Introduction
Ulcerative colitis (UC) is a type of inflammatory
bowel disease (IBD), which is a group of chronic inflammatory
disorders of the colonic mucosa (1). In recent years, the number of
patients with UC has gradually increased in China and UC has become
a common disease of the digestive system (2). The clinical features of UC include
the development of colorectal inflammation and disruption of the
normal colonic mucosal barrier (3). The tight junctions between intestinal
epithelial cells serve an important role in maintaining intestinal
mucosal permeability (4). However,
intestinal infection, inflammation, mechanical injury and other
factors induce abnormal intestinal mucosal barrier function. These
factors increase mucosal permeability, which leads to the
displacement of bacteria and antigens in the intestinal lumen to
the lamina propria, which activates immune cells and thereby causes
an abnormal immune reaction in the mucosa (5). Furthermore, inflammatory cytokines,
inflammatory mediators and reactive oxygen radicals also mediate
the abnormal expression of tight junction proteins (6). Therefore, the improvement of
intestinal mucosal barrier function is a main focus of UC treatment
and an important area in UC pathophysiological research.
Fos-like antigen 1 (FOSL1), a member of the Fos
family, serves an important role in the progression and maintenance
of certain transformed cancer cells, including in lung cancer,
prostate cancer and colon cancer (7-10).
Previous study demonstrated that FOSL1 expression levels are
significantly elevated in patients with UC, especially in mild UC
(11). However, the underlying
molecular mechanisms are complex and remain to be elucidated.
Furthermore, FOSL1 overexpression in intestinal mucosal epithelial
cells increases the risk of IBD recurrence via the attenuation of
the protective effect of the intestinal mucosa in IBD during
remission, which results in the inhibition of the damage repair
mechanism (12). FOSL1 is also one
of the major transcription factors of the activator protein-1
(AP-1) family. Blocking the AP-1 transcription factor in mice
models has been demonstrated to suppress dextran sodium sulfate
(DSS)-induced colonic inflammation (13). It could therefore be hypothesized
that FOSL1 may serve an important role in UC. Therefore the present
study aimed to understand the specific impact of FOSL1 in UC.
MMPs are protein hydrolases that degrade the
extracellular matrix and are involved in tissue damage repair
(14). A previous study reported
that MMP13 mRNA expression levels are markedly elevated in mucosal
samples from patients with IBD and are positively correlated with
tissue inflammation levels (15,16).
MMP13-mediated inflammation-related signaling pathways also have
important implications for the integrity of the intestinal mucosal
epithelial cell barrier (17).
MMP13 can indirectly regulate the barrier function of intestinal
mucosal epithelial cells via the activation of TNF-α, which
increases the permeability of the intestinal epithelium (18). According to the JASPAR database,
FOSL1 potentially binds to the MMP13 promoter and may regulate its
expression levels. It can therefore be hypothesized that FOSL1 may
interact with MMP13, which may affect inflammatory damage and
intestinal barrier damage in UC.
In the present study, a DSS-induced in vitro
cell model and an in vivo mice model of UC were created and
the expression levels of FOSL1 and MMP13 in HT29 cells and UC mice
intestinal tissues were examined. Subsequently, the association
between FOSL1 and MMP13 was investigated. Furthermore, the present
study explored the effect of FOSL1 knockdown on DSS-induced
inflammation and barrier damage in HT29 cells and the UC mice
model. The results of the present study provided a comprehensive
account of the underlying mechanism of FOSL1 and have identified
novel research areas for the development of effective therapeutic
strategies for the treatment of UC.
Materials and methods
Cell culture and treatment
McCoy's 5a (modified) medium (American Type Culture
Collection), containing a final concentration of 10% FBS (RWD Life
Science Co., Ltd.), was used to culture the human colorectal cancer
cell line HT29 cells (American Type Culture Collection) at 37˚C in
5% CO2. For the establishment of the in vitro UC
cell model, HT29 cells were cultured until 80% confluency and were
subsequently treated with 2% DSS (MP Biomedicals, LLC) at 37˚C for
24 h.
Transfection
The FOSL1 overexpression vector (oe-FOSL1; 1 µg/ml),
the MMP13 overexpression (oe) vector (oe-MMP13; 1 µg/ml), the
negative control (oe-NC; 1 µg/ml), short hairpin RNA (shRNA/sh; 100
nm)-targeting FOSL1 (sh-FOSL1-1: forward,
5'-GCCTCTGACCTACCCTCAGTA-3' and reverse,
5'-TACTGAGGGTAGGTCAGAGGC-3'; sh-FOSL1-2: forward,
5'-AGTGGATGGTACAGCCTCATT-3' and reverse,
5'-AATGAGGCTGTACCATCCACT-3') and the corresponding negative control
(sh-NC,
5'-CCGGCAACAAGATGAAGAGCACCAACTCGAGTTGGTGCTCTTCATCTTGTTGTTTTTG-3'),
were purchased from Hunan Fenghui Biotechnology Co., Ltd. The
vectors were transfected into HT29 cells seeded into 6-well plates
at a density of 1x106 cells/well using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37˚C for 24 h. Reverse
transcription-quantitative (RT-q) PCR and western blotting were
used to examine the transfection efficiency of oe-FOSL1,
sh-FOSL1-1/2 and oe-MMP13. Cells were collected for subsequent
experiments after 24 h.
Animal experimental design
For the establishment of the in vivo UC mouse
model, a total of 24 male C57BL/6 mice (age, 6-8 weeks; weight,
21-23 g; Beijing HFK Bioscience Co., Ltd.) were used. Mice were
housed in specific pathogen-free conditions at 22-26˚C, under a
12/12 h light/dark cycle with 40-70% humidity and ad libitum access
to food and water. Mice were randomly divided into four groups
(n=6/group): mice in control group were fed as usual and given
distilled water; mice in UC group were fed as usual but was given
5% DSS daily for 7 days for modeling; mice in UC+sh-FOSL1 group
were injected with adeno-associated virus (AAV) shRNA-targeting
FOSL1 (sh-FOSL1) via the tail vein, companied with 5% DSS
administration; mice in UC+sh-FOSL1+oe-MMP13 group were injected
with adeno-associated virus (AAV) shRNA-targeting FOSL1 (sh-FOSL1)
and AAV targeting MMP13 overexpression (oe-MMP13) plasmids via the
tail vein, accompanied with 5% DSS administration. By the end of
experiment, the mice were euthanized via inhalation of isoflurane
(5% for induction and 1.5-2% for maintenance; cat. no. HR135327;
Hangzhou Hairui Chemical Co., Ltd.) prior to cervical dislocation.
