Introduction
Tauroursodeoxycholic acid (TUDCA) is a bile acid
taurine conjugate derived from ursodeoxycholic acid, which is
naturally occurring in the body. TUDCA has been approved by the
U.S. Food and Drug Administration to treat primary biliary
cholangitis, and shows good safety and effectiveness in clinical
practice (1). In addition to its
choleretic activity and protective role in hepatocytes, TUDCA has
been shown to inhibit endoplasmic reticulum (ER) stress, regulate
mitochondrial dysfunction, suppress cellular apoptosis and decrease
inflammation in various disease models (2-5).
Despite evidence demonstrating the beneficial effects of TUDCA in
pre-clinical studies, the underlying mechanisms are not fully
understood.
ER stress has been demonstrated to contribute to the
development and progression of a number of diseases, including
nerve injury (6,7), degenerative diseases of the nervous
system (8), diabetes (9), cardiovascular (10) and liver diseases (11,12),
and cancer (13). Therefore, ER
stress has been recognized as an emerging target for therapy.
Evidence from preclinical studies has suggested that targeting ER
stress components or different unfolded protein response (UPR)
signaling branches through gene therapy or pharmacological
approaches can delay neurodegeneration (14-16).
Apoptosis of dorsal root ganglion (DRG) neurons has been reported
in animal models of peripheral nerve injury (17). Studies of animal models with
sciatic nerve transection have shown increased levels of caspase-3,
caspase-8, caspase-12 and caspase-7 in the DRG, and phosphorylated
(p)-protein kinase R-like ER kinase (PERK) has been observed to
mainly colocalize with isolectin B4-positive neurons in the DRG
(18). In support of this, a
previous study demonstrated that the ER stress-mediated apoptotic
pathways were activated in the injured DRG and contributed to the
development of pain hypersensitivity after nerve injury (6). Therefore, pathogenic ER stress of the
DRG has been recognized as an emerging target for peripheral nerve
injury therapy.
The present study investigated the effects of TUDCA
on ER stress-associated apoptosis induced by tunicamycin in primary
cultured rat DRG neurons. Cell viability, caspase activity,
oxidative stress-related factors and activation of ER stress
pathways in DRG cells were studied, and the anti-apoptotic effect
and potential mechanism of TUDCA in DRG neurons was reported.
Materials and methods
Chemicals and reagents
Neurobasal medium, B-27, nerve growth factor (NGF),
penicillin-streptomycin and trypsin were obtained from Thermo
Fisher Scientific, Inc. Poly-D-lysine, tunicamycin, laminin,
collagenase and 5-fluoro-2'-deoxyuridine (FUDR) were purchased from
MilliporeSigma. TUDCA and thapsigargin were obtained from Selleck
Chemicals and dissolved in DMSO. Antibodies used in
immunofluorescence staining and western blotting are listed in
Table SI.
DRG neurons culture and treatment
Pregnant Sprague-Dawley rats (10-15 weeks old) were
provided by Beijing Vitalstar Biotechnology Co., Ltd. A total of 10
pregnant rats and 107 embryonic rats were used for DRG collection
in this study. Rats were maintained in plastic cages under
controlled conditions, with an ambient temperature of 23˚C and 50%
relative humidity, with free access to food and water. Rats were
placed in a room with a standard 12-h light/dark cycle. Pregnant
rats were euthanized by CO2 inhalation in the home cage
with a flow rate of 30% displacement of cage volume/min. DRG
neurons were isolated and cultured as previously described
(19,20). In brief, the DRG was harvested from
Sprague-Dawley rats at embryonic day 15 or 16 and digested with
0.25% trypsin and 0.3% collagenase type I at 37˚C for 25 min. DRG
neurons were then dissociated by repetitive pipetting and cells
were centrifuged at 200 x g at room temperature for 5 min. Cells
were resuspended in neurobasal medium supplemented with 2% B-27, 10
ng/ml NGF and penicillin-streptomycin (both 100 µg/ml), and seeded
into 96-well plates precoated with poly-D-lysine and laminin. DRG
neurons were cultured at 37˚C with 5% CO2 and medium was
replaced every day, cycled on media with or without 20 µM FUDR for
a total of 10 days to deplete non-neuronal cells. DRG neurons were
identified with antibodies against β-tubulin and neurofilament 200
(NF200).
Cell viability detection
Cell viability of DRG neurons was evaluated using
CellTiter-Blue Cell Viability assay (Promega Corporation). DRG
neurons were cultured in 96-well plates (5x104
cells/well) and pretreated with different doses of TUDCA (50, 100,
250, 500 and 1,000 µM) at 37˚C for 24 h. Medium was then replaced
with fresh medium containing tunicamycin (0.75 µg/ml) or
thapsigargin (1 µM) and incubated at 37˚C for another 24 h.
Following treatment, CellTiter-Blue reagent was
added directly to each well (20 µl/well) and the plate was agitated
for 10 sec and incubated at 37˚C for 1 h before measuring
fluorescence (560Ex/590Em) using a
multimode plate reader (Thermo Fisher Scientific Inc.). Each assay
was repeated at least three times. Cell viability was shown as a
percentage of the control group.
Immunofluorescent staining
DRG neurons were fixed with 4% paraformaldehyde for
30 min at room temperature before immunofluorescent staining. For
β-tubulin, glial fibrillary acidic protein (GFAP), ionized calcium
binding adaptor molecule 1 (Iba1) and NF200 staining, cells were
blocked with 3% BSA (Beyotime Institute of Biotechnology)
containing 0.1% Triton X-100 and 10% goat serum (Beyotime Institute
of Biotechnology) for 1 h at room temperature to avoid non-specific
staining. The cells were then incubated with primary antibodies
against β-tubulin (1:200), GFAP (1:200), Iba1 (1:800) or NF200
(1:200) overnight at 4˚C, and incubated with Alexa Fluor 488- or
Alexa Fluor 633-conjugated secondary antibodies for 2 h at room
temperature and stained with DAPI for 10 min at room temperature.
For the TUNEL assay, DRG neurons were washed with PBS and blocked
with 3% H2O2 in methanol for 10 min at room
temperature and permeabilized with 0.5% Triton X-100 for 5 min at
room temperature. Neurons were then incubated in the dark with 50
µl TUNEL reaction mixture (Beyotime Institute of Biotechnology) for
1 h at 37˚C. TUNEL-positive cells were observed with a Leica
confocal microscope (Leica Microsystems GmbH). A minimum of 10
random fields of view was observed by microscopy.
Caspase activity assay
Caspase activity of DRG neurons was assessed using
Caspase-Glo System (Promega Corporation) according to the
manufacturer's protocol. DRG neurons were plated in white-walled
96-well plates and treated with or without TUDCA and/or
tunicamycin. Caspase-Glo Reagent was then added to the cells (100
µl/well) and mixed well. Luminescence of each sample was measured
using a luminometer (Thermo Fischer Scientific, Inc.) after
incubation at room temperature for 1 h. Results were calculated as
signal-to-noise ratios and the relative activities were shown as
ratios of the control group.
Quantification of lactate
dehydrogenase (LDH)
LDH release in the culture supernatant of DRG
neurons was determined using the LDH Cytotoxicity Detection Kit
(Takara Bio, Inc.). The cell culture supernatant was collected and
transferred to a clear, flat-bottom microtiter plate after
treatment, 100 µl reaction mixture was added to each well and the
plate was incubated in the dark for 25 min at room temperature. The
reaction was stopped by adding 1N HCl to each well. LDH release was
measured at 490 nm using a multimode plate reader (Thermo Fisher
Scientific, Inc.). Each assay was repeated at least three times.
Relative LDH level was shown as a percentage of the untreated
control.
ROS detection
For ROS detection, cell culture medium of DRG
neurons was removed and 50 µl 2',7'-dichlorofluorescein diacetate
(DCFH-DA) (10 µM; Beyotime Institute of Biotechnology) was added to
each well (20 µl/well). Cells (10,000/well) were incubated at 37˚C
for 20 min and were then washed thoroughly to remove the DCFH-DA
that did not enter the cells. Fluorescence
(490Ex/525Em) was detected using a
multimode plate reader (Thermo Fisher Scientific, Inc.).
