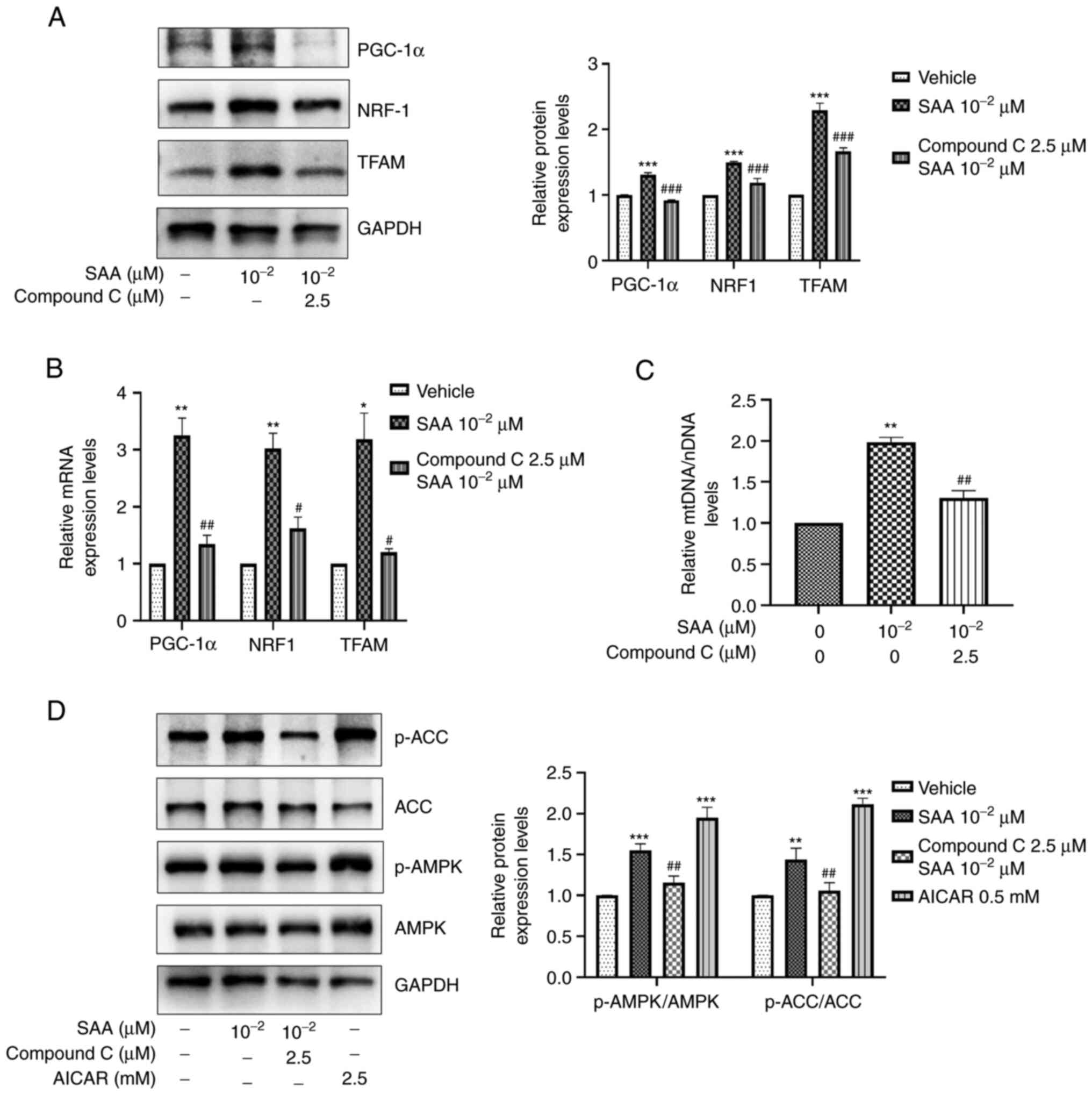 | Figure 4SAA promotes mitochondrial biogenesis
via the AMPK signaling pathway. (A) Following SAA and compound C
stimulation, western blotting was performed to assess PGC-1α, NRF1
and TFAM protein expression levels. (B) mRNA expression levels of
PGC-1α, NRF1 and TFAM were determined using RT-qPCR. (C) mtDNA/nDNA
was determined using RT-qPCR and is presented relative to vehicle.