The animals were monitored every day and the humane endpoint of
this experiment was as follows: Marked reduction in food or water
intake, labored breathing, inability to stand and no response to
external stimuli. No abnormal signs that signified the humane
endpoints of the experiment were observed from any of the rats
during the experiment. Mortality was verified by the lack of
heartbeat and a cold body. All experimental procedures were
approved by the Ethics Committee of the North China University of
Science and Technology Affiliated Hospital (approval no. Lx201896).
Colonic tissues were collected and the length was quantified. The
mice were scored using the disease activity index (DAI) based on
body weight, gross rectal bleeding and stool consistency (19). Serum and tissue samples were
collected and stored at -80˚C for further analysis.
RT-qPCR
FOSL1 and MMP13 mRNA expression levels were
determined using RT-qPCR. Briefly, total RNA was extracted from
HT29 cells (1x106 cells) using the MolPure®
Cell RNA kit (Shanghai Yeasen Biotechnology Co., Ltd.) according to
the manufacturer's protocol. Complementary DNA was synthesized from
the isolated RNA using the PrimeScript™ RT Reagent kit (Takara Bio,
Inc.) according to the manufacturer's protocols. qPCR was performed
using the One Step SYBR PrimeScript RT-PCR kit (Perfect Real Time;
TakaRa Bio Inc.) and the ABI 7500 qPCR instrument (Applied
Biosystems; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocols. The following thermocycling conditions
were used: Initial denaturation at 95˚C for 10 min; 40 cycles of
denaturation at 95˚C for 10 sec, annealing at 60˚C for 20 sec and
elongation at 72˚C for 30 sec; and a final extension at 72˚C for 7
min. Relative mRNA expression levels were determined using the
2-ΔΔCq method (20).
The experiments have been replicated for 3 times. Primers sequences
were: FOSL1, forward 5'-TGACCACACCCTCCCTAACTC-3' and reverse
5'-CTGCTGCTACTCTTGCGATGA-3'; MMP13; forward,
5'-AACATCCAAAAACGCCAGAC-3' and reverse 5'-GGAAGTTCTGGCCAAAATGA-3',
GAPDH, forward 5'-CCATGGGGAAGGTGAAGGTC-3' and reverse
5'-AGTGATGGCATGGACTGTGG-3'.
Western blotting
Total protein was extracted from HT29 cells and mice
colon tissues, from the different treatment groups, on ice using
RIPA lysis buffer (Beijing Solarbio Science & Technology Co.,
Ltd.) for 15 min. Total protein concentration was determined using
the BCA assay (Beijing Solarbio Science & Technology Co., Ltd.)
according to the manufacturer's protocol. Total protein (30 µg
protein/lane) was separated using SDS-PAGE on a 15% gel (Beijing
Leagene Biotechnology Co., Ltd.). Separated proteins were
transferred onto nitrocellulose membranes and the membranes were
blocked using 5% skimmed milk at room temperature for 1 h.
Subsequently, membranes were incubated with primary antibodies
against the following: FOSL1 (1:1,000; cat. no. ab252421; Abcam),
MMP13 (1:1,000; cat. no. ab51072; Abcam), occludin (1:1,000; cat.
no. ab216327; Abcam), zona occludens-1 (ZO-1; 1:1,000; cat. no.
ab276131; Abcam), claudin-2 (1:2,000; cat. no. ab211737; Abcam) and
GAPDH (1:2,500; cat. no. ab9485, Abcam), overnight at 4˚C. Then the
membranes were incubated with HRP-conjugated IgG secondary antibody
(1:20,000; cat. no. ab205718; Abcam) for 1 h at room temperature.
An ECL reagent (Beyotime Institute of Biotechnology) was used to
visualize the separated proteins. Image Pro Plus software version
7.0 (Media Cybernetics, Inc.) was used to analyze the protein
blots. GAPDH served as an internal control.
Dual-luciferase reporter assay
The wild-type (WT) or mutant (MUT) reporter plasmids
of MMP13 were generated by inserting WT or MUT sequences into pGL3
luciferase reporter plasmids (Promega Corporation). HT29 cells were
seeded into 24-well plates (1x105 cells/well) and
incubated at 37˚C for 24 h. Subsequently, cells were co-transfected
with reporter plasmids and oe-FOSL1/oe-NC using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Following transfection for 48 h, the luciferase
reporter activity was detected using the Dual-luciferase reporter
assay (Promega Corporation) and normalized to Renilla
luciferase activities.
Chromatin immunoprecipitation (ChIP)
assay
The interaction between FOSL1 and the MMP13 promoter
was assessed using the ChIP Assay kit (Beyotime Institute of
Biotechnology). Cells were fixed with 4% formaldehyde at room
temperature for 10 min for fixation. Subsequently, glycine solution
was added and the cells were incubated at room temperature for 5
min. The culture medium was then discarded and the pre-chilled PBS
was added to wash the cells. Following centrifugation at 4˚C at 716
x g for 5 min, SDS Lysis Buffer was added for 10 min in an ice bath
to fully lyse the cells. After sonicating the samples (20 kHz; 4
pulses of 12 sec each, followed by 30 sec rest on ice between each
pulse), 1.8 ml ChIP dilution buffer was added. Subsequently, 70 µl
protein A/G agarose was added for 30 min at 4˚C. After
centrifugation at 4,000 x g for 1 min at 4˚C, the supernatant was
collected and incubated overnight at 4˚C with anti-FOSL1 (1:30;
cat. no. ab252421; Abcam) or anti-IgG (1:100; cat. no. ab90285;
Abcam). Following incubation, 60 µl protein A/G agarose was added
for 60 min at 4˚C to precipitate the protein or corresponding
complex recognized by the primary antibody. The resulting
precipitates were detected via RT-qPCR according to the
aforementioned protocol.
ELISA
The levels of the inflammatory cytokines, TNF-α,
IL-1β and IL-6 in HT29 cells and UC mice serum were assessed using
the corresponding ELISA kits (Mouse TNF-α ELISA KIT (cat. no.
SEKM-0034), Human TNF-α ELISA KIT (cat. no. SEKM-0047), Mouse IL-1β
ELISA KIT (cat. no. SEKM-0002), Human IL-1β ELISA KIT (cat. no.
SEKM-0002), Mouse IL-6 ELISA KIT (cat. no. SEKM-0007), Human IL-6
ELISA KIT (cat. no. SEKM-0013)) from Beijing Solarbio Science &
Technology Co., Ltd. according to the manufacturer's protocol.