Malondialdehyde (MDA) detection
assay
Different concentrations of standards were used to
make a standard curve. Subsequently, 0.2 ml MDA detection working
solution (Beyotime Institute of Biotechnology) was added to each
sample. After mixing, the samples were boiled for 15 min and cooled
to room temperature in a water bath, then centrifuged at 1,000 x g
for 10 min at room temperature. The supernatant was moved to a new
96-well plate and absorbance was measured at 532 nm using a
microplate reader (Thermo Fisher Scientific, Inc.). The
concentration of MDA was calculated according to the standard curve
and the relative MDA levels were compared with the untreated
group.
Glutathione (GSH) detection assay
GSH production was determined using a GSH detection
assay kit (Beyotime Institute of Biotechnology). After treatment,
DRG neurons (10,000/well) were washed once with PBS and the
supernatant was removed. Protein removal reagent S solution (30
µl/well) was added to the cells and vortexed. Subsequently, liquid
nitrogen and a 37˚C water bath were used to freeze and thaw the
samples twice. Samples were placed at 4˚C for 5 min and centrifuged
at 10,000 x g for 10 min. The supernatant was then used for the
determination of GSH. Briefly, GSH detection working solution (150
µl/well) was added to standards and samples, mixed well and
incubated at room temperature for 5 min. Subsequently, 50 µl NADPH
solution (0.5 mg/ml) was added to wells and mixed, and absorbance
at 412 nm was immediately determined using a multimode plate
reader. Different concentrations of standards were used to make a
standard curve. GSH concentration was calculated by comparing the
sample to the standard curve.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from DRG neurons using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.) and
cDNA was generated using PrimeScript RT Reagent Kit (cat. no.
RR037B; Takara Bio, Inc.) according to the manufacturer's protocol.
Changes in mRNA expression were measured using the SYBR Green
Realtime PCR Master Mix kit (cat. no. QPK-201; Toyobo Life
Science). The amplification started with an initial denaturation
step at 95˚C for 60 sec, followed by 40 cycles of denaturation at
95˚C for 15 sec, annealing at 60˚C for 15 sec and extension at 72˚C
for 45 sec. Relative expression levels of target genes were
calculated using the 2-ΔΔCq method and normalized to
β-actin of respective samples (21). The primers used in this study are
listed in Table SII.
Western blot analysis
DRG neurons were harvested and lysed using RIPA
lysis buffer (Beyotime Institute of Biotechnology) supplemented
with a protease and phosphatase inhibitor cocktail (CoWin
Biosciences). Bicinchoninic acid assay kit (Pierce; Thermo Fisher
Scientific, Inc.) was used to quantify protein concentrations, and
equal amounts of protein (30 µg) were separated by SDS-PAGE and
transferred onto PVDF membranes (Millipore). Subsequently, the
membrane was blocked with 5% non-fat milk for 2 h at room
temperature and then incubated with primary antibodies overnight at
4˚C. After incubation with HRP-conjugated antibodies for 2 h at
room temperature, protein expression was visualized by ECL Western
Blotting Substrate (Pierce; Thermo Fisher Scientific, Inc.) and
semi-quantified using a Bio-Rad imaging system (Image Lab software
4.0; Bio-Rad Laboratories, Inc.).
Statistical analysis
All data are presented as the mean ± SEM.
Statistical analysis was performed using one-way ANOVA and Tukey
test using GraphPad Prism 7.0 (GraphPad Software, Inc.). P<0.05
was considered to indicate a statistically significant difference.
Each experiment was repeated at least three times.
Results
Effect of TUDCA on the viability of
DRG neurons
Double immunofluorescence staining of DRG neurons
derived from rat fetuses was performed to confirm the purity of the
cells. DRG neurons were positive for β-tubulin and NF200 (Fig. 1A). Iba1-labeled resident
macrophages and GFAP-positive glial cells were not observed
(Fig. 1A). The purity of DRG
neurons was >90%.
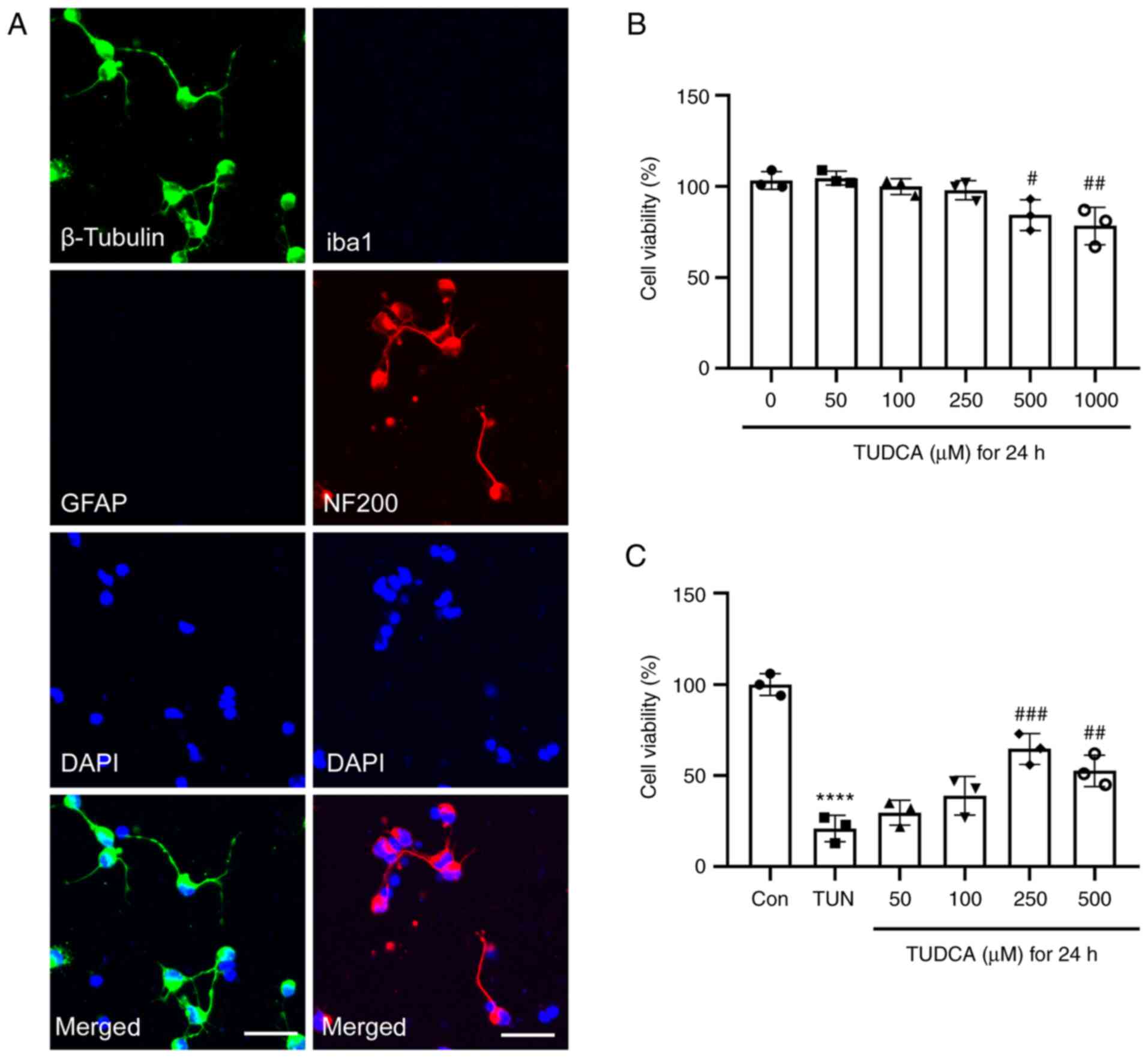 | Figure 1Effect of TUDCA on the viability of
DRG neurons. (A) Immunofluorescence staining of rat DRG neurons
in vitro. DRG neurons were stained with β-tubulin (green),
GFAP (red), Iba1 (green) and NF200 (red), and nuclei were detected
with DAPI staining (blue). Representative images from three
experiments are shown. Scale bar, 20 µm. (B) Effect of different
concentrations of TUDCA on the viability of DRG neurons. (C)
Tunicamycin-induced cytotoxicity in DRG neurons was suppressed by
TUDCA. Data were obtained from three independent experiments and
are expressed as the mean ± SEM. The control group was set at 100%.