(D) Following treatment of the cells with SAA, compound C and AICAR
stimulation, western blotting was performed for the detection of
p-AMPK and p-ACC expression levels. *P<0.05,
**P<0.01, ***P<0.001 vs. vehicle;
#P<0.05, ##P<0.01,
###P<0.001 vs. SAA treatment group. SAA, salvianolic
acid A; AICAR, 5-aminoimidazole-4-carboxamide ribonucleotide; AMPK,
5'AMP-activated protein kinase; PGC-1α, peroxisome
proliferator-activated receptor γ coactivator 1-α; NRF1, nuclear
respiratory factor 1; TFAM, mitochondrial transcription factor A;
RT-qPCR, reverse transcription-quantitative PCR; mtDNA/nDNA,
mitochondrial/nuclear DNA, p, phosphorylated; ACC, acetyl-CoA
carboxylase. |
Discussion
Vascular complications of diabetes are the most
common, and the leading cause of death and disability (19,22).
Endothelial vascular dysfunction is the main pathological basis of
diabetic vascular complications (23). Mitochondria are the most abundant
organelles in cells, and their reduced number and dysfunction serve
an important role in diabetes-related endothelial cell damage, as
well as in the occurrence and development of diabetic vascular
complications (24). Therefore,
the improvement of mitochondrial function in endothelial cells and
the induction of mitochondrial biogenesis may be an important
strategy to prevent diabetic vascular complications (25).
In the present study, SAA treatment significantly
increased mtDNA levels and mitochondrial mass in HUVECs compared
with the control group, which indicated that SAA may have promoted
mitochondrial biogenesis. Furthermore, the protein expression
levels of the mitochondrial respiratory chain complex Ш and Ⅳ, as
well as mitochondrial ATP production, were also significantly
increased by SAA treatment. These results indicated that SAA may
promote mitochondrial biogenesis and improve mitochondrial function
in HUVECs.
Mitochondrial biogenesis is a complex process.
PGC-1α is the main activator that controls the mitochondrial
biogenesis signaling pathway via activation of NRF1 and TFAM. This
signaling pathway serves an important role in mitochondrial
biogenesis (26). The
PGC-1α/NRF1/TFAM signaling pathway has previously been demonstrated
to be involved in mitochondrial biogenesis in numerous diseases
(27,28). Jiang et al (29) demonstrated that perampanel could
improve mitochondrial biogenesis in neuronal cells via activation
of the PGC-1α/NRF1 signaling pathway, which thereby exerts an
anti-epileptic effect. Sun et al (30) demonstrated that activation of the
PGC-1α signaling pathway was required to protect mitochondria and
promote mitochondrial biosynthesis in electroacupuncture
preconditioning-induced ischemia tolerance. Li et al
(31) reported that the
preservation or promotion of mitochondrial function via activation
of the PGC-1α/NRF1/TFAM signaling pathway could lead to
mitochondrial DNA and protein synthesis. The production of new
mitochondria is a potential therapeutic target for the treatment of
neurodegenerative diseases (32).
Chandrasekaran et al (33)
demonstrated that LY379268, a selective metabolomic glutamate
receptor 2/3 agonist, also upregulated the PGC-1α/TFAM signaling
pathway and prevented diabetic peripheral neuropathy via glutamate
recycling in Schwann/satellite glial cells and via the improvement
of dorsal root ganglion neuronal mitochondrial function. In the
present study, SAA significantly increased the mRNA and protein
expression levels of PGC-1α, and its downstream targets NRF1 and
TFAM, which suggested that the PGC-1α/NRF1/TFAM signaling pathway
is important for SAA-induced mitochondrial biogenesis.
AMPK is an important upstream regulatory factor of
PGC-1α and a master regulator of mitochondrial biogenesis (34). AMPK has also been demonstrated to
promote mitochondrial biogenesis and to improve endothelial
dysfunction (35,36). Previous studies have reported that
the activation of AMPK via phosphorylation promotes the expression
of PGC-1α, which thereby enhances mitochondrial biogenesis
(37). For example, ginger
(Zingiber officinale) extract has been shown to promote
mitochondrial biogenesis and improve mitochondrial function via
activation of the AMPK/PGC-1α signaling pathway (38). Similarly, naringenin can promote
mitochondrial biogenesis via activation of the AMPK/PGC-1α
signaling pathway (39). In the
present study, to explore the potential regulatory mechanism of
SAA, the phosphorylation of AMPK and downstream ACC were
investigated following treatment of HUVECs with SAA. The specific
inducer, AICAR, and inhibitor, compound C, of AMPK were used to
assess this. The results indicated that SAA significantly increased
the phosphorylation levels of AMPK and ACC in HUVECs, which was
similar to the results obtained using AICAR. Moreover, compound C
significantly inhibited the phosphorylation of AMPK and ACC
following SAA treatment of the cells. Compound C also significantly
reduced the protein expression levels of PGC-1α, and its downstream
targets NRF-1 and TFAM following SAA treatment of the cells. These
results suggested that AMPK may be a potential target of SAA, by
which the induction of PGC-1α expression and the activation of
mitochondrial biogenesis were achieved.