Trans-epithelial electrical resistance
(TEER) assay
The TEER assay was performed to identify HT29 cell
monolayer permeability. Briefly, cells were seeded into a polyester
Transwell plate (diameter, 12 mm; pore size, 0.4 µm; 24-well;
Corning, Inc.). After 24 h, initial resistance readings of all
co-cultures were obtained from an epithelial voltmeter
(EVOM2, World Precision Instruments, Inc.). The
resistance of each cell monolayer was determined by the relative
TEER value of each group, compared to the control.
Hematoxylin and eosin (H&E)
staining
Colon tissues from the UC model mice were first
washed with PBS, then fixed using 4% paraformaldehyde at room
temperature for 24 h. Next, tissues were dehydrated in a gradient
alcohol series, treated in xylene and were embedded in paraffin.
Subsequently, these tissues were made into 3-4 µm paraffin
sections. Following de-paraffinization, sections were stained using
hematoxylin for 2 min at room temperature. The samples were rinsed
under running water and were dehydrated in 95% alcohol for 30 sec
prior to being stained with eosin for 2 min at room temperature.
The sections were then washed, dehydrated and sealed with neutral
resin sealing sheets. An Olympus light microscope (BX43; Olympus
Corporation) was used to capture the images.
Statistical analysis
All data are presented as the mean ± SD. Data were
analyzed using SPSS version 20.0 (IBM Corp.). Unpaired Student's
t-test was used for statistical comparisons between two groups and
one-way ANOVA followed by Tukey's post hoc test was used for
statistical comparisons among more than two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
FOSL1 and MMP13 expression levels are
upregulated in DSS-induced HT29 cells
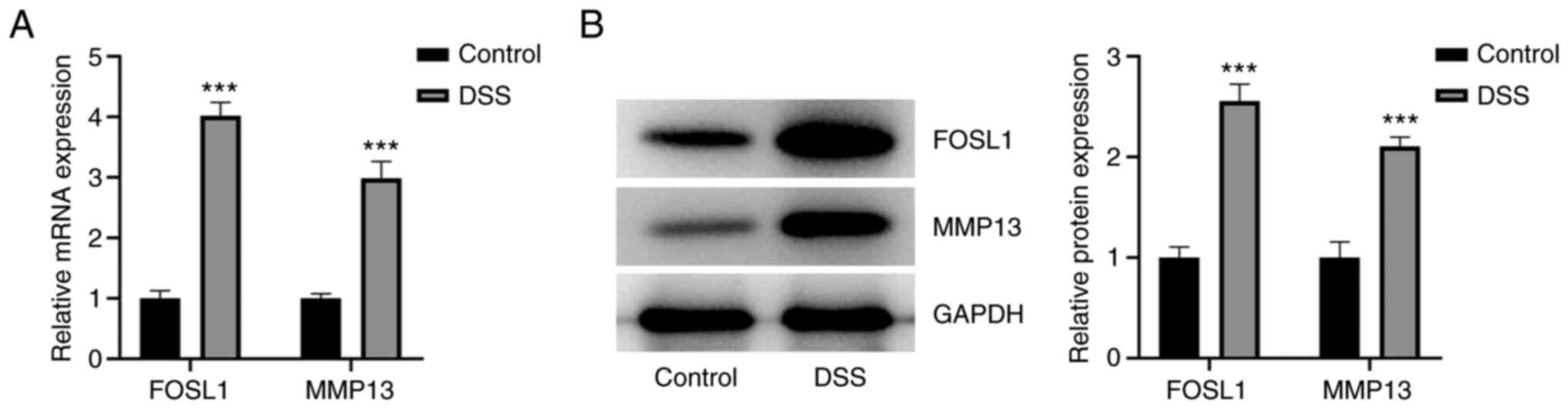
To examine the expression levels of FOSL1 and MMP13
in DSS-induced HT29 cells, an in vitro cell model of UC was
constructed and RT-qPCR and western blotting were performed. The
results demonstrated that there was an increase in the relative
FOSL1 and MMP13 mRNA and protein expression levels in DSS-induced
HT29 cells, compared with the control group (Fig. 1A and B). These results therefore indicated that
DSS potentially induced high mRNA and protein expression levels of
FOSL1 and MMP13 in HT29 cells.
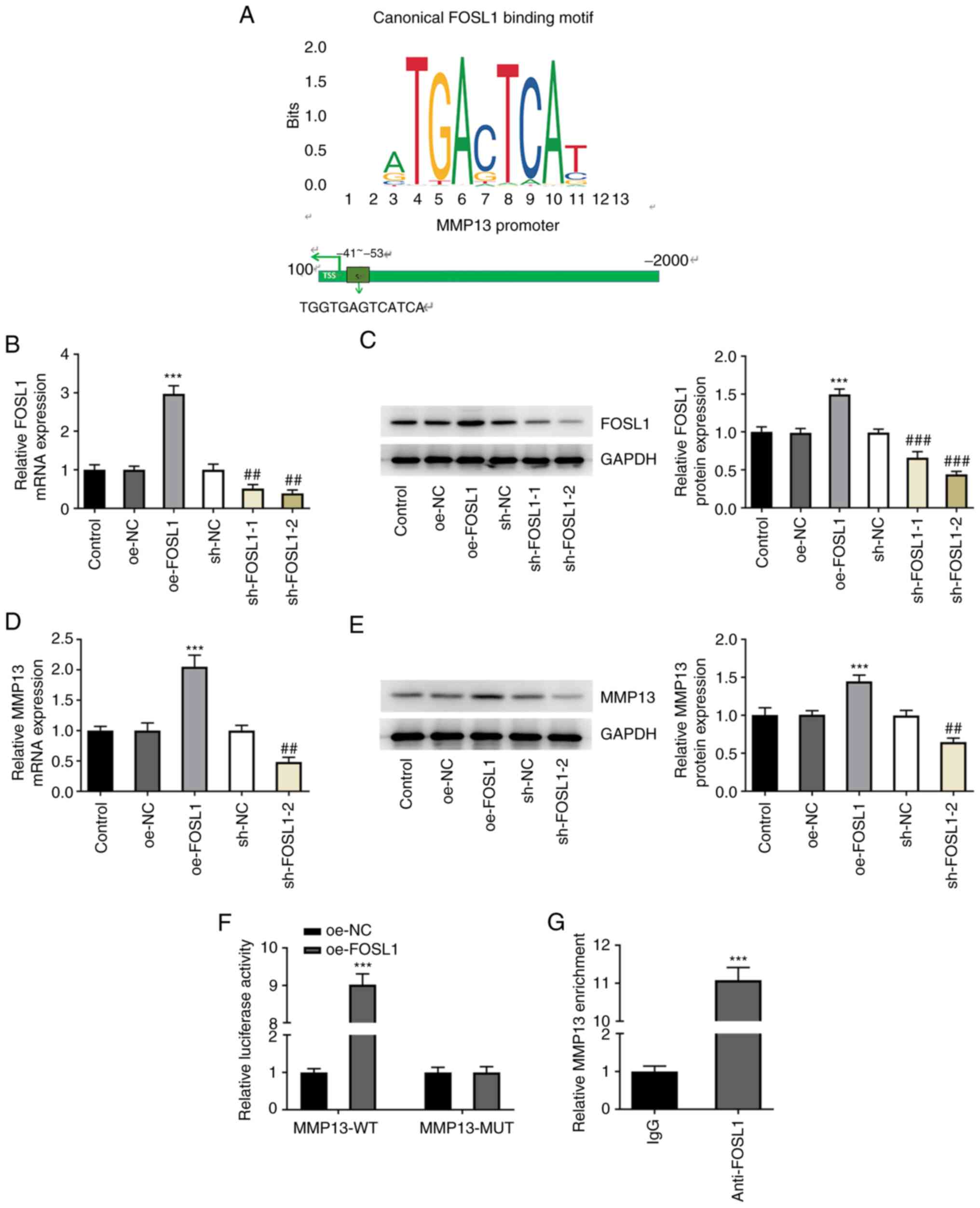
FOSL1 binds to the MMP13 promoter
region and upregulates MMP13 expression levels
To determine the relationship between FOSL1 and
MMP13, the binding site of FOSL1 and the MMP13 promoter was
predicted using the JASPAR database. This analysis demonstrated
that FOSL1 may bind to the MMP13 promoter as a transcription factor
(Fig. 2A). Furthermore, the
results demonstrated that FOSL1 mRNA and protein expression levels
were increased following transfection with oe-FOSL1 compared with
the oe-NC group. However, these expression levels were reduced
following transfection with sh-FOSL1-1/2 compared with the sh-NC
group (Fig. 2B and C). sh-FOSL1-2 was selected for use in