****P<0.0001 vs. control group;
#P<0.05, ##P<0.01,
###P<0.001 vs. TUN group. DRG, dorsal root ganglion;
TUDCA, tauroursodeoxycholic acid; GFAP, glial fibrillary acidic
protein; Iba1, ionized calcium binding adaptor molecule 1; NF200,
neurofilament 200; TUN, tunicamycin. |
To explore the effect of TUDCA on DRG neurons,
cultures were incubated with medium containing various
concentrations (0, 50, 100, 250, 500 and 1,000 µM) of TUDCA for 24
h and the cell viability was detected by CellTiter-Blue assay.
TUDCA had no significant cytotoxic effect on DRG neurons at low
concentrations (50, 100 and 250 µM). There was a marked reduction
in cell viability when the concentration of TUDCA was ≥500 µM. Cell
viability of DRG neurons decreased by 20% after exposure to 1,000
µM TUDCA compared with that of untreated cells (Fig. 1B). Subsequently, the cytoprotective
effect of TUDCA against tunicamycin-induced cytotoxicity was
evaluated. A 24-h incubation with 0.75 µg/ml tunicamycin
significantly decreased the viability of DRG neurons; this was
reversed by TUDCA at the concentrations of 250 and 500 µM (Fig. 1C). Since TUDCA had no effect on
cell viability of DRG neurons at 250 µM, this concentration was
used in the subsequent experiments.
Effect of TUDCA on oxidative stress in
tunicamycin-induced DRG neurons
The oxidative stress response of DRG neurons
following tunicamycin stimulation with or without TUDCA
pre-incubation was determined by detecting ROS, LDH, MDA and GSH
levels. As shown in Fig. 2, levels
of ROS, LDH, MDA and GSH were not influenced by TUDCA alone at 250
µM. The levels of ROS, MDA and LDH in DRG neurons were increased by
tunicamycin, whereas the levels of the antioxidant GSH were
decreased. Tunicamycin-induced free radical generation and
oxidative stress was markedly suppressed by pretreatment with TUDCA
(250 µM) for 24 h, and the decrease in GSH concentration was also
relieved (Fig. 2).
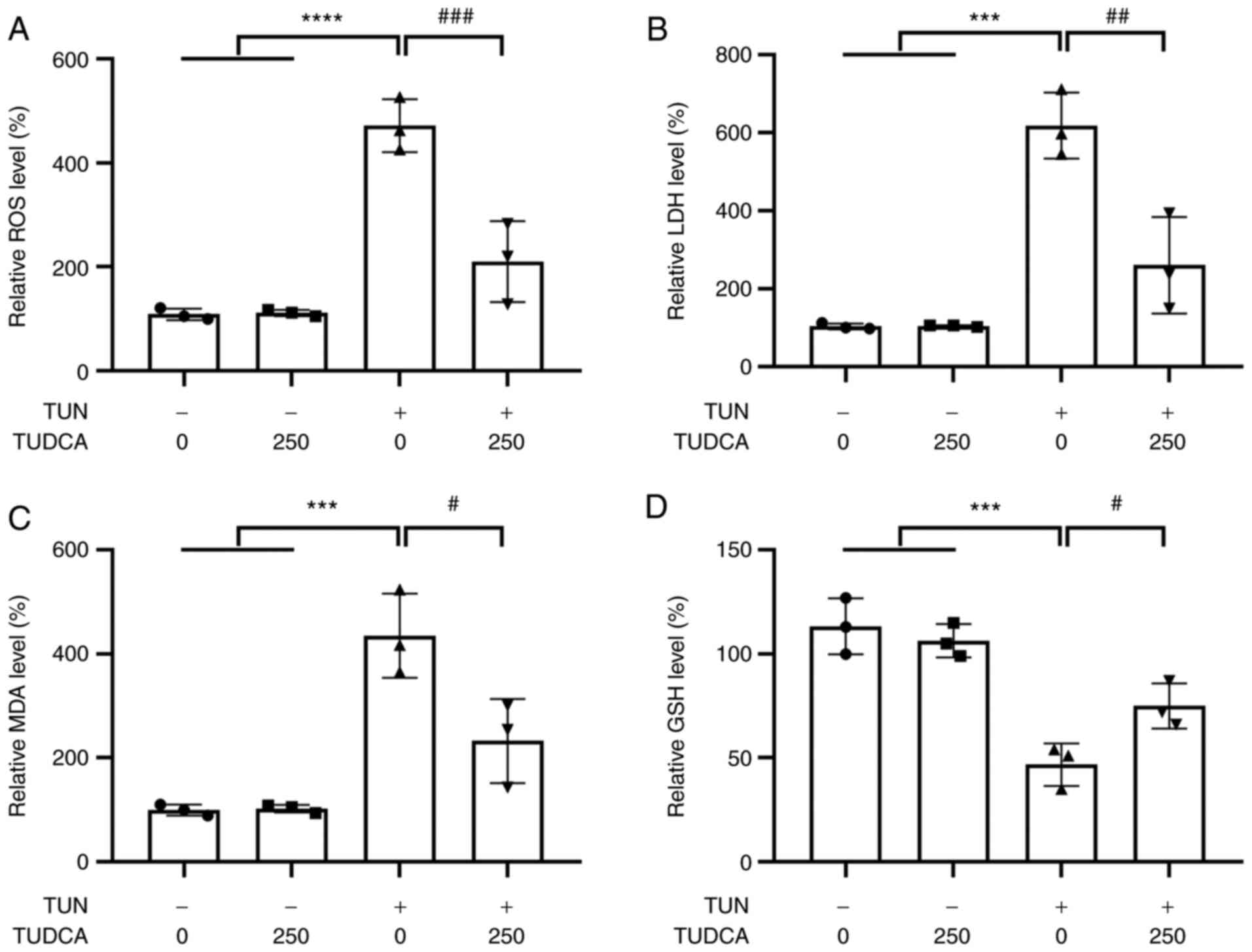 | Figure 2Effect of TUDCA on TUN-induced
oxidative damage in DRG neurons. DRG neurons were treated with 250
µM TUDCA for 24 h prior to a 24 h-stimulation with TUN (0.75
µg/ml). Relative (A) ROS, (B) LDH, (C) MDA and (D) GSH levels in
DRG neurons were detected 24 h after the indicated exposure. The
control group was set at 100%. Data were obtained from three
independent experiments and are expressed as the mean ± SEM.