In conclusion, to the best of our knowledge, the
present study demonstrated for the first time that SAA promoted
mitochondrial biogenesis in HUVECs via the AMPK/PGC-1α/NRF1/TFAM
signaling pathway. These data provided an important theoretical
basis for the use of SAA in the treatment of diabetic complications
via improving endothelial vascular dysfunction. However, there are
other pathways involved in mitochondrial biogenesis. For example,
mitochondrial biogenesis can be promoted by upregulation of
AKT/STAT3 signal to inhibit apoptosis and mitochondrial biogenesis
of adipocytes can be promoted by activation of p38/CREB pathway
(40,41). Whether SAA promotes mitochondrial
biogenesis and enhances mitochondrial function is involved in other
pathways remains to be studied.
Acknowledgements
The authors would like to thank Professor Guanhua Du
(Institute of Materia Medica, Chinese Academy of Medical Sciences,
Beijing, China) for providing SAA for use in the present study.
Funding
Funding: The present study was funded by the National Natural
Science Foundation of China (grant no. 81903872) and the Special
Scientific Research and Academic Activities Project of Hospital
Pharmacy of Shandong Pharmaceutical Association (grant no.
yyyx2021qn-01).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JS and ZH designed the study. XW was responsible for
the data collection and manuscript writing. All authors
participated in the experiments. MiZ and MeZ were responsible for
data acquisition and analysis. YH was responsible for statistical
analysis. XC and WZ were responsible for literature searches. JS
and ZH reviewed and revised the manuscript. XW and MiZ confirm the
authenticity of all the raw data. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Qingdao University (Qingdao, China; approval no.
QYFYWZLL27047).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Domingueti C, Dusse L, Carvalho M, de
Sousa L, Gomes K and Fernandes A: Diabetes mellitus: The linkage
between oxidative stress, inflammation, hypercoagulability and
vascular complications. J Diabetes Complications. 30:738–745.
2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Huang D, Shi G, Jiang Y, Yao C and Zhu C:
A review on the potential of Resveratrol in prevention and therapy
of diabetes and diabetic complications. Biomed Pharmacother.
125(109767)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Gan Q, Wang J, Hu J, Lou G, Xiong H, Peng
C, Zheng S and Huang Q: The role of diosgenin in diabetes and
diabetic complications. J Steroid Biochem Mol Biol.
198(105575)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Yaribeygi H, Butler A, Barreto G and
Sahebkar A: Antioxidative potential of antidiabetic agents: A
possible protective mechanism against vascular complications in
diabetic patients. J Cell Physiol. 234:2436–2446. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yamagishi S, Nakamura N, Suematsu M,
Kaseda K and Matsui T: Advanced glycation end products: A molecular
target for vascular complications in diabetes. Mol Med. 21 (Suppl
1):S32–S40. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Gao D, Yu P, Jing S, Yan C, Ding D, Qiao Y
and Wu G: miR-193a as a potential mediator of WT-1/synaptopodin in
the renoprotective effect of Losartan on diabetic kidney. Can J
Physiol Pharmacol. 100:26–34. 2022.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Sjølie A, Dodson P and Hobbs F: Does
renin-angiotensin system blockade have a role in preventing
diabetic retinopathy? A clinical review. Int J Clin Pract.
65:148–153. 2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Rahadian A, Fukuda D, Salim HM, Yagi S,
Kusunose K, Yamada H, Soeki T, Shimabukuro M and Sata M: Thrombin
inhibition by dabigatran attenuates endothelial dysfunction in
diabetic mice. Vascul Pharmacol. 124(106632)2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Shu A, Du Q, Chen J, Gao Y, Zhu Y, Lv G,
Lu J, Chen Y and Xu H: Catalpol ameliorates endothelial dysfunction
and inflammation in diabetic nephropathy via suppression of
RAGE/RhoA/ROCK signaling pathway. Chem Biol Interact.