subsequent experiments because of its higher efficiency. To further
determine the association between FOSL1 and MMP13, the mRNA and
protein expression levels of MMP13 were determined via RT-qPCR and
western blotting, respectively, following transfection. The results
demonstrated that FOSL1 overexpression resulted in elevated MMP13
mRNA and protein expression levels compared with the oe-NC group,
whereas sh-FOSL1-2 resulted in decreased MMP13 mRNA and protein
expression compared with the sh-NC group (Fig. 2D and E). Furthermore, the dual-luciferase
reporter assay demonstrated that there was markedly elevated MMP13
promoter activity in the MMP13-wild-type (WT) + oe-FOSL1 group
compared with the MMP13-WT + oe-NC group. However, there was no
difference between the MMP13-mutant (MUT) + oe-FOSL1 and MMP13-MUT
+ oe-NC groups (Fig. 2F). The ChIP
experiment also demonstrated that the relative enrichment abundance
of anti-MMP13 at the target gene site on anti-FOSL1 was markedly
higher compared with the NC IgG group (Fig. 2G). These results indicated that as
a transcription factor, FOSL1 may bind to the MMP13 promoter region
and therefore may have potentially upregulated MMP13 expression
levels.
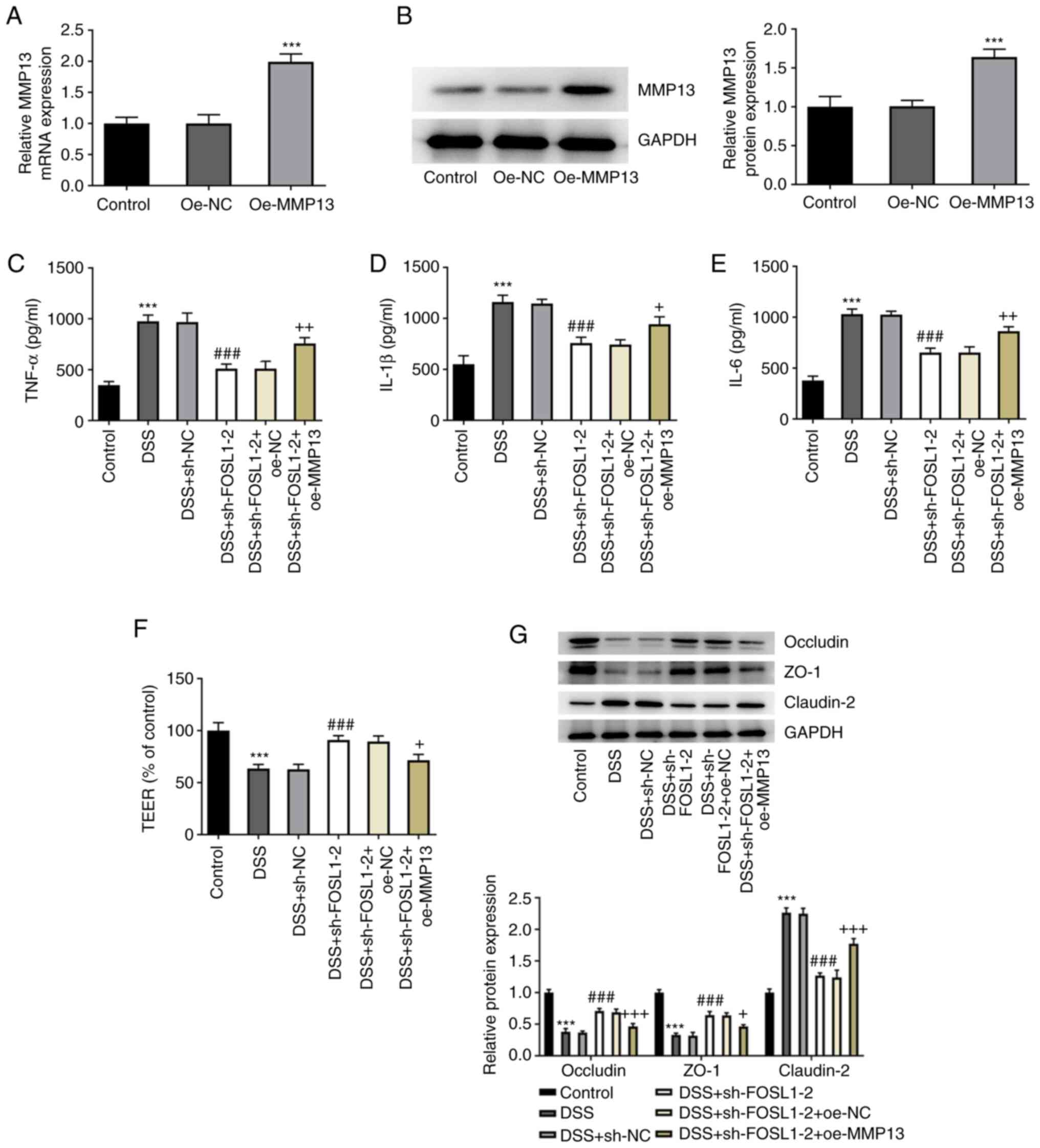
FOSL1 knockdown inhibits DSS-induced
inflammation and barrier damage in HT29 cells via MMP13
downregulation
To clarify whether FOSL1 knockdown had an effect on
DSS-induced HT29 cells MMP13 was overexpressed. The results
indicated that MMP13 was highly expressed following its
overexpression in DSS-induced HT29 cells compared with the oe-NC
group (Fig. 3A and B). Furthermore, in DSS-induced HT29 cells
there were increased levels of the inflammatory cytokines, TNF-α,
IL-1β and IL-6, compared with the control (Fig. 3C-E). Following transfection with
sh-FOSL1-2, TNF-α, IL-1β and IL-6 levels markedly decreased in
DSS-induced HT29 cells compared with the DSS + sh-NC group, whereas
MMP13 overexpression enhanced the levels of TNF-α, IL-1β and IL-6
compared with the DSS + sh-FOSL1-2 + oe-NC group. Subsequently,
TEER was performed to detect cell monolayer permeability. The
results demonstrated that DSS induced a marked decrease in cell
monolayer permeability, whereas there was a significant increase in
cell permeability following FOSL1 knockdown; however, MMP13
overexpression reversed the effect of FOSL1 knockdown (Fig. 3F). These results indicated that
FOSL1 knockdown may have effectively protected the HT29 cell
barrier from DSS-induced damage. It was also demonstrated that the
levels of tight junction-associated proteins, occludin and ZO-1, in
DSS-induced HT29 cells were markedly reduced in the DSS group
compared with the control group. However, these levels were
markedly elevated in the DSS + sh-FOSL1-2 group compared with the
DSS + sh-NC group, whereas in the DSS + sh-FOSL1-2 + oe-MMP13 group
these levels were reduced compared with the DSS + sh-FOSL1-2 +
oe-NC group (Fig. 3G). In