***P<0.001, ****P<0.0001 vs. control
group; #P<0.05, ##P<0.01 and
###P<0.001 vs. TUN group. DRG, dorsal root ganglion;
TUDCA, tauroursodeoxycholic acid; TUN, tunicamycin; ROS, reactive
oxygen species; MDA, malondialdehyde; LDH, lactate dehydrogenase;
GSH, glutathione. |
TUDCA protects DRG neurons from
apoptosis
To explore whether apoptosis of DRG neurons followed
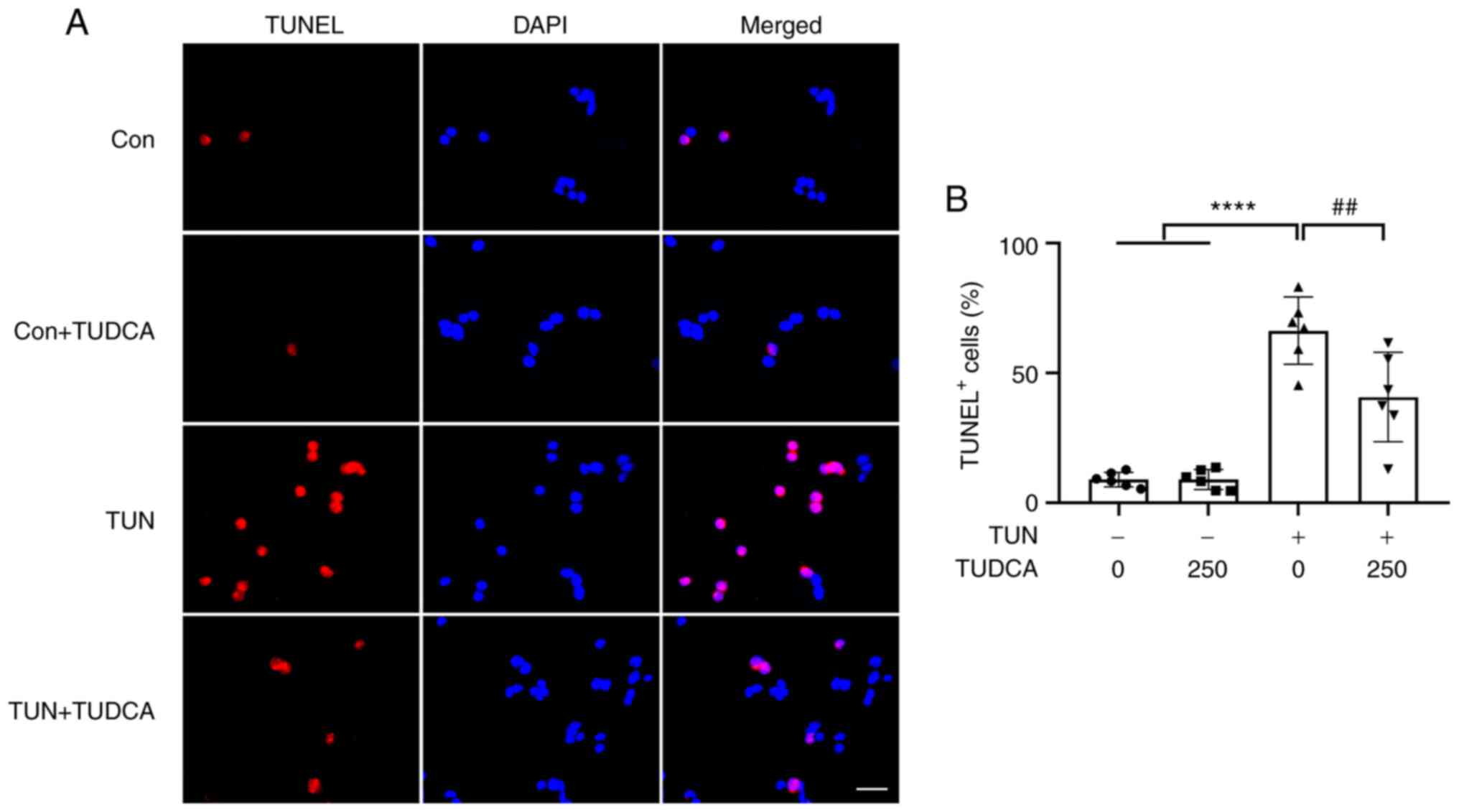
exposure to tunicamycin, TUNEL staining was performed. As shown in
Fig. 3, the percentages of
TUNEL-positive apoptotic cells were increased up to 60% in groups
exposed to tunicamycin compared with the TUN only group; however,
there was no change when cells were exposed to TUDCA alone. As
expected, pretreatment with TUDCA reduced the occurrence of
apoptosis in tunicamycin-treated DRG neurons (Fig. 3), indicating the neuroprotective
role of TUDCA.
Caspase activities and the expression levels of
apoptosis-related genes were examined by caspase-Glo assay and
RT-qPCR, respectively. A marked increase in the activities of
caspase-3 and caspase-9 was found in tunicamycin-treated DRG
neurons, whereas pretreatment with TUDCA induced a modest but
significant reduction in caspase-3 and caspase-9 activities as
compared with the tunicamycin group (Fig. 4A and B), which was consistent with the TUNEL
staining results. Similar trends were also found regarding the
expression levels of the pro-apoptotic factors Bax, DP5 and PUMA.
TUDCA suppressed the expression levels of Bax, DP5, cleaved
caspase-3 and PUMA in neurons induced by tunicamycin without
affecting the expression of total caspase-3 (Fig. 4C and E-L). By contrast, the expression levels
of Bcl-2, an apoptotic suppressor, were decreased by tunicamycin,
which was reversed by the addition of TUDCA (Fig. 4D). DRG neuronal apoptosis was also
induced by thapsigargin and a similar protective effect of TUDCA
was observed (Fig. S1).
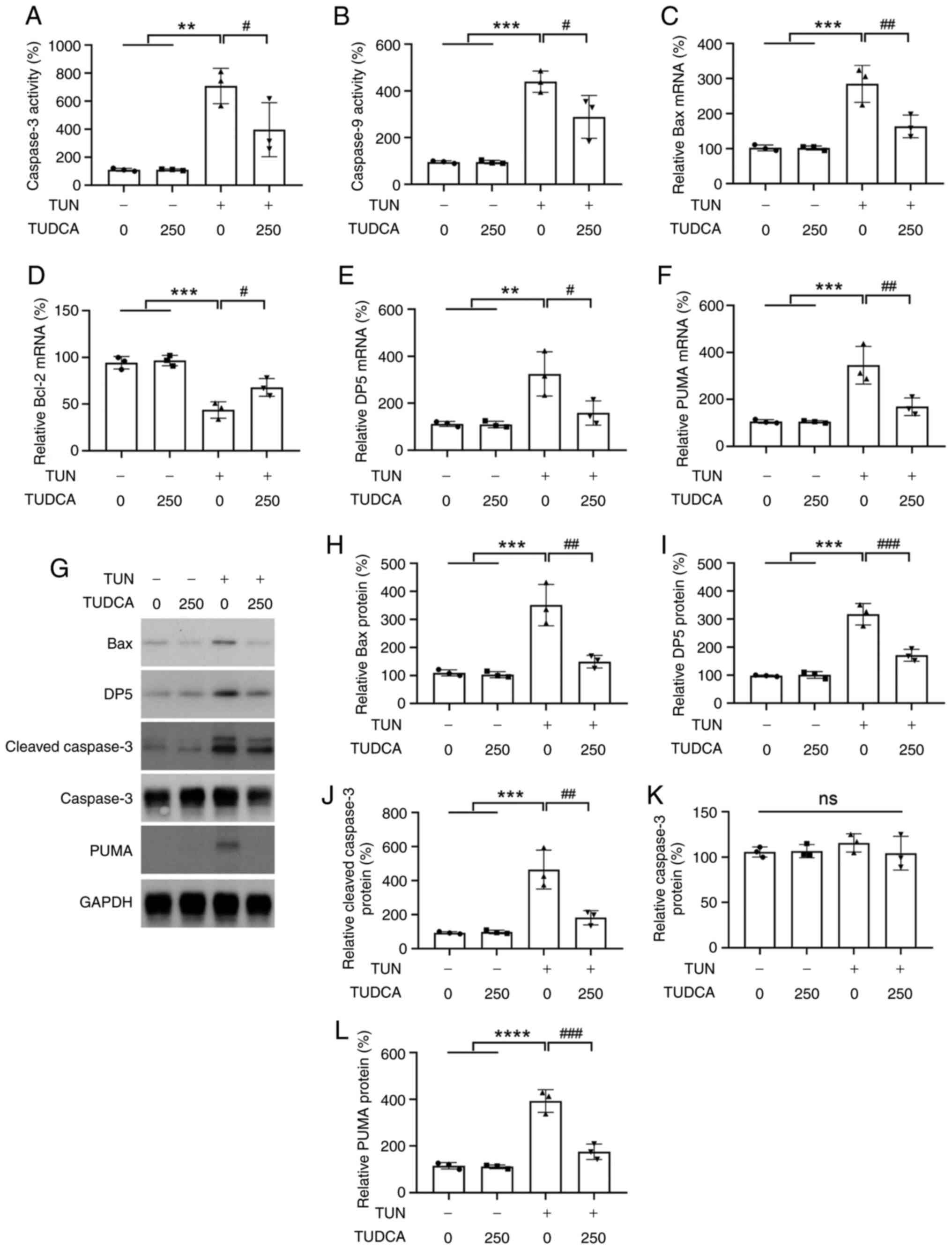 | Figure 4Changes in the levels of
apoptosis-related factors in DRG neurons after TUDCA and TUN
treatment. (A) Caspase-3 and (B) caspase-9 activity in DRG neurons
after TUDCA and TUN treatment was examined by Caspase-Glo kits.
Relative mRNA expression levels of (C) Bax, (D) Bcl-2, (E) DP5 and
(F) PUMA were analyzed by reverse transcription-quantitative PCR.
β-actin was used as an endogenous control. (G) Protein expression
levels of Bax, DP5, caspase-3 and PUMA were determined by western
blot analysis. Bar charts demonstrate the relative protein
expression levels of (H) Bax, (I) DP5, (J) cleaved caspase-3, (K)
total caspase-3 and (L) PUMA normalized to GAPDH or total caspase-3
for each group by densitometry. The control group was set at 100%.
Data were obtained from three independent experiments and are
expressed as the mean ± SEM. The control group was set at 100%.