348(109625)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Xiang H, Song R, Ouyang J, Zhu R, Shu Z,
Liu Y, Wang X, Zhang D, Zhao J and Lu H: Organelle dynamics of
endothelial mitochondria in diabetic angiopathy. Eur J Pharmacol.
895(173865)2021.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Jia L, Wang J, Cao H, Zhang X, Rong W and
Xu Z: Activation of PGC-1α and mitochondrial biogenesis protects
against prenatal hypoxic-ischemic brain injury. Neuroscience.
432:63–72. 2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Qin Q, Jin J, He F, Zheng Y, Li T, Zhang Y
and He J: Humanin promotes mitochondrial biogenesis in pancreatic
MIN6 β-cells. Biochem Biophys Res Commun. 497:292–297.
2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Miller K, Clark J and Anderson R:
Mitochondrial regulator PGC-1a-Modulating the modulator. Curr Opin
Endocr Metab Res. 5:37–44. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhu X, Wang F, Lei X and Dong W:
Resveratrol alleviates alveolar epithelial cell injury induced by
hyperoxia by reducing apoptosis and mitochondrial dysfunction. Exp
Biol Med (Maywood). 246:596–606. 2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Lian-Niang L, Rui T and Wei-Ming C:
Salvianolic acid A, a new depside from roots of Salvia
miltiorrhiza. Planta Med. 50:227–228. 1984.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wu P, Yan Y, Ma LL, Hou BY, He YY, Zhang
L, Niu ZR, Song JK, Pang XC, Yang XY and Du GH: Effects of the Nrf2
protein modulator salvianolic acid A alone or combined with
metformin on diabetes-associated macrovascular and renal injury. J
Biol Chem. 291:22288–22301. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Qiang G, Yang X, Shi L, Zhang H, Chen B,
Zhao Y, Zu M, Zhou D, Guo J, Yang H, et al: Antidiabetic effect of
salvianolic acid A on diabetic animal models via AMPK activation
and mitochondrial regulation. Cell Physiol Biochem. 36:395–408.
2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Pang W, Zhang Y, Zhao N, Darwiche S, Fu X
and Xiang W: Low expression of Mfn2 is associated with
mitochondrial damage and apoptosis in the placental villi of early
unexplained miscarriage. Placenta. 34:613–618. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Knapp M, Tu X and Wu R: Vascular
endothelial dysfunction, a major mediator in diabetic
cardiomyopathy. Acta Pharmacol Sin. 40:1–8. 2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Yao K, Zhang W, Yao L, Yang S, Nie W and
Huang F: Carvedilol promotes mitochondrial biogenesis by regulating
the PGC-1/TFAM pathway in human umbilical vein endothelial cells
(HUVECs). Biochem Biophys Res Commun. 470:961–966. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ma D, Liu X, Liu J, Li M, Chen L, Gao M,
Xu W and Yang Y: Long-term liraglutide ameliorates nigrostriatal
impairment via regulating AMPK/PGC-1a signaling in diabetic mice.
Brain Res. 1714:126–132. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Oduro PK, Fang J, Niu L, Li Y, Li L, Zhao
X and Wang Q: Pharmacological management of vascular endothelial
dysfunction in diabetes: TCM and western medicine compared based on
biomarkers and biochemical parameters. Pharmacol Res.
158(104893)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jin J, Wang X, Zhi X and Meng D:
Epigenetic regulation in diabetic vascular complications. J Mol
Endocrinol. 63:R103–R115. 2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yuan Y, Shi M, Li L, Liu J, Chen B, Chen
Y, An X, Liu S, Luo R, Long D, et al: Mesenchymal stem
cell-conditioned media ameliorate diabetic endothelial dysfunction
by improving mitochondrial bioenergetics via the Sirt1/AMPK/PGC-1α
pathway. Clin Sci (Lond). 130:2181–2198. 2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Pangare M and Makino A: Mitochondrial
function in vascular endothelial cell in diabetes. J Smooth Muscle
Res. 48:1–26. 2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhao Q, Tian Z, Zhou G, Niu Q, Chen J, Li
P, Dong L, Xia T, Zhang S, Wang A, et al: SIRT1-dependent
mitochondrial biogenesis supports therapeutic effects of
resveratrol against neurodevelopment damage by fluoride.