addition, the expression levels of claudin-2 in each group
exhibited a completely different trend from that of occludin and
ZO-1. These results indicated that FOSL1 knockdown potentially
inhibited DSS-induced inflammation and barrier damage in HT29 cells
via MMP13 downregulation.
FOSL1 knockdown ameliorates
DSS-induced colonic injury, inflammation and barrier damage in UC
mice via MMP13 downregulation
Subsequently it was investigated whether FOSL1
knockdown could ameliorate DSS-induced colonic injury in UC mice
via the downregulation of MMP13. Therefore, a DSS-induced UC mouse
model was constructed. The results demonstrated that there was a
marked increase in the protein expression levels of FOSL1 and MMP13
in UC mice, compared with the control group. Injection with
sh-FOSL1 greatly resulted in the reduction of FOSL1 and MMP13
expression in colon tissues of UC mice and oe-MMP13 elevated the
protein expression of MMP13 but had no influence on FOSL1
expression (Fig. 4A). It was
observed that the colons of UC mice were significantly shorter
compared with the control group (Fig.
4B and C). However, the colons
of UC mice injected with sh-FOSL1 were longer compared with UC
mice, whereas co-injection with oe-MMP13 and sh-FOSL1 resulted in
shorter colons of UC mice compared to injection with sh-FOSL1
alone. Furthermore, following H&E staining, the structure of
the intestinal tissues in UC mice was markedly altered compared
with the control (Fig. 4D), which
indicated that the colon tissues were diseased. However, the
intestinal tissue structure of UC mice injected with sh-FOSL1 was
markedly recovered, whereas MMP13 overexpression damaged the
structure of the intestinal tissues of UC mice injected with
sh-FOSL1. Furthermore, the DAI of the mice in the different groups
was recorded. The results demonstrated that there was a marked
increase in the DAI of UC mice compared with the control group
(Fig. 4E). However, the UC mice
injected with sh-FOSL1 exhibited a marked decrease in the DAI, but
the overexpression of MMP13 increased the DAI. Subsequently, ELISA
was performed and the results demonstrated increased levels of the
inflammatory cytokines, TNF-α, IL-1β and IL-6 in the UC group,
whereas decreased levels of TNF-α, IL-1β and IL-6 were demonstrated
in the UC-sh-FOSL1 group. In addition, elevated levels of TNF-α,
IL-1β and IL-6 were demonstrated in the UC + sh-FOSL1 + oe-MMP13
group (Fig. 4F-H). Furthermore,
the protein expression levels of occludin and ZO-1 were markedly
decreased in the UC group, but were markedly increased in the UC +
sh-FOSL1 group and slightly decreased in the UC + sh-FOSL1 +
oe-MMP13 group (Fig. 4I). The
expression levels of claudin-2 in each group exhibited a completely
different trend from that of occludin and ZO-1. These results
indicated that FOSL1 knockdown may have ameliorated DSS-induced
colonic injury, inflammation and barrier damage in UC mice via the
downregulation of MMP13.