**P<0.01, ***P<0.001,
****P<0.00001 vs. control group;
#P<0.05, ##P<0.01 and
###P<0.001 vs. TUN group. DRG, dorsal root ganglion;
TUDCA, tauroursodeoxycholic acid; TUN, tunicamycin. |
TUDCA suppresses tunicamycin-induced
ER stress activation in DRG neurons
Tunicamycin has been shown to cause ER stress in a
variety of cells (22-24).
In order to investigate whether ER stress response was involved in
the anti-apoptotic effect of TUDCA in DRG neurons, the protein
expression levels of C/EBP homologous protein (CHOP),
glucose-regulated protein 78 (GRP78) and cleaved caspase-12 were
assessed by western blotting. The CHOP pathway has been suggested
to be the main pathway through which apoptosis is induced during ER
stress (25). The protein
expression levels of CHOP and GRP78 were upregulated in
tunicamycin-treated DRG neurons as compared with the control group;
however, the upregulation of CHOP and GRP78 by tunicamycin was
suppressed by TUDCA pretreatment (Fig.
5A-C). In addition, elevated levels of cleaved caspase-12, a
key mediator of ER stress-induced apoptosis, were also observed in
DRG neurons following tunicamycin stimulation and reversed by TUDCA
pretreatment (Fig. 5A and D). There was no difference in total
caspase-12 levels in tunicamycin- or TUDCA-treated DRG neurons as
compared with that in the vehicle group (Fig. 5A and E).
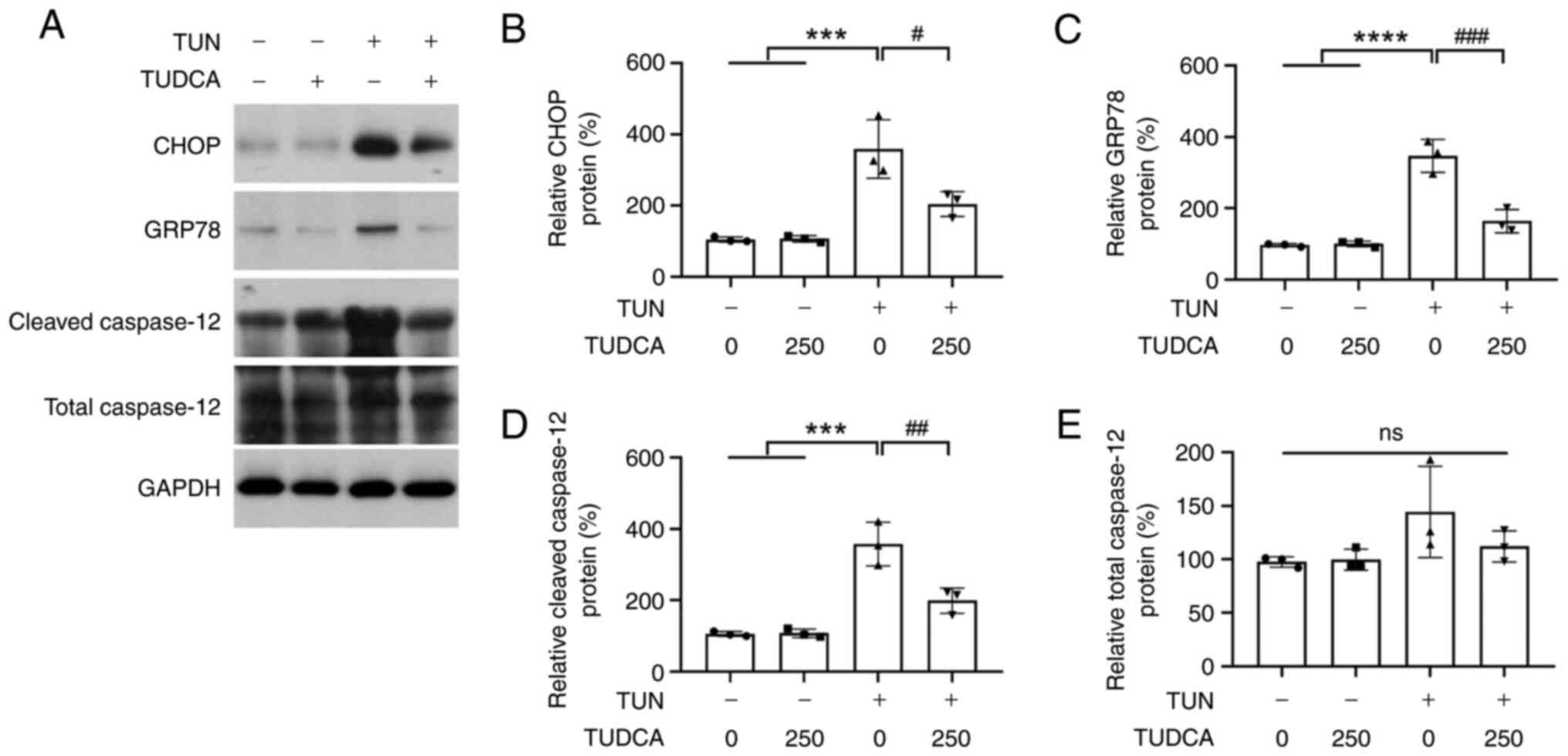 | Figure 5TUN-induced ER stress activation is
suppressed by TUDCA. (A) Western blotting was performed to detect
the protein expression levels of CHOP, GRP78, cleaved caspase-12
and total caspase-12. The bar charts demonstrate the ratio of (B)
CHOP, (C) GRP78, (D) total caspase-12 and (E) cleaved caspase-12
protein relative to GAPDH for each group by densitometry. The
control group was set at 100%. Data were obtained from three
independent experiments and are expressed as the mean ± SEM.
***P<0.001, ****P<0.0001 vs. control
group; #P<0.05, ##P<0.01 and
###P<0.001 vs. TUN group. TUDCA, tauroursodeoxycholic
acid; TUN, tunicamycin; CHOP, C/EBP homologous protein; GRP78,
glucose-regulated protein 78. |
Effect of TUDCA on the UPR pathway in
tunicamycin-induced DRG neurons
Under ER stress, the UPR is activated to maintain ER
and cellular function, and accumulation of unfolded proteins is
sensed through ER transmembrane protein sensors PERK, IRE1α and
ATF6. To better understand how the UPR pathways contribute to the
neuroprotective effect of TUDCA on DRG neurons, the activation of
PERK, IRE1α and ATF6 was investigated by western blotting. No
obvious expression changes in the phosphorylation of PERK (p-PERK)
and eIF2α (p-eIF2α; a signal downstream of PERK), as well as in
ATF6 levels, were observed among the control and 250 µM TUDCA
groups. By contrast, tunicamycin treatment increased the expression
levels of p-PERK and p-eIF2α, as well as IRE1α and ATF6 (Fig. 6). Moreover, activation of the UPR
pathways was inhibited by pre-incubation of DRG neurons with TUDCA
before tunicamycin exposure (Fig.
6).