Theranostics. 10:4822–4838. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Han H, Li X, Guo Y, Zheng M, Xue T and
Wang L: Plant sterol ester of α-linolenic acid Ameliorates high-fat
diet-induced nonalcoholic fatty liver disease in mice: association
with regulating mitochondrial dysfunction and oxidative stress via
activating AMPK signaling. Food Funct. 12:2171–2188.
2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhang X, Zhang Z, Yang Y, Suo Y, Liu R,
Qiu J, Zhao Y, Jiang N, Liu C, Tse G, et al: Alogliptin prevents
diastolic dysfunction and preserves left ventricular mitochondrial
function in diabetic rabbits. Cardiovasc Diabetol.
17(160)2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Jiang Y, Li D, Du Z, Li J, Lu R, Zhou Q,
Wang Q and Zhu H: Perampanel stimulates mitochondrial biogenesis in
neuronal cells through activation of the SIRT1/PGC-1α signaling
pathway. ACS Chem Neurosci. 12:323–329. 2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Sun S, Jiang T, Duan N, Wu M, Yan C, Li Y,
Cai M and Wang Q: Activation of CB1R-dependent PGC-1α is involved
in the improved mitochondrial biogenesis induced by
electroacupuncture pretreatment. Rejuvenation Res. 24:104–119.
2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li P, Hou X and Hao S: Mitochondrial
biogenesis in neurodegeneration. J Neurosci Res. 95:2025–2029.
2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Golpich M, Amini E, Mohamed Z, Azman Ali
R, Mohamed Ibrahim N and Ahmadiani A: Mitochondrial dysfunction and
biogenesis in neurodegenerative diseases: Pathogenesis and
treatment. CNS Neurosci Ther. 23:5–22. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Chandrasekaran K, Anjaneyulu M, Choi J,
Kumar P, Salimian M, Ho CY and Russell JW: Role of mitochondria in
diabetic peripheral neuropathy: Influencing the
NAD+-dependent SIRT1-PGC-1α-TFAM pathway. Int Rev
Neurobiol. 145:177–209. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Sun J, Song FH, Wu JY, Zhang LQ, Li DY,
Gao SJ, Liu DQ, Zhou YQ and Mei W: Sestrin2 overexpression
attenuates osteoarthritis pain via induction of
AMPK/PGC-1α-mediated mitochondrial biogenesis and suppression of
neuroinflammation. Brain Behav Immun. 102:53–70. 2022.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhang Y and Wu Q: Sulforaphane protects
intestinal epithelial cells against lipopolysaccharide-induced
injury by activating the AMPK/SIRT1/PGC-1α pathway. Bioengineered.
12:4349–4360. 2021.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zhang CL, Feng H, Li L, Wang JY, Wu D, Hao
YT, Wang Z, Zhang Y and Wu LL: Globular CTRP3 promotes
mitochondrial biogenesis in cardiomyocytes through AMPK/PGC-1α
pathway. Biochim Biophys Acta Gen Subj. 1861:3085–3094.
2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Cheng Q, Chen J, Guo H, Lu JL, Zhou J, Guo
XY, Shi Y, Zhang Y, Yu S, Zhang Q and Ding F: Pyrroloquinoline
quinone promotes mitochondrial biogenesis in rotenone-induced
Parkinson's disease model via AMPK activation. Acta Pharmacol Sin.
42:665–678. 2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Deng X, Zhang S, Wu J, Sun X, Shen Z, Dong
J and Huang J: Promotion of mitochondrial biogenesis via activation
of AMPK-PGC1α signaling pathway by ginger (Zingiber
officinale Roscoe) extract, and its major active component
6-Gingerol. J Food Sci. 84:2101–2111. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Kou G, Li Z, Wu C, Liu Y, Hu Y, Guo L, Xu
X and Zhou Z: Citrus tangeretin improves skeletal muscle
mitochondrial biogenesis via activating the AMPK-PGC1-α pathway in
vitro and in vivo: A possible mechanism for its beneficial effect
on physical performance. J Agric Food Chem. 66:11917–11925.
2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Liang N, Li S, Liang Y, Ma Y, Tang S, Ye S
and Xiao F: Clusterin inhibits Cr(VI)-induced apoptosis via
enhancing mitochondrial biogenesis through AKT-associated STAT3
activation in L02 hepatocytes. Ecotoxicol Environ Saf.
221(112447)2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Ning X, He J, Shi X, Yu T and Yang G:
Wnt3a regulates mitochondrial biogenesis through p38/CREB pathway.
Biochem Biophys Res Commun. 516:1019–1025. 2019.PubMed/NCBI View Article : Google Scholar
|