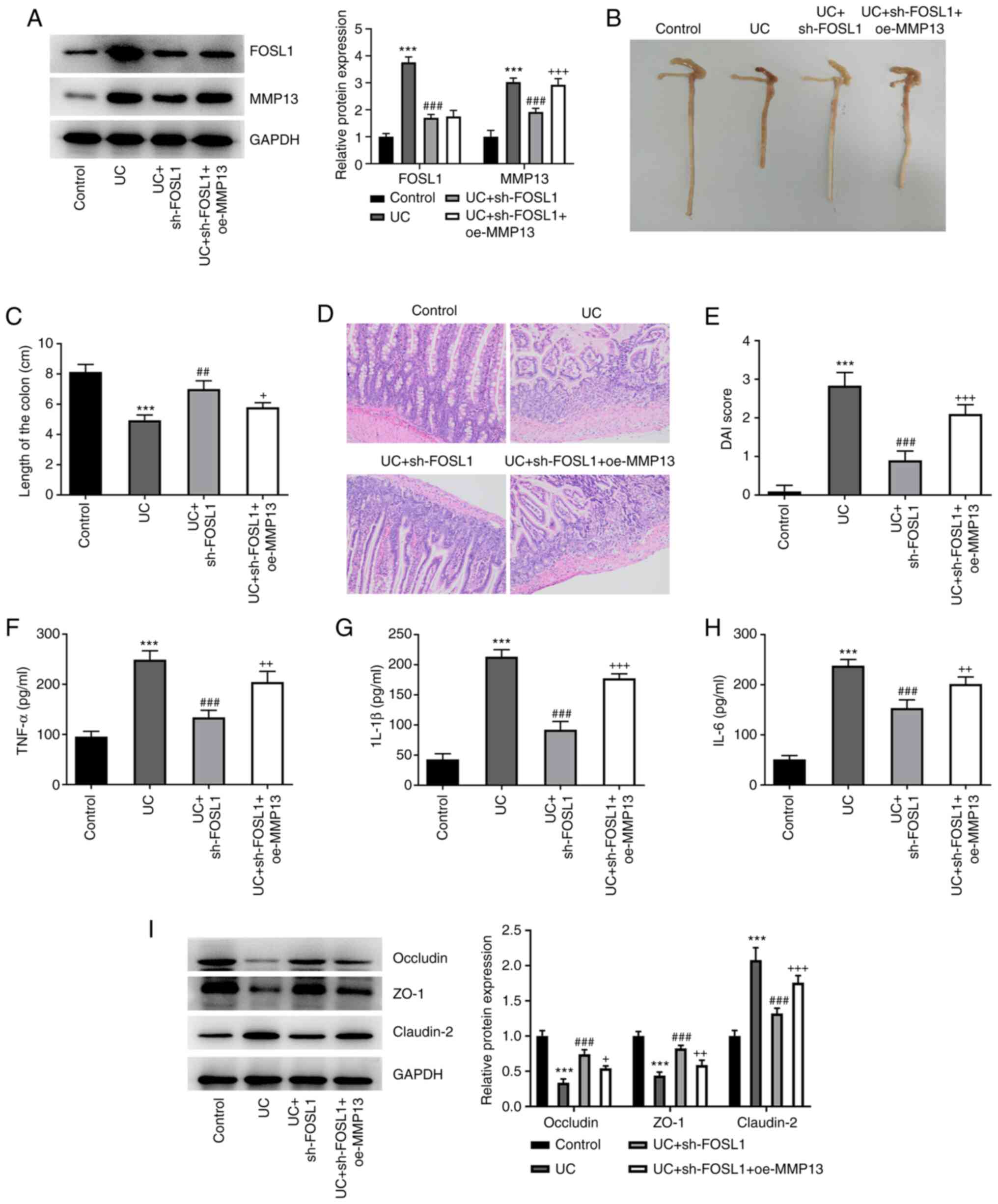 | Figure 4FOSL1 knockdown ameliorates
DSS-induced colonic injury, inflammation and barrier damage in UC
mice via the downregulation of MMP13. (A) FOSL1 and MMP13 protein
expression levels in colonic tissues were determined via western
blotting. ***P<0.001 vs. control;
###P<0.001 vs. UC; +++P<0.001 vs. UC +
sh-FOSL1. (B and C) Colon length in UC mice.
***P<0.001 vs. control; ##P<0.01 vs.
UC; +P<0.05 vs. UC + sh-FOSL1. (D) Colonic damage
level was assessed using H&E staining. Magnification, x200. (E)
Disease activity index. ***P<0.001 vs. control;
###P<0.001 vs. UC; +++P<0.001 vs. UC +
sh-FOSL1. (F-H) Serum inflammatory factors levels were determined
via ELISA. ***P<0.001 vs. control;
###P<0.001 vs. UC; ++P<0.01,
+++P<0.001 vs. UC + sh-FOSL1. (I) Tight
junction-associated protein expression levels in tissues were
detected via western blotting. ***P<0.001 vs.
control; ###P<0.001 vs. UC; +P<0.05,
++P<0.01, +++P<0.001 vs. UC + sh-FOSL1.
FOSL1, Fos-like antigen-1; DSS, dextran sodium sulfate; UC,
ulcerative colitis; sh, short hairpin RNA; NC, negative control;
oe, overexpression. |
Discussion
The incidence of UC in Asian populations has
increased rapidly over the last few decades (21). The high incidence of UC is mainly
due to inflammation and damage to the intestinal mucosal barrier
(3). At present UC is an urgent
global health problem and early prevention and treatment is
critical (22). In the present
study it was demonstrated that FOSL1 was a potentially key
biological factor in UC, as determined by the following
observations. First, the FOSL1 and MMP13 expression levels in
DSS-induced HT29 cells and UC mice were markedly increased. Second,
FOSL1 was demonstrated to bind to the MMP13 promoter region and
consequently upregulated the MMP13 expression level. Third, FOSL1
knockdown inhibited DSS-induced inflammation and barrier damage via
the downregulation of MMP13 expression levels in HT29 cells and UC
mice.
FOSL1 is one of the major transcription factors of
the AP-1 family and is upregulated in numerous malignancies,
including prostate cancer, breast cancer and colon cancer (23-25).
A previous study reported that FOSL1 was elevated in patients with
mild UC (11). In the present
study it was demonstrated that FOSL1 was highly expressed in
DSS-induced HT29 cells and the serum of UC mice colon tissues. In
addition, MMP-13 has previously been demonstrated to be a major
protease in the pathogenesis of IBD-associated mucosal ulcers
(26). A significant increase in
MMP13 mRNA expression levels has also previously been reported in
IBD biopsy specimens (15). The
results of the present study demonstrated a significant three-fold
increase in the expression levels of MMP13 in DSS-induced HT29
cells and the serum of UC mice colon tissues. Together, these data
indicated that FOSL1 and MMP13 may serve key roles in UC.
Furthermore, the promoters of most human MMP genes contain an AP-1
binding site at ~-70 bp and AP-1 has been broadly known to modulate
the expression of MMP in numerous cell types (27). In the present study, the potential
relationship between FOSL1 and MMP13 was predicted using the JASPAR
database. It was demonstrated that the expression of MMP13 was
increased following FOSL1 overexpression but decreased following
FOSL1 knockdown. Furthermore, higher promoter region activity and
relative MMP13 enrichment levels also indicated that FOSL1
interacted with the MMP13 promoter region, which upregulated MMP13
expression levels in DSS-induced HT29 cells and the serum and
tissues of UC mice. In addition, the present study investigated
colonic morphology and determined the DAI of UC mice based on their
body weight, fecal condition and occult blood. It was reported that
DSS induced a shorter colonic length, a severe level of colonic
damage and resulted in a higher DAI. However, these conditions were
alleviated by FOSL1 knockdown but were partially reversed by MMP13
overexpression. Therefore the results suggested that FOSL1
knockdown may have protected the colon of UC mice from DSS-induced
injury via the downregulation of MMP13.