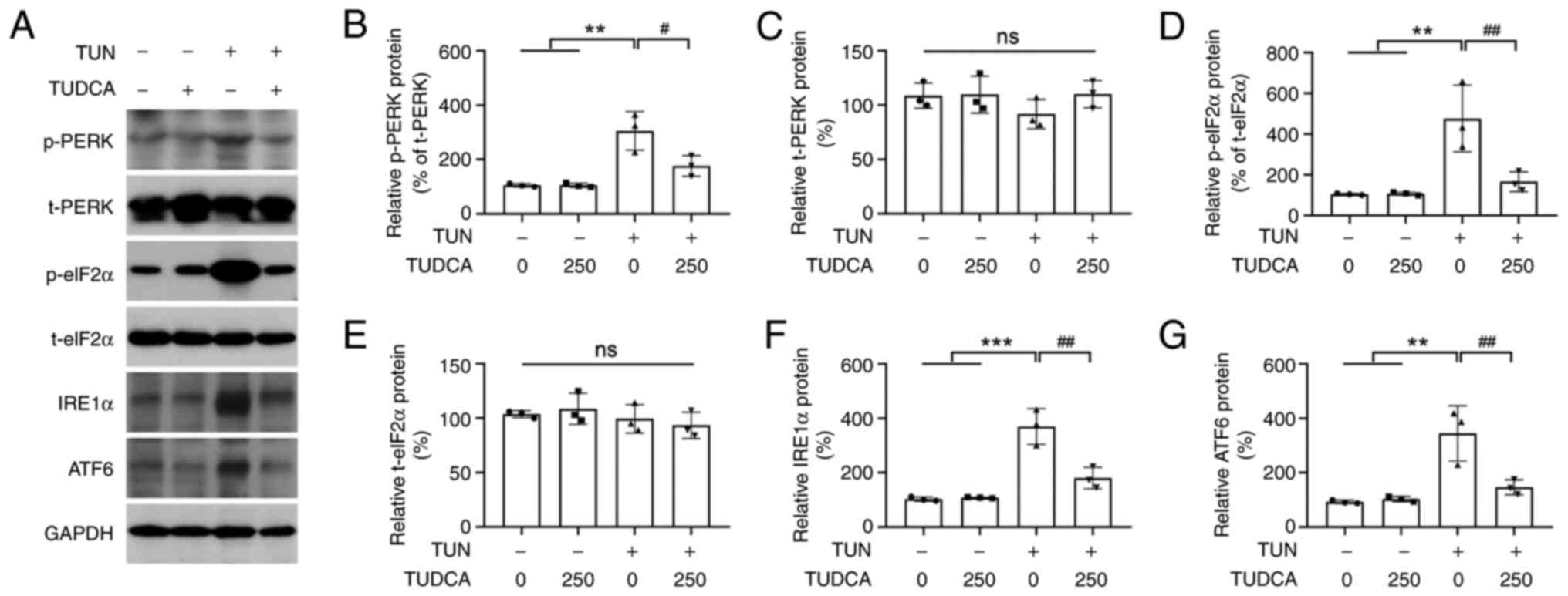 | Figure 6Effect of TUDCA on the UPR signaling
pathway in DRG neurons. (A) Protein expression levels of p-PERK,
t-PERK, p-eIF2α, t-eIF2α, IRE1α and ATF6 in DRG neurons following
TUN and TUDCA treatment were evaluated by western blotting. Bar
charts show the ratio of (B) p-PERK, (C) t-PERK, (D) p-eIF2α, (E)
t-eIF2α, (F) IRE1α and (G) ATF6, as determined by densitometry.
GAPDH was used as an internal reference; p-PERK levels were
normalized to t-PERK levels and p-eIF2α levels were normalized to
t-PERK levels. The control group was set at 100%. Data were
obtained from three independent experiments and are expressed as
the mean ± SEM. **P<0.01, ***P<0.001
vs. control group; #P<0.05 and ##P<0.01
vs. TUN group. DRG, dorsal root ganglion; TUDCA,
tauroursodeoxycholic acid; TUN, tunicamycin; PERK, protein kinase
R-like ER kinase; p, phosphorylated; t, total. |
Discussion
TUDCA has been shown to be the major active
ingredient of black bear bile, which has been utilized to treat
liver- and eye-related illnesses for centuries in traditional
Chinese medicine (2). TUDCA is
more hydrophilic and a more abundant naturally produced bile acid
in humans and bears than UDCA. A large number of potential uses for
TUDCA has been reported in studies of animal models (2,26,27).
Notably, TUDCA has attracted much attention from researchers as a
neuroprotective agent. The protective role of TUDCA in retinal
degeneration has been extensively reported in preclinical studies,
and intraperitoneal injection of TUDCA has been reported to protect
against oxidative stress-induced retinal degeneration (28), to promote neuronal survival after
experimental retinal detachment, to be closely associated with
oxidative stress and caspase activities (29), to reduce Brn311-labeled retinal
ganglion cell loss and to notably enhance GAP-43-positive axons in
a rat optic nerve crush model (30). In addition, several studies have
demonstrated neuroprotective roles of TUDCA in the brain. Chronic
feeding of TUDCA has been shown to interfere with amyloid-β peptide
production in APP/PS1 mice (14),
indicating its potential role in the treatment of Alzheimer's
disease. Wu et al (31)
reported that TUDCA administration attenuated neuronal apoptosis
and improved neurological functions in the pathogenesis of a
subarachnoid hemorrhage model. However, little is currently known
about the effect of TUDCA on rat DRG neurons. To the best of our
knowledge, the present study is the first to examine the
neuroprotective effect of TUDCA on tunicamycin-induced ER stress
and apoptosis of primary DRG neurons.
The present study showed that preincubation with
TUDCA (250 µM) markedly attenuated tunicamycin-induced cell death
of primary DRG neurons, as demonstrated by CellTiter-Blue assay,
TUNEL staining and LDH assay. When the concentration was ≤250 µM,
TUDCA exhibited no cytotoxicity in DRG neurons. These findings
indicated that TUDCA had a cytoprotective effect on DRG neurons
against tunicamycin-induced toxicity. These data were consistent
with the results of previous studies conducted in other types of
neurons, such as retinal (32),
primary cortical (33) and
dopamine neurons (34).
Despite these observations, little is currently
known about the exact molecular mechanism underlying these
phenomena. In general, inhibition of ER stress and stabilization of
the UPR pathways have been considered important mechanisms
underlying the biological functions of TUDCA (35-37).
Notably, TUDCA has been used as an inhibitor of ER stress in
previous studies (38,39). The present study revealed that
tunicamycin-induced ER stress activation and TUNEL-positive cells
were suppressed by pretreatment with TUDCA. In the presence of
tunicamycin, apoptosis of neurons was induced by ROS-mediated ER
stress, indicated by the enhancing of activities of caspase-3 and
caspase-9, and increasing of CHOP, GRP78 and ER resident cleaved
caspase-12 in DRG neurons exposed to tunicamycin. Both the
activation of ER stress and the enhanced caspase activities were
reversed by TUDCA. According to these data, it is strongly
conceivable that TUDCA rescued ER stress-associated apoptosis in
tunicamycin-treated DRG neurons.
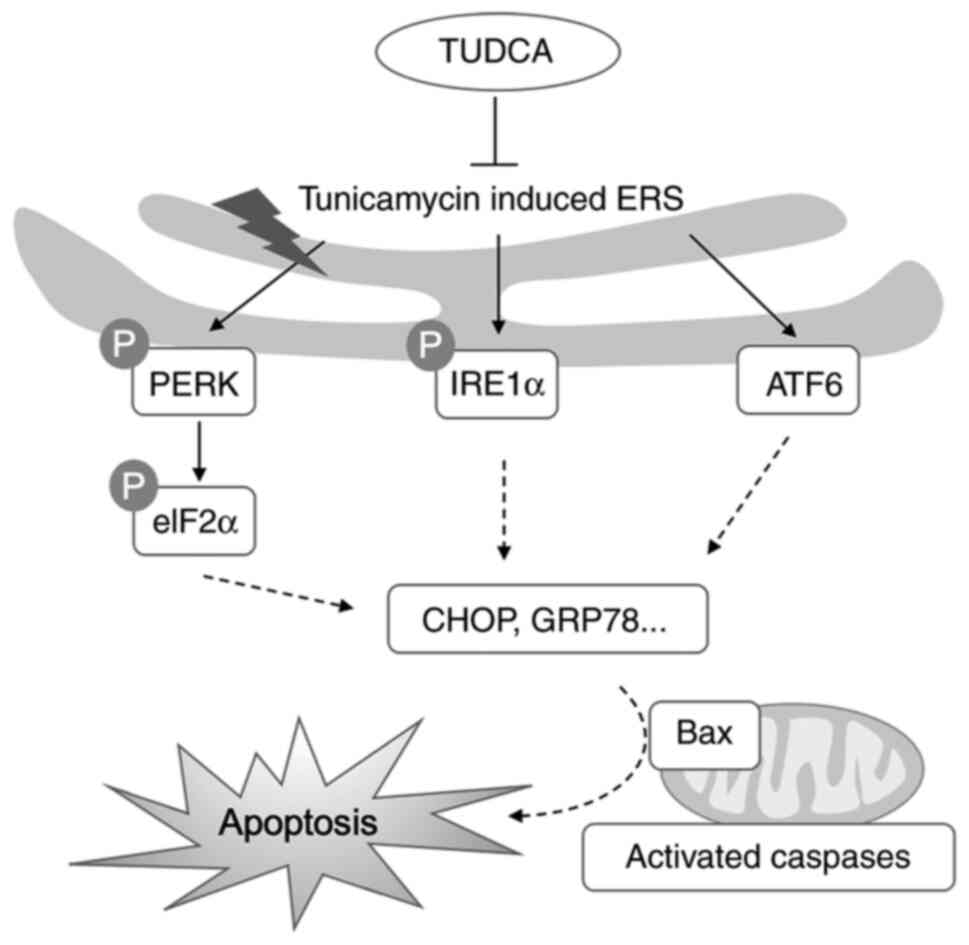
Considerable evidence has shown that ER stress is
initiated by the accumulation of unfolded or misfolded proteins in
the ER lumen. Consequently, PERK, IRE1α and ATF6 sense the fidelity
of protein folding in the ER lumen and activate the UPR to
eliminate or repair these misfolded proteins. In severe cases,
uncontrolled UPR can lead to cell death by activating the
pro-apoptotic signaling pathways. In healthy cells, the ER
environment is much more oxidative than the cytosolic compartment,
which favors protein folding (40,41).
Crosstalk between ER stress and oxidative stress has been
extensively reported in physiological and pathological conditions
(42,43). The present study evaluated the
oxidative stress in DRG neurons by detecting the levels of ROS, MDA
and the antioxidant GSH. As expected, the levels of ROS and MDA in
DRG neurons stimulated with tunicamycin were markedly higher,
whereas the levels of GSH were reduced compared with those in the
control group. However, these phenotypes could be alleviated by
pretreatment with TUDCA, suggesting that TUDCA protected primary
DRG neurons from oxidative stress in vitro. Using western
blot analysis, the activation of three branches of the UPR (PERK,
eIF2α, and ATF6) was assessed; all of these pathways were activated
in DRG neurons following tunicamycin exposure and were reversed by
TUDCA pretreatment (Fig. 7), as
determined by changes in p-PERK, p-eIF2α, IRE1α and ATF6
expression. These findings indicated that ER stress and the UPR may
be implicated in the mechanism that leads to cell apoptosis of
tunicamycin-induced DRG neurons and the neuroprotective role of
TUDCA.
The present study revealed that the expression of
the canonical transducer of the UPR, ATF6, was increased in DRG
neurons following tunicamycin exposure and was reversed by TUDCA.
These findings are consistent with the findings of Nakada et
al (44), which reported that
TUDCA may directly bind to ATF6 and reduce its protein levels. In
the present study, TUDCA also decreased the phosphorylation levels
of eIF2α and PERK, which may not only exert biological effects
through ATF6, but may also act through other upstream targets.
TUDCA has been reported to alleviate rifampicin-induced injury in
liver cancer cells by increasing the expression of bile acid
transporters and enhancing the Nrf2-mediated adaptive response
(45). The exact target of TUDCA
in DRG neurons still needs to be further investigated.
Taken together, the present study provided evidence
that TUDCA prevented ER stress-associated apoptosis in
tunicamycin-induced DRG neurons. TUDCA has been used to treat
primary biliary cholangitis, and has good safety and effectiveness
in clinical practice. However, the study of its effects on the
nervous system is still in its early stage. The protective effect
and therapeutic value of TUDCA in DRG neurons still need to be
further evaluated in spinal cord or sciatic nerve injury animal
models. Although the present study and a range of other studies
have shown the positive effects of TUDCA in cell cultures and
pre-clinical studies, there are certain limitations restraining its
clinical use, such as the relatively high TUDCA absorption capacity
of hepatocytes; therefore, researchers should consider the effects
of TUDCA on liver function when using TUDCA to treat neurological
diseases.
Supplementary Material
Effect of TUDCA on TG-induced
endoplasmic reticulum stress-associated apoptosis in DRG neurons.
Apoptotic DRG neurons were measured by TUNEL staining following
treatment with 1 μM TG and/or TUDCA. (A) Representative images of
TUNEL staining (green) in control, TUDCA, TG and TG + TUDCA groups.
Nuclear phenotype was also investigated via DAPI staining (blue).
Scale bar, 20 μm. (B) Quantitative analysis of percentages of
apoptotic cells after TUDCA treatment. (C) Protein expression
levels of cleaved caspase-3 and total caspase-3 were determined by
western blotting. (D) Relative protein expression levels of cleaved
caspase-3 to total caspase-3 for each group were calculated by
densitometry. The control group was set at 100%. Data were obtained
from three independent experiments and are expressed as the mean ±
SEM. ****P<0.0001 vs. control group;
###P<0.001 vs. TG group. DRG, dorsal root ganglion;
TUDCA, tauroursodeoxycholic acid; TG, thapsigargin.
Antibodies used in the present
study
Primers used in reverse
transcription-quantitative PCR
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by the Youth Project of
Shanghai Municipal Health and Family Planning Commission (grant no.
2018Y0141).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GC and XH designed the study and reviewed the
manuscript. FC and ZG acquired the majority of the data and drafted
the manuscript. NL and ZY analyzed the data and revised the
manuscript. RW and YZ interpreted the data, provided literature
support and revised the manuscript. All authors have read and
approved the final manuscript. RW and YZ confirm the authenticity
of all the raw data.
Ethics approval and consent to
participate
Rats were used according to the Guide for the Care
and Use of Laboratory Animals, 8th edition (46) and the present study was approved by
the Institutional Animal Care and Use Committee of Fudan University
(approval no. 20200524087).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ma H, Zeng M, Han Y, Yan H, Tang H, Sheng
J, Hu H, Cheng L, Xie Q, Zhu Y, et al: A multicenter, randomized,
double-blind trial comparing the efficacy and safety of TUDCA and
UDCA in Chinese patients with primary biliary cholangitis. Medicine
(Baltimore). 95(e5391)2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kusaczuk M: Tauroursodeoxycholate-bile
acid with chaperoning activity: Molecular and cellular effects and
therapeutic perspectives. Cells. 8(1471)2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ozcan U, Yilmaz E, Ozcan L, Furuhashi M,
Vaillancourt E, Smith RO, Görgün CZ and Hotamisligil GS: Chemical
chaperones reduce ER stress and restore glucose homeostasis in a
mouse model of type 2 diabetes. Science. 313:1137–1140.
2006.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Fonseca I, Gordino G, Moreira S, Nunes MJ,
Azevedo C, Gama MJ, Rodrigues E, Rodrigues CMP and Castro-Caldas M:
Tauroursodeoxycholic acid protects against mitochondrial
dysfunction and cell death via mitophagy in human neuroblastoma
cells. Mol Neurobiol. 54:6107–6119. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Li P, Fu D, Sheng Q, Yu S, Bao X and Lv Z:
TUDCA attenuates intestinal injury and inhibits endoplasmic
reticulum stress-mediated intestinal cell apoptosis in necrotizing
enterocolitis. Int Immunopharmacol. 74(105665)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yamaguchi Y, Oh-Hashi K, Matsuoka Y,
Takemura H, Yamakita S, Matsuda M, Sawa T and Amaya F: Endoplasmic
reticulum stress in the dorsal root ganglion contributes to the
development of pain hypersensitivity after nerve injury.
Neuroscience. 394:288–299. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tan HP, Guo Q, Hua G, Chen JX and Liang
JC: Inhibition of endoplasmic reticulum stress alleviates secondary
injury after traumatic brain injury. Neural Regen Res. 13:827–836.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yin Y, Sun G, Li E, Kiselyov K and Sun D:
ER stress and impaired autophagy flux in neuronal degeneration and
brain injury. Ageing Res Rev. 34:3–14. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Li W, Li W, Leng Y, Xiong Y and Xia Z:
Ferroptosis is involved in diabetes myocardial ischemia/reperfusion
injury through endoplasmic reticulum stress. DNA Cell Biol.
39:210–225. 2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Yang Y, Zhou Q, Gao A, Chen L and Li L:
Endoplasmic reticulum stress and focused drug discovery in
cardiovascular disease. Clin Chim Acta. 504:125–137.
2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ashraf NU and Sheikh TA: Endoplasmic
reticulum stress and oxidative stress in the pathogenesis of
non-alcoholic fatty liver disease. Free Radic Res. 49:1405–1418.
2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Lebeaupin C, Vallée D, Hazari Y, Hetz C,
Chevet E and Bailly-Maitre B: Endoplasmic reticulum stress
signalling and the pathogenesis of non-alcoholic fatty liver
disease. J Hepatol. 69:927–947. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Chen X and Cubillos-Ruiz JR: Endoplasmic
reticulum stress signals in the tumour and its microenvironment.