A dysregulated inflammatory response serves a key
role in the development of UC (1).
It has previously been reported that cells immersed in a chronic
inflammatory cytokine environment can overproduce pro-inflammatory
cytokines, such as TNF-α and IL-6(28). TNF-α is elevated in the blood and
mucosa of patients with UC (29).
Furthermore, IL-6 transduction signaling stimulates inflammatory
cells and stromal cells to induce colonic inflammation (1). The results of the present study
demonstrated that DSS induced high levels of the pro-inflammatory
cytokines, TNF-α, IL-1β and IL-6 in HT29 cells and UC mice. In
addition, it has previously been reported that the increased
secretion of IL-6 promotes the deacetylation of FOSL1 in colorectal
cancer cell lines, which leads to increased FOSL1 expression
(30). AP-1 regulates numerous
pro-inflammatory cytokine genes, including TNF-α, IL-1 and IL-6,
which perform a key role during chronic inflammation (31). It can therefore be hypothesized
that reduced FOSL1 may affect the expression of pro-inflammatory
cytokines. In the present study the knockdown of FOSL1 reduced the
levels of TNF-α, IL-1β and IL-6 in DSS-induced HT29 cells and the
serum of UC mice, which was consistent with the aforementioned
report (32). However, this effect
was partially reversed by MMP13 overexpression. These results
further indicated that FOSL1 knockdown may have ameliorated
DSS-induced inflammation in HT29 cells and the serum of UC mice via
the downregulation of MMP13.
The tight junctions between intestinal epithelial
cells serve an important role in maintaining intestinal mucosal
permeability (4). However, in UC,
damage to the epithelial barrier leads to increased permeability,
which may be due to defects in the regulation of tight junctions
(32). Furthermore, inflammatory
cytokines can also mediate the abnormal expression of tight
junction proteins due to the ability of TNF-α to abnormalize the
function of ZO-1. Subsequently, this result in the disruption of
the structure of the tight junction-associated protein claudin-1,
which in turn disrupts other tight junction proteins and leads to
the development of IBD (6). In the
present study, it was demonstrated that cell monolayer permeability
and the levels of tight junction-associated protein occludin and
ZO-1 were markedly decreased, whereas the level of claudin-2 was
markedly elevated in DSS-induced HT29 cells and the tissues of UC
mice. In addition, the activation of AP-1 reduces ZO-1 expression
in Caco-2 intestinal epithelial cells via the transcriptional
repression of the ZO-1 promoter (33). However, the reduction of AP-1 may
increase the expression levels of tight junction-associated
proteins. In the present study, the increased expression levels of
occludin and ZO-1 and decreased expression levels of claudin-2 also
demonstrated that FOSL1 knockdown may alleviate barrier damage in
DSS-induced HT29 cells and the tissues of UC mice. However, MMP13
overexpression partially reversed these effects. These results
indicated that FOSL1 knockdown may have ameliorated DSS-induced
barrier damage in HT29 cells and the tissues of UC mice via the
downregulation of MMP13.
However, limitations still exist in the present
study. Here, HT29 cell line, a colorectal cancer cell line, was
applied for constructing colitis in vitro; however, the use
of other non-cancerous cells to simulate colitis in vitro is
deserved to be explored in future work. In addition, the clinical
significance of FOSL1 in colitis needs to be verified urgently.
In conclusion, this present study assessed the
impacts of FOSL1 on UC. The results demonstrated that FOSL1
knockdown may downregulate MMP13 expression levels to improve
inflammatory damage and protect intestinal barrier integrity in
DSS-induced HT29 cells and UC mice. The present study has therefore
provided a novel approach for the diagnosis and treatment of
UC.
Acknowledgements
Not applicable.
Funding
Funding: This present study was supported by S&T Program of
Hebei (grant no. H2019209241) and Doctoral research project, North
China University of Science and Technology (grant no. bs2104).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LM designed the experiments. LM, XZ, CZ, BH and HZ
conducted the experiments. LM, XZ and CZ analyzed and interpreted
the data. LM and XZ drafted and revised the manuscript. LM and XZ
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
All experimental procedures were approved by the
Ethics Committee of the North China University of Science and
Technology Affiliated Hospital (approval no. Lx201896).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yao D, Dong M, Dai C and Wu S:
Inflammation and inflammatory cytokine contribute to the initiation
and development of ulcerative colitis and its associated cancer.
Inflamm Bowel Dis. 25:1595–1602. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Seyedian SS, Nokhostin F and Malamir MD: A
review of the diagnosis, prevention, and treatment methods of
inflammatory bowel disease. J Med Life. 12:113–122. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Porter RJ, Kalla R and Ho GT: Ulcerative
colitis: Recent advances in the understanding of disease
pathogenesis. F1000Res. 9(F1000 Faculty Rev-294)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Pruteanu M and Shanahan F: Digestion of
epithelial tight junction proteins by the commensal Clostridium
perfringens. Am J Physiol Gastrointest Liver Physiol.
305:G740–G748. 2013.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Sun M, He C, Cong Y and Liu Z: Regulatory
immune cells in regulation of intestinal inflammatory response to
microbiota. Mucosal Immunol. 8:969–978. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Shi Y, Bao CH, Wu HG, Ma XP, Yu LQ, Zhang
R and Chen WF: Effect of moxibustion on colonic TNF-alpha content
and influence of colonic supernatant of Crohn's disease rats
undergoing moxibustion on expression of occludin, claudin-1 and
zonula occludens-1 proteins and genes in cultured colonic
epithelial cells. Zhen Ci Yan Jiu. 36:235–241. 2011.PubMed/NCBI(In Chinese).
|
|
7
|
Tulchinsky E: Fos family members:
Regulation, structure and role in oncogenic transformation. Histol
Histopathol. 15:921–928. 2000.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Elangovan IM, Vaz M, Tamatam CR, Potteti
HR, Reddy NM and Reddy SP: FOSL1 Promotes Kras-induced lung cancer
through amphiregulin and cell survival gene regulation. Am J Respir
Cell Mol Biol. 58:625–635. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Luo YZ, He P and Qiu MX: FOSL1 enhances
growth and metastasis of human prostate cancer cells through
epithelial mesenchymal transition pathway. Eur Rev Med Pharmacol
Sci. 22:8609–8615. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Iskit S, Schlicker A, Wessels L and Peeper
DS: Fra-1 is a key driver of colon cancer metastasis and a Fra-1
classifier predicts disease-free survival. Oncotarget.