Nat Rev Cancer. 21:71–88. 2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Nunes AF, Amaral JD, Lo AC, Fonseca MB,
Viana RJ, Callaerts-Vegh Z, D'Hooge R and Rodrigues CM: TUDCA, a
bile acid, attenuates amyloid precursor protein processing and
amyloid-β deposition in APP/PS1 mice. Mol Neurobiol. 45:440–454.
2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Radford H, Moreno JA, Verity N, Halliday M
and Mallucci GR: PERK inhibition prevents tau-mediated
neurodegeneration in a mouse model of frontotemporal dementia. Acta
Neuropathol. 130:633–642. 2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Valenzuela V, Jackson KL, Sardi SP and
Hetz C: Gene therapy strategies to restore ER proteostasis in
disease. Mol Ther. 26:1404–1413. 2018.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lin CR, Yang CH, Huang CE, Wu CH, Chen YS,
Sheen-Chen SM, Huang HW and Chen KH: GADD45A protects against cell
death in dorsal root ganglion neurons following peripheral nerve
injury. J Neurosci Res. 89:689–699. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wiberg R, Novikova LN and Kingham PJ:
Evaluation of apoptotic pathways in dorsal root ganglion neurons
following peripheral nerve injury. Neuroreport. 29:779–785.
2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Chen F, Wu R, Zhu Z, Yin W, Xiong M, Sun
J, Ni M, Cai G and Zhang X: Wogonin protects rat dorsal root
ganglion neurons against tunicamycin-induced ER stress through the
PERK-eIF2α-ATF4 signaling pathway. J Mol Neurosci. 55:995–1005.
2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Levin E, Diekmann H and Fischer D: Highly
efficient transduction of primary adult CNS and PNS neurons. Sci
Rep. 6(38928)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yakin M, Seo B and Rich A:
Tunicamycin-induced endoplasmic reticulum stress up-regulates
tumour-promoting cytokines in oral squamous cell carcinoma.
Cytokine. 120:130–143. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wu F, Zhang R, Feng Q, Cheng H, Xue J and
Chen J: (-)-Clausenamide alleviated ER stress and apoptosis induced
by OGD/R in primary neuron cultures. Neurol Res. 42:730–738.
2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Quan X, Wang J, Liang C, Zheng H and Zhang
L: Melatonin inhibits tunicamycin-induced endoplasmic reticulum
stress and insulin resistance in skeletal muscle cells. Biochem
Biophys Res Commun. 463:1102–1107. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hu H, Tian M, Ding C and Yu S: The C/EBP
homologous protein (CHOP) transcription factor functions in
endoplasmic reticulum stress-induced apoptosis and microbial
infection. Front Immunol. 9(3083)2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zangerolamo L, Vettorazzi JF, Solon C,
Bronczek GA, Engel DF, Kurauti MA, Soares GM, Rodrigues KS, Velloso
LA, Boschero AC, et al: The bile acid TUDCA improves glucose
metabolism in streptozotocin-induced Alzheimer's disease mice
model. Mol Cell Endocrinol. 521(111116)2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lu Q, Jiang Z, Wang Q, Hu H and Zhao G:
The effect of tauroursodeoxycholic acid (TUDCA) and gut microbiota
on murine gallbladder stone formation. Ann Hepatol.
23(100289)2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Oveson BC, Iwase T, Hackett SF, Lee SY,
Usui S, Sedlak TW, Snyder SH, Campochiaro PA and Sung JU:
Constituents of bile, bilirubin and TUDCA, protect against
oxidative stress-induced retinal degeneration. J Neurochem.
116:144–153. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Mantopoulos D, Murakami Y, Comander J,
Thanos A, Roh M, Miller JW and Vavvas DG: Tauroursodeoxycholic acid
(TUDCA) protects photoreceptors from cell death after experimental
retinal detachment. PLoS One. 6(e24245)2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Kitamura Y, Bikbova G, Baba T, Yamamoto S
and Oshitari T: In vivo effects of single or combined topical
neuroprotective and regenerative agents on degeneration of retinal
ganglion cells in rat optic nerve crush model. Sci Rep.
9(101)2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wu H, Yu N, Wang X, Yang Y and Liang H:
Tauroursodeoxycholic acid attenuates neuronal apoptosis via the
TGR5/SIRT3 pathway after subarachnoid hemorrhage in rats. Biol Res.
53(56)2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Bikbova G, Oshitari T, Baba T and Yamamoto
S: Combination of neuroprotective and regenerative agents for
AGE-induced retinal degeneration: In vitro study. Biomed Res Int.
2017(8604723)2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Hou Y, Luan J, Huang T, Deng T, Li X, Xiao
Z, Zhan J, Luo D, Hou Y, Xu L and Lin D: Tauroursodeoxycholic acid
alleviates secondary injury in spinal cord injury mice by reducing
oxidative stress, apoptosis, and inflammatory response. J
Neuroinflammation. 18(216)2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Duan WM, Rodrigures CMP, Zhao LR, Steer CJ
and Low WC: Tauroursodeoxycholic acid improves the survival and
function of nigral transplants in a rat model of Parkinson's
disease. Cell Transplant. 11:195–205. 2002.PubMed/NCBI
|
|
35
|
Lee JH, Yoon YM and Lee SH: TUDCA-treated
mesenchymal stem cells protect against ER stress in the hippocampus
of a murine chronic kidney disease Model. Int J Mol Sci.
20(613)2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zhao H, Wang Y, Li B, Zheng T, Liu X, Hu
BH, Che J, Zhao T, Chen J, Hatzoglou M, et al: Role of endoplasmic
reticulum stress in otitis media. Front Genet.
11(495)2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Wen Y, Zong S, Liu T, Du P, Li H and Xiao
H: Tauroursodeoxycholic acid attenuates cisplatin-induced
ototoxicity by inhibiting the accumulation and aggregation of
unfolded or misfolded proteins in the endoplasmic reticulum.
Toxicology. 453(152736)2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kim EH and Park PH: Globular adiponectin
protects rat hepatocytes against acetaminophen-induced cell death
via modulation of the inflammasome activation and ER stress:
Critical role of autophagy induction. Biochem Pharmacol.
154:278–292. 2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Yu X, Wang T, Zhu M, Zhang L, Zhang F,
Jing E, Ren Y, Wang Z, Xin Z and Lin T: Transcriptome and
physiological analyses for revealing genes involved in wheat
response to endoplasmic reticulum stress. BMC Plant Biol.
19(193)2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Magallón M, Carrión AE, Bañuls L, Pellicer
D, Castillo S, Bondía S, Navarro-García MM, González C and Dasí F:
Oxidative stress and endoplasmic reticulum stress in rare
respiratory diseases. J Clin Med. 10(1268)2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Hwang C, Sinskey AJ and Lodish HF:
Oxidized redox state of glutathione in the endoplasmic reticulum.
Science. 257:1496–1502. 1992.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Victor P, Umapathy D, George L, Juttada U,
Ganesh GV, Amin KN, Viswanathan V and Ramkumar KM: Crosstalk
between endoplasmic reticulum stress and oxidative stress in the
progression of diabetic nephropathy. Cell Stress Chaperones.
26:311–321. 2021.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Cansler SM and Evanson NK: Connecting
endoplasmic reticulum and oxidative stress to retinal degeneration,
TBI, and traumatic optic neuropathy. J Neurosci Res. 98:571–574.
2020.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Nakada EM, Bhakta NR, Korwin-Mihavics BR,
Kumar A, Chamberlain N, Bruno SR, Chapman DG, Hoffman SM, Daphtary
N, Aliyeva M, et al: Conjugated bile acids attenuate
allergen-induced airway inflammation and hyperresponsiveness by
inhibiting UPR transducers. JCI Insight. 4(e98101)2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Zhang W, Chen L, Feng H, Wang W, Cai Y, Qi
F, Tao X, Liu J, Shen Y, Ren X, et al: Rifampicin-induced injury in
HepG2 cells is alleviated by TUDCA via increasing bile acid
transporters expression and enhancing the Nrf2-mediated adaptive
response. Free Radic Biol Med. 112:24–35. 2017.PubMed/NCBI View Article : Google Scholar
|
|
46
|
National Research Council (US): Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals. Guide for the care and use of laboratory animals. 8th
edition. National Academies Press, Washington, DC, 2011.
|