6:43146–43161. 2015.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Sabzevary-Ghahfarokhi M, Shohan M, Shirzad
H, Rahimian G, Bagheri N, Soltani A, Deris F, Ghatreh-Samani M and
Razmara E: The expression analysis of Fra-1 gene and IL-11 protein
in Iranian patients with ulcerative colitis. BMC Immunol.
19(17)2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wang X, Guo R, Lv Y and Fu R: The
regulatory role of Fos related antigen1 in inflammatory bowel
disease. Mol Med Rep. 17:1979–1985. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Moriyama I, Ishihara S, Rumi MA, Aziz MD,
Mishima Y, Oshima N, Kadota C, Kadowaki Y, Amano Y and Kinoshita Y:
Decoy oligodeoxynucleotide targeting activator protein-1 (AP-1)
attenuates intestinal inflammation in murine experimental colitis.
Lab Invest. 88:652–663. 2008.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Balzan S, de Almeida Quadros C, de Cleva
R, Zilberstein B and Cecconello I: Bacterial translocation:
Overview of mechanisms and clinical impact. J Gastroenterol
Hepatol. 22:464–471. 2007.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Rath T, Roderfeld M, Graf J, Wagner S,
Vehr AK, Dietrich C, Geier A and Roeb E: Enhanced expression of
MMP-7 and MMP-13 in inflammatory bowel disease: A precancerous
potential? Inflamm Bowel Dis. 12:1025–1035. 2006.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Vizoso FJ, Gonzalez LO, Corte MD, Corte
MG, Bongera M, Martinez A, Martin A, Andicoechea A and Gava RR:
Collagenase-3 (MMP-13) expression by inflamed mucosa in
inflammatory bowel disease. Scand J Gastroenterol. 41:1050–1055.
2006.PubMed/NCBI View Article : Google Scholar
|
|
17
|
O'Sullivan S, Gilmer JF and Medina C:
Matrix metalloproteinases in inflammatory bowel disease: An update.
Mediators Inflamm. 2015(964131)2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Vandenbroucke RE, Dejonckheere E, Van
Hauwermeiren F, Lodens S, De Rycke R, Van Wonterghem E, Staes A,
Gevaert K, Lopez-Otin C and Libert C: Matrix metalloproteinase 13
modulates intestinal epithelial barrier integrity in inflammatory
diseases by activating TNF. EMBO Mol Med. 5:1000–1016.
2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Chaudhary G, Mahajan UB, Goyal SN, Ojha S,
Patil CR and Subramanya SB: Protective effect of Lagerstroemia
speciosa against dextran sulfate sodium induced ulcerative
colitis in C57BL/6 mice. Am J Transl Res. 9:1792–1800.
2017.PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Burisch J and Munkholm P: Inflammatory
bowel disease epidemiology. Curr Opin Gastroenterol. 29:357–362.
2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhang YZ and Li YY: Inflammatory bowel
disease: Pathogenesis. World J Gastroenterol. 20:91–99.
2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zerbini LF, Wang Y, Cho JY and Libermann
TA: Constitutive activation of nuclear factor kappaB p50/p65 and
Fra-1 and JunD is essential for deregulated interleukin 6
expression in prostate cancer. Cancer Res. 63:2206–2215.
2003.PubMed/NCBI
|
|
24
|
Belguise K, Kersual N, Galtier F and
Chalbos D: FRA-1 expression level regulates proliferation and
invasiveness of breast cancer cells. Oncogene. 24:1434–1444.
2005.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Diesch J, Sanij E, Gilan O, Love C, Tran
H, Fleming NI, Ellul J, Amalia M, Haviv I, Pearson RB, et al:
Widespread FRA1-dependent control of mesenchymal
transdifferentiation programs in colorectal cancer cells. PLoS One.
9(e88950)2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Rath T, Roderfeld M, Blocher S, Rhode A,
Basler T, Akineden O, Abdulmawjood A, Halwe JM, Goethe R, Bulte M
and Roeb E: Presence of intestinal Mycobacterium avium
subspecies paratuberculosis (MAP) DNA is not associated with
altered MMP expression in ulcerative colitis. BMC Gastroenterol.
11(34)2011.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Yan C and Boyd DD: Regulation of matrix
metalloproteinase gene expression. J Cell Physiol. 211:19–26.
2007.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wang C, Li W, Wang H, Ma Y, Zhao X, Zhang
X, Yang H, Qian J and Li J: Saccharomyces boulardii alleviates
ulcerative colitis carcinogenesis in mice by reducing TNF-α and
IL-6 levels and functions and by rebalancing intestinal microbiota.
BMC Microbiol. 19(246)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ordas I, Eckmann L, Talamini M, Baumgart
DC and Sandborn WJ: Ulcerative colitis. Lancet. 380:1606–1619.
2012.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Malcomson FC, Willis ND, McCallum I, Xie
L, Shivappa N, Wirth MD, Hebert JR, Kocaadam-Bozkurt B,
Ozturan-Sirin A, Kelly SB, et al: Diet-Associated Inflammation
Modulates Inflammation and WNT Signaling in the Rectal Mucosa, and
the Response to Supplementation with Dietary Fiber. Cancer Prev Res
(Phila). 14:337–346. 2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Ip YT and Davis RJ: Signal transduction by
the c-Jun N-terminal kinase (JNK)-from inflammation to development.
Curr Opin Cell Biol. 10:205–219. 1998.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Heller F, Florian P, Bojarski C, Richter
J, Christ M, Hillenbrand B, Mankertz J, Gitter AH, Burgel N, Fromm
M, et al: Interleukin-13 is the key effector Th2 cytokine in
ulcerative colitis that affects epithelial tight junctions,
apoptosis, and cell restitution. Gastroenterology. 129:550–564.
2005.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Chen J, Xiao L, Rao JN, Zou T, Liu L,
Bellavance E, Gorospe M and Wang JY: JunD represses transcription
and translation of the tight junction protein zona occludens-1
modulating intestinal epithelial barrier function. Mol Biol Cell.
19:3701–3712. 2008.PubMed/NCBI View Article : Google Scholar
|